1 1. VZNIK A VÝVOJ MNOHOBUNĚČNÝCH ŽIVOČICHŮ, RÝHOVÁNÍ ZYGOTY KOPINATCE, TKÁNĚ 1.1. Živočišná buňka Obecné znaky buňky Definice buňky Buňka ( cellula, cyton) je základní stavební a funkční jednotka všech živých organismů. Je to nejmenší útvar schopný samostatné existence a reprodukce (viz buněčná teorie - Th. Schwann 1839). Studiem buňky se zabývá cytologie (buněčná biologie). Studuje biologii buňky komplexně - tj. z hlediska biofyziky, biochemie, molekulární biologie, fyziologie, genetiky a morfologie. OBECNÉ ZNAKY BUŇKY Buňka má: • vlastní genetický a proteosyntetický aparát • vlastní energetický metabolismus • je ohraničena membránou (reguluje pohyb látek dovnitř a ven) Tyto znaky odlišují buňky od nebuněčných forem živých soustav - virů. Výše uvedené znaky má buňka prokaryotická i eukaryotická. Kromě buněk se na stavbě organismů podílejí mezibuněčné hmoty (nacházejí se vně buňky hlavně v živočišných pojivech - kost, chrupavka) a soubuní, což jsou mnohojaderné útvary vzniklé buď opakovaným dělením jádra bez dělení buňky nebo také splýváním buněk. Struktura buňky Pod pojmem struktura rozumíme velikost, tvar a vnitřní uspořádání buňky. Rozlišujeme strukturu: mikroskopickou - to, co vidíme světelným mikroskopem submikroskopickou - to, so pozorujeme elektronovým mikroskopem (také ultrastruktura) Typy buněk prokaryotická buňka - viz dříve eukaryotická buňka - a) rostlinná, b) živočišná, c) buňka hub Prokaryotická buňka je jednodušší než eukaryotická. Obsahuje minimální množství biomembrán. Nikdy nevytváří mnohobuněčný organismus, nanejvýše kolonie. Stavba živočišné buňky: buněčné povrchy cytoplazma buněčné organely a) glykokalyx + cytoplazmatická membrána b) cytoplazma c) buněčné jádro d) endoplazmatické retikulum e) Golgiho komplex f) lyzozómy g) mitochondrie h) vakuoly i) cytoskelet
Transcript
1
1. VZNIK A VÝVOJ MNOHOBUNĚČNÝCH ŽIVOČICHŮ,
RÝHOVÁNÍ ZYGOTY KOPINATCE, TKÁNĚ 1.1. Živočišná buňka Obecné znaky buňky Definice buňky Buňka ( cellula, cyton) je základní stavební a funkční jednotka všech živých organismů. Je to nejmenší útvar schopný samostatné existence a reprodukce (viz buněčná teorie - Th. Schwann 1839). Studiem buňky se zabývá cytologie (buněčná biologie). Studuje biologii buňky komplexně - tj. z hlediska biofyziky, biochemie, molekulární biologie, fyziologie, genetiky a morfologie.
OBECNÉ ZNAKY BUŇKY
Buňka má: • vlastní genetický a proteosyntetický aparát • vlastní energetický metabolismus • je ohraničena membránou (reguluje pohyb látek dovnitř a ven) Tyto znaky odlišují buňky od nebuněčných forem živých soustav - virů. Výše uvedené znaky má buňka prokaryotická i eukaryotická. Kromě buněk se na stavbě organismů podílejí mezibuněčné hmoty (nacházejí se vně buňky hlavně v živočišných pojivech - kost, chrupavka) a soubuní, což jsou mnohojaderné útvary vzniklé buď opakovaným dělením jádra bez dělení buňky nebo také splýváním buněk.
Struktura buňky Pod pojmem struktura rozumíme velikost, tvar a vnitřní uspořádání buňky. Rozlišujeme strukturu: � mikroskopickou - to, co vidíme světelným mikroskopem � submikroskopickou - to, so pozorujeme elektronovým mikroskopem (také ultrastruktura)
Typy buněk � prokaryotická buňka - viz dříve � eukaryotická buňka - a) rostlinná, b) živočišná, c) buňka hub Prokaryotická buňka je jednodušší než eukaryotická. Obsahuje minimální množství biomembrán. Nikdy nevytváří mnohobuněčný organismus, nanejvýše kolonie.
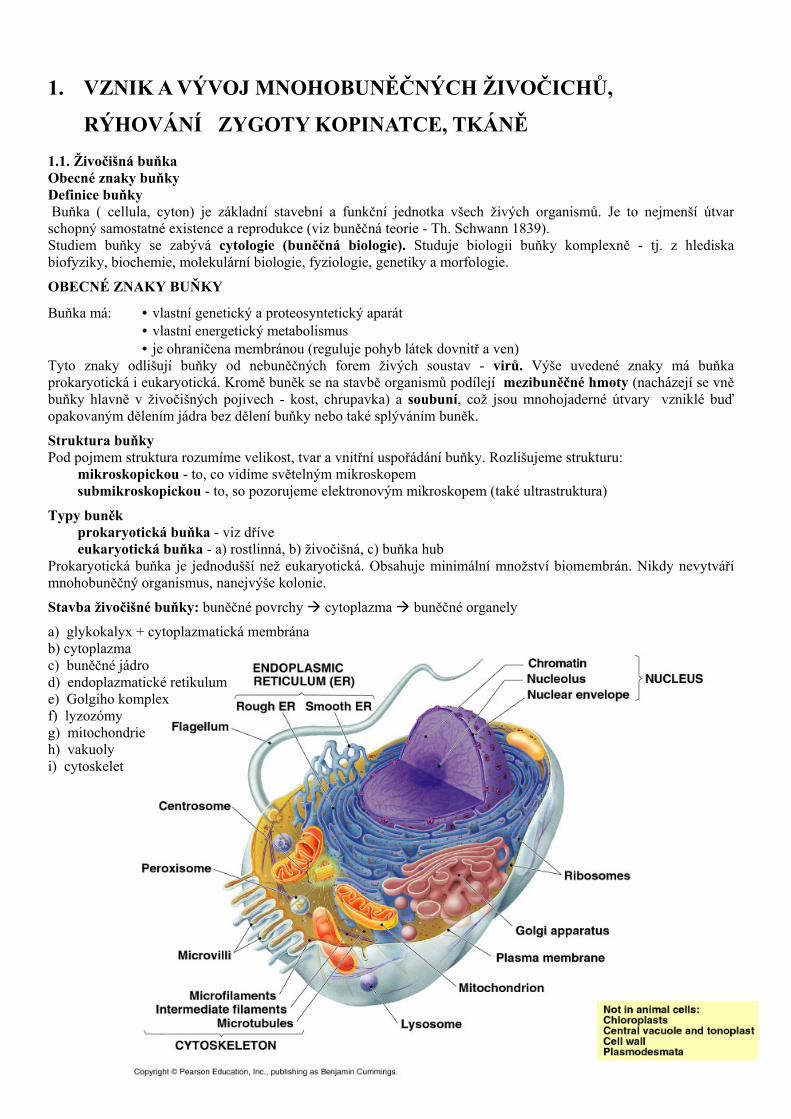
Stavba živočišné buňky: buněčné povrchy cytoplazma buněčné organely
a) glykokalyx + cytoplazmatická membrána b) cytoplazma c) buněčné jádro d) endoplazmatické retikulum e) Golgiho komplex f) lyzozómy g) mitochondrie h) vakuoly i) cytoskelet
2
BIOMEMBRÁNY - biomembrány jednotlivých typů organel se vzájemně liší obsahem některých látek, rozdílnými funkcemi i
různou tloušťkou. Na jejich utváření se jako základní látky podílejí bílkoviny a složené tuky (fosfolipidy, glykolipidy a steroidy). Pro složené tuky je charakteristické, že mají část své molekuly hydrofilní, např. PO4 skupiny, a část hydrofóbní tvořenou zbytky mastných kyselin. Díky tomu mohou vytvářet ve vodném prostředí blanku označovanou jako vrstvový koacervát, jehož molekuly se orientují hydrofóbními částmi k sobě a hydrofilními k okolnímu vodnímu prostředí. Takto vzniklá blanka ohraničuje vždy kulovitý nebo geometricky od něj odvozený prostor, který tvoří hranici mezi dvěma různými systémy vodných roztoků a je základem membrán ohraničujících vnitřní prostor buňky (cytoplazmatická membrána) nebo obsah různých typů buněčných organel - tato membrána se jeví jako polopropustná (semipermeabilní) tj. schopná propouštět jen molekuly vody a nikoli látky ve vodě rozpuštěné vně a uvnitř membránové struktury. Zatímco lipidická vrstva slouží jako ostrá hranice mezi roztoky různých látek vně a uvnitř membránové struktury, bílkovinná složka zabezpečuje výběrově některým látkám volný průchod.
- podle pevnosti vazby bílkovin na membránové lipidy a polohy v biomembráně rozlišujeme bílkoviny periferní (povrchové), které jsou na vnější nebo vnitřní straně lipidické vrstvy a plní různé enzymatické funkce a na propustnost lipidické vrstvy nemají vliv, a integrální zabudované do vrstev lipidů. Enzymatické funkce plní intergální bílkoviny chemicky vázané na membránové lipidy.
Membránové váčky mají schopnost vzájemného splývání (fúze biomembrán) a dělení. Takto je umožněno množení buněčných organel, jejich zvětšování, zmenšování a přeprava látek z jedné organely do druhé - tzv. cytózy. CYTOPLAZMATICKÁ MEMBRÁNA - přítomna na povrchu všech typů buněk jako hraniční vrstva mezi vnitřním a vnějším prostřením buňky. Kromě lipoproteinové vrstvy o síle 5 nm je zevně kryta vrstvou cukerných látek - sacharidový plášť = glykokalyx. Tyto látky nesou a svých volných koncích funkční skupiny schopné reagovat s různými látkami v okolí buňky a slouží k příjmu látkových signálů a podílejí se na přenosu informací do vnitřního prostředí buňky, k vazbám částic a v buněčných koloniích a mnohobuněčných organismech se podílejí na tvorbě mezibuněčných spojů. Součástí cytoplazmat. membrány je také svrchní vrstva prostoupená hustou sítí cytoskeletu, jehož vlákna, zejména aktinová mikrofilamenta pevně vázána na bílkoviny , slouží ke zpevnění povrchu buňky, umožňují změny tvaru buňky a jiné povrchové funkce. Spolu s cytoskeletem se cytoplazmatická membrána podílí také na utváření trvalých i dočasných pohybových organel buňky - např. panožky, bičíky, řasinky nebo mikroklky, sloužících k pohybu buňky, přijímání potravy, výměně tekutin v okolí buňky atd. - je také v úzkých funkčních a morfologických vztazích s nitrobuněčnými strukturami (s endoplasmatickou sítí, s Golgiho komplexem, dyktyozómy, vakuolami apod.) Její nejdůležitější funkcí je polopropustnost látek, která zabezpečuje ochranu vnitřního prostředí buňky a umožňuje její metabolické funkce. Vzhledem k tomu, že je lipidická vrstva biomembrán polopropustná, mohou skrze ni pronikat kromě molekul vody jen látky rozpustné v tucích. Jelikož je tato membrána prostoupena také drobnými póry vyplněnými vodou, mohou skrze ni pronikat i hydrofilní částice o velikosti 0,4 - 0,8 nm (některé ionty). Větší částice (nízkomolekulární organ. látky aj. ) se mohou vázat s integrálními bílkovinami a pomocí nich se dostávat na druhou stranu polopropustné vrstvy. Tyto transporty se dějí na základě fyzikálních zákonů kinetiky hmotných soustav – zákona difúze a osmotického zákona.
3
NITROBUNĚČNÉ MEMBRÁNOVÉ ORGANELY - podle významu se mohou dělit na tři skupiny: a) buněčné jádro - řídící centrum buňky b) semiautonomní organely (mitochondrie a plastidy) c) ostatní membránové struktury a) buněčné jádro (karyon = nucleus)
Jádro je nedílnou součástí téměř všech buněk. Jen zcela výjimečně, u buněk vysoce specializovaných, může dojít k jeho vymizení (např. v červených krvinkách savců). Stává se tak jen u buněk, které se po ukončení diferenciace jež nerozmnožují a po funkčním opotřebení odumírají.
Tvar jádra bývá nejčastěji kulovitý nebo oválný, vzácněji růžencovitý, vláknitý, podkovovitý, skobovitý, větvený aj. Jádro je největší organelou buňky, zaujímá asi 10% jejího celkového objemu , v savčích buňkách má průměr okolo 6 mikrometrů, ale u buněk různě specializovaných, různých vývojových stadií, různých druhů organismů se může velikost jádra značně lišit.
Většina buněk má jádro jen jedno (monoenergidní buňky), vzácněji se setkáváme i s vícejadernými (polyenergidními) buňkami, které dělíme podle způsobu vzniku do dvou skupin:
a) plazmódia - vznikají opakovaným dělením jádra, aniž by se dělila buňka jako celek (bez dělení plazmy) b) syncytia - vznikají rozrušením přepážek mezi několika sousedními buňkami a splynutím jejich cytoplazmy (fúzí jejich cytoplazmatických membrán.) V plazmódiích a syncytiích jsou zpravidla všechny jádra tvarově a funkčně stejná, jen u nálevníků (prvoci) se setkáváme s buňkami o dvou velikostně i funkčně rozlišitelných jádrech. Velké jádro (makronukleus) řídí metabolické funkce a malé jádro (mikronukleus) se uplatňuje při dělení buňky.
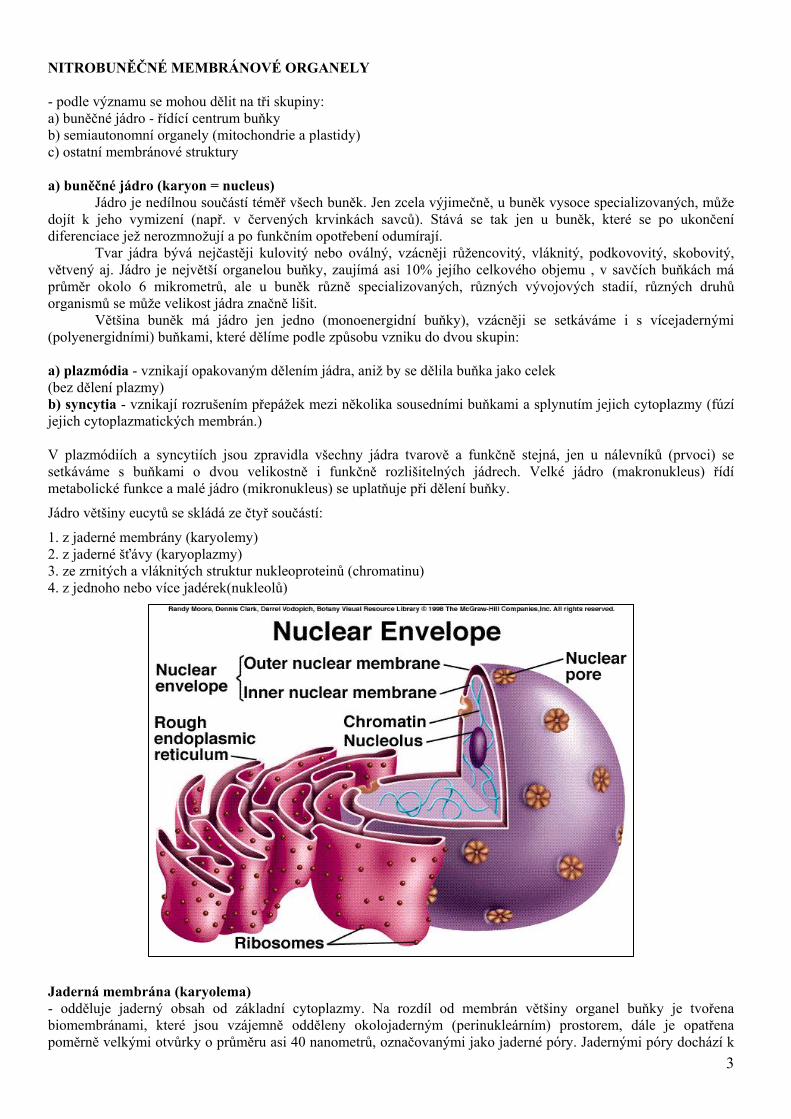
Jádro většiny eucytů se skládá ze čtyř součástí:
1. z jaderné membrány (karyolemy) 2. z jaderné šťávy (karyoplazmy) 3. ze zrnitých a vláknitých struktur nukleoproteinů (chromatinu) 4. z jednoho nebo více jadérek(nukleolů) Jaderná membrána (karyolema) - odděluje jaderný obsah od základní cytoplazmy. Na rozdíl od membrán většiny organel buňky je tvořena biomembránami, které jsou vzájemně odděleny okolojaderným (perinukleárním) prostorem, dále je opatřena poměrně velkými otvůrky o průměru asi 40 nanometrů, označovanými jako jaderné póry. Jadernými póry dochází k
4
řízené výměně makromolekulárních látek (RNA a bílkovin) mezi karyoplazmou a cytoplazmou. Ostatní molekuly a ionty mohou procházet biomembránami jaderné membrány a okolojaderným prostorem osmoticky, difúzí a aktivním transportem.
Jaderná šťáva (karyoplazma ) - obsahuje zpravidla více vody než základní cytoplazma. Lze v ní rozlišit složku rozpustnou, směs koloidních a krystaloidních roztoků, a nerozpustnou, tvořenou sítí bílkovinných vláken označovanou jako jaderný skelet.
Jaderný chromatin - jeho základem jsou nukleoproteiny i volné bílkoviny a nukleové kyseliny, nositelé genetické informace. Velké shluky chromatinu, tvořící silně spiralizovaná vlákna DNA proteinů, která se nepodílejí na transkripci, se označují jako heterochromatin. Aktivní molekuly DNA jsou despiralizované a uvolněné z vazby s histony a tvoří tzv. euchromatin. V dělících se buňkách se veškerý genetický materiál soustřeďuje v chromozómech, kde každý chromozóm je tvořen silně spiralizovanou molekulou DNA s navázanými histony i jinými bílkovinami. Jadérko (nukleolus) - je zpravidla kulovitý, silně světlolomný útvar uložený v jaderné šťávě asi ve středu jádra nebo pod jadernou membránou. Jeho velikost se pohybuje v rozmezí 1-5 mikrometrů. Byla v něm rozlišena vláknitá a zrnitá složka. Zrnitá složka jsou již hotové ribozomální podjednotky tvořené komplexy r-RNA a bílkovin, které jsou do jadérka dopravovány ze základní cytoplazmy. Ribozomální podjednotky se z jadérka uvolňují a jadernými póry pronikají do cytoplazmy, kde se z nich vytvářejí kompletní ribozómy. V jádře se zpravidla vyskytuje jedno nebo dvě jadérka, vzácněji jich může být v jednom jádře několik desítek až stovek. Oproti jaderné šťávě není jadérko ohraničeno biomembránou. V období buněčného dělení jadérko jako samostatná struktura mizí a zachovává se jen úsek molekuly DNA v podobě organizátoru jadérka. b) semiautonomní organely buňky Podle tzv. endosymbiotické teorie docházelo ve vývoji eucytů ke zdokonalování jejich metabolických funkcí také soužitím s protocyty, které měly některé z metabol. drah vyvinuty již dokonaleji než primitivní eucyt. Z protocytů schopných dokonalým způsobem využívat světelnou energii k syntéze organických látek cestou fotosyntézy se vyvinuly plastidy a z protocytů, u kterých byla nejdokonaleji vyvinuta schopnost získávat potřebnou energii rozkladem organ. látek až na CO2 a H2O za spoluúčasti kyslíku se vyvinuly mitochondrie. Tyto organely si uvnitř eucytu uchovaly schopnost samostatně se rozmnožovat replikací DNA a dělením. Biomembrány na jejich povrchu mají oproti biomemránám karyolemy různý původ. Vnitřní membrána má podobné funkce a látkové složení jako měla cytoplazmatické membrána původního protocytu, vnější membrána odvozuje svůj původ podle chemického složení nejspíše od cytoplazmat. Membrány eucytu. MITOCHONDRIE - jsou zpravidla kulovité, oválné nebo tyčinkovité organely vyskytující se téměř u všech eucytů v počtu od 50 do
5000. Dosahují šířky okolo 0,5 mikrometru, délky až 7 mikrometrů. Obal je tvořen dvěma vrstvami, vnější a vnitřní mitochondriální membránou, mezi kterými je vnější neplazmatický prostor mitochondrie. Z vnitřní membrány se tvoří do vnitřního četné membránové vchlípeniny zvané mitochondriální kristy podobné tylakoidům, od kterých se ale liší funkcí a tím, že jejich vnitřní prostor přechází v neplazmatický prostor mitochondrie. Plazmatický prostor je vyplněn matrixem, jejíž součástí je molekula DNA, RNA a větší zrnité útvar, dále se zde vyskytují enzymy tzv. Krebsova cyklu. Vnější biomembrána je volně průchodná pro ionty i molekuly, včetně menších molekul bílkovin. Vnitřní membrána je průchodná jen pro molekuly látek, pro které je vybaveny specifickými přenašeči. Podílí se na řízeném transportu O2 a CO2.
Hlavní funkcí mitochondrií je oxidace a fosforylace organ. látek za účelem uvolnění energie z jejich chemických vazeb a převedení této energie do tzv. makroergních vazeb v molekulách ATP podle rovnice:
5
ADP + H3PO4 + energie = ATP + H2O ATP pak slouží k přenosu energie do všech míst v buňce, kde je potřebná. Mitochondrie jsou schopny syntetizovat část bílkovin potřebných k plnění jejích funkcí a mitochondriální granula slouží k ukládání zásob vápníku v buňce. c) ostatní membránové struktury ENDOPLAZMATICKÁ SÍŤ (= RETIKULUM)
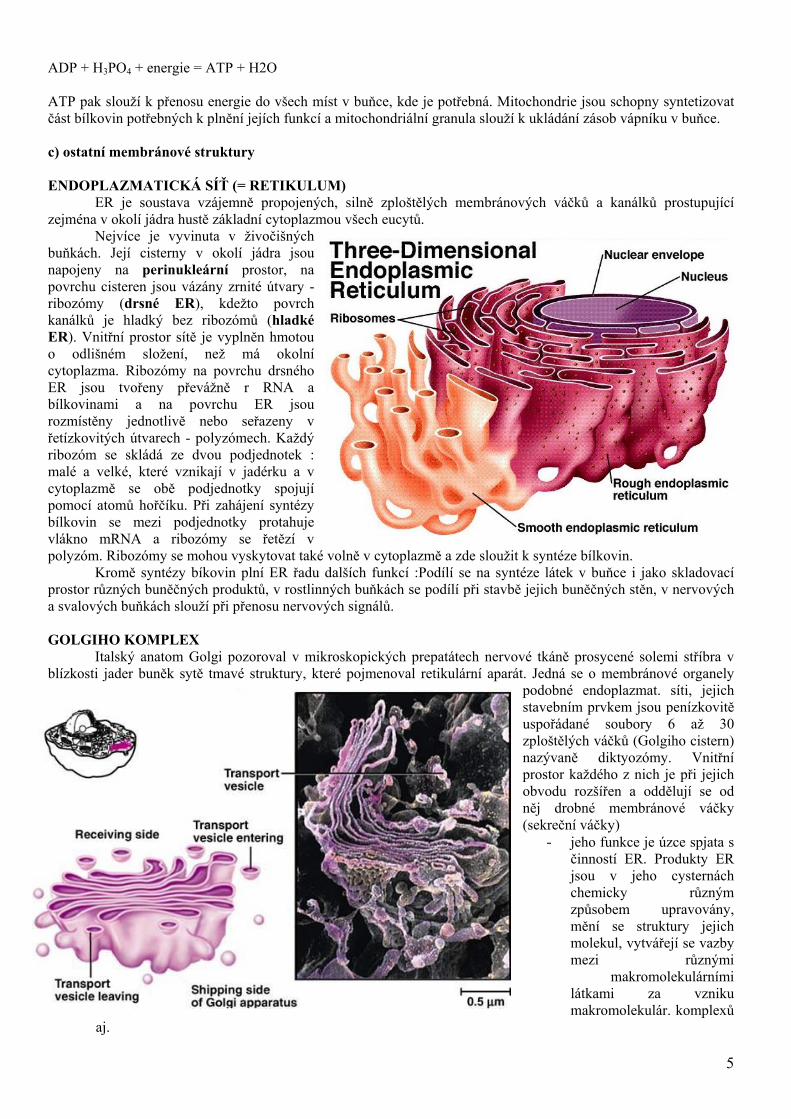
ER je soustava vzájemně propojených, silně zploštělých membránových váčků a kanálků prostupující zejména v okolí jádra hustě základní cytoplazmou všech eucytů.
Nejvíce je vyvinuta v živočišných buňkách. Její cisterny v okolí jádra jsou napojeny na perinukleární prostor, na povrchu cisteren jsou vázány zrnité útvary - ribozómy (drsné ER), kdežto povrch kanálků je hladký bez ribozómů (hladké ER). Vnitřní prostor sítě je vyplněn hmotou o odlišném složení, než má okolní cytoplazma. Ribozómy na povrchu drsného ER jsou tvořeny převážně r RNA a bílkovinami a na povrchu ER jsou rozmístěny jednotlivě nebo seřazeny v řetízkovitých útvarech - polyzómech. Každý ribozóm se skládá ze dvou podjednotek : malé a velké, které vznikají v jadérku a v cytoplazmě se obě podjednotky spojují pomocí atomů hořčíku. Při zahájení syntézy bílkovin se mezi podjednotky protahuje vlákno mRNA a ribozómy se řetězí v polyzóm. Ribozómy se mohou vyskytovat také volně v cytoplazmě a zde sloužit k syntéze bílkovin.
Kromě syntézy bíkovin plní ER řadu dalších funkcí :Podílí se na syntéze látek v buňce i jako skladovací prostor různých buněčných produktů, v rostlinných buňkách se podílí při stavbě jejich buněčných stěn, v nervových a svalových buňkách slouží při přenosu nervových signálů. GOLGIHO KOMPLEX
Italský anatom Golgi pozoroval v mikroskopických prepatátech nervové tkáně prosycené solemi stříbra v blízkosti jader buněk sytě tmavé struktury, které pojmenoval retikulární aparát. Jedná se o membránové organely
podobné endoplazmat. síti, jejich stavebním prvkem jsou penízkovitě uspořádané soubory 6 až 30 zploštělých váčků (Golgiho cistern) nazývaně diktyozómy. Vnitřní prostor každého z nich je při jejich obvodu rozšířen a oddělují se od něj drobné membránové váčky (sekreční váčky)
- jeho funkce je úzce spjata s činností ER. Produkty ER jsou v jeho cysternách chemicky různým způsobem upravovány, mění se struktury jejich molekul, vytvářejí se vazby mezi různými
makromolekulárními látkami za vzniku makromolekulár. komplexů
aj.
6
- takto upravené látky jsou sekrečními váčky uvolňovány do cytoplazmy a touto dopravovány na určená místa v buňce, kde plní své funkce, nebo jsou z buňky vylučovány. Cestou odškrcování membrán. váčků od Golgiho cistern vznikají také samostatné membrán. organely buňky jako jsou lyzozómy nebo cytozómy.
Z ER se dopravují látky do Golgiho aparátu prostřednictvím váčků, které se odškrcují od biomembrán ER, putují cytoplazmou k povrchové membráně Golgiho cisteren, se kterou splývají. Golgiho aparát produkty ER také zahušťuje a přebytečnou vodu vylučuje z buňky pomocí kanálků, které jsou napojeny na cytoplazmat. membránu (osmoregulační funkce GA) - mezi GA, ER a plazmat. membránou existuje úzká morfologická souvislost a funkční návaznost. LYZOZÓMY A NITROBUNĚČNÉ TRÁVENÍ
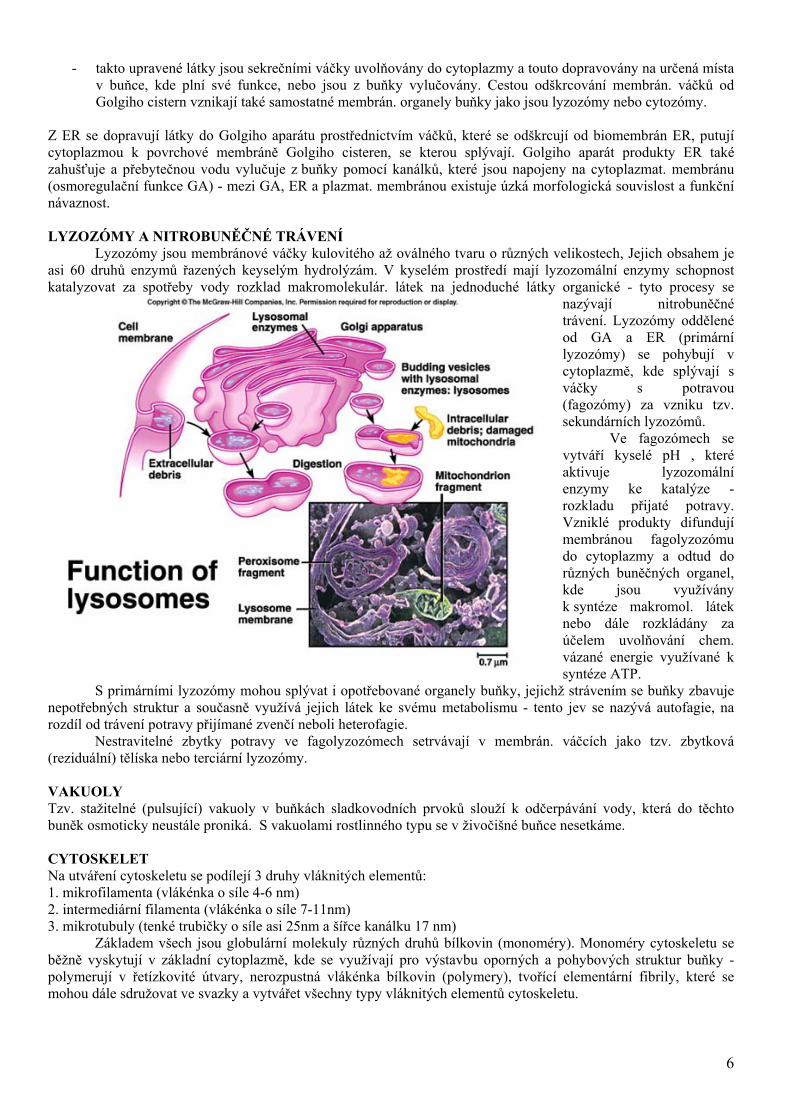
Lyzozómy jsou membránové váčky kulovitého až oválného tvaru o různých velikostech, Jejich obsahem je asi 60 druhů enzymů řazených keyselým hydrolýzám. V kyselém prostředí mají lyzozomální enzymy schopnost katalyzovat za spotřeby vody rozklad makromolekulár. látek na jednoduché látky organické - tyto procesy se
nazývají nitrobuněčné trávení. Lyzozómy oddělené od GA a ER (primární lyzozómy) se pohybují v cytoplazmě, kde splývají s váčky s potravou (fagozómy) za vzniku tzv. sekundárních lyzozómů.
Ve fagozómech se vytváří kyselé pH , které aktivuje lyzozomální enzymy ke katalýze - rozkladu přijaté potravy. Vzniklé produkty difundují membránou fagolyzozómu do cytoplazmy a odtud do různých buněčných organel, kde jsou využívány k syntéze makromol. látek nebo dále rozkládány za účelem uvolňování chem. vázané energie využívané k syntéze ATP.
S primárními lyzozómy mohou splývat i opotřebované organely buňky, jejichž strávením se buňky zbavuje nepotřebných struktur a současně využívá jejich látek ke svému metabolismu - tento jev se nazývá autofagie, na rozdíl od trávení potravy přijímané zvenčí neboli heterofagie.
Nestravitelné zbytky potravy ve fagolyzozómech setrvávají v membrán. váčcích jako tzv. zbytková (reziduální) tělíska nebo terciární lyzozómy.
VAKUOLY Tzv. stažitelné (pulsující) vakuoly v buňkách sladkovodních prvoků slouží k odčerpávání vody, která do těchto buněk osmoticky neustále proniká. S vakuolami rostlinného typu se v živočišné buňce nesetkáme. CYTOSKELET Na utváření cytoskeletu se podílejí 3 druhy vláknitých elementů: 1. mikrofilamenta (vlákénka o síle 4-6 nm) 2. intermediární filamenta (vlákénka o síle 7-11nm) 3. mikrotubuly (tenké trubičky o síle asi 25nm a šířce kanálku 17 nm)
Základem všech jsou globulární molekuly různých druhů bílkovin (monoméry). Monoméry cytoskeletu se běžně vyskytují v základní cytoplazmě, kde se využívají pro výstavbu oporných a pohybových struktur buňky - polymerují v řetízkovité útvary, nerozpustná vlákénka bílkovin (polymery), tvořící elementární fibrily, které se mohou dále sdružovat ve svazky a vytvářet všechny typy vláknitých elementů cytoskeletu.
7
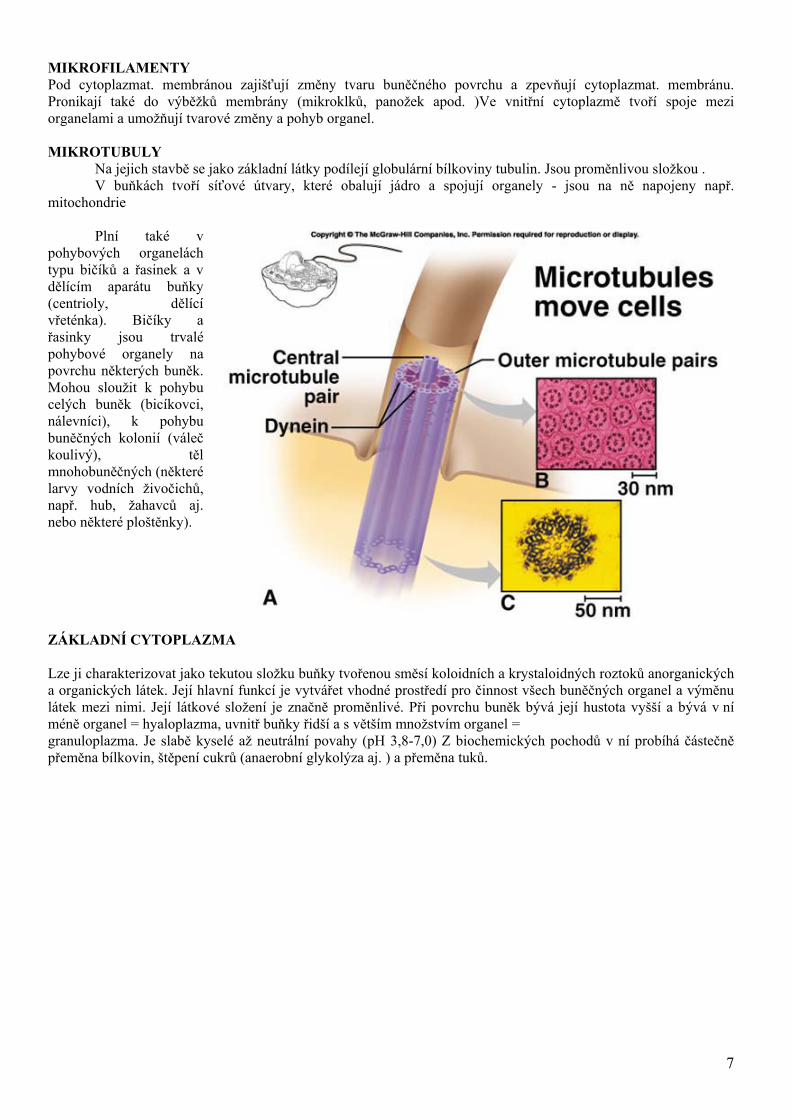
MIKROFILAMENTY Pod cytoplazmat. membránou zajišťují změny tvaru buněčného povrchu a zpevňují cytoplazmat. membránu. Pronikají také do výběžků membrány (mikroklků, panožek apod. )Ve vnitřní cytoplazmě tvoří spoje mezi organelami a umožňují tvarové změny a pohyb organel. MIKROTUBULY Na jejich stavbě se jako základní látky podílejí globulární bílkoviny tubulin. Jsou proměnlivou složkou .
V buňkách tvoří síťové útvary, které obalují jádro a spojují organely - jsou na ně napojeny např. mitochondrie
Plní také v
pohybových organelách typu bičíků a řasinek a v dělícím aparátu buňky (centrioly, dělící vřeténka). Bičíky a řasinky jsou trvalé pohybové organely na povrchu některých buněk. Mohou sloužit k pohybu celých buněk (bicíkovci, nálevníci), k pohybu buněčných kolonií (váleč koulivý), těl mnohobuněčných (některé larvy vodních živočichů, např. hub, žahavců aj. nebo některé ploštěnky).
ZÁKLADNÍ CYTOPLAZMA Lze ji charakterizovat jako tekutou složku buňky tvořenou směsí koloidních a krystaloidných roztoků anorganických a organických látek. Její hlavní funkcí je vytvářet vhodné prostředí pro činnost všech buněčných organel a výměnu látek mezi nimi. Její látkové složení je značně proměnlivé. Při povrchu buněk bývá její hustota vyšší a bývá v ní méně organel = hyaloplazma, uvnitř buňky řidší a s větším množstvím organel = granuloplazma. Je slabě kyselé až neutrální povahy (pH 3,8-7,0) Z biochemických pochodů v ní probíhá částečně přeměna bílkovin, štěpení cukrů (anaerobní glykolýza aj. ) a přeměna tuků.
8
1.2 VÝVOJ JEDINCE začíná oplozením vajíčka a končí zánikem jedince 2 části : 1) embryonální období – vyvíjí se a roste
2) postembryonální období – jedinec dospívá a rozmnožuje se 1.2.1 EMBRYONÁLNÍ VÝVOJ - EMBRYOGENEZE - oplozené vajíčko (zygota, synkaryon) – 2 sady chromozomů hned po oplození vstupuje do I .BLASTOGENEZE (vývoj zárodečného listu)
a) rýhování – může nastat už několik minut po oplození (mitóza) → vznikají 2 dceřiné buňky – blastomery → navazuje několik navazujících mitóz (→ odpadá interfáze) ⇒ počet buněk narůstá ale velikost útvaru zůstává stejná (buňky se zmenšují) → vzniká morula (může dojít k různému dělení na pólech → různá velikost buněk) → morula se vyvíjí vzniká blastula (uvnitř se začíná vytvářet dutina blastocel)
b) gastrulace- (vmáčknutí) ⇒ vzniká gastrula
vzniká i) přesouváním (imigrace) buněk ii) vchlipování (invaginace) u různých živočichů různé způsoby ⇒ vznikají 2-3 zárodečné listy
• pokud živočichové zůstávají na tomto stupni vývoje diblastika (gastrulovci) – 2 zárodečné listy (uvnitř endoderm, povrch ektoderm)
• může se vytvořit mezoderm (3.zárodečný list) → triblastika • mezoderm – vytváří se mezi ektodermem a endodermem, může mít povahu • parenchymu, epitelární → mezenchym (slouží jako výstelky), z mezodermu vznikají pohlavní orgány,
ledviny, svalstvo
c) nerulace – typické u obratlovců nebo strunatců, vytváří se chorda a mícha
II. ORGANOGENEZE zahájena intenzivním růstem zárodku, který začíná dostávat rysy dospělce dochází k diferenciaci buněk ( v souvislosti s vývojem zárodečných listů) dochází k protahování a ohýbání těla do určitého tvaru z jednotlivých zárodečných listů vznikají orgány
1) EKTODERM – z něj vzniká pokožka + její deriváty, dřeň nadledvin, vylučovací orgány bezobratlých,
vzdušnice, vnější žábra (u pavoukovců), výstelka přední a zadní části trávicí trubice, nervová soustava 2) ENDODERM – výstelky střední části střeva, trávicí žlázy, plíce obratlovců, štítná žláza, chorda dorsalis 3) MEZODERM – nejdříve se rozliší celomové váčky, které se rozdělí na hřbetní části na tzv. somity a v břišní
POSTEMBRYONÁLNÍ VÝVOJ začíná uvolněním zárodku z vaječných obalů, což probíhá 2 způsoby
A) NEPŘÍMÝ VÝVOJ (vývoj s proměnou) – mezi embryem a dospělcem je stádium larvy
Vývoj hmyzu z vajíčka → larva → svlékání, po posledním svlékáním vzniká imago (dospělec) = larva podobná dospělci – menší, bez pohlavních orgánů i) hmyz s proměnou nedokonalou – (rovnokřídlí, švábi) – larvy (najády) – nymfy ii) hmyz s proměnou dokonalou – vloženo stadium pupy (kukly) – dochází k metamorfóze (proměně) →
rozpuštění tkání → přestavba → dospělec → kukla proměna → dospělec s jinými znaky (motýli) Typy larev : housenky, housnice, ponravy, strusky
B) PŘÍMÝ VÝVOJ – zárodek podobný dospělci
• Zárodek je připraven na samostatný život, podobný dospělci, roste, dospívá, vyvíjí se pohlavní orgány • Vývoj v těle matky – aby zárodek byl co nejlépe připraven k samostatnému životu
9
Zygota se mění v trotoblast a amnioblast, z části buněk vznikají zárodečné obaly a z části plodová voda Vytváří se amniová dutina –ohraničena blánami, bývá naplněna tekutinami (u obratlovců – plodová voda), 2 blány : amnion (vnitřní) chorion (seroza) – vnější
• vytváří se až u amniot = plazi, ptáci, savci (plazi, ptáci –vajíčka vyvíjející se mimo tělo matky, savci-přímý porod + mateřské mléko)
savci – vyvíjí se uvnitř matky → porod → v prvních dnech vývoje mimo tělo matky – výživa pomocí mateřského mléka (typické složení pro jednotlivé druhy – bílo/masožravci) • porod –blány prasknou → zárodek vypuzen z těla ven s placentou, která zajišťuje výživu zárodku před
porodem (vzniká propojením sliznice s výběžky chorionu) Jednobuněčnost je předstupněm mnohobuněčnosti. Přechod od prvoků k mnohobuněčným živočichům představují buněčné kolonie (např. váleč koulivý, trubénky atd.) Při pohlavním rozmnožování je na počátku ontogenetického vývoje každého mnohobuněčného živočicha jediná buňka – oplozená vaječná buňka, zygota. Ta se opakovaně mitoticky dělí, což nazýváme rýhování zygoty. Buňky nadále nerostou, ale opakovaně se mitoticky dělí a jsou stále menší. Nazývají se blastomery, vytváří kulovitý zárodek z jediné vrstvy buněk, tzv. blastulu.
Představu o vzniku mnohobuněčnosti podává několik teorií: • Invaginační teorie – vchlipováním ektodermu (Ernst Haeckel) • Imigrační teorie – vcestováním buněk ektodermu do blastocelu (Ilja Mečnikov)
10
Dvouvrstevní živočichové mají vyvinuty dva zárodečné listy: ektoderm a entoderm (houby, žahavci, žabernatci). Tělní souměrnost kulovité dvouvrstevné gastruly je paprsčitá (radiální). Kulovitá gastrula se protahuje do délky, vzniká dvouvrstevná válcovitá gastrula (ploštěnci, hlísti). Tělní souměrnost je dvoustranná (bilaterální). Živočichové, u kterých se ústní otvor vyvíjí ze zbytku prvoúst gastruly a řitní otvor vzniká na opačném konci těla se nazývají prvoústí (protostomia). Živočichové, u kterých se v místě prvoúst vyvíjí otvor řitní a na opačném konci zárodku vzniká ústní otvor se nazývají druhoústí (deuterostomia). U vyšších živočichů vývoj pokračuje dále. Gastrula zůstává válcovitá, ale vyvíjí se třetí zárodečný list mezoderm. Z hřbetní části válcovitého entodermu se tvoří mezodermální váčky. Vznikají ve třech párech za sebou. Článkovaní živočichové mají v každém tělním článku jeden pár coelomových (mezodermálních) váčků. Základem zárodku jsou zárodečné listy: ektoderm, entoderm, mezoderm. Z nich se vyvíjí tkáně a orgány.
- výstelka tělních dutin - svalová a pojivová tkáň - pohlavní a močové orgány - škára, kostra
11
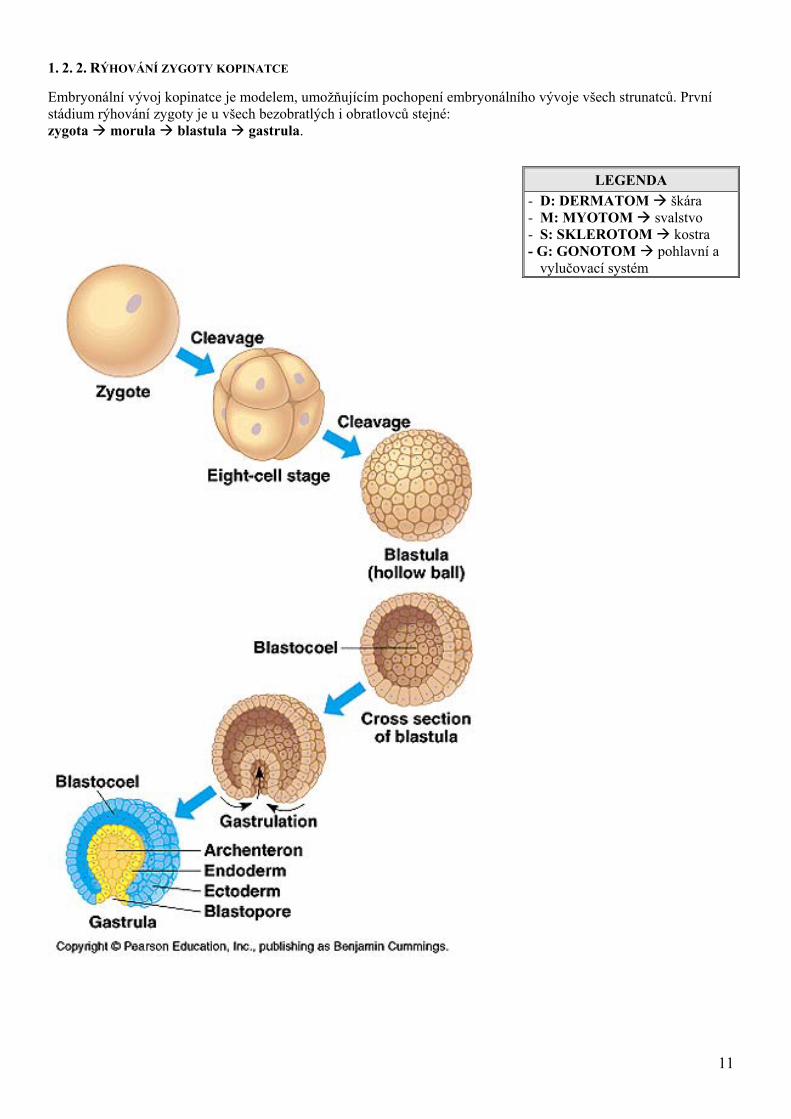
1. 2. 2. RÝHOVÁNÍ ZYGOTY KOPINATCE
Embryonální vývoj kopinatce je modelem, umožňujícím pochopení embryonálního vývoje všech strunatců. První stádium rýhování zygoty je u všech bezobratlých i obratlovců stejné: zygota morula blastula gastrula.
LEGENDA - D: DERMATOM škára - M: MYOTOM svalstvo - S: SKLEROTOM kostra - G: GONOTOM pohlavní a
vylučovací systém
12
1. 3. Tkáně Tkáně = soubor buněk stejného tvaru a fce (x nervová tkáň - b. hlavní a pomocné) - soudržnost buněk ve tkáních zajištěna fyzikálními, chemickými a fyzikálně
chemickými vazbami a mezibuněčnými spoji
- začínají se formovat z buněk zárodečných listů v časných fázích embr. vývoje - rozlišovat se začínají ve stadiu neuruly
- v prostorách mezi buňkami - tkáňový mok - zprostředk. látkovou výměnu mezi tělními tekutinami a tkáněmi
- činností buněk vzniká buněčná hmota : a) fibrilární - tvořena kolagenními, retikulárními a elastickými vlákny b) amorfní - tvořena glukózaminoglykany - želatinovitá, polotekutá
- ektoderm. původu - epitely krycí, smyslové, počátek a konec trávicí trubice, žlázové epitely, zubní tkáň, nervová, epitely dých. org. - entoderm. původu - epitely tráv. trubice, jater a slinivky břišní, plíce - mezoderm - epitely ledvin, gonád, výstelky tělních dutin, svalová tkáň
1. 3. 1. EPITELY
- fce - kryje povrch těla a orgánů, vystýlá dutiny - nejvíce se podobají zárodeč. listům, většina je ektoderm. a entoderm. původu - málo mezib. hmoty, buňky těsně vedle sebe - dělení podle : tvaru buněk - plochý = dlaždicovitý - kubický - cylindrický
počtu vrstev - jednovrstevný - víceřadý - vrstevnatý - přechodný Dělení podle funkce:
a) Krycí – na povrchu, chrání organismus před různými vlivy (např. pokožka)
b) Výstelkové- podobná fce jako krycí – výstelky všech dutin uvnitř těla (sliznice)
c) Epitely žlázové- speciálně upraveny , velké buňky hruškovitého tvaru, obsahují speciální látky- sekrety 3 základní typy sekrece : merokrinní – sekrety odváděny spec. kanálky- sekrece plynulé nebo v intervalech ( potní žlázy) apokrinní- sekret se shromažďuje na jednom póli buňky, odstraňuje se i s výměšky ( mléčné žlázy) holokrynní- sekret se hromadí v celé buňce,, naplněná buňka je vypuzena z epitelu a rozpadnutím uvolňuje sekret do okolí žlázy – vznikají shlukováním žlázových buněk, rozeznáváme: alveolární, tubulární, tubuloalveolární exokrynní- (s vnější sekrecí, na povrch epitelu) endokrynní- (s vnitřní sekrecí. Do krevního oběhu
d) Resorpční- různá fce (dých,..),vylučovací soustava e) Zárodečné- vyživují vyvíjející se zárodečné buňky f) Smyslové- schopny přijímat podráždění z okolí, např. u ploštice
Buňky pojivové tkáně jsou odděleny mezibuněčnou hmotou, která je jejich produktem. Spojují nebo izolují orgány, vyplňují prostor mezi nimi a poskytují oporu měkkým tkáním. 1. 3. 2. 1. Vazivová tkáň Podle mezibuněčné hmoty rozeznáváme 3 typy vaziv: • Řídká: převládá rosolovitá hmota • Hustá: převládá vláknitá složka (pouzdra orgánů, fascie…) • Uspořádaná hustá: tvoří šlachy a vazy Lymfoidní vaziva jsou vaziva obsahující bílé krvinky (lymfocyty). 1. 3. 2. 2. Chrupavčitá tkáň Je tužší než vazivo, odolná vůči tlaku. Chrupavky se vyznačují silným rozvojem mezibuněčné hmoty. Základem chrupavčité buňky jsou chondrocyty, které jsou uzavřené ve zvláštních schránkách. Chrupavka je obvykle bezcévná. Výživa probíhá z kosti, k níž je chrupavka přirostlá, a vazivové blanky, tzv. ochrustavice (perichondrium), která chrupavku pokrývá. Regenerační schopnost chrupavky je velmi malá. Podle mezibuněčné hmoty rozeznáváme 3 typy chrupavek: • Chrupavčitá tkáň sklovitá (hyalinní) V tenkých plátcích je průhledná, ve větší vrstvě bílá. Tvoří kloubní chrupavky, konce žeber, výstuže dýchacích cest, je základem kostí. V základní hmotě, která je stejnorodá, se nachází skupiny chrupavčitých buněk. Mezibuněčná hmota se nazývá kolagen. • Chrupavčitá tkáň vazivová Je matně bílá, v tenkých vrstvách neprůhledná. V mezibuněčné hmotě se nachází četné fibrily kolagenního typu. Tvoří meziobratlové ploténky a vnitrokloubní destičky. • Chrupavčitá tkáň elastická Má žlutobílou barvu, v tenkých vrstvách je neprůhledná. V mezibuněčné hmotě se nachází elastické fibrily dodávající pružnost. Tvoří např. podklad ušního boltce, hrtanovou příklopku aj. 1. 3. 2. 3. Kostní tkáň Kostní tkáň je nejtvrdší z pojiv. Vyskytuje se pouze u obratlovců. Je tvořena z kostních buněk, tzv. osteocytů. Obsahuje: - látky organické: asi 1/3, dodávají kosti pružnost, bílkovina osein - látky anorganické: asi 2/3, dodávají kosti pevnost, Ca3(PO4)2, CaCO3, Stářím se složení mění, přibývá anorganické hmoty snadná lámavost.
• Kostní tkáň houbovitá (spongiózní) Má strukturu jako mycí houba, obsahuje systém trámečků. Nachází se v hlavicích dlouhých kostí, v kostech krátkých, plochých aj. • Kostní tkáň hutná (kompaktní) Má lamelární strukturu. Výskyt v tělech dlouhých kostí. Nejběžnějším typem lamel jsou Haversovy systémy:lamely tvořené osteocycy jsou koncentricky uspořádány kolem kanálků, probíhajících rovnoběžně s osou kosti. V těchto kanálcích, jsou uloženy cévy. Kolem kanálků 8-15 lamel. Haversovy kanálky jsou navzájem spojeny šikmými spojkami a s povrchem kosti jsou spojeny tzv. Volkmanovými kanálky. Stavba dlouhé kosti: Kost je zevně kryta vazivovou blanou okosticí (periost), ta je bohatě prokrvena a obstarává růst kosti do šířky přirůstáním nových lamel. Kostní dřeň má v mládí červenou barvu. S přibývajícím věkem žloutne až šedne. V červené kostní dřeni se tvoři krevní elementy. V dospělosti zůstává jen v těle obratlů a v hrudní kosti. Uspořádání trámečků houbovité kosti je závislé na tlaku a tahu (architektonika kosti). Trámečky nevystavované tlaku a tahu se odbourávají.
Vývoj kostí: Kosti se vytváří z chrupavčitého nebo vazivového základu kostnatěním (osifikací) již během embryonálního vývoje. Osifikace se šíří z tzv. osifikačních jader charakteristických pro danou kost. Růst kostí: - do šířky: pomocí okostice - do délky: uskutečňuje se na tzv. růstové chrupavce. Růst je ukončen mezi 18-20 rokem. U žen asi o 2 roky dříve. 1. 3. 2. 4. Trofické tkáně • Krev Krev je tekutý orgán, který zabezpečuje stálost vnitřního prostředí organismu. Je to neprůzračná červená tekutina, tvořená suspenzí krevních tělísek v krevní plazmě Složení krve: - tekutá krevní plasma - tuhé elementy: - erytrocyty = červené krvinky - leukocyty = bílé krvinky - trombocyty = krevní destičky Krevní plasma Krevní plasma je nažloutlá tekutina, slouží k přepravě glukózy, tuků…
Složení: - 91% H2O – volná (rozpouštědlo) a vázaná na bílkoviny - 8% organické látky: 1) krevní bílkoviny 7%: - albuminy - globuliny - fibrinogen Bílkoviny mají význam pro transport látek, které se na ně Váží: minerály, tuky, hormony. Poutají vodu v krvi.
Fibrinogen má význam pro srážení krve. 2) glukóza 1% Koncentrace glukózy v plasmě se nazývá glykémie (0,8 – 1,2 g/l). Její hladina je stálá. Glukóza slouží jako zdroj energie. Z krve je neustále odčerpávána, ale její hladina je neustále doplňována ze zásob glykogenu v játrech. Glykogen – polysacharid, tzv. živočišný škrob - 1% anorganické látky
Hlavně NaCl a NaHCO3. Jsou to osmoticky aktivní látky. Řídí osmotický tlak, který závisí na množství vody. Udržují i konstantní hodnotu pH. Další ionty, např.: Fe2+ - pro krvetvorbu Ca2+ - pro svalový stah a srážení krve I- - pro činnost štítné žlázy
Tuhé elementy Všechny se vyvíjí z krvetvorných buněk kostní dřeně
Červené krvinky (erytrocyty) Okrouhlé bezjaderné buňky obalené polopropustnou membránou. Bezjadernost snižuje jejich vlastní spotřebu kyslíku. Jsou to primitivní buňky bez vnitřních struktur (schránky na hemoglobin). Tvar: na průřezu piškotovitý (bikonkávní) zvětšení povrchu proti kouli stejného objemu o 30%. Funkce: přenos kyslíku z plic do tkání. Kyslík se v krvinkách váže na krevní barvivo, tzv. hemoglobin. Vzniká tzv. oxihemoglobin. Tato vazba je slabá kyslík se z ní snadno uvolňuje. Hemoglobin obsahuje ve své molekule ionty Fe2+, na které se váže kyslík jako magnet. Hemoglobin má schopnost vázat také CO, vzniká tzv. karboxyhemoglobin. Tato vazba je pevná, CO z ní nelze uvolnit. Hemoglobin zůstává trvale zablokován nastává smrt z nedostatku kyslíku. Vznik: v červené kostní dřeni (v dospělosti hrudní kost a těla obratlů). Životnost: 120 dnů Červené krvinky zanikají ve slezině, játrech a kostní dřeni. Po odštěpení železa se zbytek molekuly oxiduje na barvivo bilirubin, který se vylučuje játry do žluči. Krevní destičky (trombocyty)
15
Nemají jádro, tvar je nepravidelný, jedná se jen o úlomky kostní dřeně. 200 000 – 500 000 v 1mm3 krve. Obsahují látky nepostradatelné pro srážení krve (Ca2+, seratonin, enzymy). Vznik: v červené kostní dřeni.
Podstata srážení krve: bílkovina krevní plasmy, rozpustný fibrinogen přechází na nerozpustný fibrin (sítovitá struktura, v níž se zachycují krevní elementy)
enyzm trombin fibrinogen fibrin
Trombin je v plasmě přítomen jako neúčinný protrombin. Do krve se dostává z jater, kde se tvoří za přítomnosti vitamínu K. trombokináza
protrombin trombin Trombokináza se uvolňuje rozpadem krevních destiček. Bílé krvinky (leukocyty) Bezbarvé buňky s jádrem. Vznik v červené kostní dřeni, funkce obranná. Jsou obsaženy v krvi, tkáňovém moku, míze a v některých typech vaziva. Životnost různá: několik hodin, let i celý život. V 1mm3 7 000 – 8 000 leukocytů. Typy leukocytů: 1) Granulocyty:obsahují barvitelná zrníčka granuly 2) Agranulocyty: bez zrníček
GRANULOCYTY (70% leukocytů) se dělí dále podle typu barvitelnosti na: - granulocyty neutrofilní: zrna jsou barvitelná kyselými i zásaditými barvivy. Mají největší fagocytární
schopnost z granulocytů. - granulocyty eozinofilní: barvitelné kyselým barvivem eozinem. Mají menší fagocytární schopnost než
neutrofily. Zvýšená hladina při parazitických a alergických chorobách. - granulocyty bazofilní: barví se zásaditými barvivy. Působí jako miny – vybuchují a uvolňují chemické látky,
spouštějící zánětlivé procesy. Jejich granule produkují heparin, který zabraňuje srážení krve uvnitř cév.
AGRANULOCYTY dělí se na: - lymfocyty: jsou odpovědné za imunitu = odolnost organismu (T-Lymfocyty, B-Lymfocyty) - monocyty: největší bílé krvinky. mají schopnost diapedeze = pronikat mimo krevní řečiště. Dostávají se do
tkání jako tzv. makrofágy. Mají velkou fagocytární schopnost , pohlcují i větší částice.
• Míza Míza má podobné složení jako krevní plazma. Je chudší na bílkoviny, obsahuje proměnlivé množství lymfocytů. Má důležitou roli při vstřebávání tuků. • Tkáňový mok Vztah mezi krví, mízou a tkáňovým mokem: ve vlásečnicích dochází k výměně plynů, vody a látek mezi krví a tkáňovým mokem. Na začátku kapiláry prostupuje tekutina, obsahující živiny a jiné potřebné látky z krve do mezibuněčných prostor. Vytváří tkáňový mok. Na konci kapiláry se vrací tekutina obsahující zplodiny metabolismu zpět do krve.
16
1. 3. 3. SVALOVÁ TKÁŇ
Charakteristickou vlastností svalové tkáně je schopnost smršťování (kontrakce). Ta je umožněna přítomností myofibril. 1. 3. 3. 1. Hladká svalová tkáň Hladká svalová tkáň je tvořena buňkami vřetenovitého tvaru s jedním jádrem uprostřed. Délka buňky je 100 – 200 µm. Myofibrily probíhají přes celou buňku. V mikroskopu se proto buňky jeví jako stejnorodé, hladké. Buňky se přikládají k sobě a tvoří tenčí nebo silnější vrstvičky ve stěnách orgánů (střevo, cévy, kůže). Charakteristická je pomalá, déle trvající kontrakce. Hladká svalovina je řízena nervstvem, které nepodléhá naší vůli (vegetativní, útrobní nervstvo). 1. 3. 3. 2. Srdeční svalová tkáň Srdeční svalová tkáň tvoří svalovou stěnu srdce, tzv. myokard. Tato svalovina je schopna kontrakce i bez nervových popudů. Má vlastní automacii. Vegetativní vlákna její činnost nevyvolávají, pouze ji zpomalují nebo zrychlují. Srdeční svalová tkáň je tvořena prostorovou sítí buněk, která je rozdělená pomocí interkalárních disků na úseky s jedním nebo dvěma jádry. Prostory mezi síťovinou jsou vyplněny vazivem, které tvoří kostru srdce. 1. 3. 3. 3. Příčně pruhovaná svalová tkáň Příčně pruhovaná svalová tkáň se upíná na kostru a umožňuje pohyb. Skládá se z mnohojaderných svalových vláken. Svalové vlákno představuje svalovou buňky, která vznikla splýváním mnoha zárodečných buněk, tzv. myoblastů, již v embryonálním vývoji. Svalové vlákno je dlouhé několik milimetrů až centimetrů. Na povrchu svalového vlákna je obal, tzv. sarkolem, pod kterým se nachází četná jádra. Uvnitř svalového vlákna probíhají myofibrily. Každé svalové vlákno jich obsahuje několik set až tisíc. Nejsou jako u hladké svaloviny stejnorodé, ale skládají se z úseků temnějších a světlejších. Tyto úseky se pravidelně střídají a sval se tak jeví jako příčně pruhovaný. Podle struktury rozeznáváme dva typy svalových vláken: • Svalová vlákna červená (tonická) Obsahují více svalového hemoglobinu, tzv. myoglobinu. Tato vlákna jsou pomalejší, ale vytrvalejší. • Svalová vlákna bledá (fázická) Obsahují méně myoglobinu. Jsou rychlejší, ale rychleji se také unaví. Svaly člověka obsahují oba typy těchto vláken. Stavba příčně pruhovaného svalu: několik svalových vláken, obklopených vazivem, tvoří snopeček svalový. Více snopečků vytváří snopec. Snopce tvoří svalové bříško. To přechází ve šlachu, kterou se sval upíná na kost. Na povrchu snopečků, snopců a bříšek se nachází vazivo, kterým prochází cévy a nervy. Na povrchu svalu je svalová povázka. Činný sval je bohatě prokrvený.
17
1. 3. 4. NERVOVÁ TKÁŇ
Základní stavební i funkční jednotkou nervové soustavy je nervová buňka, tzv. neuron. Její tvar je rozmanitý. Nejčastější tvar: Uvnitř neuronů probíhají vlákna neurofibrily. Axon je obalen pláštěm z lipidů, který vytváří myelinovou pochvu. Zevně se nachází Schwannova pochva tvořená ze Schwannových buněk. Obě pochvy jsou spirálovitě svinuty kolem axonu. Pochvy jsou rozděleny tzv. Ranvierovými zářezy na segmenty asi 1 mm dlouhé.
Seskupením těl a dendritů vzniká šedá hmota nervová, seskupením axonů vzniká bílá hmota nervová. Základní funkcí je vznik a šíření vzruchů (impulzů). Jsou to elektrické změny zprostředkované pohybem iontů, nikoliv elektronů. Uvnitř neuronů je převaha záporně nabitých iontů, zevně převažují ionty kladné. Mezi vnitřkem a vnějškem existuje rozdíl potenciálů se záporným potenciálem uvnitř neuronu. Jeho hodnota –70 mV = klidový membránový potenciál. Uvnitř buňky je převaha iontů K+, ale celkově převažuje záporný náboj (bílkoviny, Cl-). Ve vnějším prostředí jsou přítomny ve velké koncentraci Na+ ionty. Při podráždění neuronu se mění propustnost plazmatické membrány pro Na+, ty vnikají do buňky a vyrovnávají záporný náboj. Ionty K+ vystupují z buňky. Vzniká akční potenciál, který se šíří po celé délce neuritu. (Rychlost šíření vzruchu až 120 m/s.) Obnovení původního stavu iontů se děje mechanismem aktivního transportu (sodíkodraslíková pumpa). Neurony se vzájemně propojují a dotýkají se svými výběžky. Místo tohoto spojení se nazývá zápoj (synapse). Činností synapsí se vzruchy přenáší z jednoho neuronu na druhý. (Na jednom neuronu 1000 i více synapsí.) Přenos vzruchu je možný jen jedním směrem – z presynaptické na postsynaptickou membránu. Dorazí-li vzruch na synapsi, váčky s mediátorem se rozpadnou a mediátor se dostává do štěrbiny mezi oběma neurony. Na membráně druhého neuronu nastává depolarizace (K+ vystupují z buňky, Na+ vstupují do buňky.) Mediátory: - tlumivé (inhibiční): kyselina gamaaminomáselná - budivé (excitační): acetylcholin - na synapsích sympatiku: noradrenalin Nervová tkáň obsahuje kromě neuronů i buňky gliové – mají funkci podpůrnou.
u jednoduchých → dělení buňky z mateřského jedince vznikají 2 nebo více jedinců, dceřiní jedinci mají geneticky shodný materiál jako mateřský
jedinec amitóza, mitóza
1) Prvoci – mitóza a) dělením – buňka se rozdělí na 2 stejné části nálevníci – dělí se příčně bičíkovci – dělí se podélně b) pučením – na mateřském jedinci se vytvoří pupen → zvětšení→ oddělení c) rozpadem (schizogonie) – několikanásobné dělení jádra, kolem jader okrsky cytoplazmy → náhlé rozdělení → velké množství jedinců 2) Mnohobuněční – tělo se může rozpadnout, pupen,…
a) dělení (fisiparie) – mateřský jedinec se rozdělí na 2 či více částí → dceřiné organismy ( žahavci, kroužkovci,..)
b) pučení (gemiparie) – typické pro houby, žahavce, pláštěnce mateřský + dceřiný jedinec zůstávají propojeni (korály) Pučení vnější (gematio) – kdekoli na těle pupen → roste
Pučení vnitřní (gaulatio) – sladkovodní houby, vnitřní pupeny, kam se soustředí část těla, vše se obalí amfidisky
c) Fragmentace – (houby, polypy, sasanky), z mateřského jedince se oddělují části těla, ze kterých dorůstají noví jedinci
1.4.2 POHLAVNÍ ROZMNOŽOVÁNÍ
- většina živočichů - vytvoření speciálních buněk (gamety), vznikají meiózou (redukční dělení) v pohlavních orgánech
1) Prvoci – k obohacení genetického materiálu → umožněn vývoj a) konjugace (spojení) – 2 buňky se k sobě připojí buněčnými ústy, makronukleus se rozpadá, mikronukleus se dělí → vzniká: menší jádro – samčí (migratorní) větší jádro – samičí (stacionární) dojde k výměně migratorních jader a následnému splynutí migratorního a stacionárního jádra → oddělí se → objeví se makronukleus → navazují mitózy b) kopulace – 2 jedinci se začínají chovat jako jeden – gomonti (splynutí těchto 2 buněk) , splynutím vzniká zygota, ta se dál nepohlavně rozmnožuje – vždy schizogonie
2) Mnohobuněční – OOGAMIE – pohlavní buňky jsou morfologicky odlišné (vajíčko, spermie) se vyvíjí v pohlavních orgánech (gonády)
19
SEPRMATOGENEZE – vývoj spermie V semenných kanálcích, které jsou vystlány Sertoliho vrstvou (zárodečný epitel), slouží k výživě. Nad Sertoliho vrstvou jsou pohlavní buňky, ty se dále dělí na spermatogonie. Ta se dělí mitózou a vznikají 2 spermatocyty prvního řádu. Ty vstupují do prvního zracího dělení a vznikají spermatocyty druhého řádu. Následuje 2. zrací dělení a vznikají spermatidy (spermie bez bičíku) dále se diferencují ve spermie Typy spermií – podobné bičíkovcům, vyjímečně bezbičíkaté spermie (korýši, roztoči, škrkavky) Bičíkaté – hlavička = buněčné jádro s genetickým materiálem krček –obsahuje mitochondrie bičík – obyčejná stavba např- explosivní spermie u mořských plžů – „autobusové spermie“ – spermie rozvážející ty pravé
OOGENEZE – vývoj vajíček , nižší počet vajíček Probíhá v Gráfově falikulu ve vaječnících Začátek – primordiální gonocyty z nich se vytváří oogonie (malé buňky s velkými jádry). U plazů, ptáků a savců se už během zárodečného vývoje oogonie mění mitózou na oocyty 1. řádu. Takto jsou uchovány až do pohlavní zralosti. Pak se postupně až 10 000x zvětší, potom meiotickým dělením vznikají oocyty 2. řádu a buňka pólová. Oocyty 2. řádu vstupují do druhého zracího dělení a vzniká ootida a buňka pólová a z první buňky pólové vznikají další 2 buňky pólové (využívají se pro výživu), ootida se mění na zralé vajíčko
Typy vajíček – Liší se tím jakým způsobem se do vajíček ukládají zásobní látky, ty jsou uloženy ve žloutku (žloutek obsahuje zásobní látky, které vyživují vyvíjející se zárodek – zrna vitelinu, nukleové kyseliny, bílkoviny, tuky), na žloutku vzniká animální ( budoucí zárodek) a vegetativní pól (výživa – soustředěn žloutek)
Vaječné obaly – mimo tělo matky
a) primární obaly – vylučovány vlastním oocytem, pružné membrány (žloutková blána – ptačí, žíhaná blána – savčí)
b) sekundární obaly – vznikají činností buněk orgánů , ve kterých se vajíčka vyvíjí, např. chorion (chitinózní obal vajíček hmyzu)
c) terciální obaly – vznikají až po oplození, činností přídatných žláz v pohlavních orgánech (skořápky, kožovité blány, bílek,..)
OSEMENĚNÍ – INSEMINACE Střetnutí spermie s vajíčkem
a) oplození vnější – spermie i vajíčka vypuštěna do vody v obrovské kvantitě b) vnitřní oplození – probíhá uvnitř těla samičky, spermie vnikají do pohlavního ústrojí samičky (menší
množství vajíček, hodně spermií) , biologický boj – jen nejschopnější spermie oplodní vajíčko
OPLOZENÍ
splynutí vajíčka a spermie, probíhá u většiny živočichů stejně spermie se dostane k vajíčku → přichytí na povrchu vajíčka → vajíčko vypouští speciální chemické látky podporující positivní chemotaxi spermií = gamony vajíčko vylučuje gynogamon I – urychluje pohyb spermií směrem k vajíčku, umožňuje
přilepení spermie k vajíčku ve chvíli, kdy pronikne první spermie gynogamon II – zabraňuje pronikání dalších spermií spermie vylučují androgamon I – funkce: tlumí pohyb ⇒ prodlužuje život spermií (zásobní látky se při pohybu
spotřebovávají – vyčerpání → odumření), je vylučován když gynogamon I není přítomen androgamon II – vylučován když se spermie dostane k vajíčku, umožňuje její proniknutí do vajíčka – rozpouští
jaderný obal ve vajíčku se rozpouští obal spermie → splynutí samiččího a samčího jádra ⇒ SYNKARYON
PARTENOGENEZE – vývoj neoplozeného vajíčka Vyskytuje se u mnohých živočichů ve chvíli, kdy jsou příhodné podmínky → vajíčka se začnou vyvíjet i bez
oplození Typické pro mšice, pakobylky, perloočky
20
Z genetického hlediska – nepohlavní rozmnožování
HERMAFRODITISMUS + GONOCHORISMUS
Hermafrodité – vytváří pohlavní buňky gonochorismus – jeden jedinec vytváří pouze samčí nebo samičí pohlavní buňky