Fyziologie a molekulová biologie živočišné buňky Pro magisterské studium biologie Katedra biologie Přírodovědecká fakulta UJEP Ústí n.L Obrazová příloha Strukturální a funkční kompartmentalizace (Endomembránový systém, mitochondrie, cytoskelet, jádro, cytosol) Pouze pro osobní potřebu účastníka konzultace Šíření v tištěné nebo digitální formě je v rozporu se zákonem o autorských právech citovaných zdrojů Mareš V Rupert M Aktualizováno 16.2.2016 Pokud není uveden jiný zdroj, materiál je z archivu autora
Transcript
Fyziologie a molekulová biologie živočišné buňkyPro magisterské studium biologie
Katedra biologiePřírodovědecká fakulta UJEP
Ústí n.L
Obrazová příloha Strukturální a funkční kompartmentalizace (Endomembránový systém, mitochondrie, cytoskelet, jádro, cytosol)
Pouze pro osobní potřebu účastníka konzultaceŠíření v tištěné nebo digitální formě je v rozporu se zákonem
o autorských právech citovaných zdrojů
Mareš VRupert M
Aktualizováno 16.2.2016 Pokud není uveden jiný zdroj, materiál je z archivu autora
Co je molekulová (molekulární?) biologie?
Předmět zabývající se makromolekulami buněk, především NK a bílkovin.Složením a stavbou, syntézou, metabolizmem, funkcemi atd.
Molekulání genetika
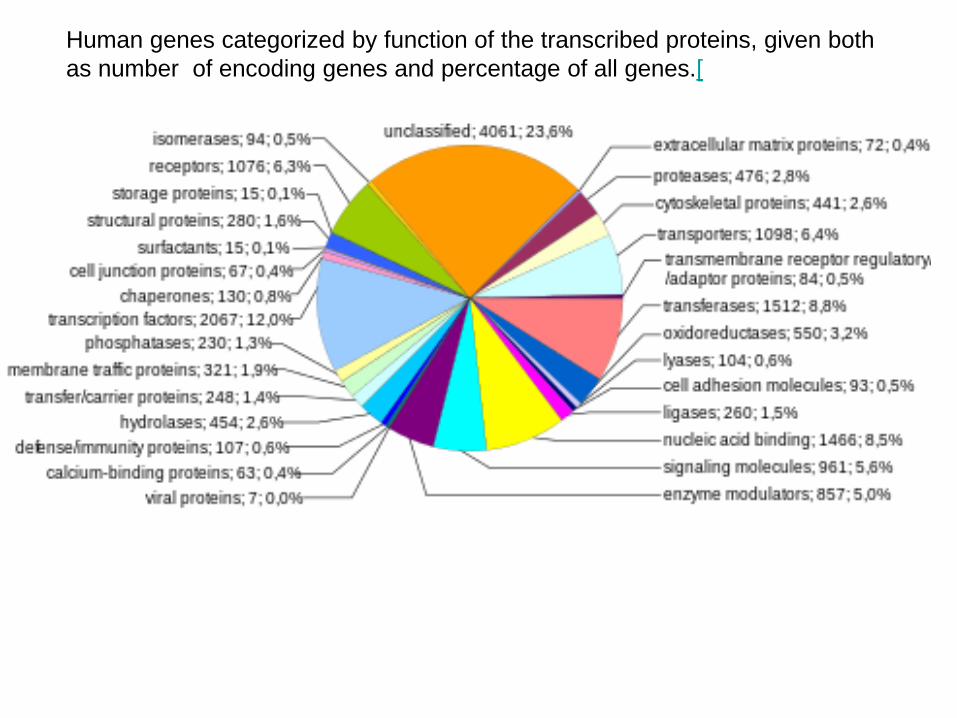
Funkce bílkovin jsou velmi rozsáhlé a stále málo známé
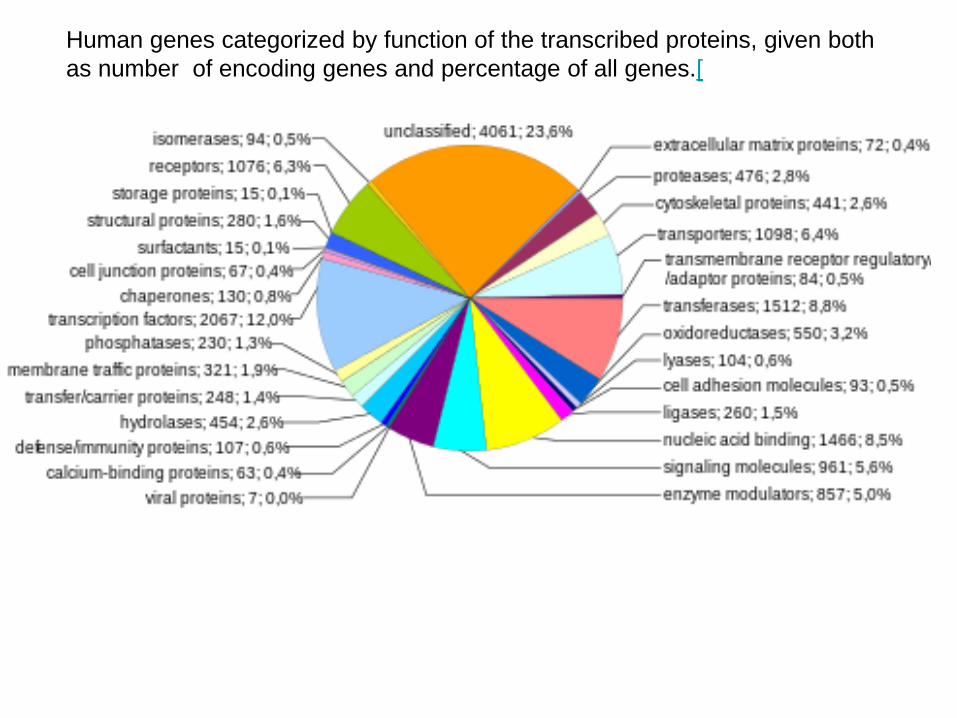
Human genes categorized by function of the transcribed proteins, given both as number of encoding genes and percentage of all genes.[
Společné znaky: Jednotková membrána (membrane unit)Tok látek a membrán mezi jednotlivými organelamiMetabolická kooperace „Dělba/specializace práce v buňce“
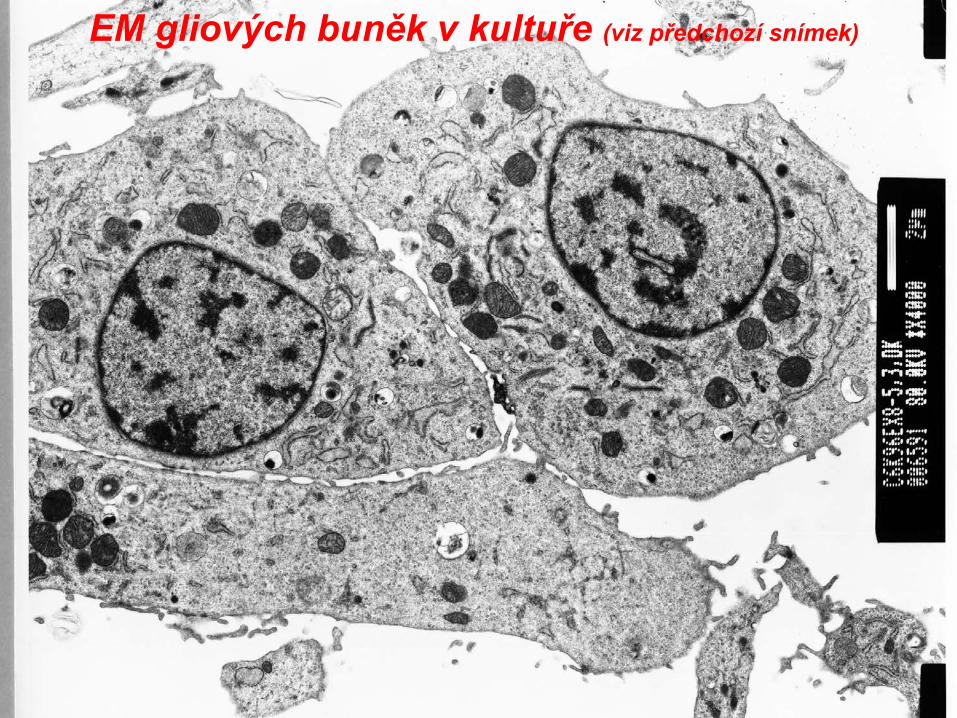
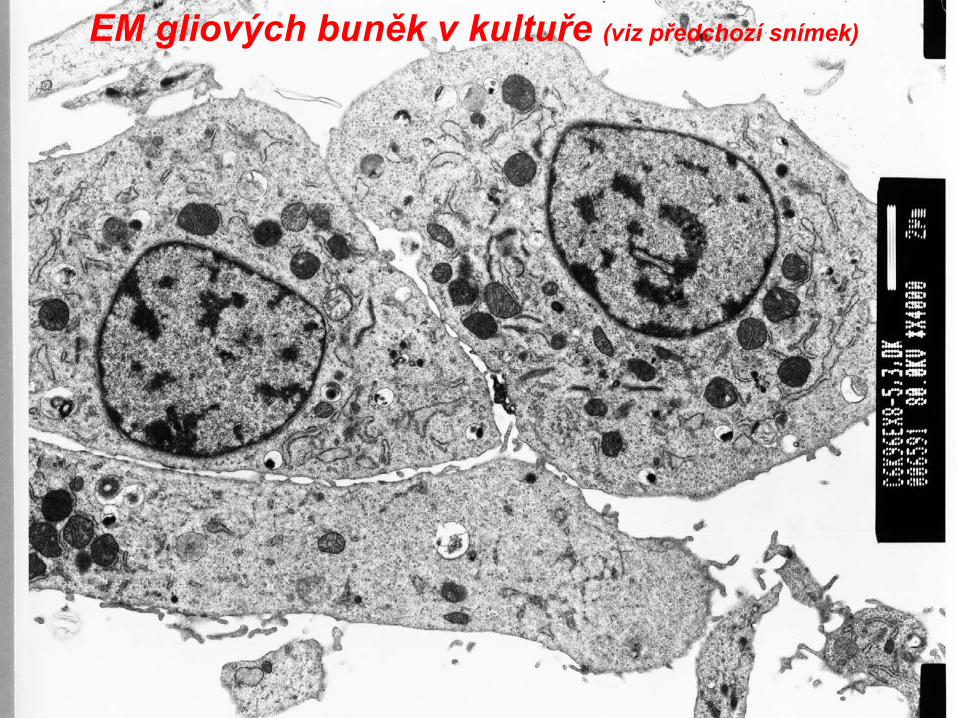
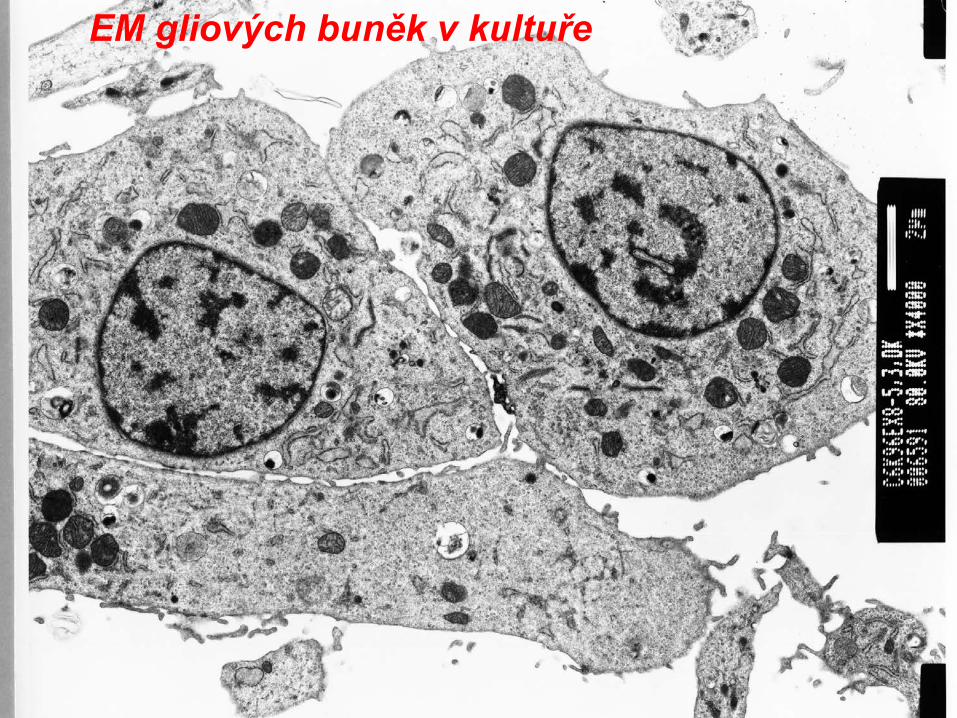
EM gliových buněk v kultuře (viz předchozí snímek)
EM gliových buněk v kultuře(část jádra,viz mitochondrie, hrER)
Ferenčík 2000
2 /um
0.05 /um
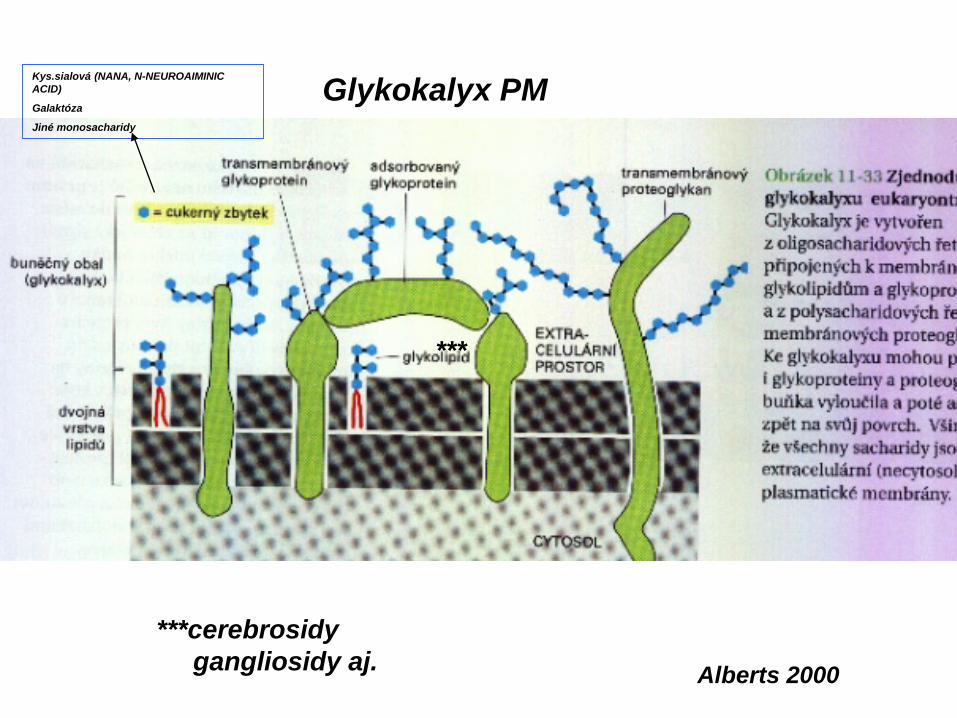
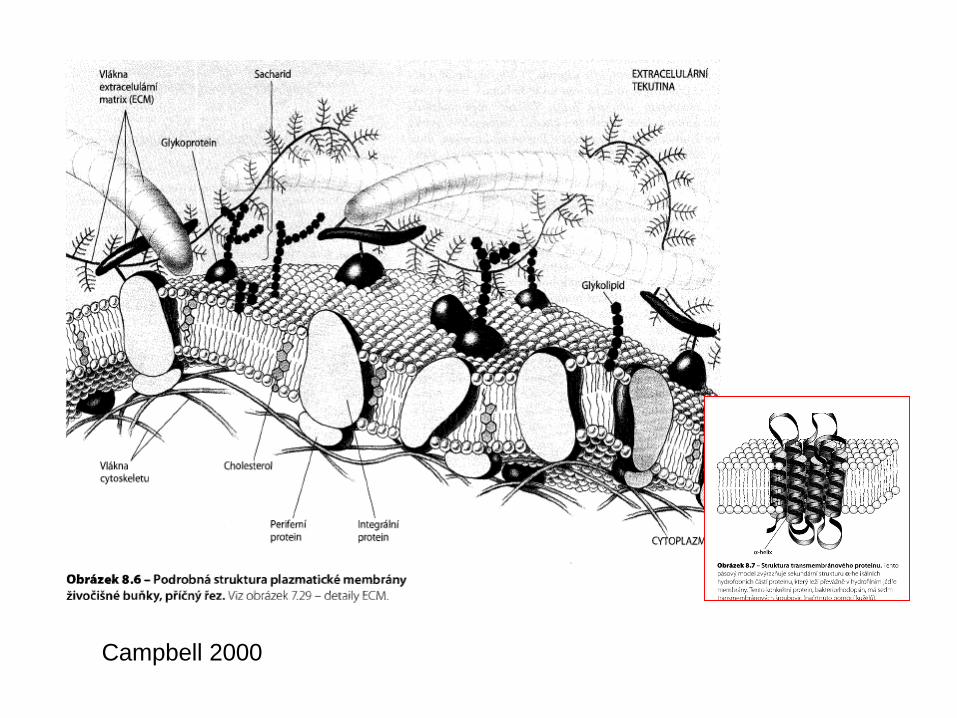
Plazmatická membrána
The cell membrane, also called the plasma membrane or plasmalemma, is a selectively permeable lipid bilayer which comprises the outer layer of a cell.
The plasma membrane consists of 1/3 cholesterol and 2/3 phospholipids (65-80%) andsphingolipids (20-35%). The outer leaflet contains 5% glycolipids.
Phospholipid molecules in the cell membrane are "fluid," in the sense that they are free to diffuse and exhibit rapid lateral diffusion. Lipid rafts and caveolae are examples of cholesterolenrichedmicrodomains in the cell membrane.
The cell membrane contains a large amount of protein. Individualproteins may be adjacent to the membrane or may be embeddedwithin it; such proteins are called peripheral and integralmembrane proteins, respectively.
Many integral proteins are not free to diffuse. The cytoskeleton undergirds the cell membraneand provides anchoring points for integral membrane proteins. Anchoring restricts them to a particular cell face or surface — for example, the "apical" surface of epithelial cells that line thevertebrate gut — and limits how far they may diffuse within the bilayer. Rather than presenting always a formless and fluid contour, the plasma membrane surface of cells may show
Returning to the example of epithelial cells in the gut, the apical surfaces of many such cells are dense with involutions,all similar in size. The finger-like projections, called microvilli, increase cell surface area and facilitate the absorption of molecules from the outside.structure. Returninge cell membrane contains a large amount of protein.
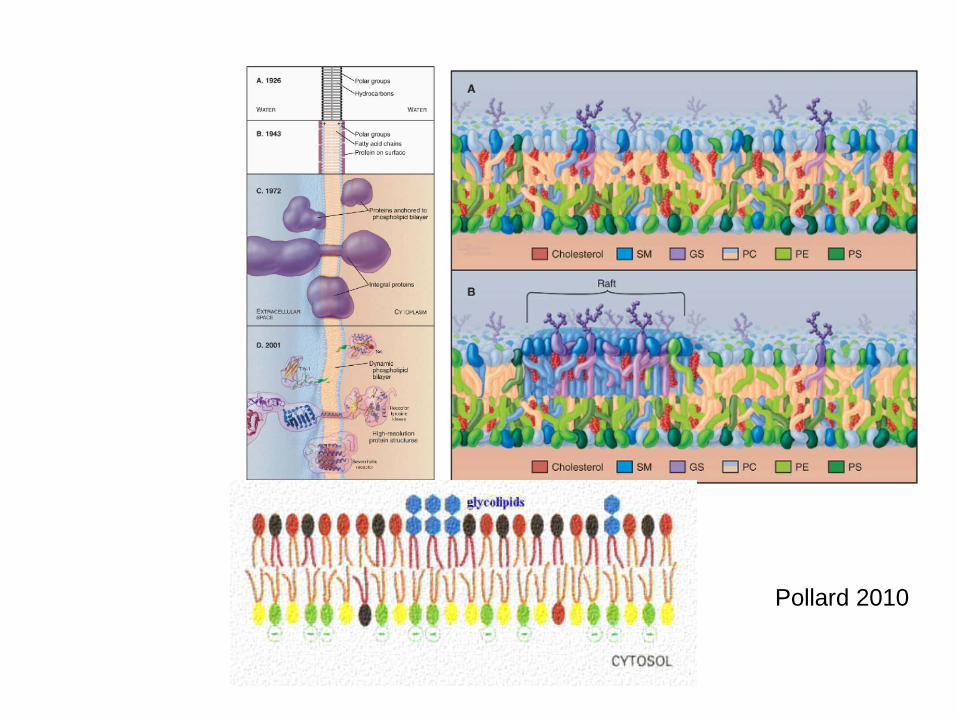
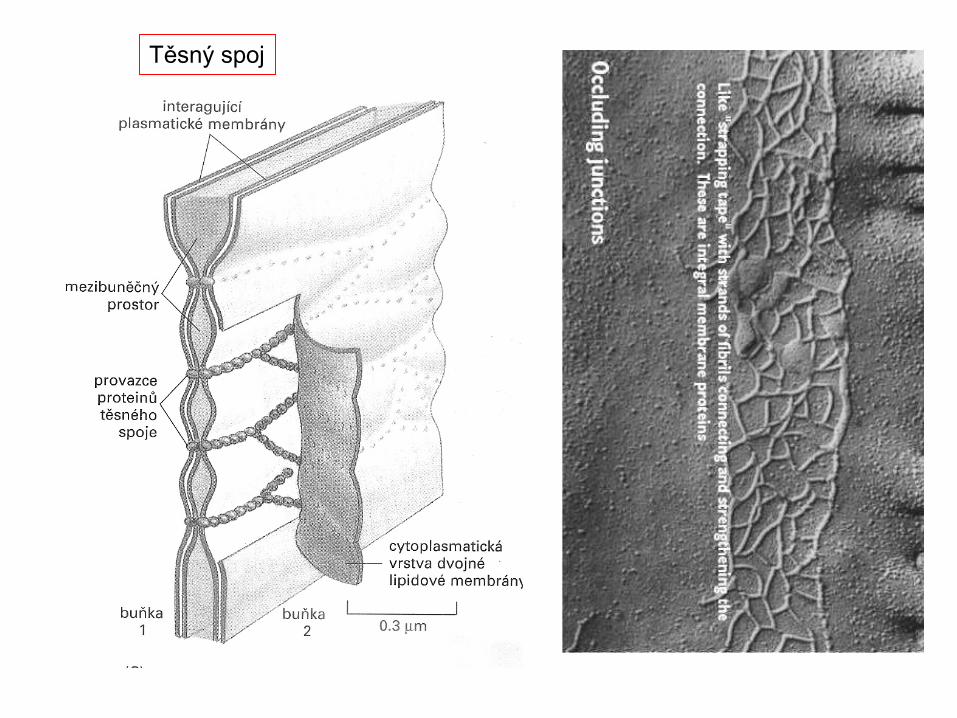
In the 1950's Robertson: He hypothesized that the railroad track appearance came from the binding of osmium tetroxide to proteins and polar groups of lipids.
V r 1972 Singer and Nicolson: PM je tekutou mozaikou kde proteiny jsou kromě hydrofilních konců zanořeny do lipidické dvojvrstvy, ve které se mou přemisťovat. Model fluidní mozaiky (viz shora a níže)
In the early 1930's-40's, Danielli and Davson studied triglyceride lipid bilayers over a water surface. They found that they arranged themselves with the polar heads
facing outward. However, they always formed droplets (oil in water) and the surface tension was much higher than that of
cells. However, if you added proteins, the surface tension was reduced and the membranes flattened out.
Historický vývoj struktury PM
Top Figure modified from Alberts et al. Molecular Biology of the Cell, Garland Publishing, N.Y., 1994, Third Edition, Figure 10-11
100 nm=10 mm
Top Figure modified from Alberts et al. Molecular Biology of the Cell, Garland Publishing, N.Y., 1994, Third Edition, Figure 10-11
Zevní vnitřní zevní - list lipidické dvojvrstvy PM
Cholesterol
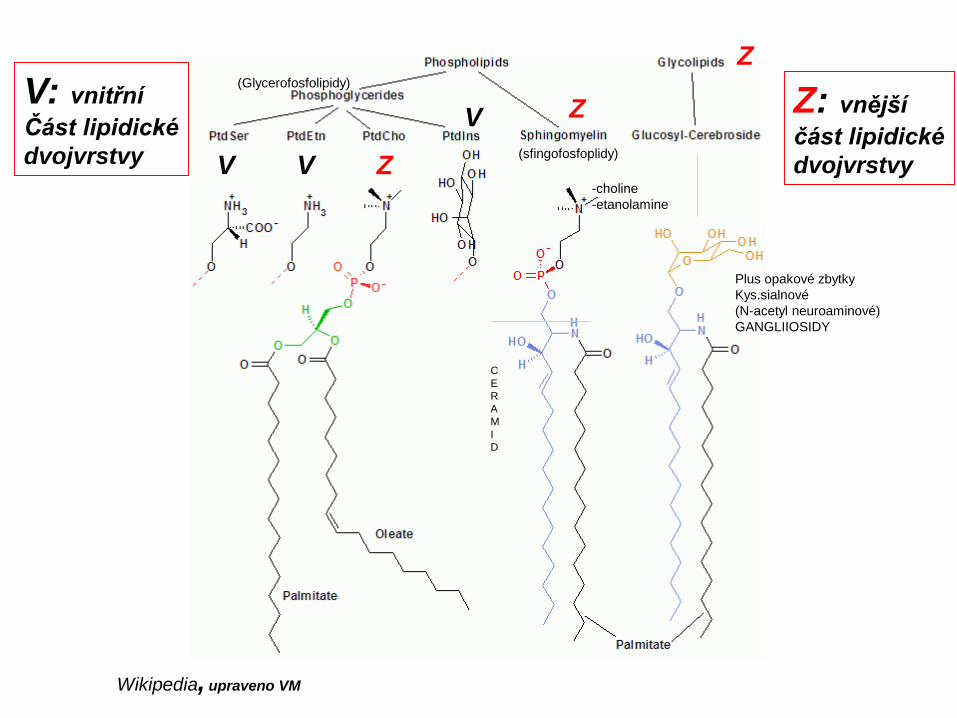
Lipidy biomembrán
Wikipedia, upraveno VM
Z
Z
Z
V V
VV: vnitřníČást lipidickédvojvrstvy
Z: vnějšíčást lipidickédvojvrstvy
-choline-etanolamine
(sfingofosfoplidy)
(Glycerofosfolipidy)
Plus opakové zbytky Kys.sialnové (N-acetyl neuroaminové)GANGLIIOSIDY
CERAMI D
§
§
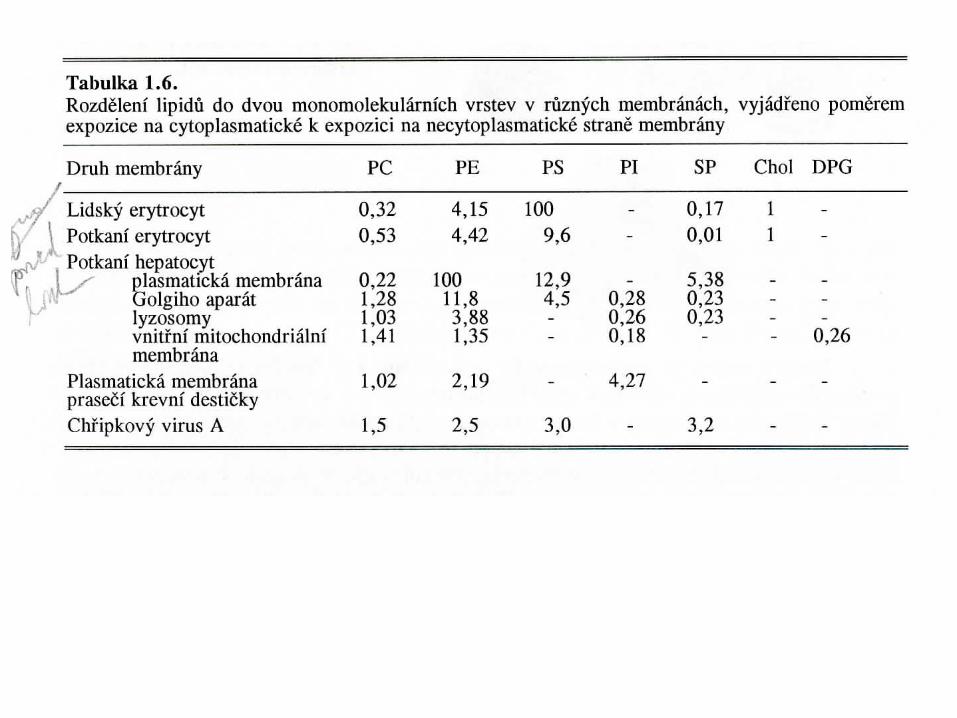
Fenotypické rozdíly ve výskytu membránových lipidů
Fenotypické rozdíly v poločasech membránových lipidů
Bílkoviny PMIntegrální 1 a 2 typu – dle lokalizace N a C konce peptidůPeriferní (asociované) – zakotvené do lipidické fáze- viz nížeIntralipidické – zanořené - rhomboidní
Rozdíly v distribuci a motilitě bílkovinEmbedding do lipidů- pohyblivost Fixace cytoskeletem a EXCM
Specifické struktury a molekuly PM dané buněčnou fenotypií Mezibunečné spojeFilipodia, lamelipodiaMicrovilliCilie, bičíky
Patofyziologie mikroheterogenity PM, vč. apoptózy
Formation of "Microdomains"Sphingolipids and cholesterol work together to help cluster proteins in a region called a "microdomain". They function as "rafts" or platforms for the attachment of proteins as membranes are moved around the cell and also during signal transduction.For more information about the rafts and "microdomains" see: Simons and Ikonen, Nature 387: 569, 1997. Also, R.E. Brown, J. Cell Science 111: 1-9, 1998
Top Figure modified from Alberts et al. Molecular Biology of the Cell, Garland Publishing, N.Y., 1994, Third Edition, Figure 10-11
(1)Standartní lipidická dvojvrstva(2)Raft. (3) lipid raft is a cholesterol and sphingolipid-enriched microdomain or platform found in ell
A lipid raft is a cholesterol and sphingolipid-enriched microdomain in cell membranes.[1] ------serving as organizing centers for the assembly of signaling molecules, influencing membrane fluidity and membrane protein trafficking, and regulating neurotransmission and receptor trafficking.[1][2] Lipid rafts are more ordered and tightly packed than the surrounding bilayer, but float freely in the membrane bilayer.[3] Wikipedia
In eukaryotes, cytoplasmic dynein must be activated by binding of dynactin, another multisubunit protein that is essential for mitosis. Dynactin may regulate the activity of dynein, and possibly facilitates the attachment of dynein to its cargo.
Each cilium (and flagellum) grows out from, and remains attached to, a basal bodyembedded in the cytoplasm. Basal bodies are identical to centrioles and are, in fact, produced by them. For example, one of the centrioles in developing sperm cells — after it has completed its role in the distribution of chromosomes during meiosis — becomes a basal body and produces the flagellum.
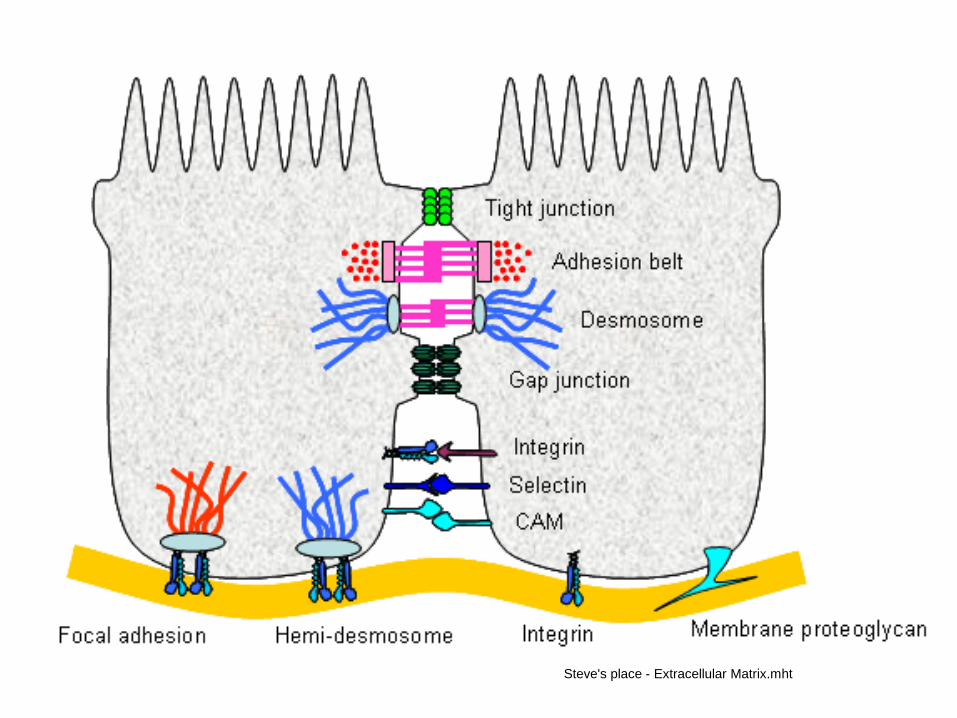
Cell-cell and cell-matrix adhesion involves a vast array of different systems
Tight junctions (formed by claudins) form impermeable barriers. Adhesion belts (adherens junctions) and desmosomes (formed by cadherin proteins) serve to link the actin and intermediate filament components of the cytoskeleton respectively.Gap junctions formed by connexins allow ion exchange between cells. CAMs (immunoglobulin-like receptors), selectins and integrins form non-communicating junctions. Integrins also link the cell to the basal lamina, either alone, or by interaction with actin (focal adhesions), or the intermediate filaments (hemidesmosomes). Membrane proteoglycans also bind the cell to the ECM. Integrins are mostly found in focal adhesions and hemidesmosomes. The binding of integrins to the ECM promotes activity of FAK (focal adhesion kinase) and the Src tyrosine kinase. This promotes cell survival by altering gene expression. The BL can self-assemble, but this is faster when it is bound to integrins
„Kotvící“ –“zpevňující“(desmomy bodové a pásové syn.syn. macula adherens a zonula adherens-adhesion belt) fokální spoje zpevňující a informační
Vodivé -pohyb látek a informací mezi buňkami (nexus syn. gap junction, elektrické synapse, chemické
synapse)
Adhezní a SignalizačníFunkce integrinů
Cadheriny
Homodimery, Ca++ dependentní membránové proteiny
Klasické: P-kadherin (placentální) VE …(endotheliálníE-kadherin(epiteliální) N…(neuronální, svalový, pojivový)
Neklasické: Desmocollin, demoglein* aj. ( *blisters!!)
Sigma/Learning Center
E-cadherin is a type 1 membrane protein. It is a member of the large family of cadherins, calcium dependent cell adhesion proteins. These proteins are involved in many morphoregulatory processes including the establishment of tissue boundaries, tissue rearrangement, cell differentiation, and metastisis. Cadherins typically consist of a large extracellular domain containing DXD and DXNDN repeats responsible for calcium-dependent adhesion, a single pass transmembrane domain, and a highly conserved, short C-terminal cytoplasmic domain responsible for interacting with catenins. E-cadherins contain five extracellular calcium-binding domains, each of ~110 amino acids. The extracellular domain of E-cadherin tends to bind in a homophilic manner; however, heterophilic binding occurs under certain conditions. The binding of extracellular cadherin is the basis for cell-cell adhesion and tends to be prevalent at adherins junctions and is structurally associated with actin bundles. The disassembly of adherens junctions is dependent on the internalization of E-cadherin via vesicle transport into the cytoplasm. The N-cadherin/ Fc chimera has been shown to retain structural and functional properties of the cadherins. Sigma/Learning Center
Integrins are adhesive membranereceptors. All integrins are heterodimeric transmembrane proteins. They differ from hormone receptors in that they have lower affinity for their ligands, and are present in higher numbers. They exhibit a Velcro effect: strength in numbers, but individually easy to disrupt. They require Ca2+ or Mg2+ to bind, and their job is to link the ECM to the cytoskeleton.
Steve's place - Extracellular Matrix.mht
Integrins are heterodimers. Most cell-matrix adhesion involves the β1 subunit, and specificity is achieved by different α subunits.
There are 24 α types and 9 β types - β1 binds ECM proteins which combine to form many different integrins with varied roles and cell specificities:α5β1 binds fibronectin. α6β1 binds laminin. α7β1 binds laminin in muscle.
Steve's place - Extracellular Matrix.mht
Fibronectin
LamininEXCM
Integrins can also perform inside out signalling: the β subunit binds to cytoplasmic talin, and thence to actin, which is how integrins link the ECM to the cytoskeleton. Phosphorylation of the integrin cytoplasmic tails prevents binding to talin, and dissociates integrin from both cytoskeleton and ECM. Anchoring to the ECM or basal lamina is critical for cell survival. Cells that fail to anchor often apoptose. For example, during embryogenesis, ectoderm cells are induced to apoptose by chemical signals from endoderm cells in the formation of a proamniotic cavity. Only those endoderm cells attached to the basal lamina survive. Steve's place - Extracellular Matrix.mht
Fibronectin aj. EXCM
P
P
Steve's place - Extracellular Matrix.mht
Top Figure modified from Alberts et al. Molecular Biology of the Cell, Garland Publishing, N.Y., 1994, Third Edition, Figure 10-11
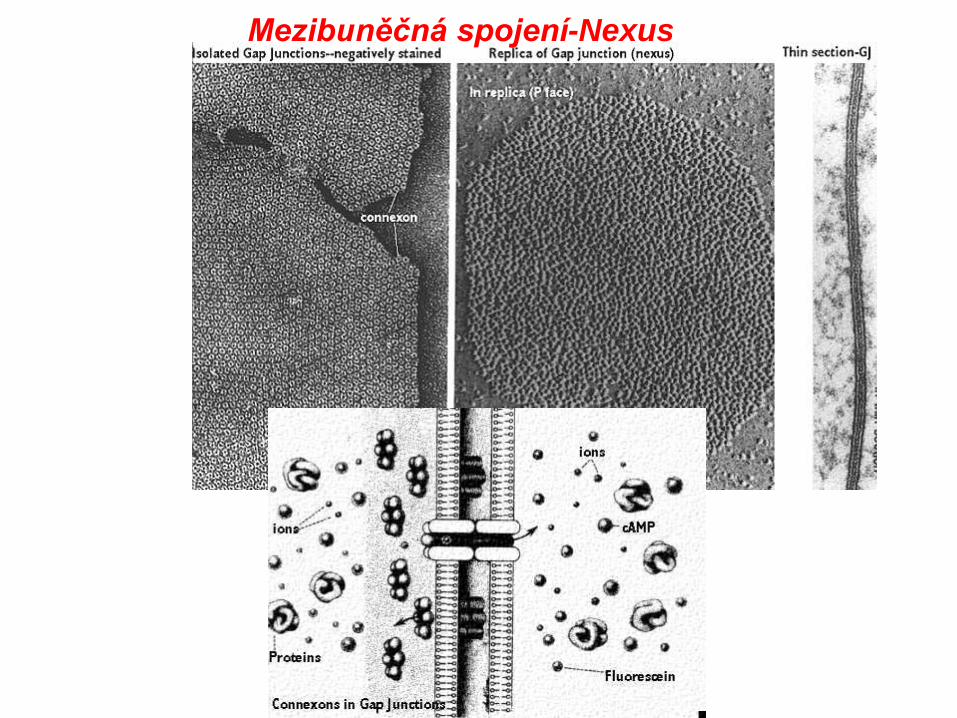
Nexus, syn. dírkatý spoj, vodivý nebo mezerový spoj
Top Figure modified from Alberts et al. Molecular Biology of the Cell, Garland Publishing, N.Y., 1994, Third Edition, Figure 10-11
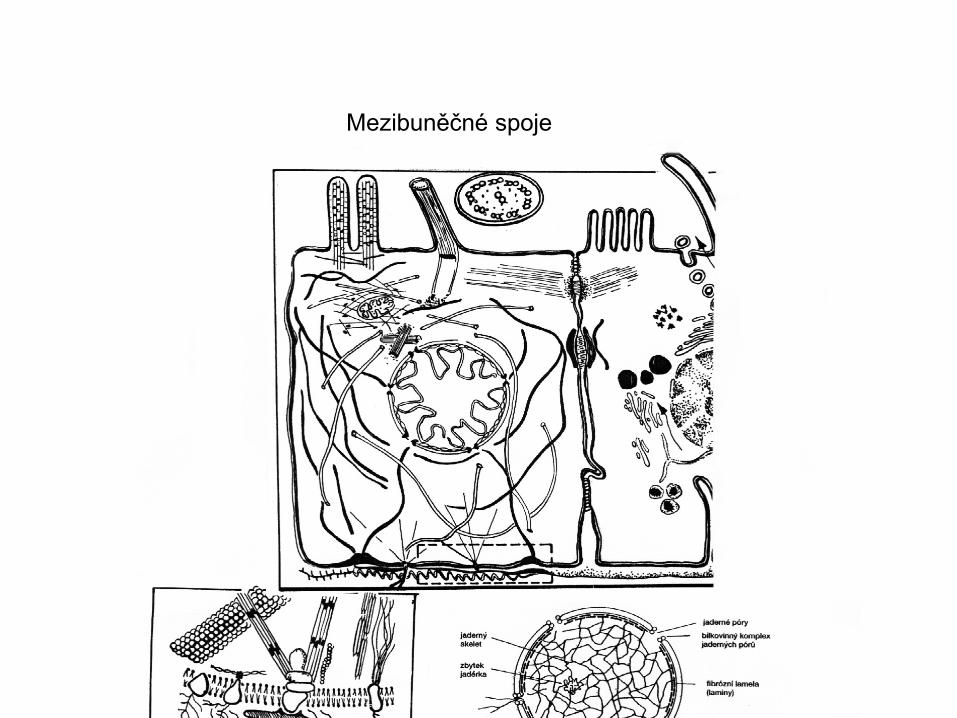
Mezibuněčné spoje
Mezibuněčná spojení
Gannong 2005
Těsnící
Kotvící-zpevňující
Komunikační
*
*
Integriny HeterodimeryReceptorová adhese aSignalizace mezi buňkami abuňkami a EXCMIntegrins integrate the ECM (here fibronectin) and the cytoskeleton (here an actin microfilament). Regulation is possible from both directions: binding of fibronectin to integrins allows the action of FAK(Focal Adhesion Kinase); but phosphorylation of the integrin causes it to dissociate from cytoskeleton-binding proteins such as talin. It seem that this also causes conformational changes in the extracellular portion of the integrin, loosening its grip on the ECM.
Anchoring to the ECM or basal lamina is critical for cell survival Cells that fail to anchor often apoptose. For example, during embryogenesis, ectoderm cells are induced to apoptose by chemical signals from endoderm cells in the formation of a proamniotic cavity. Only those endoderm cells attached to the basal lamina survive. Steve's place - Extracellular Matrix.mht
Integrins are obligate heterodimers containing two distinct chains, called the α (alpha) and β (beta) subunits. In mammals, 19 α and 8 β subunits have been characterized, whereas the Drosophila genome encodes only five α and two β subunits, and the Caenorhabditis nematodes possess two α and one β genes.[1] Both the α and β subunits contain two separate tails, both of which penetrate the plasma membrane and possess small cytoplasmic domains.[2] Wikipedia
Integrins can also perform inside out signalling: the β subunit binds to cytoplasmic talin, and thence to actin, which is how integrins link the ECM to the cytoskeleton. Phosphorylation of the integrin cytoplasmic tails prevents binding to talin, and dissociates integrin from both cytoskeleton and ECM. Steve's place - Extracellular Matrix.mht Wikipedia
Beta1 integrins interact with many alpha integrin chains. Gene knockouts of integrins in mice are not always lethal. It proves that during embryonal development, one integrin may substitute its function for another, to allow survival. Some integrins are on the cell surface in an inactive state, and can be rapidly primed, or put into a state capable of binding their ligands, by cytokines. Integrins can assume several different well defined shapes, or "conformational states". Once primed, the conformational state changes to stimulate ligand binding which then activat
Mezibuněčná spojení-Nexus
Mezibuněčná spojení-Nexus
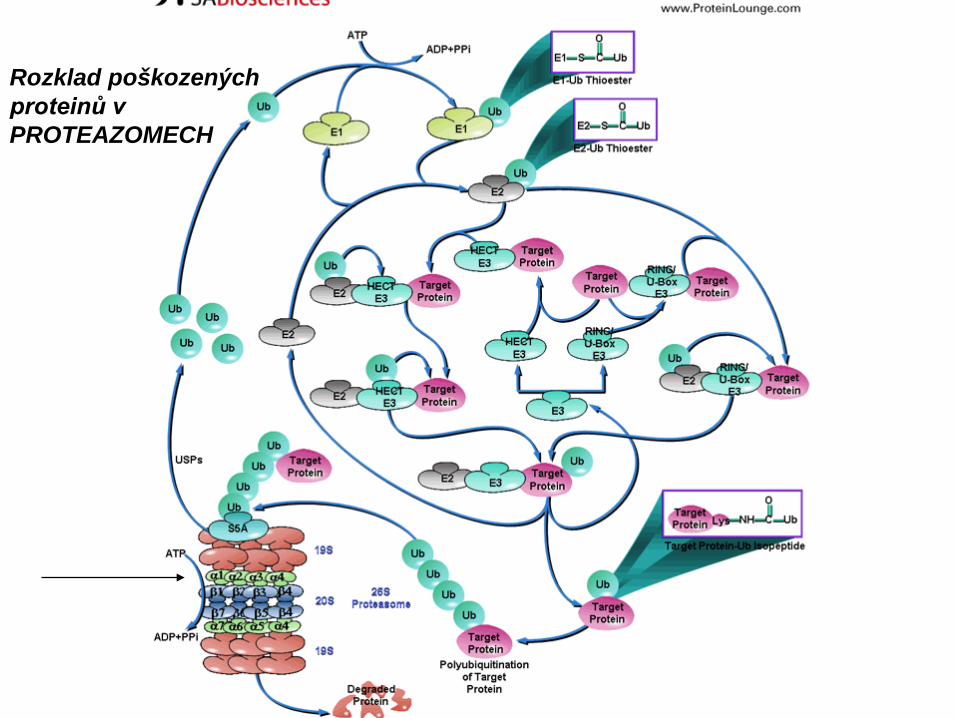
Další organely membránového systému
Síť gliových buněk s výběžky v kultuře(fázový kontrast)
Výřez viz EM na dalším snímku)
EM gliových buněk v kultuře
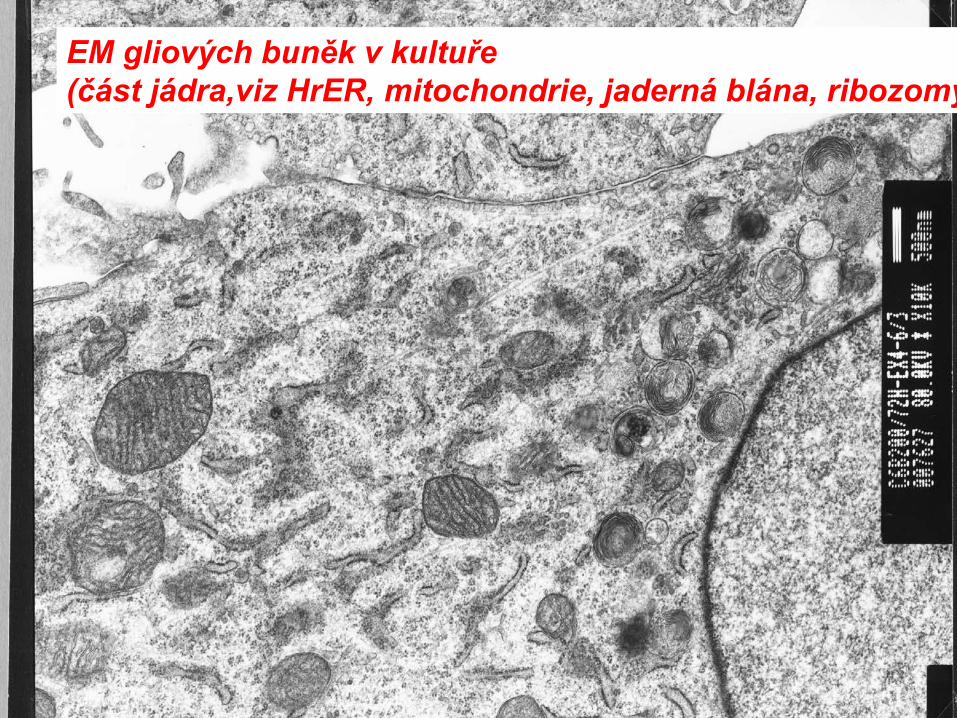
EM gliových buněk v kultuře(část jádra,viz HrER, mitochondrie, jaderná blána, ribozomy
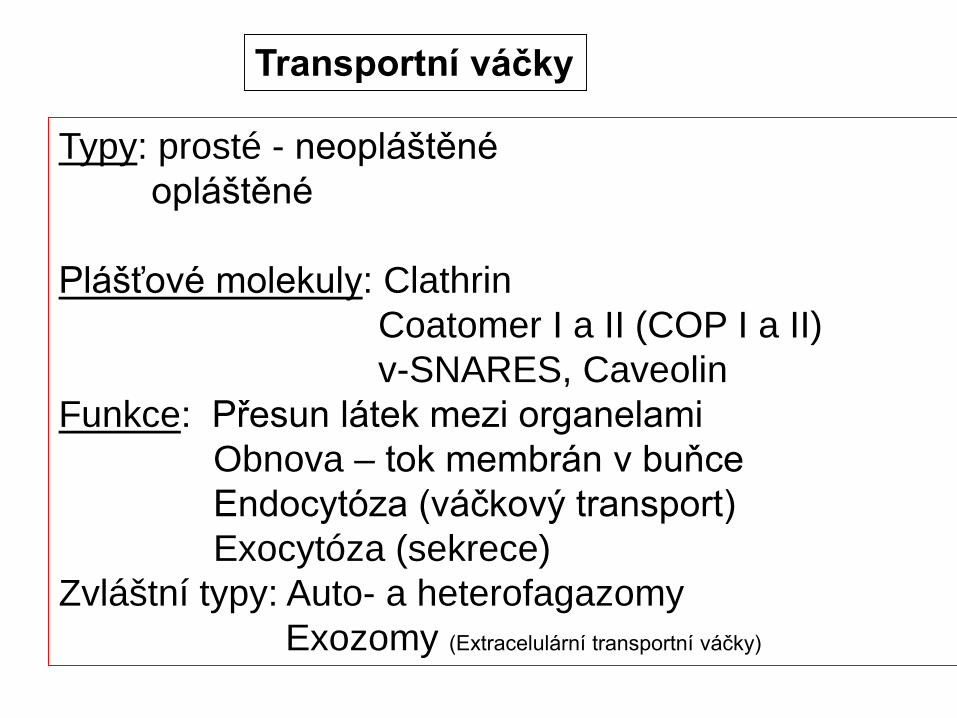
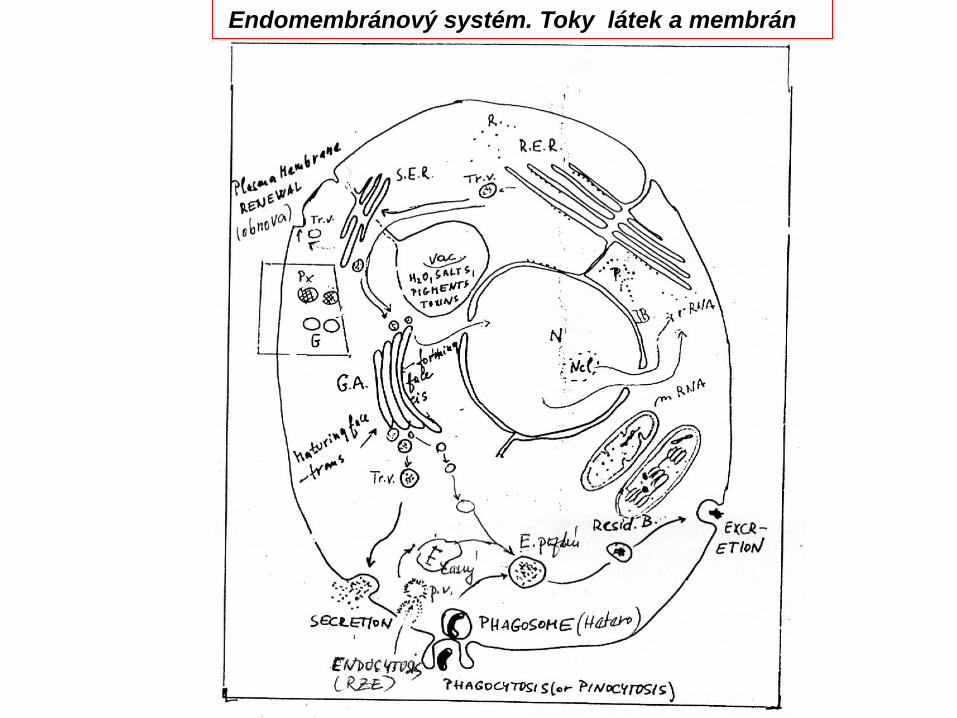
Plášťové molekuly: ClathrinCoatomer I a II (COP I a II)v-SNARES, Caveolin
Funkce: Přesun látek mezi organelamiObnova – tok membrán v buňceEndocytóza (váčkový transport)Exocytóza (sekrece)
Zvláštní typy: Auto- a heterofagazomyExozomy (Extracelulární transportní váčky)
Transportní váčkyEndo-exocytózaHrER
Exocytoza (sekrece) a endocytoza
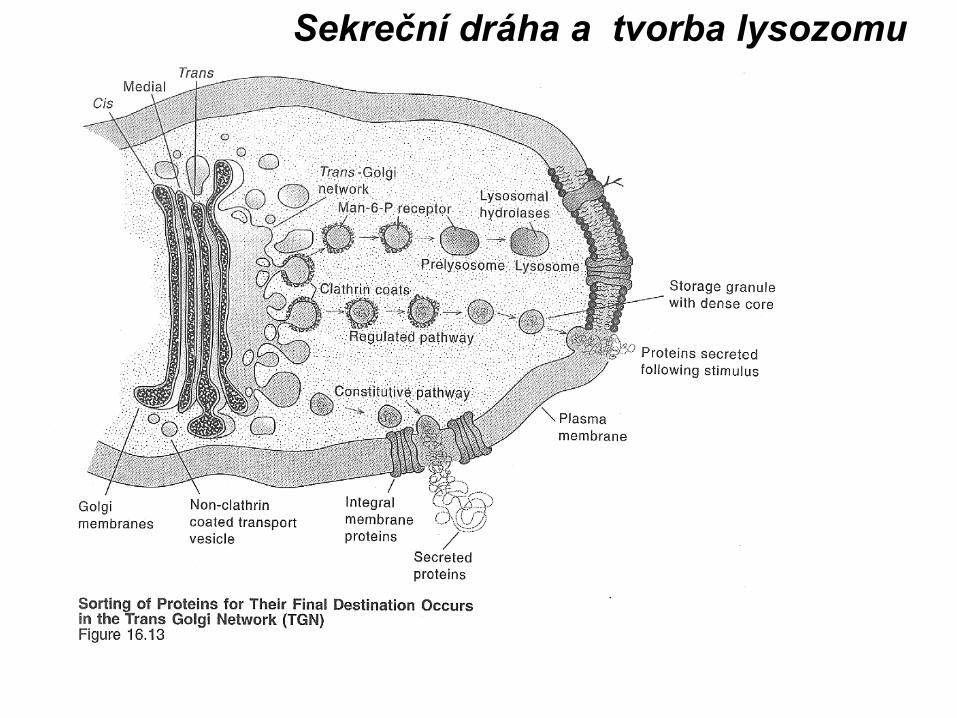
Sekreční dráhaProstáReguovaná
EndocytoProstá-pinReceptoro
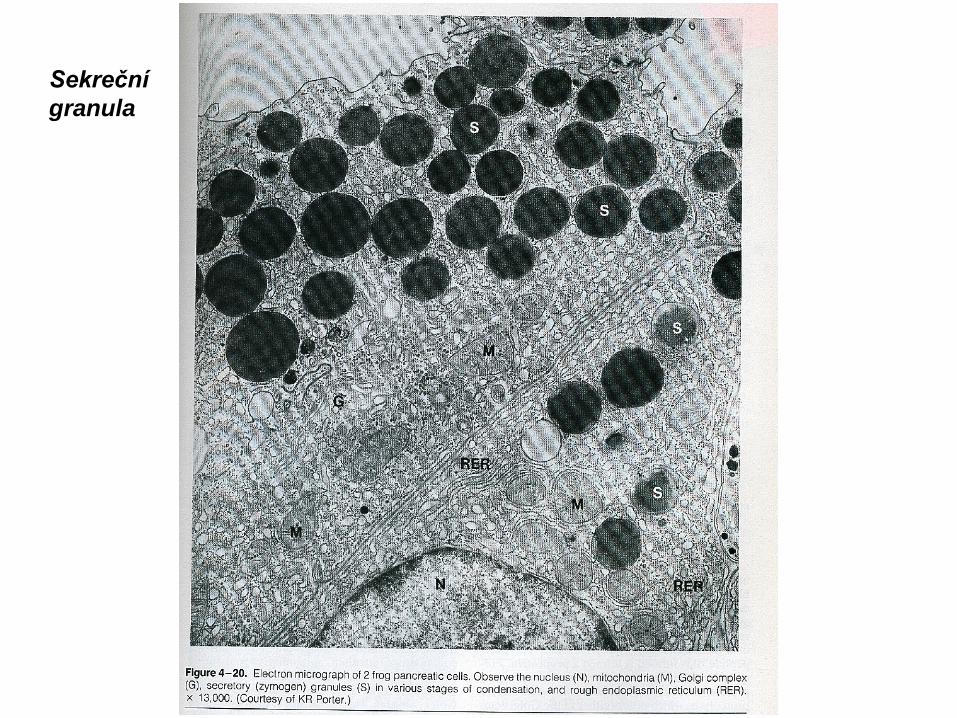
Sekreční granula
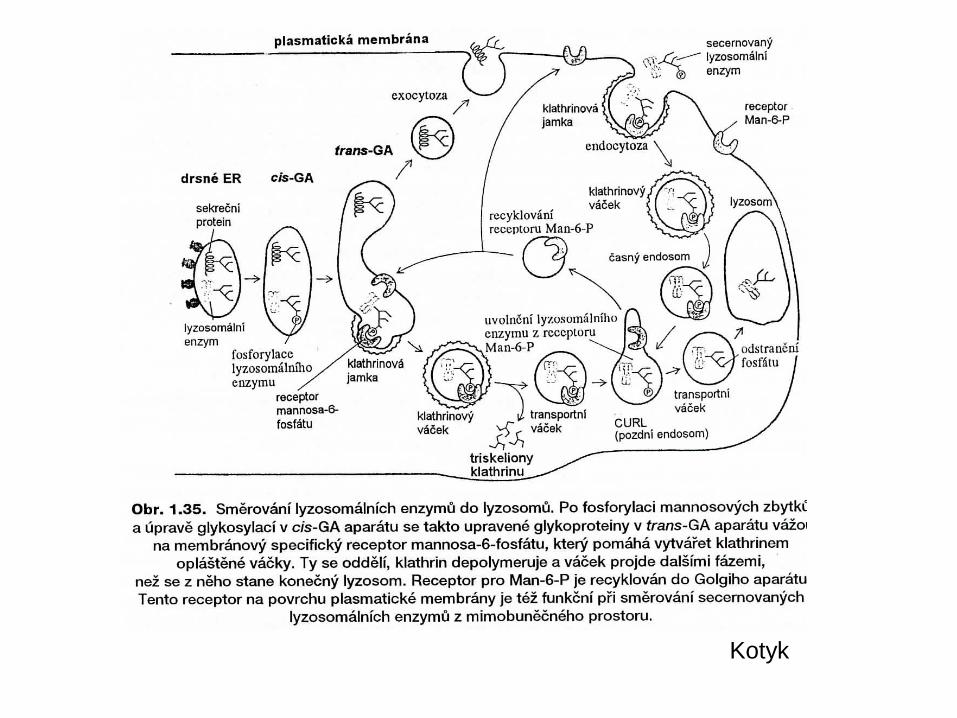
Kotyk
Endomembránový systém. Toky látek a membrán
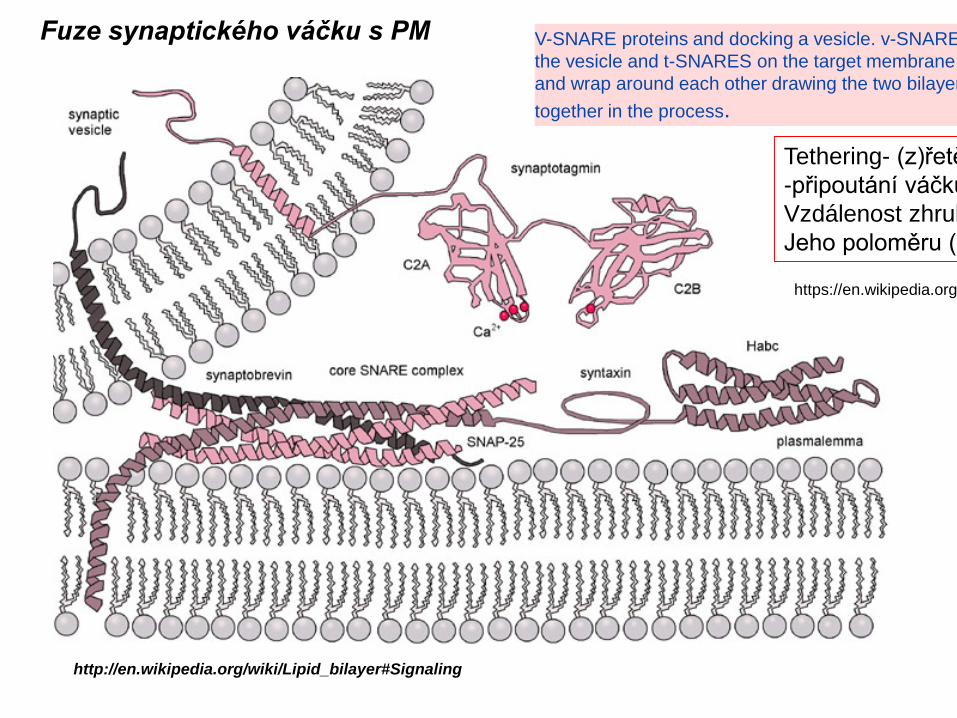
Vesicle trafficking[edit]Certain vesicle-trafficking steps require the transportation of a vesicle over a moderately small distance. For example, vesicles that transport proteins from the Golgi apparatus to the cell surface area, will be likely to use motor proteins and a cytoskeletal track to get closer to their target. Before tethering would have been appropriate, many of theproteins used for the active transport would have been instead set for passive transport, because the Golgi apparatus does not require ATP to transport proteins. Both the actin- and the microtubule-base are implicated in these processes, along with several motor proteins. Once the vesicles reach their targets, they come into contact with tetheringfactors that can restrain them.Vesicle tethering[edit]It is useful to distinguish between the initial, loose tethering of vesicles to their objective from the more stable, packing interactions. Tethering involves links over distances of more than about half the diameter of a vesicle froma given membrane surface (>25 nm). Tethering interactions are likely to be involved in concentrating synaptic vesicles at the synapse.Tethered vesicles are also involved in regular cell's transcription processes.Vesicle docking[edit]Secretory vesicles transiently dock at the cell plasma membrane, preceding the formation of a tight t-/v-SNARE complex, leading to priming and the establishment of continuity between the opposing bilayers.Vesicle priming[edit]In neuronal exocytosis, the term priming has been used to include all of the molecular rearrangements and ATP-dependent protein and lipid modifications that take place
V-SNARE proteins and docking a vesicle. v-SNARE the vesicle and t-SNARES on the target membrane and wrap around each other drawing the two bilayer together in the process.
Fuze synaptického váčku s PM
Tethering- (z)řetě-připoutání váčku Vzdálenost zhrubJeho poloměru (-
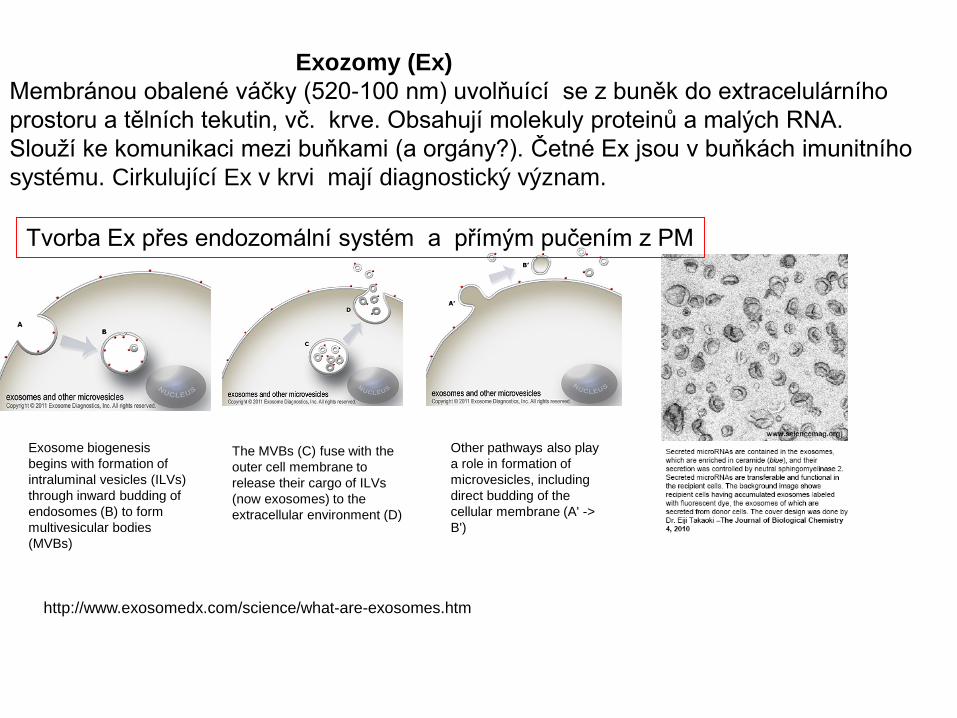
Exozomy (Ex)Membránou obalené váčky (520-100 nm) uvolňuící se z buněk do extracelulárního prostoru a tělních tekutin, vč. krve. Obsahují molekuly proteinů a malých RNA. Slouží ke komunikaci mezi buňkami (a orgány?). Četné Ex jsou v buňkách imunitního systému. Cirkulující Ex v krvi mají diagnostický význam.
Exosome biogenesis begins with formation of intraluminal vesicles (ILVs) through inward budding of endosomes (B) to form multivesicular bodies (MVBs)
The MVBs (C) fuse with the outer cell membrane to release their cargo of ILVs (now exosomes) to the extracellular environment (D)
Other pathways also play a role in formation of microvesicles, including direct budding of the cellular membrane (A' -> B')
…..a hydrogenosomyHydrogenosomy jsou, podobně jako mitochondrie, malé organely rovněž obalené dvojitou membránou. Vnitřní membrána však netvoří kristy ani tubuly pravděpodobně ani DNA. Vyskytuje se u jednobuněčných anerobních prvoků, žijících většinou parasiticky (Trichomonas vaginalis, bičíkovci žijící v bachoru přežvýkavců, některé nižší houby). Mají schopnost oxidovat některé organické molekuly, především pyruvát, který přeměňuji na acetát a plynný molekulární vodík (H2). Oxidací uvolněnou energii přeměňují na ATP, nikoliv však cestou chemiosmotické, ale tzv. substrátové fosforylace. V hydrogenozómech chybí některé enzymy mitochondriálního dýchacího řetězce a ATP syntáza.
Kolem původu této málo časté organely je mnoho spekulací. Dle některých autorů hydrogenozomy představují anerobního prokaryontního prapředka asimilovaného v aerobní prabuňce. Dle jiných, hydrogenozómy jsou zjednodušenou formou mitochondrií, adaptovaných na anerobní podmínky a parasitický způsob života nositelů těchto organel. Nevylučuje se ani jejich vývojový vztah k peroxisomům. Pozoruhodné je rovněž to, že hydrogenozómy byly objeveny v r. 1973 dvěma pracovníky v zahraničí a současně i českým přírovědcem Jiřím Čerkasovem.
A hydrogenosome is a membrane-enclosed organelle of some anaerobic ciliates, trichomonads and fungi. The hydrogenosomes of trichomonads (the most studied of the hydrogenosome-containing microorganisms) produce molecular hydrogen, acetate, carbon
dioxide and ATP by the combined actions of pyruvate:ferredoxin oxido-reductase, hydrogenase, acetate:succinate CoA transferase and succinate thiokinase. Superoxide dismutase, malate dehydrogenase (decarboxylating), ferredoxin, adenylate kinase and
NADH:ferredoxin oxido-reductase are also localized in the hydrogenosome. This organelle is thought to have evolved from anaerobic bacteria or archaea, though in the case of trichomonad hydrogenosomes the question remains open.
In 2010, scientists reported their discovery of the first known anaerobic metazoans with hydrogenosome-like organelles.[2]
CytoskeletKomplex vláknitých struktur několika základních typů, které zajišťují tvar a oporu buňky, pružnost a pohyb jejich organel v buňce i celé buňky, podílí se na příjmu í informací z okolní mezibuněčné hmoty a jejich odpovědí v buňce
Cytoskelet dle lokalizace: CytoplazmatickýMembránovýJaderný
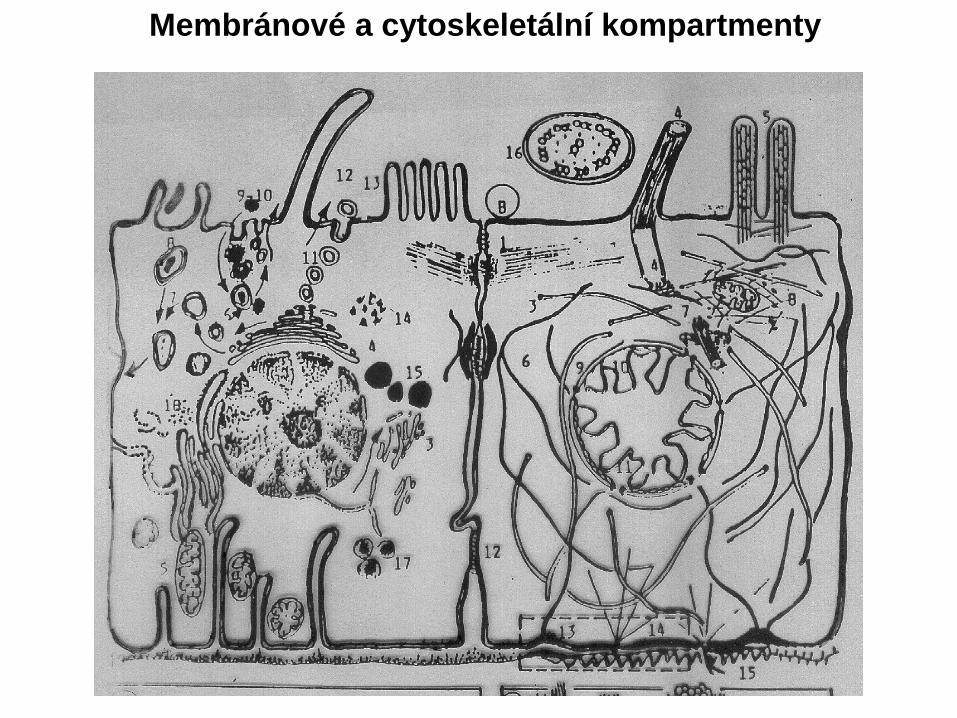
Membránové a cytoskeletální kompartmenty
Cytoskelet a jeho složky a distribuce v buňce
Alberts 2000
Alberts 2000
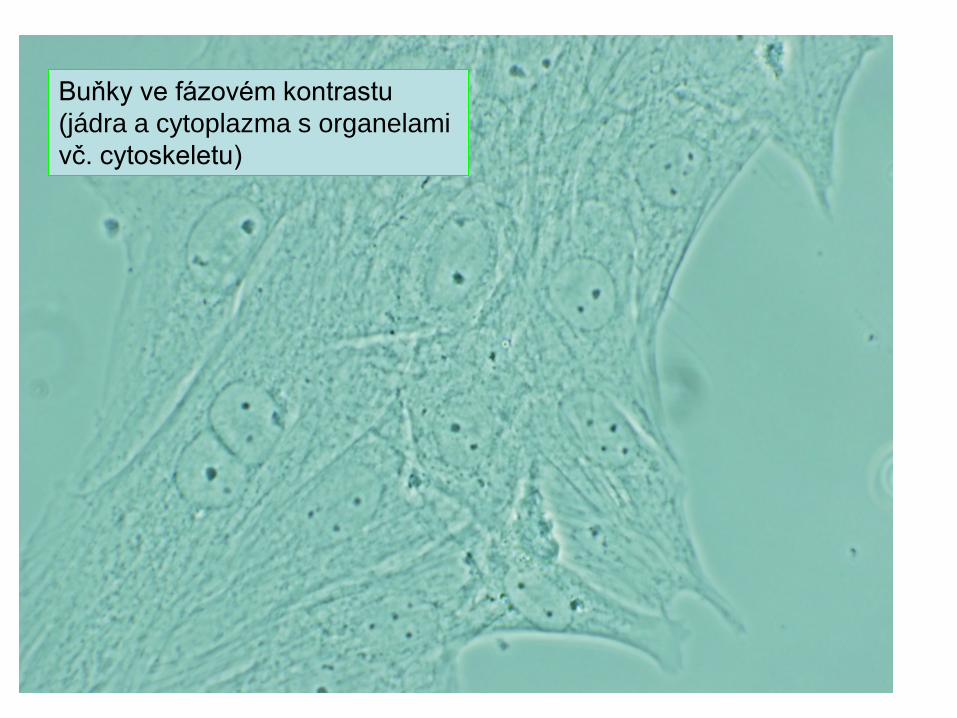
Buňky ve fázovém kontrastu (jádra a cytoplazma s organelami vč. cytoskeletu)
Mikrofilamenta -protilátka proti aktinu, fluorescenční technika
Endothelial cells. Nuclei are stained blue with DAPI, microtubles are marked green by an antibody bound to FITC and actin filaments are labelled red with phalloidin bound to TRITC.
Stavba a funkce mikrotubulů (MT)(cytoplazmatické, bičíkové, centriolové)
Stavba bičíku a cilií
Ganong 2005
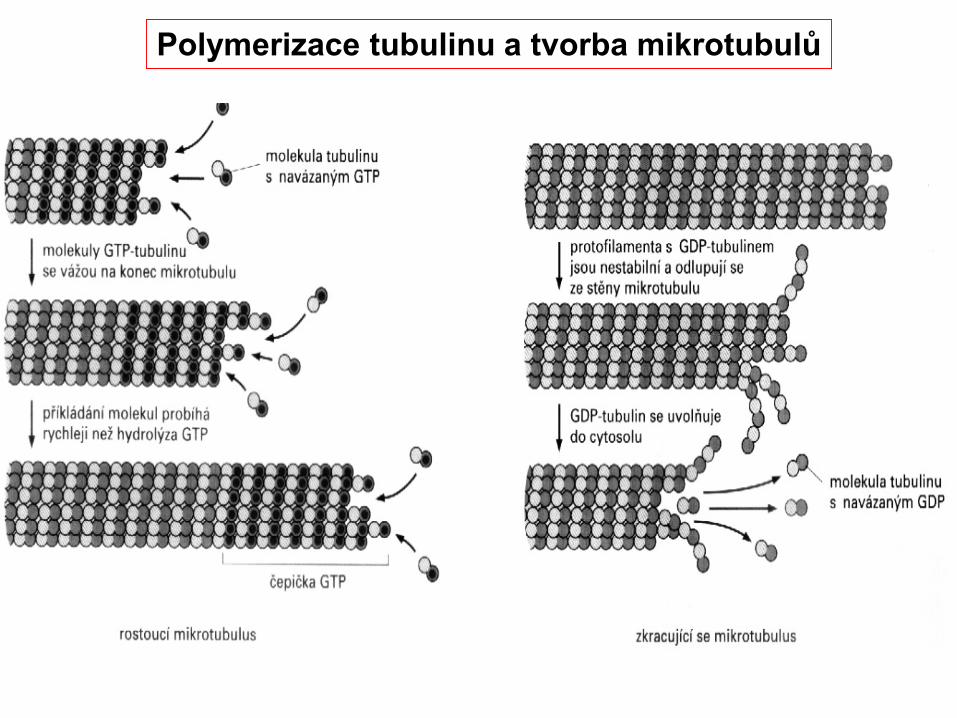
Polymerizace tubulinu a tvorba mikrotubulů
Organizace polymerace mikrotubulů centriolou
+
-
GTP
GDP
GTP čepička stabilizuje MTGDP podporuje depolymeraci
Střední filamenta a mikrofilamenta
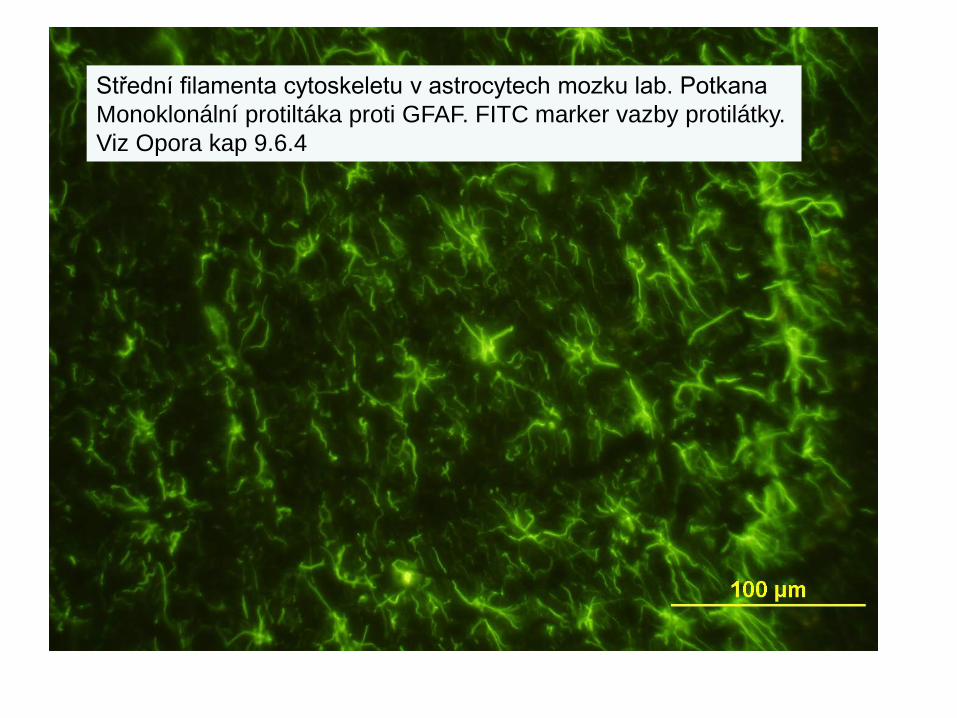
Střední filamenta cytoskeletu
Střední filamenta cytoskeletu v astrocytech mozku lab. PotkanaMonoklonální protiltáka proti GFAF. FITC marker vazby protilátky.Viz Opora kap 9.6.4
Střední filamenta cytoskeletu v astrocytech mozku lab.potkanaMonoklonální protiltáka proti GFAF. Marker vazby protilátky diaminobenzidinová reakce s křenovou peroxidázou. Viz Opora kap 9.6.4.
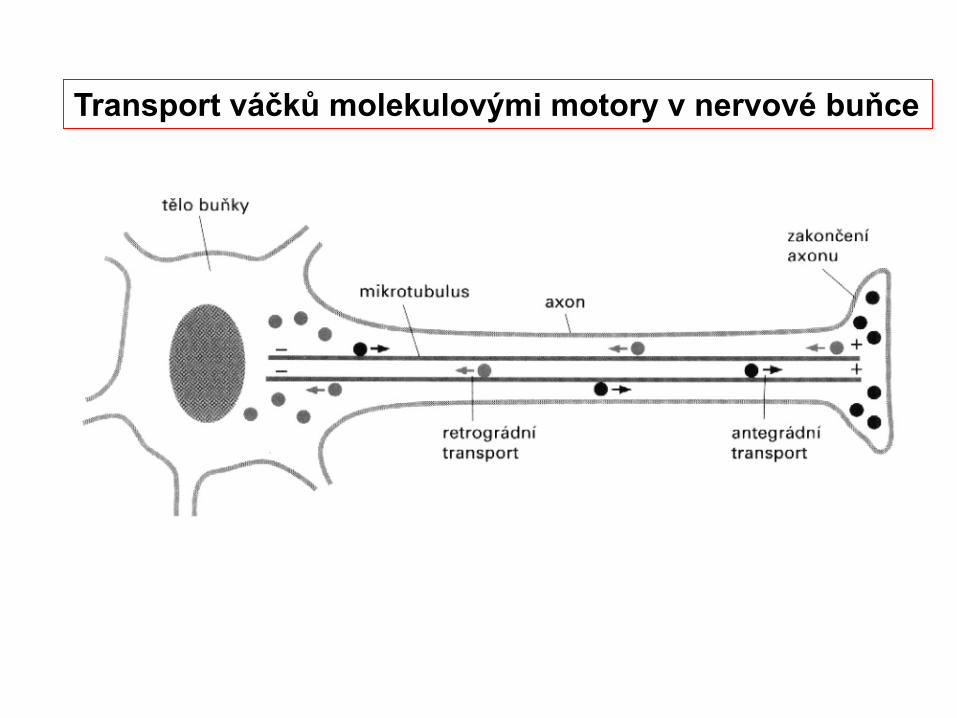
Transport váčků molekulovými motory v nervové buňce
Molekulové motory asociované s mikrotubuly
Molekulové motory asociované s mikrotubuly
Molekulové motory asociované s aktinovými mikrofilamenty
Ganong 2005
Membránový (kortikální) cytoskelet
Membránovýcytoskelet
Ganong 2005
A fourth eukaryotic cytoskeletal element, microtrabeculae, was proposed by Keith Porter based on images obtained from high-voltage electron microscopy of whole cells in the 1970s.[13]The images showed short, filamentous structures of unknown molecular composition associated with known cytoplasmic structures. Porter proposed that this microtrabecular structure represented a novel filamentous network distinct from microtubules, filamentous actin, or intermediate filaments. It is now generally accepted that microtrabeculae are nothing more than an artifact of certain types of fixation treatment, although we have yet to fully understand the complexity of the cell's cytoskeleton.[14] http://en.wikipedia.org/wiki/Cytoskeleton
The prokaryotic cytoskeleton http://en.wikipedia.org/wiki/CytoskeletonMain article: prokaryotic cytoskeletonThe cytoskeleton was previously thought to be a feature only of eukaryotic cells, but homologues to all the major proteins of the eukaryotic cytoskeleton have recently been found in prokaryotes.[9] …..[10]….[11]
FtsZ was the first protein of the prokaryotic cytoskeleton to be identified. Like tubulin, FtsZ forms filaments in the presence of GTP, but these filaments do not group into tubules. During cell division, FtsZ ….. to move to the division site, and is essential for recruiting other proteins for synthesis the new cell wall between the dividing cells.
Prokaryotic actin-like proteins, such as MreB, are involved in the maintenance of cell shape….form a helical network beneath the cell membrane that guides the proteins involved in cell wall biosynthesis.Some plasmids encode a actin-like protein ParM. Filaments of ParM exhibit dynamic instability, and may partition plasmid DNA into the dividing daughter cells by a mechanism analogous to that used by microtubules during eukaryotic mitosis.The bacterium Caulobacter crescentus contains a third protein, crescentin, that is related to the intermediate filaments of eukaryotic cells. Crescentin is also involved in maintaining cell shape, such as helical and vibrioid forms of bacteria, but the mechanism by which it does this is currently unclear.[12]Microtrabeculae
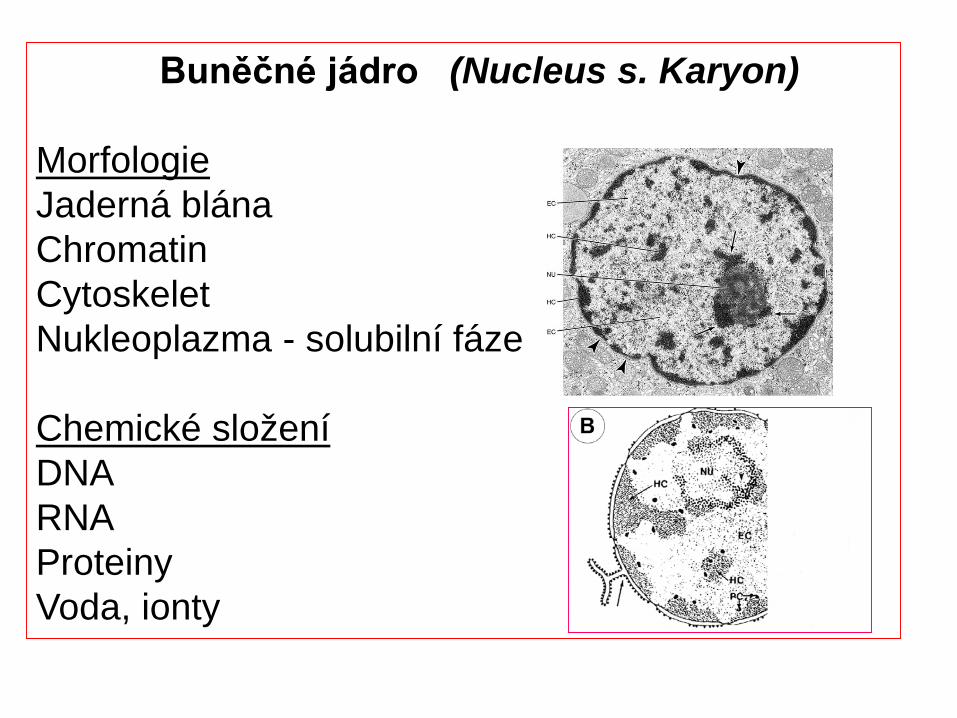
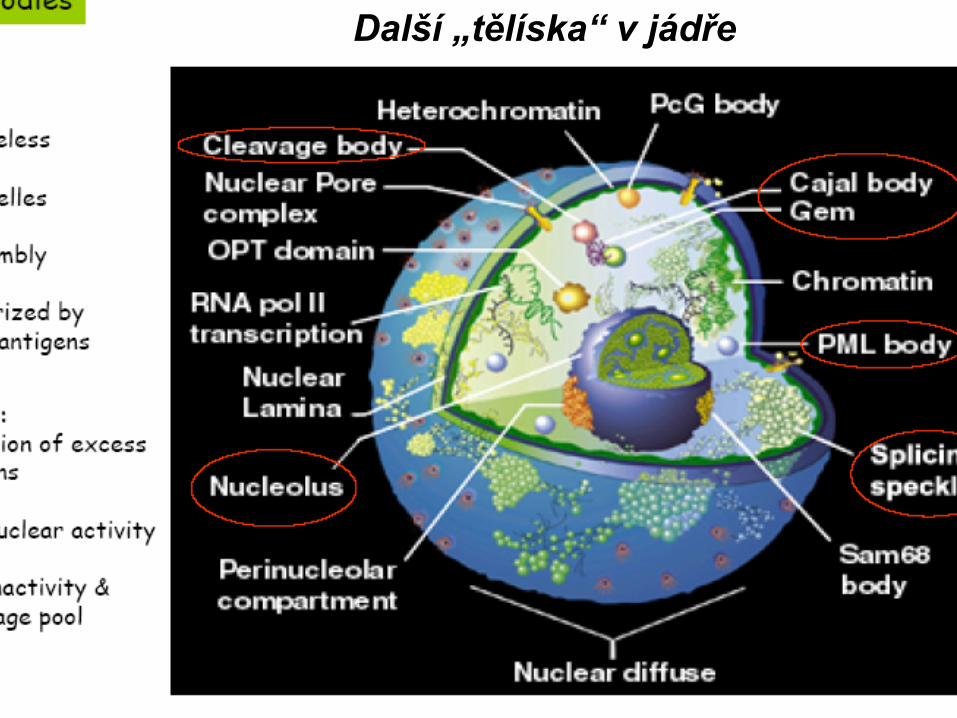
MorfologieJaderná blánaChromatinCytoskeletNukleoplazma - solubilní fáze
Chemické složeníDNARNAProteinyVoda, ionty
Romanovský a kol. 1985Viz též obrazová část Opory OZ
Krevní buňky
Author: Sherri Wick, Coordinator and Instructor - Human Physiology and Anatomy LaboratoriesUniversity of Nebraska at OmahaAllwine Hall 211E, [email protected]
Nervové, gliové a cévní buňky(přední mozek)
Neuron
Glie
Endothelie kapiláry
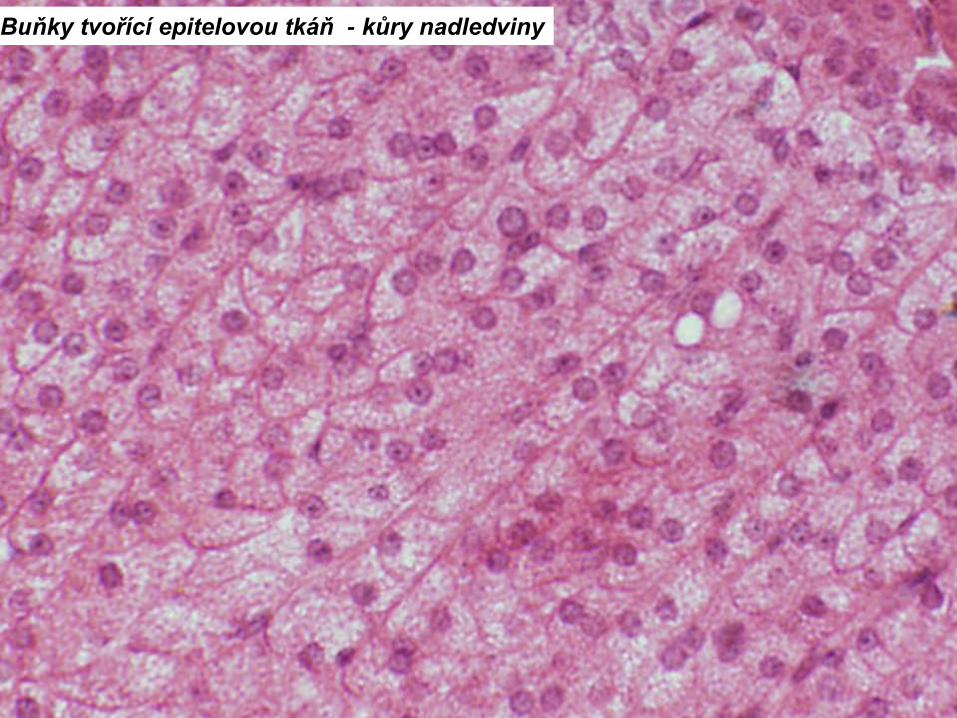
Buňky tvořící epitelovou tkáň - kůry nadledviny
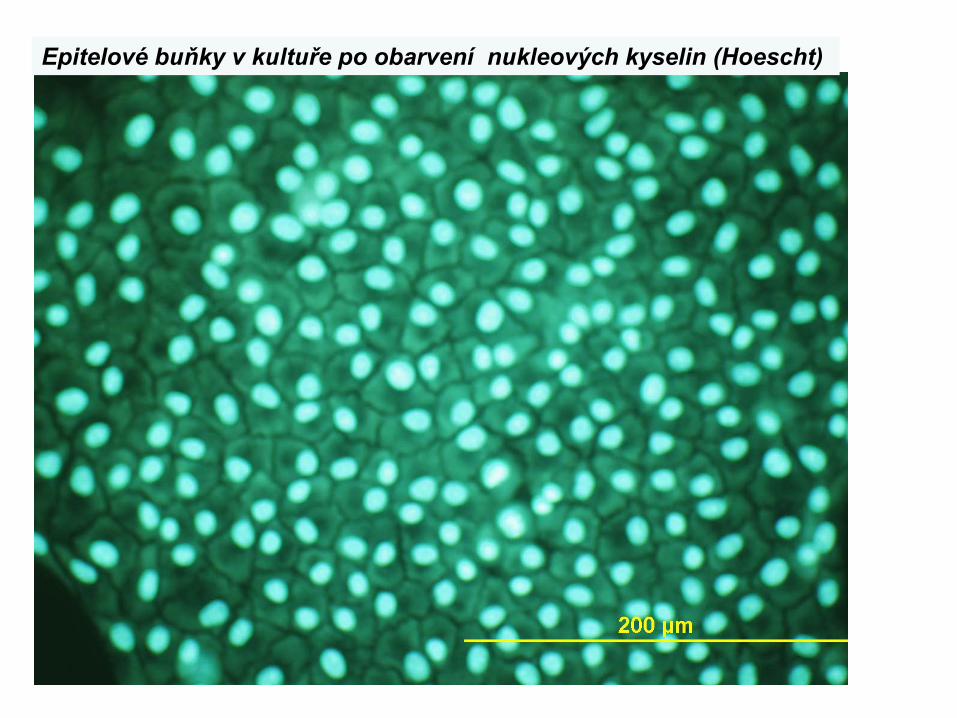
Epitelové buňky v kultuře po obarvení nukleových kyselin (Hoescht)
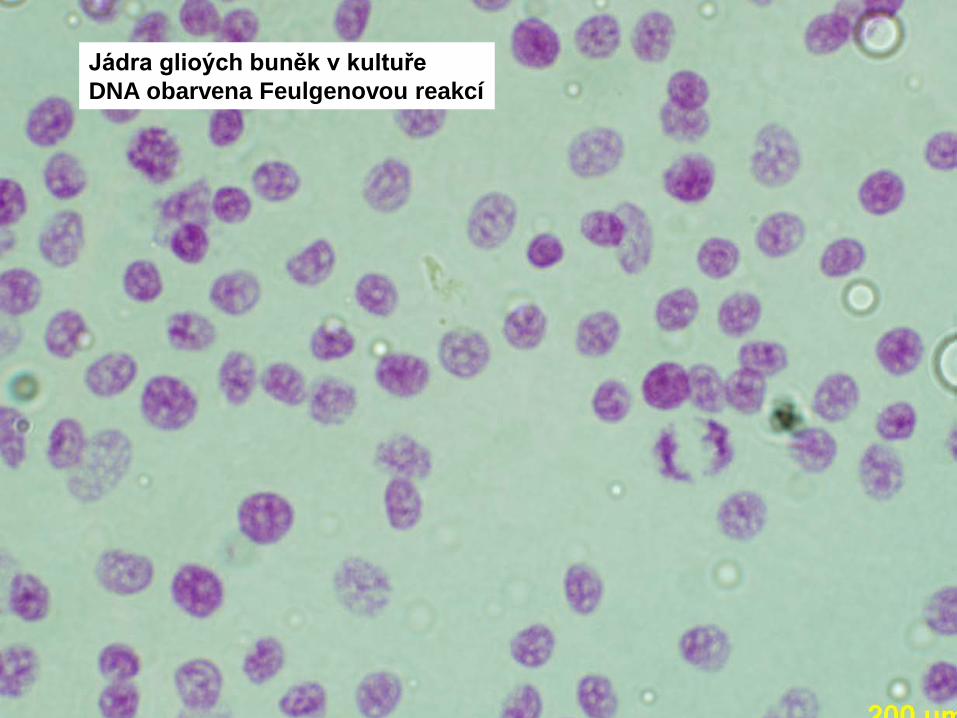
Jádra glioých buněk v kultuřeDNA obarvena Feulgenovou reakcí
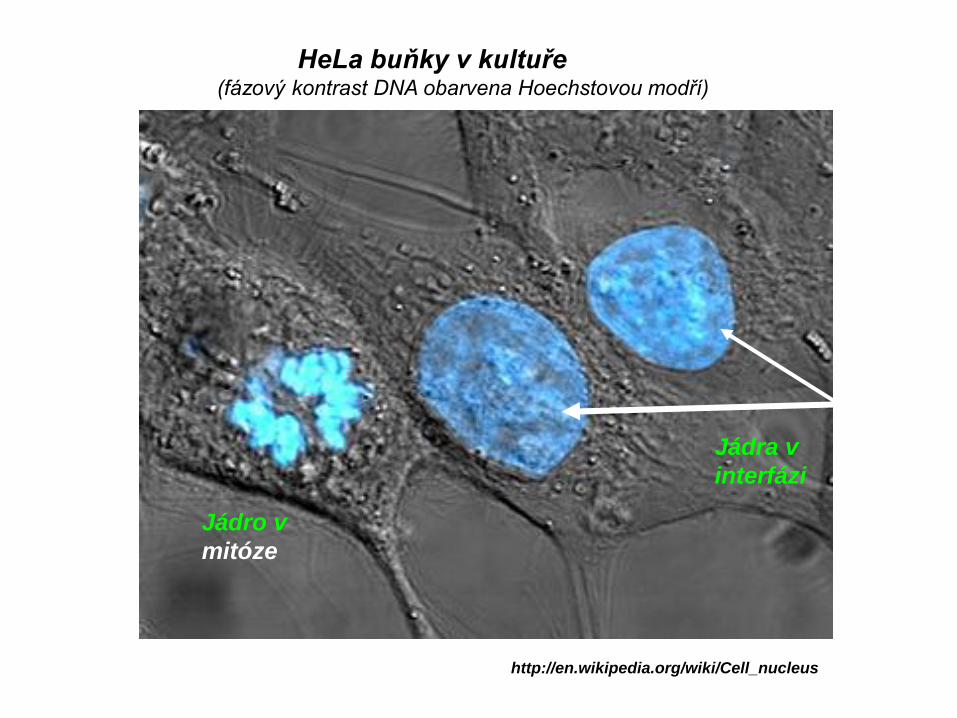
HeLa buňky v kultuře(fázový kontrast DNA obarvena Hoechstovou modří)
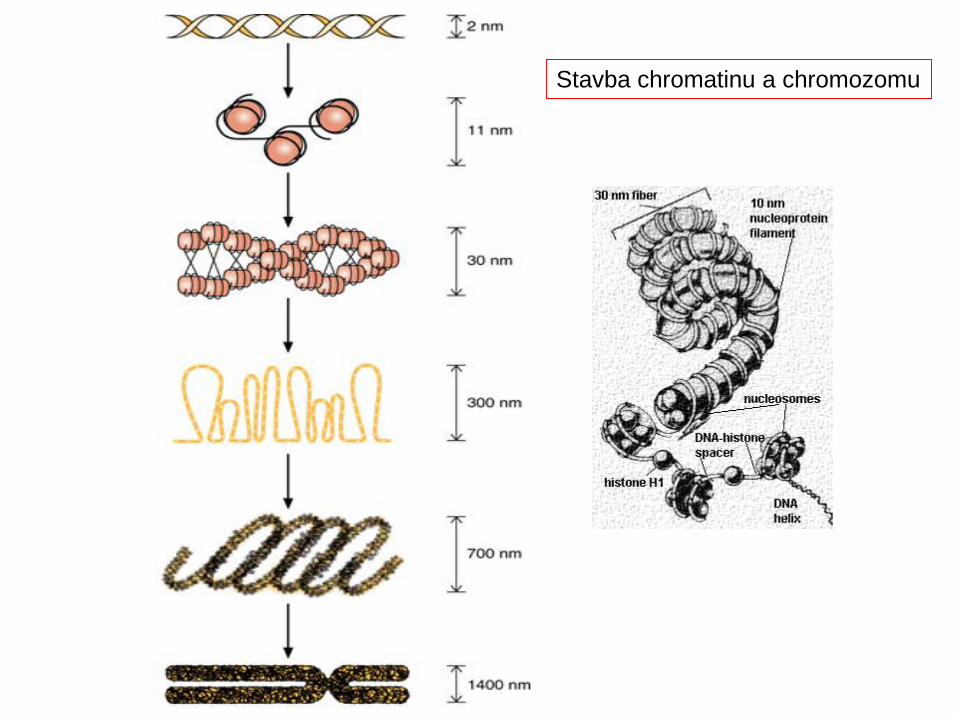
Struktura chromatinu má několik úrovní. Základní jednotkou je nukleozóm, struktura tvořená histonovými molekulami obtočemými DNA (asi 80 párů bází). Vyšší strukturou je solenoid, spiralizované uspořádání nukleozómů (1 závit tvoří asi 6 nukleozómů a nese 1200 párů bází). Solenoidy se uspořádávají do smyček, z nichž každá obsahuje okolo 50 otoček solenoidu a nese statisíce párů bází. 18 smyček pravidelně uspořádaných okolo základní proteinové matrice vytváří základní segment chromozómu
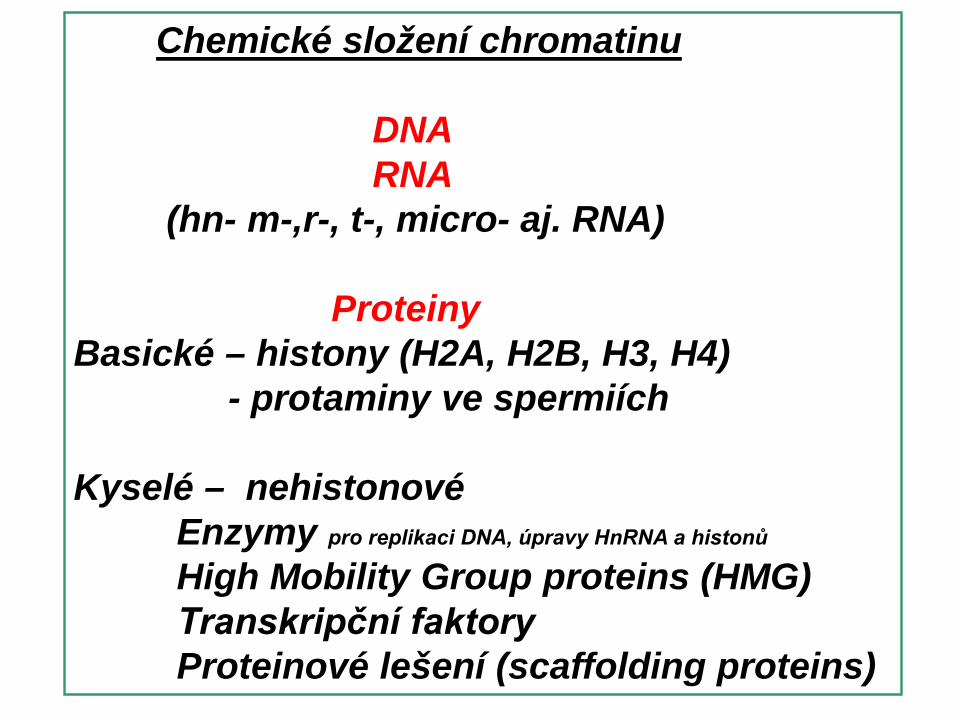
Kyselé – nehistonovéEnzymy pro replikaci DNA, úpravy HnRNA a histonů
High Mobility Group proteins (HMG)Transkripční faktoryProteinové lešení (scaffolding proteins)
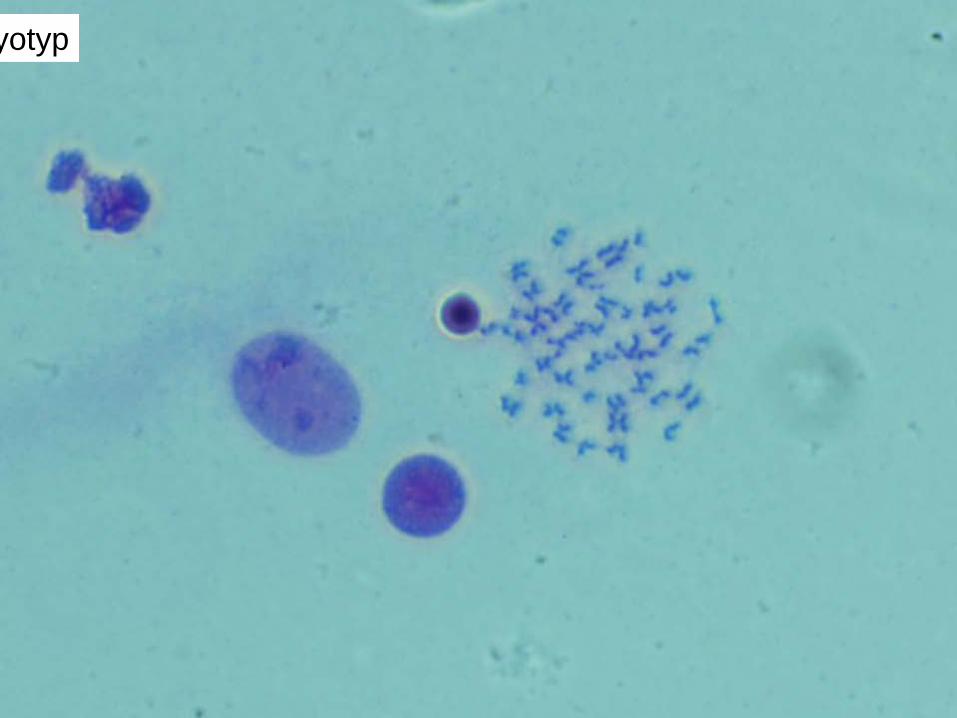
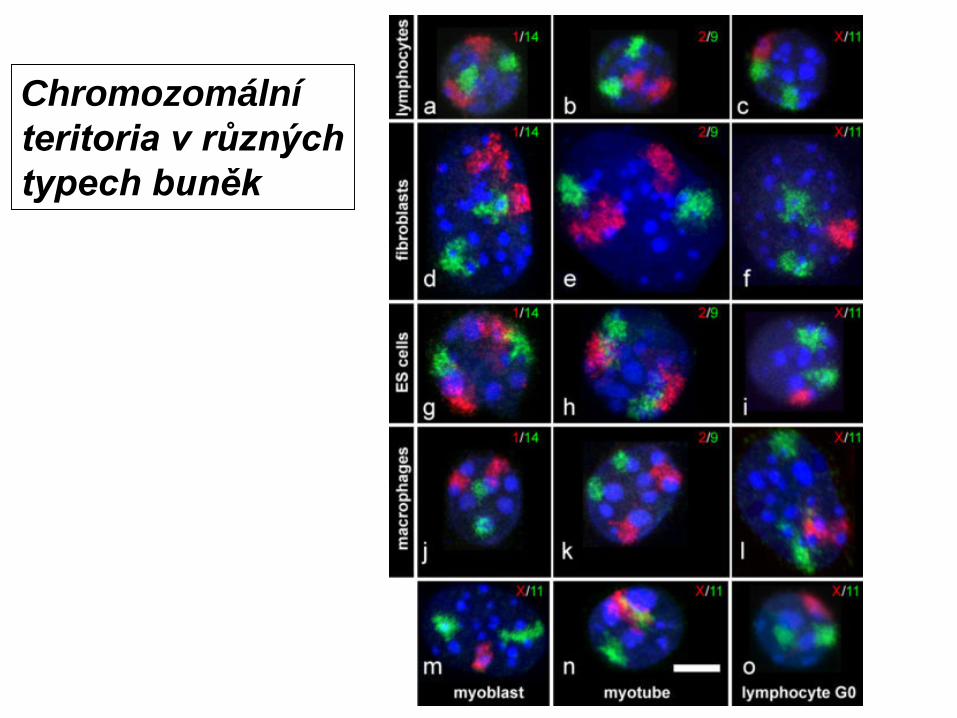
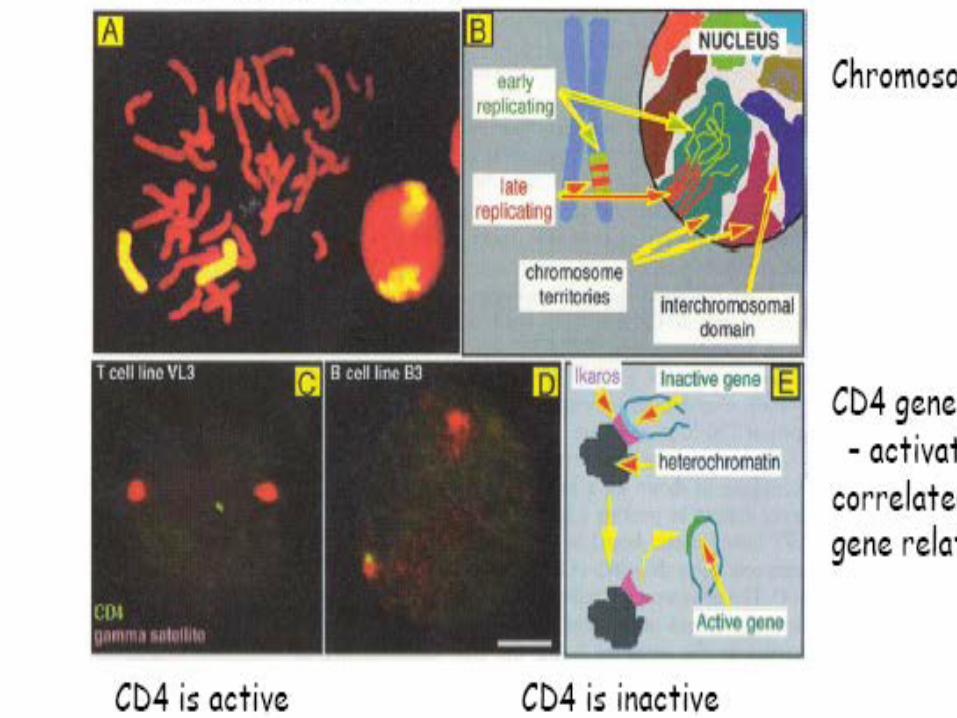
Human lymphocyte nucleus stained with DAPI with chromosome 13 (green) and 21 (red) centromere probes hybrydized (fluorescent in situ hybridization, FISH)
Main articles: Mechanical properties of DNA and Z-DNAThe vast majority of DNA within the cell is the normal DNA structure.
However in nature DNA can form three structures, A-, B- and Z-DNA. A and B chromosomes are very similar, forming right-handed helices, while Z-DNA is a more unusual left-handed helix with a zig-zag phosphate backbone. Z-
DNA is thought to play a specific role in chromatin structure and transcriptionbecause of the properties of the junction between B- and Z-DNA.
At the junction of B- and Z-DNA one pair of bases is flipped out from normal bonding. These play a dual role of a site of recognition by many proteins and as a sink for torsional stress from RNA polymerase or nucleosome binding.
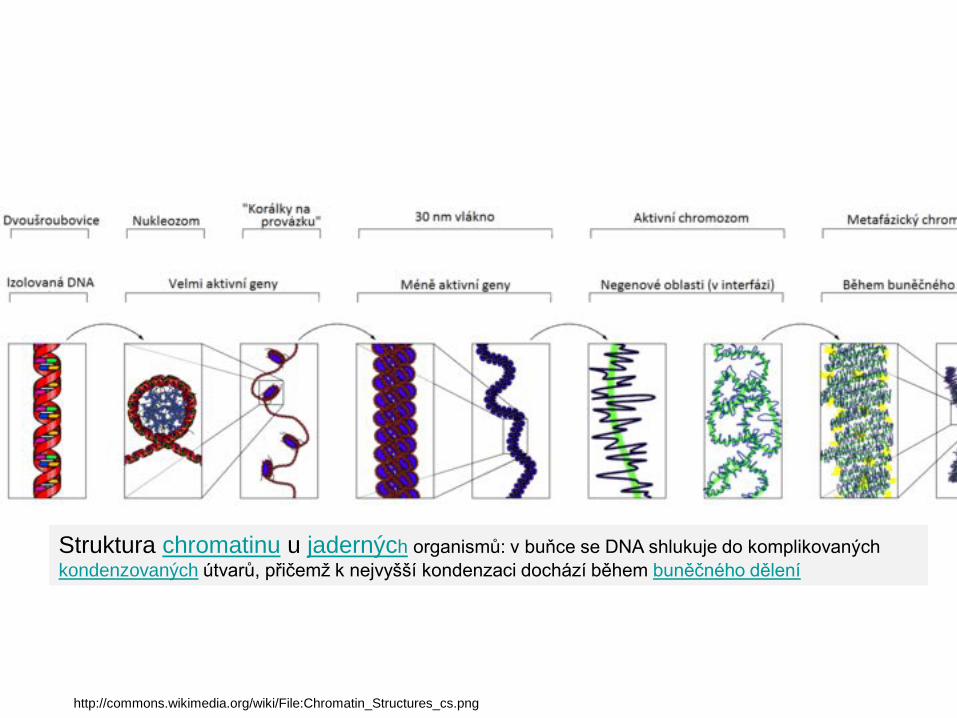
Struktura chromatinu u jaderných organismů: v buňce se DNA shlukuje do komplikovaných kondenzovaných útvarů, přičemž k nejvyšší kondenzaci dochází během buněčného dělení
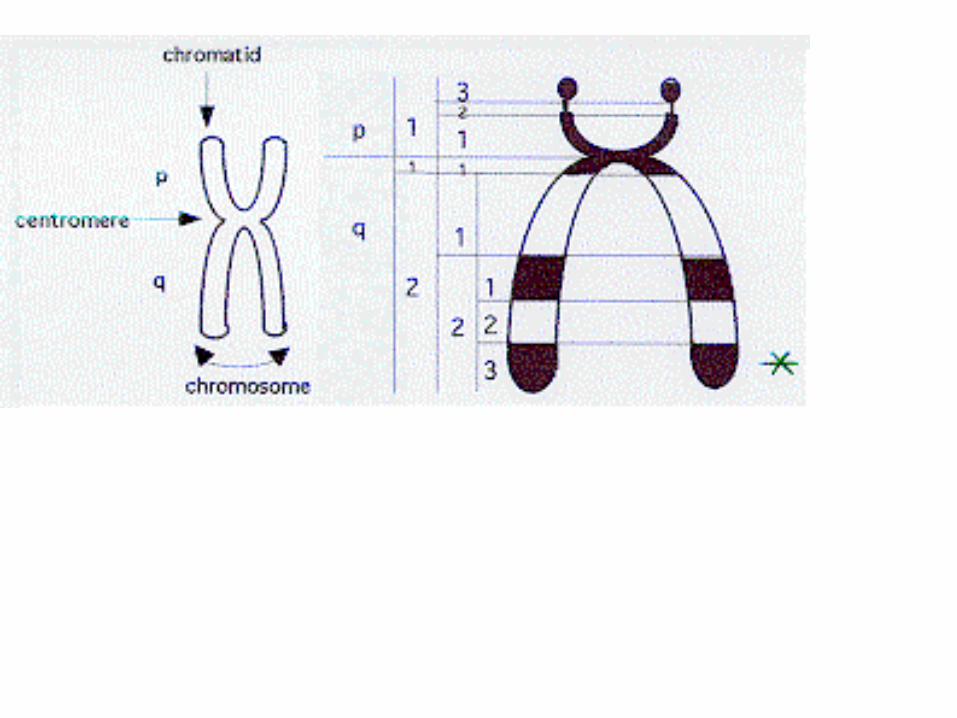
Morfologie ….chromozómu je nejlépe pozorovatelná ve stadiu metafáze nebo rané anafáze, v jiných fázích jaderného dělení je již zkreslován despiralizací (dekondezací). Chromozóm je …. tvořený zpravidla 2 podélně orientovanými chromatidami (výjimkou jsou tzv. polyténní chromozómy), které jsou propojeny centromerou v oblasti tzv. primární konstrikce, která rozděluje chromatidy na dvě ramena. U některých … chromozómů je ještě tzv. sekundární konstrikce, která odděluje z jedněch ramen tzv. satelit. Vnější koncové oblasti ramen chromatid (ty odvrácené od centromery) se nazývají telomery a u živočichů hrají důležitou úlohu v procesu stárnutí. Telomery, centromera a oblasti konstrikce mají strukturní funkci a neobsahují genetickou informaci.
Na základě umístění centromery rozlišujeme základní morfologické typy chromozómů: Metacentrický chromozóm - centromera je umístěna zhruba uprostřed chromozómu a dělí chromatidy na dvě zhruba stejně dlouhá ramena. Chromozóm má po replikaci DNA tvar blížící se písmenu X. Submetacentrický chromozóm - centromera je výrazněji posunuta směrem ke konci jedněch ramen.Akrocentrický chromozóm - centromera dělí chromatidy na jedno rameno velké a jedno malé uzlovité. Telocentrický chromozóm - centromera je umístěna u oblasti telomer a chromozóm se tak i v G2 fázi opticky jeví jako jednoramenný a připomíná písmeno V. ….u člověka se na rozdíl od ostatních nevyskytuje.
U chromozómů s různě dlouhými rameny se kratší rameno označuje p (původně z francouzského petit) a delší q (písmeno následující v abecedě po p). Ramena se dělí na oblasti a pruhy. Pro určení konkrétního místa v karyotypu se používá čtyřmístný kód, v němž první znak určuje chromozóm, druhý rameno, třetí oblast
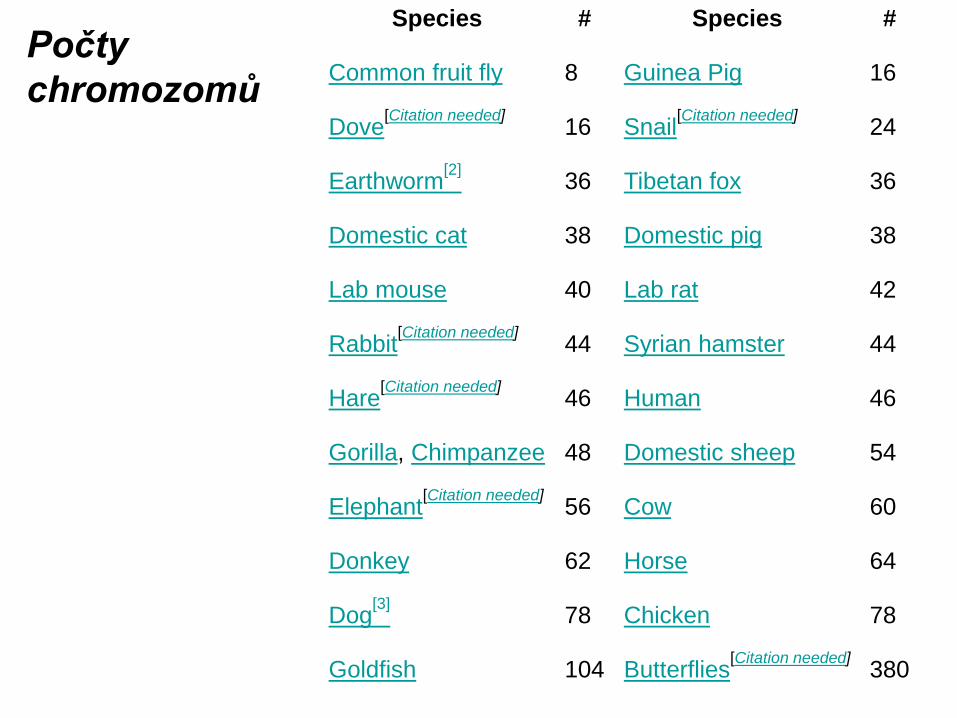
…buňky v organismu jsou zpravidla tzv. diploidní (2n), tj.chromozomy jsou v párech, každý chromozom se v buňce vyskytuje dvakrát. Jedná-li se o autozómy, mají oba chromozómy v páru alely stejných genů ve stejných lokusech, tyto alely ovšem mohou být různé (každá alela může být dominantní nebo recesivní). Jedná-li se o heterozómy (pohlavních chromozómy, u organismů typu Drosophila X a Y), platí totéž pouze pro jejich homonymní část, zatímco heteronymní části heterozómů jsou zcela různé (jak morfologií, tak geny, jejichž alely obsahují) - v této části jsou zakódovány znaky vázané na pohlaví.Kromě diploidních buněk existují běžně i buňky haploidní (n, každý chromozom jen jednou), zejména během dělení buněk nebo jako výsledek redukčního dělení. Umělou manipulací s genetickou informací mohou vznikat i buňky či celé organismy, které jsou polyploidní (tedy více než diploidní). Běžně se vysazuje například tetraploidní (4n) kukuřice. ….. chromozómy studuje genetika, manipulací s nimi se zabývá genetické inženýrství. cs.wikipedia.org/wiki/Chromozom
Vlákno nukleozomů je zhruba 10 nm silné. Oproti nahé DNA je 7x kratší.Vyšší struktura- chromatinové vlákno je silné 30 nm. Délku DNA zkracuje zhruba 40x. Dalším stáčením vzniká chromatida, která je zhruiba 700 nm silná
Průměrná délka DNA 1 chromozom člověka je 6 cm. Všech 46 chromozomů 2 metry
Chemické úpravy histonů, především acetylace jejich lyzinových N-konců, trčících z nukloeozomů, vedou k dekondenzaci chromatinu a „obnažení“ přiléhající DNA,která se stává přístupnou RNA polymeráze, zahajující transkribci genu.
Diagram of a duplicated and condensed metaphase eukaryotic chromosome. (1) Chromatid – one of the two identical parts of the chromosome after S phase. (2) Centromere – the point where the two chromatids touch, and where the icrotubules
attach. (3) Short arm. (4) Long arm. http://en.wikipedia.org/wiki/Chromosome
Dle polohy centromeryCh. metacentrické
akrocentrickételocentrické
holocentrické (kvasinky, C.elegans
Heterochromatin, DNA centromery je repetitivní-satelitní
Metacentric chromosomes have the centromere located near the middle so there are equal sized p and q arms. An example is chromosome 1.
Submetacentric chromosomes have the centromere located slightly off center so the q arm is slightly longer than the p arm. An example is chromosome 4.Acrocentric –
Acrocentric chromosomes have their centromeres very near one end so there is not much of a p arm. An example is chromosome 21.
Table of Denver Chromosome Classification Chromosome Group
Chromosome number
Description of chromosomes
A 1 - 3 Large, metacentric B 4 - 5 Large, submetacentric C 6 - 12 Medium-sized, submetacentric; X chromosome closely
resembles pair number 6, longest in group D 13 - 15 Medium-sized, acrocentric; all three pairs with satellites on
short arm E 16 - 18 Smallest of the medium-sized; pair 16 metacentric; others
submetacentric F 19 - 20 Short, metacentric G 21 - 22 very short; acrocentric; both with satellites
..human chromosomes in metaphase … with chromatid pairs. The telomeres stained with a green fluorescent dye on both chromatids in each replicated chromosome. http://www.ncsu.edu/project/bio183de/Lab/genetics1_lab/genetics1.html#act1
Humans do not have that many more genes than other organisms.E. coli = 4000 genes.Yeast = 6000 genes.Fruit fly = 12,000 genes.C. elegans = 19,000 genes.Humans = 30-40,000 genes.
Why are humans more sophisticated than other organisms?Human genes contain more introns than that of lower eukaryotes.Human proteins undergo more extensive protein modifications (phosphorylation/dephosphorylation).Humans share many genes with other organisms.Humans and chimp DNA is 98% identical.Humans and mice have almost identical genes.Flies have at least 100 genes that are also found in humans and cause human disease.Human chromosomes have undergone extensive chromosomal duplications.This may be a major driving force of human evolution.Chromosomal duplications result in large gene families.Paralogs are the genes that make up a gene family.http://pages.cabrini.edu/ddunbar/BIO263/chapter_13.htm
The term karyotype refers to a display of the chromosomes of a cell by lining them up, beginning with the largest chromosome and with the short arm of the chromosome oriented toward the top of the karyotype sheet
Proužkování (banding) chromozomů. Giemsa is a stain that stains chromosomal DNA. A-T rich areas of a chromosome stain more intensely than G-C rich areas. Therefore, chromosomes will contain dark-staining and light-staining bands
The banding pattern is unique for a given chromosome and can be used to identify a specific chromosome.
Další barvící/PROUŽKOVACÍ metody
Human ancestors had 48 chromosomes rather than 46. In the evolution of the human genome it is thought that two acrocentric chromosomes fused to create human chromosome 2.
The fusion of two non-homologous acrocentric chromosomes is called a Robertsonian translocation.
DNA of the human chromosome 2 was labeled, applied to Orang-Utan (left) and human metaphase chromosomes (right) by fluorescence in situ hybridization and detected in green. While in the human metaphase spread only the two copies of chromosome 2 were detected, in the Orang-Utan metaphase spread the two original chromosome pairs were painted: Human chromosome 2 is the evolutionary derived fusion product from two separate ancestral chromosomes. In non-human primates - like for example in the Orang-Utan (chromosomes on the left side) - as well as in several other mammals, two chromosomes are observed which carry human chromosome 2 orthologous genes and syntenic segments of orthologous DNA sequences The fusion in human 2q13-14 at approximately 114
Human karyotype is made of 46 chromosomes: 22 pairs of autosomes, numbered from 1 to 22 by order of decreasing length 1 pair of gonosomes, or sex chromosomes: XX in the female, XY in the male. Each chromosome has a centromere (CEN), region which contains the kinetochore, a micro-tubule organising centre (MTOC) responsible for attachment of the chromosome to the spindle apparatus at mitosis. The 2 sister-chromatids are principally held together at the para-centric heterochromatin at opposite ends of the centromeric region. CEN divides the chromosome into two arms: the short arm (p arm) and the long arm (q arm). Convention places the p-arm at the top in diagrammatic representations. Each arm terminates (pter, qter) in a telomere, a highly conserved repetitive gene sequence which inhibits end-end fusion, and which is important for attachments of chromosome ends to the nuclear envelope, particularly during meiosis. It is thought by many that diminution of telomere size is associated with cell ageing. When the short arm is nearly as long as the long arm, the chromosome is said metacentric; if it is shorter, the chromosome is said sub-metacentric; when it is very short, but still visible, the chromosome is said to be sub-telocentric; when extremely short, virtually invisible, the chromosome is said acrocentric. In the human karyotype, chromosome pairs 13, 14, 15, 21, 22 are acrocentric, and Y is sub-telocentric. In mammalian cells, the p-arm of many acrocentric chromosomes carry nucleolar organising regions (NORs) which contain genes coding for ribosomal RNA. This is true for all 5 pairs of acrocentrics in human cells.
Certain staining techniques cause the chromosomes to take on a banded appearance, each arm presenting a sequence of dark and light bands of varying intensities. Patterns are specific and repeatable for each chromosome, allowing unambiguous identification and longitudinal mapping for locating gene positions and characterising structural changes. The number of bands observed is not fixed but is related dynamically to the state of chromosome contraction. Thus, prophase chromosomes have many more bands than metaphase ones. Patterns, and the nomenclature for defining positional mapping have been standardised to allow cytogeneticists to communicate and archive information for medical purposes. Numbering begins from the centromere and continues outward to the end of each arm. The arms are divided into a number of regions by means of "land-mark" bands, and bands numbered sequentially within each. Sub-bands are catered for by using a decimal system. e.g. the place of the star in the Figure 2 is: 21q22.3http://atlasgeneticsoncology.org/Educ/PolyMecaEng.html
Vložit
LambrushGiant
EM Krajčí
Vložit další z Wikipedie vč. USB G
TEM póry naše vč. tabulky
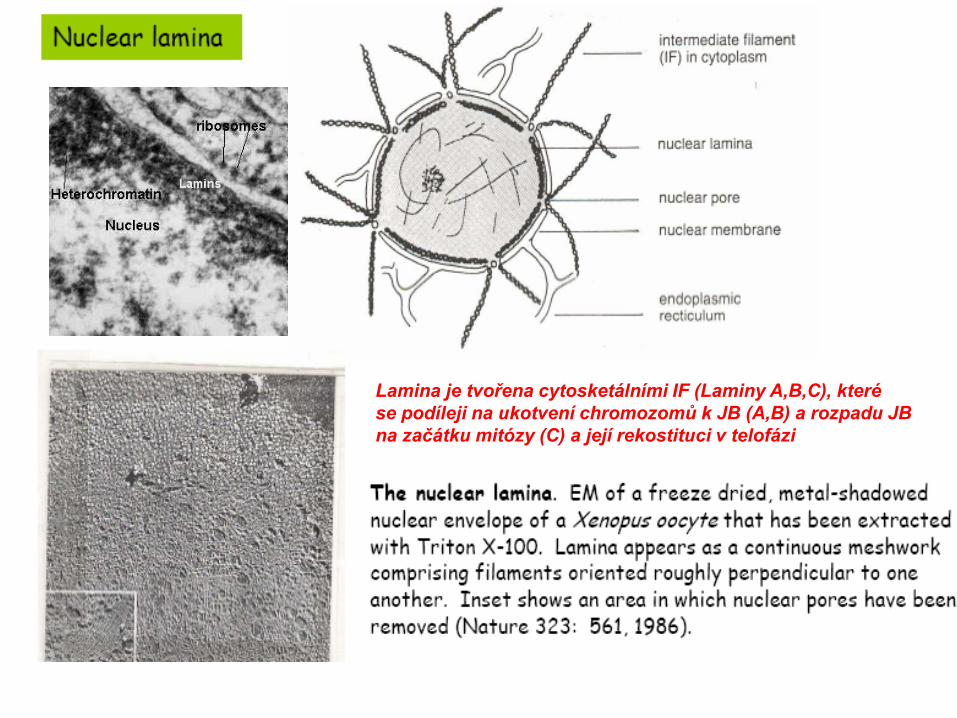
Lamina je tvořena cytosketálními IF (Laminy A,B,C), kterése podíleji na ukotvení chromozomů k JB (A,B) a rozpadu JB na začátku mitózy (C) a její rekostituci v telofázi
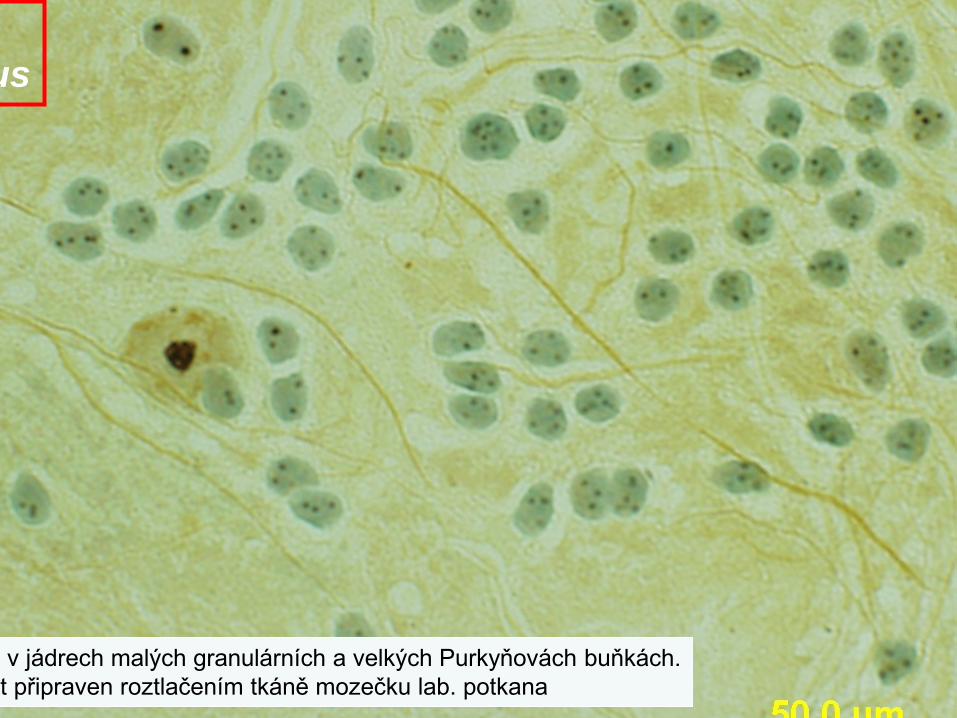
v jádrech malých granulárních a velkých Purkyňovách buňkách.t připraven roztlačením tkáně mozečku lab. potkana
us
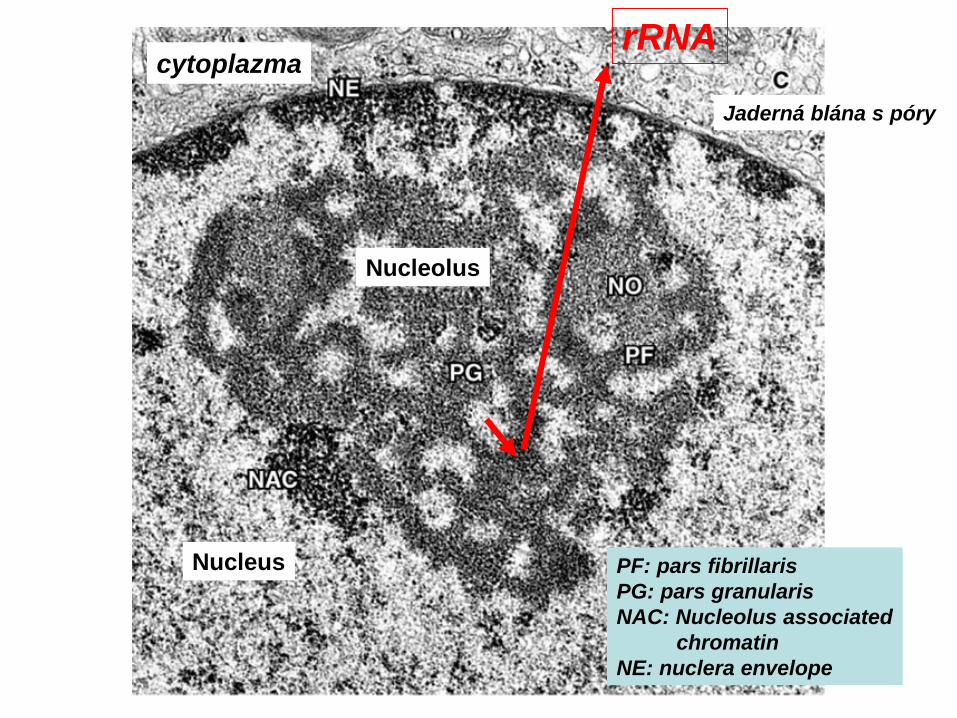
Nucleus
Nucleolus
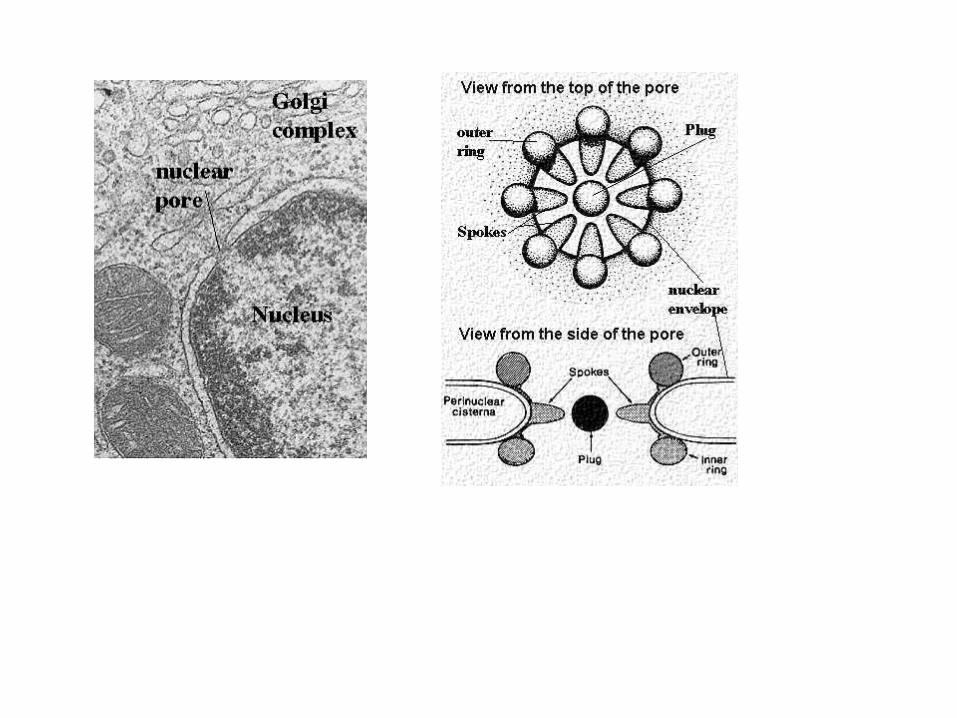
Jaderná blána s póry
rRNAcytoplazma
PF: pars fibrillarisPG: pars granularisNAC: Nucleolus associated
chromatinNE: nuclera envelope
Transkripce r-DNA genů v jadérku„Christmas trees“
Other DNA http://en.wikipedia.org/wiki/Human_genome#Other_DNA
Protein-coding sequences (specifically, coding exons) comprise less than 1.5% of the human genome.[3] Aside from genes and known regulatory sequences, the human genome contains vast regions of DNA the function of which, if any, remains unknown. These regions in fact comprise the vast majority, by some estimates 97%, of the human genome size. Much of this is composed of Repeat elements:Tandem repeats
http://pages.cabrini.edu/ddunbar/BIO263/new_page_1.htm Viz F USB, Ad b. cyklus
In normal cells, p53 levels are low. Mdm2 removes p53 from the nucleus and leads to its degradation by the proteasome.
Damage to DNA results in p53 phosphorylation (P) and acetylation (Ac), and activation of p53 as a transcription factor.There are three major checkpoints in the cell cycle.
1. A DNA damage checkpoint.2. A centrosome duplication checkpoint.
P53 is a tumor-suppressor gene. Tumor-supressor genes are genes that normally negatively control cell proliferation or that activate the apoptotic pathway, in which loss-of-function mutations contribute to cancer progression.Some well characterized tumor-suppressor genes are p53, p21, p16/p19ARF, RB, Bax and Bub1.Oncogenes are gain-of-function mutations associated with cancer progression. They derive from normal genes called proto-oncogenes. Oncogenes improperly enhance the expression of genes that promote cell proliferation or inhibit apoptosis.Cancer is not one disease but rather many diseases that share similar attributes: The cells show uncontrolled growth as a result of mutations that affect a limited number of genes. It is a disease of somatic cells.About 1% of cancer cases are familial, inherited. The other 99% of cases are sporadic, and are the result of genetic changes only in somatic cells.Cancer cells lose the property of contact inhibition. In normal cells, cell-to-cell contact inhibits further growth and cell division. This is referred to as contact inhibition.
Normal cells also cease to divide in culture after about 50 doublings (a process called cell senescence). Cancer cells in culture divide indefinitely. Most cancer cells re-activate the enzyme telomerase which allows them to divide indefinitely.Two Hits, Two Errors hypothesis for cancer development. Both alleles of a particular gene must be mutated in a somatic cell in order for it to potentially become cancerous.