372
Příroda Č Í S L O 3 1 P R A H A • 2 0 1 1
| Date post: | 09-Feb-2017 |
| Category: |
Documents |
| Upload: | truongkhanh |
| View: | 270 times |
| Download: | 2 times |

PřírodaČ Í S L O 3 1
P R A H A • 2 0 1 1

ZuZana MünZbergová, Lucie Černá & JarMiLa gabrieLová [eds]
PRioRity dRuHové ocHRAnycévnAtýcH Rostlin

ZuZana MünZbergová, Lucie Černá & JarMiLa gabrieLová [eds]
PRioRity dRuHové ocHRAnycévnAtýcH Rostlin


Příroda, Praha, 31: 5–9, 2011
5
Priority druhové ochrany cévnatých rostlin České republiky
ZuZana MünZbergová1, 2, JarMiLa gabrieLová1, 3 & Lucie Černá3
1Katedra botaniky, Přírodovědecká fakulta Univerzity Karlovy, Benátská 2, CZ-128 01 Praha 2; [email protected]; [email protected]ý ústav Akademie věd České republiky, v. v. i., Zámek 1, CZ-252 43 Průhonice3Agentura ochrany přírody a krajiny České republiky, Nuselská 34, CZ-140 00 Praha 4; [email protected]
Prezentované monotematické číslo sborníku Příroda se věnuje výsledkům projektu Priority druhové ochrany cévnatých rostlin ČR (VaV 2B06178) řešeného Botanickým ústavem AV ČR, Agenturou ochrany přírody a krajiny ČR a Sdružením pro ochranu přírody střední Moravy Sagittaria. Prvotní myšlenkou a hlavním cílem projektu, který probíhal v letech 2006–2011, bylo utřídit naše znalosti o kriticky ohrožených druzích rostlin České republiky (podle Černého a Červeného seznamu cévnatých rostlin České republiky, Procházka 2001). Těchto druhů se v naší flóře vyskytuje více než 470, což početně silně převyšuje personální a finanční možnosti monitoringu a ochrany všech těchto druhů. Naším cílem bylo vytvořit přehled o biologických vlastnostech (Gabrielová et al. 2011a), stanovištních nárocích a současném a historickém rozšíření všech těchto druhů (Černá et al. 2011). Z důvodu ohromného množství potřebných dat bylo při naší práci nutné spoléhat z velké části na již existující údaje, tyto údaje shromáždit, utřídit a vyhodnotit. Zejména v případě dat o současném a historickém rozšíření druhů v ČR práce představovala kompilaci ohromného počtu jednotlivých údajů (Černá et al. 2011) a je zřejmé, že výsledný soubor není zcela kompletní a může obsahovat chyby. Naše výsledky proto nepovažujeme za konečný přehled stavu našich kriticky ohrožených druhů. Spíše se jedná o první ucelenější přehled, jehož cílem je mimo jiné vyvolat diskuzi na toto téma a přimět osoby disponující podrobnějšími znalostmi o jednotlivých druzích k doplnění našich přehledů. Získaný materiál byl upraven také do formy databáze přístupné na stránkách AOPK ČR a bude tak přímo k dispozici pracovníkům ochrany přírody (http://portal.nature.cz/c1/rostliny/).
Výsledky práce týkající se vlastností a rozšíření všech kriticky ohrožených druhů nám umožnily stanovit, které druhy jsou vzácné, avšak s víceméně stabilním počtem lokalit v čase (např. Agrostis alpina, Aposeris foetida, Hypericum pulchrum, Minuartia corcontica, Primula minima a Saxifraga oppositifolia), a odlišit je od druhů, které sice byly dříve hojněji rozšířené, ale v čase výrazně ubyly (např. Arnoseris minima, Cyperus flavescens, Centaurium littorale subsp. compressum, Gentianella germanica subsp. germanica, Pseudognaphalium luteo-album a Radiola linoides) a zaslouží si tedy aktivní ochranu (Gabrielová et al. 2011c).
Důležitou součástí dat o kriticky ohrožených druzích byly i údaje o celkovém areálu druhů a jejich ohrožení v okolních zemích (Gabrielová et al. 2011b). Identifikovali jsme druhy, které jsou v ČR vzácné, ale mají velký areál, v řadě ostatních zemí jsou velmi hojné (např. Carex atrata, Luzula spicata, Poa alpina, Spergularia salina nebo Xanthium strumarium) a jejich ochrana tak není z celosvětového hlediska velkou prioritou. Na druhou

6
Münzbergová et al.: Priority druhové ochrany cévnatých rostlin
stranu jsme identifikovali druhy s malým areálem nebo vzácné v celém svém areálu (např. Artemisia pancicii, Dracocephalum austriacum, Epipactis nordeniorum, Galium sudeticum, Ligularia sibirica, Pedicularis exaltata, Pedicularis sudetica, Tephroseris longifolia subsp. moravica), za jejichž ochranu má Česká republika velkou zodpovědnost.
Posledním významným kritériem pro hodnocení, které druhy si zasluhují prioritní ochranu, byla informace o tom, zda se druhy vyskytují na stále stejných místech, nebo zda se jejich lokality obměňují. Obě skupiny těchto druhů samozřejmě zasluhují velkou pozornost, nicméně přístupy k jejich ochraně se budou lišit. U druhů s výskytem na stále stejných místech lze však ochrany dosáhnout s menším úsilím a jejich ochrana tak bude pravděpodobně úspěšnější. Mezi prioritní druhy u nás silně ubývající, výrazně ohrožené v ostatních evropských zemích a vyskytující se na stále stejných lokalitách patří např. Gentianella campestris subsp. baltica, Hammarbya paludosa, Pulsatilla patens a Scorzonera parviflora. Druhy jako Botrychium matricariifolium, Campanula cervicaria, Littorella uniflora, Spiranthes spiralis a Tillaea aquatica jsou ohrožené do stejné míry jako druhy zmíněné výše. Jejich ochrana je však komplikována faktem, že lokality jejich výskytu se v čase obměňují a jejich ochranu tak není vždy možné zajistit pouhou ochranou jejich stanovišť (Gabrielová et al. 2011c).
Práce pouze se souborem kriticky ohrožených druhů v sobě nese nebezpečí, že při našem hodnocení prioritních druhů přehlédneme druhy, které jsou na silném ústupu a zasloužily by si prioritní aktivní ochranu, ale v současnosti jsou klasifikované v kategorii silně ohrožené či kategorii nižší. Zachycení těchto druhů by samozřejmě bylo velmi zajímavé a cenné, nicméně vzhledem k jejich počtu to již překračuje rámec našeho projektu a zůstává tak námětem pro další práci.
Druhou velkou částí prezentovaného projektu jsou modelové studie věnující se populační dynamice, stanovištním nárokům a genetické diverzitě vybraných kriticky ohrožených druhů. V rámci této části jsme se pokusili sebrat maximum informací o životním cyklu vybraných druhů a získat tak představu o očekávaném vývoji populací těchto druhů do budoucna. Současně jsme studovali stanovištní podmínky, v nichž se druhy vyskytují, a genetickou diverzitu populací vybraných druhů. Získané informace nám umožnily identifikovat, které populace daných druhů jsou stabilní či dokonce rostoucí a které jsou naopak ve fázi silného poklesu. Oproti pouhému sledování vývoje počtu jedinců v populacích jsme tímto přístupem získali informace o očekávaném poklesu populací některých druhů bez toho, aby došlo ke skutečnému poklesu počtu dospělých jedinců v populaci během sledovaného období. Tímto nám data o populační dynamice poskytují nástroj časného varování, protože nám umožní identifikovat neblahé procesy v populacích dlouho předtím, než dojde ke skutečnému pozorovatelnému úbytku druhu. Současně jsme mohli zhodnotit stanovištní podmínky, za kterých dochází k poklesu velikosti populací druhu, a identifikovat tak vhodný management lokalit.
Výsledky populačních studií ukázaly, že populace druhů Orchis tridentata a Spiranthes spiralis jsou stabilní a stávající management na jejich studovaných lokalitách je vyhovující (Dostalík et al. 2011, brabec et al. 2011b). Podobně stabilní se jeví i populace druhu Dracocephalum austriacum. V tomto případě se ovšem ukazuje, že pro udržení stabilních populací je kritické zachovat alespoň stávající genetickou diverzitu populací. Její pokles totiž významně snižuje produkci semen, která je klíčová pro dlouhodobé přežití druhu na lokalitách (Dostálek & MünzberGová 2011a). Relativně stabilní populace má i druh Artemisia pancicii. Druh však velmi málo kvete a přesto, že produkuje životaschopný pyl, nebyla na lokalitách nikdy zaznamenána vyvinutá semena. To ukazuje, že tento druh přežívá na lokalitách pouze vegetativně. Jakákoliv větší disturbance lokalit tak může vést k jejich zániku (Gillová et al. 2011).

Příroda, Praha, 31: 5–9, 2011
7
Stabilní populace mají i zástupci kapradin, modelové druhy Asplenium adulterinum a A. cuneifolium. Stabilita těchto populací je do velké míry dána dlouhověkostí jednotlivých rostlin. Navzdory obecné představě o všudypřítomnosti spor se ovšem ukazuje, že v krajině existují vhodné, leč neobsazené lokality pro oba tyto druhy. To na jednu stranu naznačuje, že existuje teoretický potenciál k dalšímu rozšíření těchto druhů. Na druhou stranu ale nízký skutečný kolonizační potenciál druhů naznačuje, že ztráta každé jednotlivé populace se může významně projevit na schopnosti druhu přežít v této krajině. Jako největší nebezpečí se jeví výsadba smrku na hadcových skalkách, v jehož opadu druhy nedokáží růst (bucharová et al. 2011).
Relativní stabilitu ukazují i populace druhu Gentianella praecox subsp. bohemica (brabec et al. 2011a), ale pouze v případě, kdy jsou lokality sečeny a disturbovány. Tento typ managementu sice přímo poškozuje jednoleté růžice druhu, přesto však vede k výraznému zvýšení růstové rychlosti oproti pouhé seči díky vytváření vhodných mikrostanovišť pro klíčení.
Zatímco populace výše zmíněných druhů jsou víceméně stabilní, další studované druhy ukazují klesající tendence. Studie poloparazita Thesium ebracteatum naznačuje, že počet prýtů v jediné velké české populaci v posledních letech klesá (nedávno byla u nás nalezena druhá malá populace druhu; Prach & zajíČková 2009). Krátká časová řada spolu s problémy při odlišování semenáčků od vegetativních prýtů však neumožňuje stanovit, zda se jedná o dlouhodobý trend či pouze fluktuaci. Zajímavým výsledkem této studie je zjištění, že poloparazité z rodu Thesium si svoje hostitele nijak specificky nevybírají a parazitují na všech hojných druzích ve společenstvu. Díky této vlastnosti lze očekávat, že druhy tohoto rodu budou méně citlivé na změnu hostitelského společenstva než (polo)parazitické druhy s pevnou vazbou na hostitele (Dostálek & MünzberGová 2011b).
Nejméně početným druhem mezi modelovými druhy je druh Lathyrus pisiformis, který se na dvou lokalitách v ČR vyskytuje celkově v počtu kolem 200 prýtů. Zatímco počet prýtů byl po celou dobu sledování poměrně stabilní, celková produkce semen na lokalitách je velmi variabilní a naznačuje možné problémy při generativní reprodukci druhu (Štefánek 2011). Z dlouhodobého hlediska tedy můžeme očekávat jeho další úbytek a vzhledem k jeho početnosti je zde velké riziko vymření na našem území.
Klesající trend je patrný i u některých populací Ligularia sibirica. Zde se jasně ukazuje kritický význam udržení vhodných stanovištních podmínek pro přežití druhu. Malá velikost populace ani snížená genetická diverzita nemají na přežití druhu výrazný negativní vliv. Nárůst obsahu dusíku v půdě však vede k silnému poklesu růstové rychlosti populací a ochrana tohoto vzácného druhu by se tak měla zaměřit hlavně na obnovení stanovištních podmínek degradovaných lokalit (heinken-ŠMíDová & MünzberGová 2011).
Populace tří studovaných druhů kuřiček, Minuartia smejkalii, M. corcontica a M. caes-pi tosa, se vyznačují velkou variabilitou. Přestože některé trsy všech studovaných druhů jsou velmi dlouhověké, populace se vyznačují velkým obratem a každý rok je více než polovina jedinců v populaci obměněna. V případě poklesu produkce semen tak může dojít k rychlému úbytku druhů. To je nebezpečné zejména u druhu M. corcontica, který netvoří vytrvalou semennou banku. V průběhu sledování byl navíc u tohoto druhu pozorován pokles počtu kvetoucích jedinců (Pánková & MünzberGová 2011).
Na rozdíl od předchozích studií nebylo hlavním zájmem studie modelového poddruhu Thymus pulegioides subsp. carniolicus popsat dynamiku populace, ale zejména zhodnotit nebezpečí křížení tohoto poddruhu s ostatními příbuznými taxony. Výsledky ukazují poměrně časté křížení zájmového taxonu s Thymus pulegioides subsp. chamaedrys. Velkým problémem je fenologická synchronizace obou taxonů. Jediným vhodným managementem se tak jeví ruční vytrhávání Thymus pulegioides subsp. chamaedrys (ČáP & kleinová).

8
Münzbergová et al.: Priority druhové ochrany cévnatých rostlin
Jednotlivé modelové studie spolu s přehledem vlastností všech kriticky ohrožených druhů rostlin ukazují, že zkoumaný druhový soubor je velmi heterogenní. Jeho heterogenita jednoznačně vede k tomu, že pro ochranu těchto druhů není bohužel možné využít jediný standardizovaný přístup. Každý druh se totiž jeví jako unikát, jehož ochrana vyžaduje speciální postupy. Možnosti přenosu konkrétních poznatků mezi blízce příbuznými či morfologicky a stanovištně podobnými druhy tak zůstává námětem pro další výzkumy.
Celý předkládaný sborník představuje hlavní výsledky získané v prezentovaném projektu. Oficiálním koncem projektu a vydáním tohoto sborníku však naše práce věnující se kriticky ohroženým druhům ČR zdaleka nekončí. Proto pokud vás přečtení tohoto sborníku inspirovalo k otevření šuplíků a vytažení starých zápisníků s údaji o některém ze studovaných druhů, naše práce splnila svůj účel. Neváhejte, prosím, a určitě nás kontaktujte. Za údaje ohledně opomenutých lokalit, špatně lokalizovaných populací a jejich velikostech či ohledně dalších detailů z biologie našich kriticky ohrožených druhů budeme velmi vděční. Naší snahou bude výsledky tohoto projektu průběžně aktualizovat a získat tak živý zdroj informací o kriticky ohrožených druzích květeny ČR, který by do budoucna zajistil skutečně smysluplnou a efektivní ochranu těchto druhů.
Poděkování
Celý projekt by nevznikl bez prvotního nadšení Martiny Fialové, které tímto velmi děkujeme. Tato práce byla podpořena projektem MŠMT VaV 2B06178. Částečně vznikla také díky projektům MŠMT 0021620828 and AV0Z60050516.
literatura
brabec j., bucharová a. & Štefánek M. (2011a): Vliv obhospodařování na životní cyklus hořečku mnohotvarého českého (Gentianella praecox subsp. bohemica). – Příroda, Praha, 31: 85–109.
brabec j., iPser z., jersáková j., Malinová t., balounová z., Plesková l., soMol v., brabec j., krinke l., Štefánek M. & nesvaDbová j. (2011b): Populační dynamika, energetická náročnost kvetení a přežívání švihlíku krutiklasu (Spiranthes spiralis) na tradičně obhospodařované lokalitě „NPP Pastviště u Fínů“. – Příroda, Praha, 31: 185–200.
bucharová a., tájek P. & MünzberGová z. (2011): Dynamika dvou vzácných druhů kapradin (Asplenium adulterinum a A. cuneifolium) na lokální a krajinné úrovni. – Příroda, Praha, 31: 33–52.
ČáP j. & kleinová H. (2011): Současné poznatky o mateřídoušce vejčité kraňské (Thymus pulegioides subsp. carniolicus) v České republice. – Příroda, Praha, 31: 227–246.
Černá l., Gabrielová j., MünzberGová z. & rybka v. (2011): Změny v rozšíření kriticky ohrožených druhů rostlin v České republice na základě síťových map. – Příroda, Praha, 31: 281–298.
Dostálek t. & MünzberGová z. (2011a): Populační dynamika a genetická diverzita kriticky ohroženého druhu Dracocephalum austriacum. – Příroda, Praha, 31: 53–83.
Dostálek t. & MünzberGová z. (2011b): Stanovištní nároky a výběr hostitelů u poloparazitů rodu lněnka (Thesium). – Příroda, Praha, 31: 201–225.
Dostalík s., rybka v., zeMánková-bartková r. & baláž M. (2011): Monitoring vstavače trojzubého (Orchis tridentata) v Národní přírodní rezervaci Strabišov-Oulehla v období 1997-2010. – Příroda, Praha, 31: 167–183.
Gabrielová j., MünzberGová z., husáková i. & chrtek J. (2011a): Kriticky ohrožené druhy rostlin České republiky: Jak se odlišují svými biologickými a ekologickými vlastnostmi od hojných druhů? – Příroda, Praha, 31: 247–279.
Gabrielová j., fialová t. & MünzberGová z. (2011b): Jak je to s ohrožením kriticky ohrožených druhů rostlin České republiky v jiných evropských zemích? – Příroda, Praha, 31: 299–345.
Gabrielová j., Černá l. & MünzberGová z. (2011c): Jak souvisí změny v rozšíření kriticky ohrožených druhů rostlin České republiky s jejich biologickými vlastnostmi a ekologickými nároky? – Příroda, Praha, 31: 345–370.

Příroda, Praha, 31: 5–9, 2011
9
Gillová l., kitner M., Majeský Ľ., skálová D. & vyMyslický t. (2011): Nové poznatky o druhu pelyněk Pančićův (Artemisia pancicii). – Příroda, Praha, 31: 11–31.
heinken-ŠMíDová A. & MünzberGová z. (2011): Faktory ovlivňující populační dynamiku kriticky ohroženého druhu Ligularia sibirica. – Příroda, Praha, 31: 127–147.
Pánková h. & MünzberGová z. (2011): Populační biologie a stanovištní nároky druhů z okruhu Minuartia verna agg. – Příroda, Praha, 31: 149–165.
Prach j. & zajíČková l. (2009): Thesium ebracteatum Hayne. − In: haDinec j. & lustyk P. [eds], Additamenta ad floram Reipublicae Bohemicae. VIII. Zpr. Čes. Bot. Spol. 44, pp. 309−310.
Štefánek M. (2011): Hrachor hrachovitý (Lathyrus pisiformis) v České a Slovenské republice. – Příroda, Praha, 31: 111–126.

10
Münzbergová et al.: Priority druhové ochrany cévnatých rostlin

Příroda, Praha, 31: 11–31, 2011
11
nové poznatky o druhu pelyněk Pančićův (Artemisia pancicii)
new knowledge on Artemisia pancicii
Lenka giLLová1, MiLosLav kitner2, Ľuboš MaJeský2, dagMar skáLová2 & toMáš vyMysLický3
1Agentura ochrany přírody a krajiny České republiky, Lafayettova 13, CZ–772 00, Olomouc; [email protected]; [email protected] botaniky Přírodovědecké fakulty, Univerzita Palackého Olomouc, Šlechtitelů 11, CZ-783 71 Olomouc; [email protected]; [email protected]; [email protected] 3Zemědělský výzkum, spol. s r. o., Zahradní 1, CZ-664 41 Troubsko; [email protected]
Abstract: The Pannonian endemic species Artemisia pancicii is one of the most endangered plant species. Its area is very fragmented and its current occurrence is known only from ten localities in the Czech Republic, Austria and Serbia. Historically, only six localities are known in the Czech Republic, and its recent occurrence has been observed only in three localities in Southern Moravia. The species is recorded from six localities in Austria – from Niederösterreich (Lower Austria) and Burgenland. One locality is to be found in Serbian Vojvodina, Deliblatska peščara. Artemisia pancicii forms compact polycormons of vegetative spreading shoots in the majority of localities. The Czech and Austrian populations rarely flower. Since 2004, the species populations have been regularly monitored on seven permanent plots (since 2007 on nine plots). Artemisia pancicii was chosen among the model plants in the project VaV 2B06178 – Protection Priorities in Vascular Plants in 2006. Under this project, the viability of pollen grains and pollen development stages were tested on sixteen plants selected from all flowering populations. The possibility of generative reproduction was tested. In order to gain an understanding of the ecological relationships the phytosociological relevés were recorded and soil samples were taken in all Czech localities. The genetic variability was analysed on 82 plants from the Liščí kopec I locality using the AFLP method.Keywords: AFLP, Artemisia pancicii, endangered species, Natura 2000, viability of pollen grains
Abstrakt: Panonský endemit Artemisia pancicii (pelyněk Pančićův) patří mezi nejohroženější druhy středoevropské flóry. Jeho areál je značně fragmentován a v současnosti se jeho výskyt uvádí z deseti lokalit na území České republiky, Rakouska a Srbska. V České republice je historicky evidováno pouze šest lokalit na jižní Moravě, recentně jsou známy pouze tři lokality. Z Rakouska je recentně druh uváděn z šesti lokalit v Dolních Rakousích a v Burgenlandu. Jedna lokalita leží v srbské Vojvodině na lokalitě Deliblatska peščara. Artemisia pancicii na většině lokalit vytváří kompaktní polykormony vegetativně se rozrůstajících prýtů. České a rakouské populace kvetou velmi zřídka. Od roku 2004 jsou na sedmi (od roku 2007 na devíti) trvalých plochách tyto populace pravidelně monitorovány. V roce 2006 byl druh vybrán mezi modelové druhy projektu VaV 2B06178 - Priority druhové ochrany cévnatých rostlin. V rámci tohoto projektu byla u 16 vy-bra ných rostlin ze všech kvetoucích populací testována životnost a vývojové stadium pylových zrn. Byla studována možnost generativní reprodukce. Pro pochopení ekologických vazeb druhu byly na všech českých lokalitách zapsány fytocenologické snímky a odebrány půdní vzorky. Metodou AFLP byla analyzována genetická variabilita 82 jedinců z lokality Liščí kopec I.

12
Gillová et al.: Nové poznatky o Artemisia pancicii
Klíčová slova: AFLP, Artemisia pancicii, Natura 2000, ohrožený druh, životnost pylových zrn nomenklatura: hejný & slavík 1988, 1990, 1992; slavík 1995, 1997, Moravec 1995
Úvod
Artemisia pancicii patří k jednomu z nejvzácnějších druhů středoevropské flóry. Podle klasického pojetí Rabinowitze (rabinowitz (1981) patří A. pancicii do skupiny přirozeně vzácných druhů s malým areálem, s úzkou ekologickou valencí a s převládající nízkou početností jednotlivých populací. Recentní areál druhu zahrnuje deset lokalit v České republice, Rakousku a Srbsku. Druh vyhynul na třech lokalitách v České republice (Danihelka 1995), historické rozšíření druhu v Srbsku a Rakousku není autorům článku známé. Většina populací je malá, izolovaná, zahrnující desítky až stovky prýtů. Pouze populace na Liščím kopci I a na Bisambergu jsou bohatší, čítající tisíce ramet.
Publikovaných dat o tomto druhu je velmi málo. Výjimkou jsou práce zabývající se nomenklaturou druhu (Danihelka & MarholD 2003), rozšířením druhu v České republice (Danihelka 1995), v Rakousku (naGler 2010) a v Srbsku (boža 1999), diplomová práce zaměřena na biologii, ekologii a rozšíření druhu (bulová 2002) a souhrnné informace v Květeně ČR (Grulich 2004) a Červené knize ČR a SR (holub & Grulich 1999). Podrobnější monitoring českých populací začal v roce 2004 v rámci úkolu sledování evropsky významných druhů (tzv. naturových). V roce 2006 byl druh vybrán mezi modelové druhy projektu VaV 2B06178 Priority druhové ochrany cévnatých rostlin. V rámci tohoto projektu se začala sledovat populační dynamika druhu na třech lokalitách, studovaly se ekologické nároky druhu, provedl se výzkum genetické variability našich a rakouských populací, sledovala se životaschopnost pylových zrn a možnost generativní reprodukce druhu. Cílem tohoto článku je shrnout dílčí výsledky tohoto projektu. Výsledky studia genetické variability českých a rakouských populací jsou shrnuty v samostatné práci (kitner et al. in prep.).
studované lokality
Liščí kopec I
Lokalita je součástí Národní přírodní rezervace Dunajovické kopce, druh roste na západním svahu jižního dvojvrcholu Liščího kopce (271,7 m). Druh zde byl objeven Podpěrou v roce 1923 (Danihelka 1995). Geologický podklad území tvoří vápnité jíly s proužky písku, hlavním půdním typem je zde černozem pelická (bulová 2002). Druh se vyskytuje ve vegetaci svazu Bromion erecti Koch 1926. Lokalita je nepravidelně kosena, naposledy bylo ca 90 % plochy s výskytem A. pancicii pokoseno v srpnu roku 2006.
Liščí kopec II
Lokalita je součástí Národní přírodní rezervace Dunajovické kopce, leží cca 90 m se vero-západně od kóty Liščího kopce (271,7 m), ca 185 m severovýchodně od lokality Liščí kopec I. Studovaný druh roste na jihozápadním svahu Liščího kopce, těsně pod zlomem svahu. Geo lo gický podklad území tvoří vápnité jíly s proužky písku, hlavním půdním typem je zde černozem pelická (bulová 2002). Artemisia pancicii se vyskytuje ve vegetaci svazu Bromion erecti Koch 1926. Plocha byla ručně pokosena v letech 2005–2007 ve druhé polovině vegetační sezóny.

13
Příroda, Praha, 31: 11–31, 2011
Liščí kopec III
Lokalita byla objevena J. Danihelkou v roce 1994 (Danihelka 1995), leží ca 60 m jihovýchodně od kóty Liščího kopce (271,7 m), ca 200 m VSV od lokality Liščí kopec I, těsně za hranicemi NPR Dunajovické kopce. Druh roste na severovýchodním svahu Liščího kopce, na zarůstajících terasách, těsně pod rozbitým plotem. Geologickým podkladem území jsou stejně jako u lokalit Liščí kopec I–II vápnité jíly s proužky písku, hlavním půdním typem je zde černozem pelická (Bulová 2002). Druh roste ve vegetaci svazu Bromion erecti Koch 1926. Plocha s výskytem A. pancicii je dlouhodobě nekosena, zarůstá druhy Calamagrostis epigejos, Dactylis glomerata, Bromus inermis a Prunus fruticosa, na severovýchodním okraji do plochy expanduje Prunus spinosa.
Pouzdřany
Lokalita leží na jihovýchodním svahu Pouzdřanské stepi, je součástí Národní přírodní rezervace Pouzdřanská step – Kolby. Pelyněk Artemisia pancicii zde objevil R. Picbauer v roce 1924 (Danihelka 1995). Druh roste v horních partiích svahu, v okolí turistické stezky vedoucí k lesu Kolby. Geologické podloží lokality tvoří vápnité jíly, slíny a pískovce, z půdních typů zde převládá lehčí střední půda typu černozem (bulová 2002). Artemisia pancicii roste ve vegetaci svazu Bromion erecti Koch 1926. Populace byla na konci osmdesátých let na pokraji zániku (Grulich 1987), po roce 1988, kdy lokalita vyhořela, došlo k velkému nárůstu početnosti druhu (Danihelka 1995). Na lokalitě probíhá nepravidelný management, v letech 2006, 2007 a 2009 byla plocha extenzivně pasena stádem ovcí, horní část populace je ale stále ohrožena zarůstáním Prunus spinosa.
Prostřední Špidlák
Lokalita je součástí Evropsky významné lokality Čejkovické Špidláky. Studovaný druh roste v horní třetině severovýchodního svahu Prostředního Špidláku, těsně na okraji akátiny a stepního trávníku, v nadmořské výšce 228 m n. m. První literární údaje o výskytu A. pancicii na Čejčsku pocházejí ze čtyřicátých let 20. století (wenDelberGer 1960 sec. Danihelka 1995). V té době zde druh rostl na dvou lokalitách. První lokalita ležela v širším okolí Špidláku mezi Čejčí a Mutěnicemi a druhá lokalita byla zřejmě na Prostředním Špidláku. Tato lokalita byla v roce 1993 ověřena (Danihelka 1995). Geologický podklad území je tvořen třetihorními vápnitými jíly a písky, z půdních typů na lokalitě převažuje lehčí střední půda typu černozem (bulová 2002). Vegetace území se dá přiřadit ke svazu Bromion erecti Koch 1926. Plocha s výskytem A. pancicii byla dlouhodobě neobhospodařována, zarůstá druhy Bromus erectus, Arrhenatherum elatius, Dactylis glomerata, pod plochou zmlazuje Junglans regia, v blízkosti populace je akátový háj, Robinia pseudacacia zatím do plochy neexpanduje. V roce 2010 byla lokalita poprvé pokosena.
Popis druhu
Artemisia pancicii je vytrvalý druh s tenkými podzemními výběžky, rozrůstající se v roz vol-něné až velmi husté porosty sterilních prýtů 5–10 cm vysokých, s 3–5 přízemními šedobíle až zlatožlutě chlupatými peřenosečnými listy. České a rakouské populace kvetou velmi zřídka, kvetoucí lodyhy jsou (20–)30–90(–95) cm vysoké. Latovité květenství je 8–15 cm vy soké, složené ze zvonkovitých úborů 2–3 mm v průměru, nažky jsou elipsoidní, mír ně smáčknuté, ca 1 mm dlouhé (Grulich 2004). Druh ojediněle kvete od počátku srpna do

14
Gillová et al.: Nové poznatky o Artemisia pancicii
září, zřídka i v říjnu, květy jsou opylovány větrem. Zaznamenáno je pouze vegetativní rozmnožování rozrůstáním podzemních oddenků (holub & Grulich 1999, bulová 2002).
Druh preferuje teplomilné stepní trávníky, těžiště výskytu má v přechodových porostech mezi vegetací svazů Bromion erecti a Festucion valesiacae (Danihelka 1995, holub & Grulich 1999). V Rakousku druh roste také ve vegetaci svazu Geranion sanguinei a ojediněle také ve vegetaci svazu Quercion pubescentis-sessiliflorae (naGler 2010). Druh roste na minerálně bohatých, vápnitých půdách na karbonátových píscích, slínitých jílech až slínech (wenDelberGer 1959, bulová 2002).
Metodika
Monitoring populací, odhad velikostí populací
Pro sledování změn v početnosti populací bylo v roce 2004 na lokalitách Liščí kopec I, na Prostředním Špidláku a na Pouzdřanech vytyčeno sedm trvalých ploch o velikosti 1 × 1 metr. Rozmístění jednotlivých trvalých ploch bylo vybráno tak, aby co nejrepre-zentativněji pokrylo celou plochu populací. Počet ploch byl stanoven vzhledem k celkové ploše jednotlivých populací, tzn. na lokalitě Liščí kopec I tři plochy, na Prostředním Špidláku dvě plochy a na Pouzdřanech dvě plochy. V roce 2007 byla vytyčena jedna trvalá plocha na lokalitách Liščí kopec II a Liščí kopec III. Základní monitorovací jednotkou je nadzemní prýt (tzn. rameta). Pomocí kovového rámu s rastrem 10 × 10 cm jsou na těchto plochách každoročně v době květu (tzn. v září) sledovány počty nadzemních prýtů. Dále jsou na lokalitách dohledávány všechny květonosné lodyhy.
Pro stanovení celkové velikosti populací jsme vycházeli z předpokladu, že sledovaný druh vytváří ± kompaktní polykormony. Na každé lokalitě došlo v roce 2007 k pečlivému prozkoumání místa výskytu druhu. Okrajové body polykormonu byly označeny dřevěnými kolíky a pomocí provázku byla vytyčena celková plocha populace. Velikost plochy v m2 byla vypočítána po přenesení plochy v odpovídajícím měřítku na milimetrový papír. Velikost populací byla spočtena vynásobením průměrného počtu nadzemních prýtů v trvalých čtvercích plochou populace.
Počty nadzemních prýtů v trvalých plochách mezi jednotlivými populacemi byly testovány pomocí analýzy variance, za průkazný výsledek byla považována dosažená hladina významnosti 5 %. Statistické analýzy a grafy byly zpracovány v programu NSCC 2007 (hintze 2007).
Životnost pylových zrn
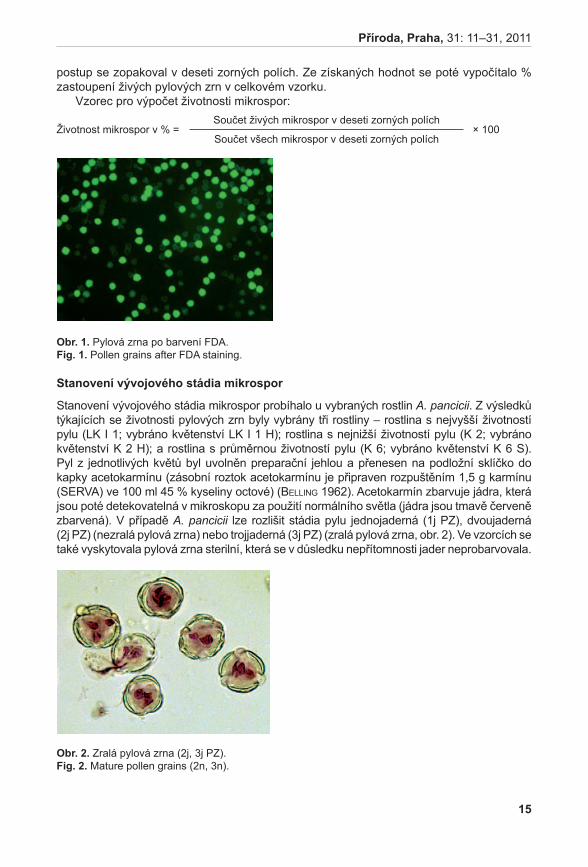
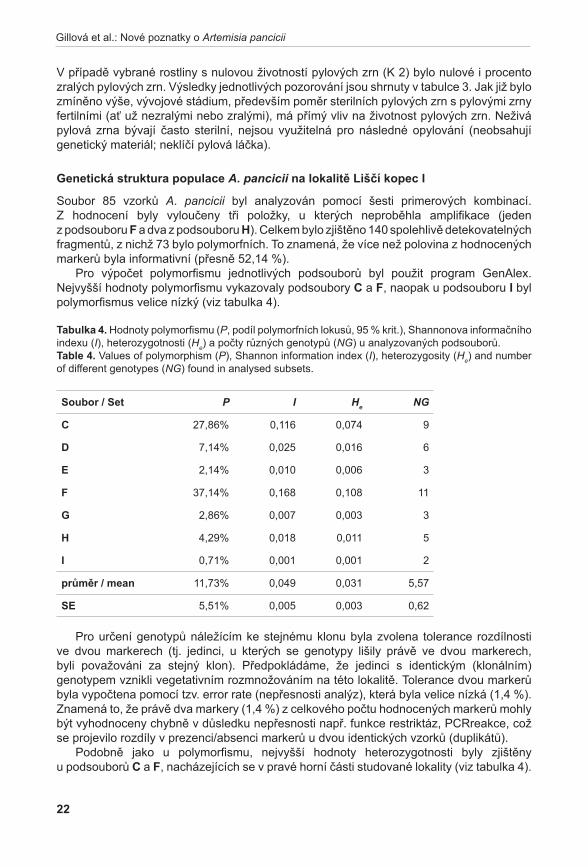
Životnost pylu byla hodnocena u šestnácti rostlin A. pancicii z lokalit Pouzdřany (P 1–3), Liščí kopec I (LK I 1–5) a Liščí kopec III (LK III 1–2) a z rostlin pěstovaných v Troubsku v kultivaci (K 1–6). Z každé rostliny byly odebrány tři květenství – z horní (H), střední (S) a dolní části (D). Pyl z jednotlivých květů byl uvolněn preparační jehlou a přenesen na podložní sklíčko do kapky promývacího roztoku (NLN promývací roztok – osmoticky vyhovující pylovým zrnům, která v čisté vodě praskají, lichter 1981). K rozvolněnému pylu v promývacím roztoku bylo přidáno 20 µl roztoku s fluorescein diacetátem (FDA) (pracovní roztok FDA připraven z 1 ml promývacího roztoku NLN a 20 µl FDA, larkin 1976). Životnost pylových zrn byla hodnocena pomocí fluorescenčního mikroskopu Olympus BX60. V prvním kroku se počítalo množství všech pylových zrn při procházejícím světle, ve druhém kroku se počítala živá pylová zrna při použití BW filtru (fluorescence; viz vzorec). Živá pylová zrna se pomocí FDA obarví a v případě použití BW filtru jasně fluoreskují (viz obr. 1). Tento
15
Příroda, Praha, 31: 11–31, 2011
postup se zopakoval v deseti zorných polích. Ze získaných hodnot se poté vypočítalo % zastoupení živých pylových zrn v celkovém vzorku.
Vzorec pro výpočet životnosti mikrospor:
Životnost mikrospor v % =Součet živých mikrospor v deseti zorných polích
× 100Součet všech mikrospor v deseti zorných polích
obr. 1. Pylová zrna po barvení FDA.Fig. 1. Pollen grains after FDA staining.
stanovení vývojového stádia mikrospor
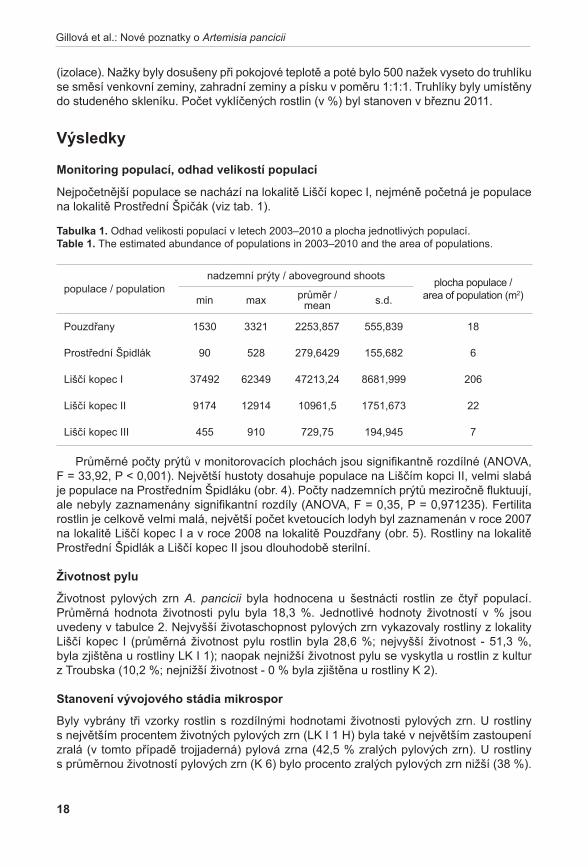
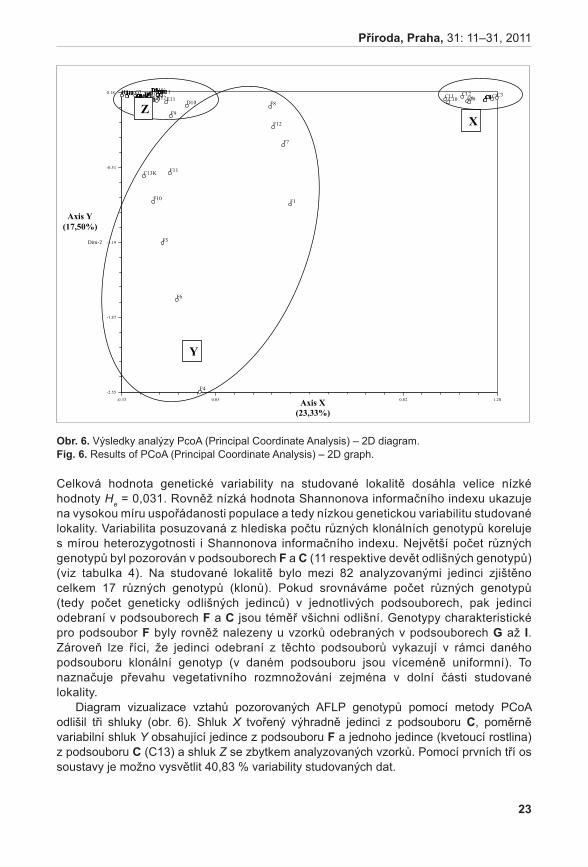
Stanovení vývojového stádia mikrospor probíhalo u vybraných rostlin A. pancicii. Z výsledků týkajících se životnosti pylových zrn byly vybrány tři rostliny – rostlina s nejvyšší životností pylu (LK I 1; vybráno květenství LK I 1 H); rostlina s nejnižší životností pylu (K 2; vybráno květenství K 2 H); a rostlina s průměrnou životností pylu (K 6; vybráno květenství K 6 S). Pyl z jednotlivých květů byl uvolněn preparační jehlou a přenesen na podložní sklíčko do kapky acetokarmínu (zásobní roztok acetokarmínu je připraven rozpuštěním 1,5 g karmínu (SERVA) ve 100 ml 45 % kyseliny octové) (bellinG 1962). Acetokarmín zbarvuje jádra, která jsou poté detekovatelná v mikroskopu za použití normálního světla (jádra jsou tmavě červeně zbarvená). V případě A. pancicii lze rozlišit stádia pylu jednojaderná (1j PZ), dvoujaderná (2j PZ) (nezralá pylová zrna) nebo trojjaderná (3j PZ) (zralá pylová zrna, obr. 2). Ve vzorcích se také vyskytovala pylová zrna sterilní, která se v důsledku nepřítomnosti jader neprobarvovala.
obr. 2. Zralá pylová zrna (2j, 3j PZ).Fig. 2. Mature pollen grains (2n, 3n).

16
Gillová et al.: Nové poznatky o Artemisia pancicii
Genetická struktura populace Artemisia pancicii na lokalitě liščí kopec i
Rostlinný materiálNa lokalitě Liščí kopec I byly nalezeny mezní body, ve kterých se A. pancicii vyskytoval. Tyto body byly spojeny provazem, přičemž vznikl obdélník s délkou stran 17 × 19,5 m. Každá strana vytvořeného obdélníku byla rozdělena na třetiny a ty byly opět spojeny provazem. Tímto způsobem byla lokalita rozdělena na devět podsouborů s velikosti stran 5,7 × 6,5 m a o zna čených písmeny A až i (obr. 3). Pro extrakci DNA byl v každém podsouboru proveden odběr 1–2 listů z devíti náhodně vybraných jedinců. Při odběru vzorků se vybíraly rostliny vzdáleny od sebe nejméně 50 cm, aby se minimalizovala pravděpodobnost sběru vzorků náležícím ke stejnému kormonu. Protože v podsouboru A a B nebyli jedinci A. pancicii nalezeni, byly vzorky doplněny ze zbývajících obdélníků. Celkem bylo odebráno 85 vzorků. Označení podsouborů s příslušným počtem odebraných vzorků je uvedeno na obr. 3.
Poznámka: / Note:GPS souřadnice mezních bodů: / GPS coordinates of the boundary points:1: N 48°50’12,1” E 16°33’49,0” 3: N 48°50’12,0” E 16°33’47,8”2: N 48°50’11,7” E 16°33’48,6” 4: N 48°50’12,5” E 16°33’48,3”
obr. 3. Rozdělení lokality Liščí kopec, označení podsouborů (A – I), počty odebraných vzorků (n).Fig. 3. Division of the Liščí kopec I locality with designation of subdivided parts (A – I) and number of samples taken (n).

17
Příroda, Praha, 31: 11–31, 2011
DNA extrakceExtrakce genomické DNA byla provedena ze 100 mg rostlinného materiálu podle modifikované CTAB metody (kuMP & javornik 1996). Kvalita a integrita DNA byla ověřena na 0,8 % agarózovém gelu. Koncentrace DNA byla zjištěna pomocí UV-spektrofotometru NanoDrop ND - 1000 Spectrophotometer (NanoDrop Technologies).
AFLPProtokol použitý pro provedení metody AFLP na studovaném souboru vzorků byl upraven podle publikace vos et al. (1995) s dílčími modifikacemi dle kitner et al. (2008). Amplifikované fragmenty byly separovány na 6 % denaturačním polyakrylamidovém gelu s následnou vizualizací fragmentů barvením stříbrem. K analýze bylo vybraných šest primerových kombinací. Jejich sekvence současně s příslušnými počty detekovaných fragmentů jsou dostupné u autorů.
Analýza datAFLP fragmenty byly hodnoceny vizuálně, a to pouze bandy spolehlivě rozlišitelné a de-te kovatelné. Na základě takto získaných dat byla v programu MS Excel vytvořena binární matice (1 = přítomnost, 0 = nepřítomnost bandu), která byla dále použita pro vizualizaci příbuznosti analyzovaných položek PCoA analýzou (Principal Coordinate Analysis) pomocí programu NTSYS-pc (Jaccard’s similarity matrix) (rohlf 1998). Pro posouzení genetické diverzity populace byl vypočítán index heterozygotnosti (He = 2 * p * q), Shannonův informační index (I = -1* (p * Ln (p) + q * Ln(q))) a polymorfizmus pomocí programu GenAlex (Peakall & sMouse 2006). Pomocí R-skriptu AFLPdat (ehrich 2006) byl stanoven počet klonů (jedinců nesoucích shodný genotyp, NG – number of genotypes) jednotlivě pro každý podsoubor a dále celkový počet klonů vyskytujících se na lokalitě.
Ekologické nároky druhu
Pro zjištění ekologických nároků druhu byly na všech českých lokalitách zapsány v roce 2008 fytocenologické snímky dle standardní metodiky (cf. Moravec et al. 1994), pokryvnost byla odhadnuta v sedmičlenné Braun-Blanquetově stupnici. Pro jejich zápis byla zvolena jednotná plocha 16 m2. Všechny snímky byly převedeny do fytocenologické databáze v programu TURBOVEG for Windows 2.66 (hennekens & schaMinée 2001) a byly analyzovány v programu Juice 7.0 metodou Twinspan (tichý 2002).
V roce 2007 byly jednorázově odebrány půdní vzorky na analýzy z následujících lokalit: Pouzdřany, Liščí kopec I a z rakouské lokality Neusiedl am See. V roce 2010 byly odebrány a analyzovány půdní vzorky z lokalit: Prostřední Špidlák, Liščí kopec I – část u lesíku, Liščí kopec I – hrana svahu, Liščí kopec II, Liščí kopec III, Pouzdřany – část nad cestou, Pouzdřany – část pod cestou.
Klíčivost semen
Na lokalitě Liščí kopec I byla pomocí pytlíku z mlynářského hedvábí v průběhu července 2010 zaizolována květenství u pěti z celkově osmi kvetoucích rostlin. Podobně byla zakryta květenství u 18 rostlin v kultivaci (z lokalit Neusiedl am See a Liščí kopec III). V průběhu listopadu 2010 byly z pěti plodných rostlin z lokality Liščí kopec I (izolace) a z pěti rostlin z lokality Liščí kopec III (bez izolace) odebrány zralé úbory, a to vždy po třech z horní, střední a spodní části květenství. Podobně byly odebrány úbory i z rostlin pěstovaných v kultivaci, které pocházely z lokalit Neusiedl am See (izolace, bez izolace) a lokality Liščí kopec III
18
Gillová et al.: Nové poznatky o Artemisia pancicii
(izolace). Nažky byly dosušeny při pokojové teplotě a poté bylo 500 nažek vyseto do truhlíku se směsí venkovní zeminy, zahradní zeminy a písku v poměru 1:1:1. Truhlíky byly umístěny do studeného skleníku. Počet vyklíčených rostlin (v %) byl stanoven v březnu 2011.
výsledky
Monitoring populací, odhad velikostí populací
Nejpočetnější populace se nachází na lokalitě Liščí kopec I, nejméně početná je populace na lokalitě Prostřední Špičák (viz tab. 1).
tabulka 1. Odhad velikosti populací v letech 2003–2010 a plocha jednotlivých populací. table 1. The estimated abundance of populations in 2003–2010 and the area of populations.
populace / populationnadzemní prýty / aboveground shoots plocha populace /
area of population (m2)min max průměr / mean s.d.
Pouzdřany 1530 3321 2253,857 555,839 18
Prostřední Špidlák 90 528 279,6429 155,682 6
Liščí kopec I 37492 62349 47213,24 8681,999 206
Liščí kopec II 9174 12914 10961,5 1751,673 22
Liščí kopec III 455 910 729,75 194,945 7
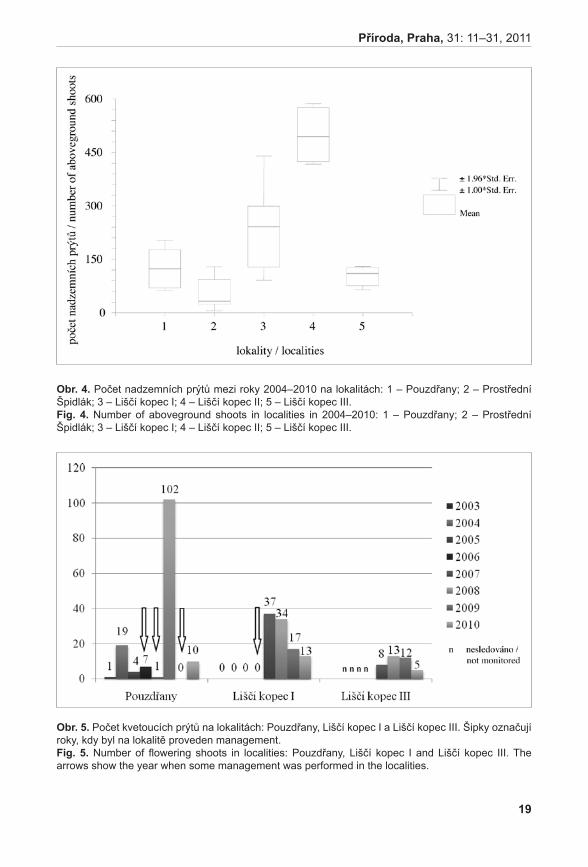
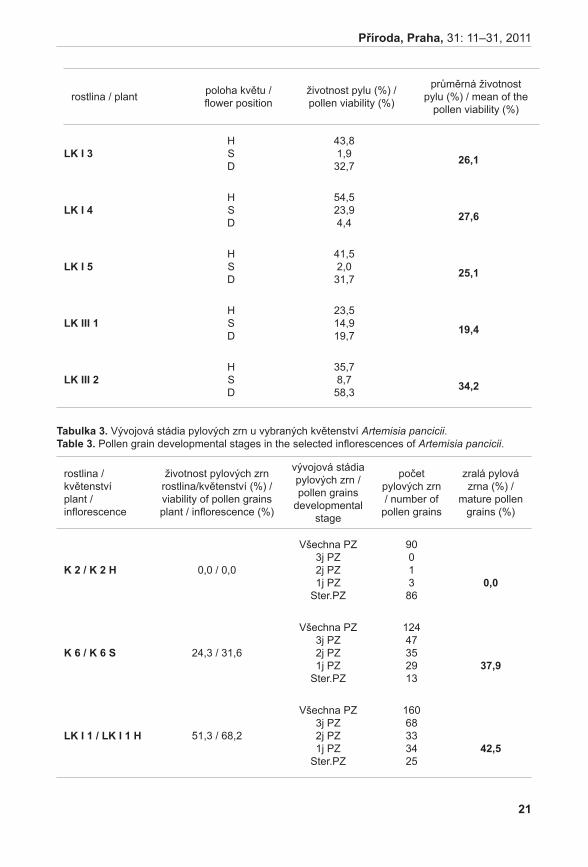
Průměrné počty prýtů v monitorovacích plochách jsou signifikantně rozdílné (ANOVA, F = 33,92, P < 0,001). Největší hustoty dosahuje populace na Liščím kopci II, velmi slabá je populace na Prostředním Špidláku (obr. 4). Počty nadzemních prýtů meziročně fluktuují, ale nebyly zaznamenány signifikantní rozdíly (ANOVA, F = 0,35, P = 0,971235). Fertilita rostlin je celkově velmi malá, největší počet kvetoucích lodyh byl zaznamenán v roce 2007 na lokalitě Liščí kopec I a v roce 2008 na lokalitě Pouzdřany (obr. 5). Rostliny na lokalitě Prostřední Špidlák a Liščí kopec II jsou dlouhodobě sterilní.
Životnost pylu
Životnost pylových zrn A. pancicii byla hodnocena u šestnácti rostlin ze čtyř populací. Průměrná hodnota životnosti pylu byla 18,3 %. Jednotlivé hodnoty životností v % jsou uvedeny v tabulce 2. Nejvyšší životaschopnost pylových zrn vykazovaly rostliny z lokality Liščí kopec I (průměrná životnost pylu rostlin byla 28,6 %; nejvyšší životnost - 51,3 %, byla zjištěna u rostliny LK I 1); naopak nejnižší životnost pylu se vyskytla u rostlin z kultur z Troubska (10,2 %; nejnižší životnost - 0 % byla zjištěna u rostliny K 2).
stanovení vývojového stádia mikrospor
Byly vybrány tři vzorky rostlin s rozdílnými hodnotami životnosti pylových zrn. U rostliny s největším procentem životných pylových zrn (LK I 1 H) byla také v největším zastoupení zralá (v tomto případě trojjaderná) pylová zrna (42,5 % zralých pylových zrn). U rostliny s průměrnou životností pylových zrn (K 6) bylo procento zralých pylových zrn nižší (38 %).
19
Příroda, Praha, 31: 11–31, 2011
obr. 4. Počet nadzemních prýtů mezi roky 2004–2010 na lokalitách: 1 – Pouzdřany; 2 – Prostřední Špidlák; 3 – Liščí kopec I; 4 – Liščí kopec II; 5 – Liščí kopec III.Fig. 4. Number of aboveground shoots in localities in 2004–2010: 1 – Pouzdřany; 2 – Prostřední Špidlák; 3 – Liščí kopec I; 4 – Liščí kopec II; 5 – Liščí kopec III.
obr. 5. Počet kvetoucích prýtů na lokalitách: Pouzdřany, Liščí kopec I a Liščí kopec III. Šipky označují roky, kdy byl na lokalitě proveden management.Fig. 5. Number of flowering shoots in localities: Pouzdřany, Liščí kopec I and Liščí kopec III. The arrows show the year when some management was performed in the localities.

20
Gillová et al.: Nové poznatky o Artemisia pancicii
tabulka 2. Životnost pylu (%) u vybraných rostlin Artemisia pancicii.table 2. Pollen viability (%) in selected plants of Artemisia pancicii.
rostlina / plant poloha květu / flower position
životnost pylu (%) / pollen viability (%)
průměrná životnost pylu (%) / mean of the
pollen viability (%)
K 1HSD
11,10,00,7 3,9
K 2HSD
0,00,00,0 0,0
K 3HSD
2,90,01,7 1,5
K 4HSD
0,00,02,8 0,9
K 5HSD
6,328,356,7 30,4
K 6HSD
4,531,636,7 24,3
P 1HSD
29,44,31,4 11,7
P 2HSD
1,00,01,4 0,8
P 3HSD
53,90,014,2 22,7
lK i 1HSD
68,261,524,2 51,3
lK i 2HSD
1,00,037,0 12,7
21
Příroda, Praha, 31: 11–31, 2011
rostlina / plant poloha květu / flower position
životnost pylu (%) / pollen viability (%)
průměrná životnost pylu (%) / mean of the
pollen viability (%)
lK i 3HSD
43,81,932,7 26,1
lK i 4HSD
54,523,94,4 27,6
lK i 5HSD
41,52,031,7 25,1
lK iii 1 HSD
23,514,919,7 19,4
lK iii 2 HSD
35,78,758,3 34,2
tabulka 3. Vývojová stádia pylových zrn u vybraných květenství Artemisia pancicii.table 3. Pollen grain developmental stages in the selected inflorescences of Artemisia pancicii.
rostlina / květenstvíplant / inflorescence
životnost pylových zrnrostlina/květenství (%) / viability of pollen grainsplant / inflorescence (%)
vývojová stádia pylových zrn / pollen grains
developmental stage
počet pylových zrn / number of
pollen grains
zralá pylová zrna (%) /
mature pollen grains (%)
K 2 / K 2 H 0,0 / 0,0
Všechna PZ 3j PZ2j PZ 1j PZ
Ster.PZ
9001386
0,0
K 6 / K 6 s 24,3 / 31,6
Všechna PZ 3j PZ2j PZ 1j PZ
Ster.PZ
12447352913
37,9
lK i 1 / lK i 1 H 51,3 / 68,2
Všechna PZ 3j PZ2j PZ 1j PZ
Ster.PZ
16068333425
42,5
22
Gillová et al.: Nové poznatky o Artemisia pancicii
V případě vybrané rostliny s nulovou životností pylových zrn (K 2) bylo nulové i procento zralých pylových zrn. Výsledky jednotlivých pozorování jsou shrnuty v tabulce 3. Jak již bylo zmíněno výše, vývojové stádium, především poměr sterilních pylových zrn s pylovými zrny fertilními (ať už nezralými nebo zralými), má přímý vliv na životnost pylových zrn. Neživá pylová zrna bývají často sterilní, nejsou využitelná pro následné opylování (neobsahují genetický materiál; neklíčí pylová láčka).
Genetická struktura populace A. pancicii na lokalitě liščí kopec i
Soubor 85 vzorků A. pancicii byl analyzován pomocí šesti primerových kombinací. Z hodnocení byly vyloučeny tři položky, u kterých neproběhla amplifikace (jeden z podsouboru F a dva z podsouboru H). Celkem bylo zjištěno 140 spolehlivě detekovatelných fragmentů, z nichž 73 bylo polymorfních. To znamená, že více než polovina z hodnocených markerů byla informativní (přesně 52,14 %).
Pro výpočet polymorfismu jednotlivých podsouborů byl použit program GenAlex. Nejvyšší hodnoty polymorfismu vykazovaly podsoubory c a F, naopak u podsouboru i byl polymorfismus velice nízký (viz tabulka 4).
tabulka 4. Hodnoty polymorfismu (P, podíl polymorfních lokusů, 95 % krit.), Shannonova informačního indexu (I), heterozygotnosti (He) a počty různých genotypů (NG) u analyzovaných podsouborů. table 4. Values of polymorphism (P), Shannon information index (I), heterozygosity (He) and number of different genotypes (NG) found in analysed subsets.
soubor / set P I He NG
c 27,86% 0,116 0,074 9
d 7,14% 0,025 0,016 6
E 2,14% 0,010 0,006 3
F 37,14% 0,168 0,108 11
G 2,86% 0,007 0,003 3
H 4,29% 0,018 0,011 5
i 0,71% 0,001 0,001 2
průměr / mean 11,73% 0,049 0,031 5,57
sE 5,51% 0,005 0,003 0,62
Pro určení genotypů náležícím ke stejnému klonu byla zvolena tolerance rozdílnosti ve dvou markerech (tj. jedinci, u kterých se genotypy lišily právě ve dvou markerech, byli považováni za stejný klon). Předpokládáme, že jedinci s identickým (klonálním) genotypem vznikli vegetativním rozmnožováním na této lokalitě. Tolerance dvou markerů byla vypočtena pomocí tzv. error rate (nepřesnosti analýz), která byla velice nízká (1,4 %). Znamená to, že právě dva markery (1,4 %) z celkového počtu hodnocených markerů mohly být vyhodnoceny chybně v důsledku nepřesnosti např. funkce restriktáz, PCRreakce, což se projevilo rozdíly v prezenci/absenci markerů u dvou identických vzorků (duplikátů).
Podobně jako u polymorfismu, nejvyšší hodnoty heterozygotnosti byly zjištěny u podsouborů c a F, nacházejících se v pravé horní části studované lokality (viz tabulka 4).
23
Příroda, Praha, 31: 11–31, 2011
Dim-1-0.33 0.05 0.44 0.82 1.20
Dim-2
-2.55
-1.87
-1.19
-0.51
0.18 C1C2 C3C4C5C6 C7C8C9C10C11 C12
C13K
D1 D2D3D4D5D6D7D8D9
D10
D11D12E1E2
E3E4E5
E6E7E8
E9E10E11
E12
F1
F3
F4
F5
F6
F7
F8
F9
F10
F11
F12
G1G2G3G4G5G6G7G8G9G10G11G12H1H2H4H5H6H7H9H10H11 H12I1I2I3I4I5I6I7I8I9I10I11I12
Obr. 6. Výsledky analýzy PcoA (Principal Coordinate Analysis) – 2D diagram.
Fig. 6. Results of PCoA (Principal Coordinate Analysis) - 2D graph.
X
Y
Z
Axis Y (17,50%)
Axis X (23,33%)
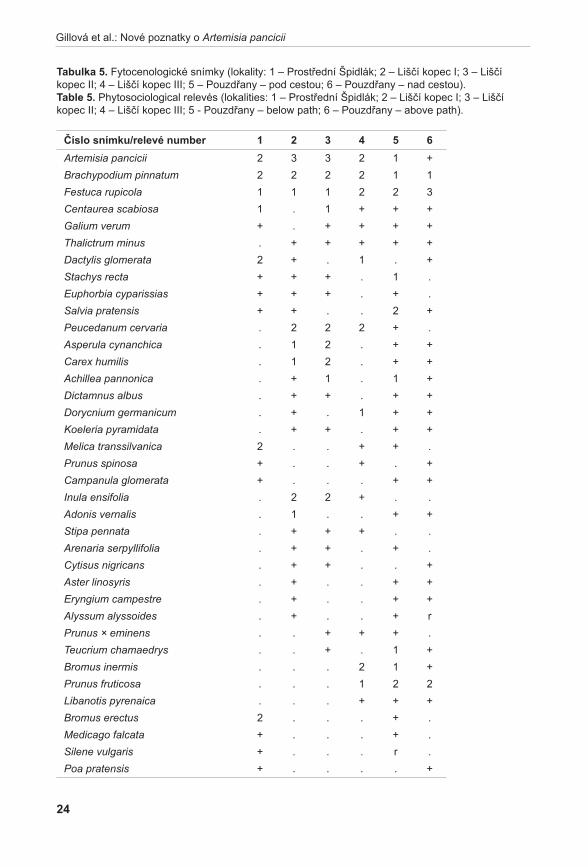
obr. 6. Výsledky analýzy PcoA (Principal Coordinate Analysis) – 2D diagram. Fig. 6. Results of PCoA (Principal Coordinate Analysis) – 2D graph.
Celková hodnota genetické variability na studované lokalitě dosáhla velice nízké hodnoty He = 0,031. Rovněž nízká hodnota Shannonova informačního indexu ukazuje na vysokou míru uspořádanosti populace a tedy nízkou genetickou variabilitu studované lokality. Variabilita posuzovaná z hlediska počtu různých klonálních genotypů koreluje s mírou heterozygotnosti i Shannonova informačního indexu. Největší počet různých genotypů byl pozorován v podsouborech F a c (11 respektive devět odlišných genotypů) (viz tabulka 4). Na studované lokalitě bylo mezi 82 analyzovanými jedinci zjištěno celkem 17 různých genotypů (klonů). Pokud srovnáváme počet různých genotypů (tedy počet geneticky odlišných jedinců) v jednotlivých podsouborech, pak jedinci odebraní v podsouborech F a c jsou téměř všichni odlišní. Genotypy charakteristické pro podsoubor F byly rovněž nalezeny u vzorků odebraných v podsouborech G až i. Zároveň lze říci, že jedinci odebraní z těchto podsouborů vykazují v rámci daného podsouboru klonální genotyp (v daném podsouboru jsou víceméně uniformní). To naznačuje převahu vegetativního rozmnožování zejména v dolní části studované lokality.
Diagram vizualizace vztahů pozorovaných AFLP genotypů pomocí metody PCoA odlišil tři shluky (obr. 6). Shluk X tvořený výhradně jedinci z podsouboru c, poměrně variabilní shluk Y obsahující jedince z podsouboru F a jednoho jedince (kvetoucí rostlina) z podsouboru c (C13) a shluk Z se zbytkem analyzovaných vzorků. Pomocí prvních tří os soustavy je možno vysvětlit 40,83 % variability studovaných dat.
24
Gillová et al.: Nové poznatky o Artemisia pancicii
tabulka 5. Fytocenologické snímky (lokality: 1 – Prostřední Špidlák; 2 – Liščí kopec I; 3 – Liščí kopec II; 4 – Liščí kopec III; 5 – Pouzdřany – pod cestou; 6 – Pouzdřany – nad cestou).table 5. Phytosociological relevés (lokalities: 1 – Prostřední Špidlák; 2 – Liščí kopec I; 3 – Liščí kopec II; 4 – Liščí kopec III; 5 - Pouzdřany – below path; 6 – Pouzdřany – above path).
Číslo snímku/relevé number 1 2 3 4 5 6Artemisia pancicii 2 3 3 2 1 +Brachypodium pinnatum 2 2 2 2 1 1Festuca rupicola 1 1 1 2 2 3Centaurea scabiosa 1 . 1 + + +Galium verum + . + + + +Thalictrum minus . + + + + +Dactylis glomerata 2 + . 1 . +Stachys recta + + + . 1 .Euphorbia cyparissias + + + . + .Salvia pratensis + + . . 2 +Peucedanum cervaria . 2 2 2 + .Asperula cynanchica . 1 2 . + +Carex humilis . 1 2 . + +Achillea pannonica . + 1 . 1 +Dictamnus albus . + + . + +Dorycnium germanicum . + . 1 + +Koeleria pyramidata . + + . + +Melica transsilvanica 2 . . + + .Prunus spinosa + . . + . +Campanula glomerata + . . . + +Inula ensifolia . 2 2 + . .Adonis vernalis . 1 . . + +Stipa pennata . + + + . .Arenaria serpyllifolia . + + . + .Cytisus nigricans . + + . . +Aster linosyris . + . . + +Eryngium campestre . + . . + +Alyssum alyssoides . + . . + rPrunus × eminens . . + + + .Teucrium chamaedrys . . + . 1 +Bromus inermis . . . 2 1 +Prunus fruticosa . . . 1 2 2Libanotis pyrenaica . . . + + +Bromus erectus 2 . . . + .Medicago falcata + . . . + .Silene vulgaris + . . . r .Poa pratensis + . . . . +

25
Příroda, Praha, 31: 11–31, 2011
Číslo snímku/relevé number 1 2 3 4 5 6Galium aparine + . . . . rAnthericum ramosum . 2 2 . . .Inula hirta . 2 + . . .Poa angustifolia . 1 . + . .Senecio jacobaea . + + . . .Pyrethrum corymbosum . + + . . .Thymus pannonicus . + + . . .Bupleurum falcatum . + . + . .Genista tinctoria . + . + . .Pseudolysimachion spicatum . + . . + .Veronica prostrata . + . . + .Centaurea stoebe . + . . + .Astragalus onobrychis . + . . + .Festuca valesiaca . + . . . 1Melampyrum arvense . . 1 r . .Thlaspi perfoliatum . . + + . .Galium glaucum . . + . + .Oxytropis pilosa . . + . + .Potentilla arenaria . . + . + .Onobrychis viciifolia . . + . . +Falcaria vulgaris . . . + + .Artemisia pontica . . . + . +Erysimum diffusum . . . r r .Stipa pulcherrima . . . . 2 1Elytrigia intermedia . . . . 1 +Cytisus procumbens . . . . + +Vincetoxicum hirundinaria . . . . + +Securigera varia . . . . r +
druhy přítomné v jednom snímku / species occurring in one relevé:Arrhenatherum elatius 1: 3; Fragaria moschata 1: 1; Filipendula ulmaria 1: 1; Vicia cracca 1: +; Festuca rubra agg. 1: +; Lotus corniculatus 1: +; Juglans regia juv. 1: +; Hypericum elegans 1: +; Geum urbanum 1: +; Campanula moravica 1: +; Centaurea jacea 1: +; Primula veris 1: +; Ranunculus polyanthemos 1: +; Rumex acetosa 1: +; Galium boreale 1: +; Achillea millefolium agg. 1: +; Anthoxanthum odoratum 1: +; Arabis hirsuta agg. 1: +; Trifolium montanum 1: r; Pimpinella saxifraga 1: r; Rosa canina agg. juv. 1: r; Geranium sanguineum 2: 1; Dianthus pontederae 2: +; Crepis praemorsa 2: +; Orthantha lutea 2: r; Scabiosa canescens 2: r; Crambe tataria 2: r; Chamaecytisus virescens 3: +; Arabis hirsuta 3: r; Calamagrostis epigejos 4: 1; Vicia tenuifolia 4: +; Viola hirta 4: +; Fragaria vesca 4: +; Verbascum austriacum 4: +; Lotus corniculatus 4: +; Euphorbia virgata 4: +; Salvia verticillata 4: +; Acinos arvensis 4: r; Rhamnus cathartica juv. 4: r; Artemisia campestris 5: +; Allium vineale 5: +; Potentilla recta 5: +; Viola odorata 5: +; Scabiosa ochroleuca 5: +; Iris pumila 5: +; Koeleria macrantha 5: +; Origanum vulgare 5: +; Hypericum perforatum 5: +; Camelina microcarpa 5: r; Campanula sibirica 5: r; Nonea pulla 5: r; Reseda lutea 5: r; Plantago media 6: +; Pyrus pyraster juv. 6: +; Avenula pratensis 6: +; Phleum phleoides 6: +; Trifolium alpestre 6: r; Quercus pubescens juv. 6: r.
26
Gillová et al.: Nové poznatky o Artemisia pancicii
tabulka 6. Výsledky chemických rozborů půdy z lokalit: 1 - Pouzdřany 2008; 2 - Liščí kopec I 2008; 3 - Neusiedl am See 2008; 4 - Čejkovice 2010; 5 - Liščí kopec I - u lesíka 2010; 6 - Liščí kopec II 2010; 7 - Liščí kopec III 2010; 8 - Pouzdřany - pod cestou 2010; 9 - Pouzdřany - nad cestou 2010; 10 - Liščí kopec I – hrana svahu 2010.table 6. The results of soil chemical analyses from localities: 1 - Pouzdřany 2008; 2 - Liščí kopec I 2008; 3 - Neusiedl am See 2008; 4 - Čejkovice 2010; 5 - Liščí kopec I – by a small wood 2010; 6 - Liščí kopec II 2010; 7 - Liščí kopec III 2010; 8 - Pouzdřany - below path 2010; 9 - Pouzdřany - above path 2010; 10 - Liščí kopec I - edge of slope 2010.
lokalita / locality 1 2 3 4 5 6 7 8 9 10
pH/KCl 7 6,8 6,5 7,2 7,2 7,1 7,1 7,4 7,1 7,4
P (mg/kg) 7 9 13 16 8 5 11 5 5 3
K (mg/kg) 295 308 185 169 284 162 589 280 365 102
Mg (mg/kg) 236 266 385 373 302 245 275 239 268 200
Ca (mg/kg) 7376 9794 3688 6456 6958 10383 7583 9498 7495 13870
Humus (%) 4,75 7,53 10,15 6,32 7,21 7,02 8,09 4,52 8,41 2,25
Nc (%) 0,625 0,495 0,545 0,51 0,59 0,34 0,66 0,31 0,51 0,24
Ekologické nároky druhu
Pro pochopení ekologických vazeb druhu byly zapsány fytocenologické snímky na lo ka-litách s výskytem tohoto druhu. Publikované snímky (viz tab. 5) pocházejí z roku 2008.
Po provedení statistických analýz programem TWINSPAN se jako signifikantně odlišný oddělil snímek č. 1 z lokality Prostřední Špidlák. Nicméně jeho odlišnost je jen velmi malá. Na základě výčtu typických druhů (Artemisia pancicii, Brachypodium pinnatum, Festuca rupicola, Centaurea scabiosa, Galium verum, Thalictrum minus, Dactylis glomerata, Stachys recta, Euphorbia cyparissias, Salvia pratensis, Peucedanum cervaria, Asperula cynanchica, Carex humilis, Achillea pannonica, Dictamnus albus, Dorycnium germanium) se jedná v případě všech lokalit o typickou vegetaci svazu Bromion erecti Koch 1926 s vysokým podílem typických panonských druhů – v našich snímcích: Artemisia pancicii, Dorycnium germanicum, Inula ensifolia, Erysimum diffusum, Hypericum elegans, Dianthus pontederae, Crambe tataria, Chamaecytisus virescens, Iris pumila, Campanula sibirica aj.
Výsledky půdních rozborů jsou uvedeny v tab. 6. Všechny analyzované vzorky patří do skupiny černozemních půd. Z výsledků analýz vyplývá, že se jedná o půdy s neutrální až alkalickou půdní reakcí, obsah draslíku je dostatečný až velmi vysoký, obsah hořčíku dobrý až vysoký, obsah vápníku vysoký, obsah dusíkatých látek střední až dobrý a obsah humusu vysoký. Mezi jednotlivými vzorky nebyly zaznamenány zásadní rozdíly. Limitací pro generativní reprodukci může být velmi nízký obsah fosforu.
Klíčivost semen
Klíčivost semen byla velmi nízká, celkově dosahovala pouze 0,72 %. Vyklíčily nažky ze všech lokalit, a to jak z rostlin izolovaných, tak i bez izolace (viz tab. 7). Nejvíce semenáčků bylo zjištěno u izolovaných rostlin pěstovaných v kultivaci z lokality Liščí kopec III (devět

27
Příroda, Praha, 31: 11–31, 2011
semenáčků). Pouze jeden semenáček vyklíčil ze semen sebraných z izolovaných rostlin z lokality Neusiedl am See pěstovaných v kultivaci a z neizolovaných rostlin z lokality Liščí kopec III.
tabulka 7. Klíčivost semen (%) z lokalit: 1 - Liščí kopec I (izolace); 2 - Liščí kopec III (bez izolace); 3 – Liščí kopec III (izolace, kultivace), 4 - Neusiedl am See (bez izolace, kultivace); 5 - Neusiedl am See (izolace, kultivace).table 7. Germination (%) of seeds from localities: 1 - Liščí kopec I (isolation); 2 - Liščí kopec III (without isolation); 3 – Liščí kopec III (isolation, cultivation); 4 - Neusiedl am See (without isolation, cultivation); 5 - Neusiedl am See (isolation, cultivation).
lokalita / locality 1 2 3 4 5
klíčivost (%) / germination (%) 0,6 0,2 1,8 0,8 0,2
diskuse
Monitoring populací, odhad velikostí populací
Rozdíly ve velikostech populací jsou s největší pravděpodobností způsobeny zachovalostí biotopu na lokalitách. Na lokalitách Liščí kopec I a II a Pouzdřany se provádí nepravidelný management, naproti tomu nejmenší populace na Prostředním Špidláku a Liščím kopci III byly dlouhodobě bez údržby. Lokalita Prostřední Špidlák byla posečena až v roce 2010. Obě dvě lokality zarůstají expanzními travinami (Arrhenatherum elatius, Bromus erectus, Bromus inermis, Calamagrostis epigejos, Dactylis glomerata) a hromadí se zde stařina.
Podobně i meziroční fluktuace v počtu sterilních a kvetoucích prýtů jsou pravděpodobně výsledky prováděných managementových opatření. Nejvíce kvetoucích lodyh bylo zaznamenáno na lokalitě Liščí kopec I a Pouzdřany, a to rok po pokosení a pastvě lokalit – viz obr. 5.
Artemisia pancicii preferuje teplomilné stepní trávníky, těžiště výskytu má v přechodových porostech mezi vegetací svazů Bromion erecti a Festucion valesiacae (Danihelka 1995, holub & Grulich 1999, Grulich in slavík & ŠtěPánková 2004). To jsou biotopy, jejichž druhová bohatost je závislá na víceméně pravidelném managementu (chytrý 2007). Pravidelné sečení i vypalování má pozitivní vliv na druhovou bohatost společenstev svazu Festucion valesiaceae (MünzberGová 2001). bulová (2002) ve své diplomové práci experimentálně testovala vliv sečení na počet nadzemních prýtů. Na lokalitě Prostřední Špidlák byl počet nadzemních prýtů v posečené ploše vyšší než v roce, kdy plocha nebyla posečena. Na lokalitě Liščí kopec pozitivní efekt posečení nebyl prokázán. Jednalo se ale o jednoroční výzkum a design experimentu nebyl vybrán standardně, výsledky tedy nejsou příliš průkazné. Pozitivní efekt vypalování byl pozorován po roce 1988, kdy vyhořela Pouzdřanská step a kdy v následujících letech došlo k velkému nárůstu kvetoucích prýtů (V. Grulich, úst. sdělení).
Celkové velikosti populací jsou pouze hrubým odhadem. Artemisia pancicii vytváří na většině lokalit kompaktní polykormony, přesné sečtení nadzemních prýtů by bylo velmi pracné. Daná metodika byla zvolena především pro sběr dat pro hodnotící zprávy o stavu evropsky významných lokalit. Přesné počty prýtů vypovídající o trendu vývoje populací jsou sledovány v rámci každoročního monitoringu v trvalých plochách.

28
Gillová et al.: Nové poznatky o Artemisia pancicii
Životnost pylu, stanovení vývojového stádia mikrospor a klíčivost semen
Životnost pylu je ovlivňována řadou vnějších i vnitřních faktorů. Nejdůležitějším vnějším faktorem jsou aktuální teplotní a především vlhkostní podmínky. Vlhkost a nižší teploty mají na pyl, jeho životnost, a následně schopnost oplození (prorůstání pylové láčky) negativní vliv. Co se týče vnitřních faktorů ovlivňujících životaschopnost pylu, důležitý je celkový stav rostliny (napadení rostliny chorobami, parazity, herbivory) a zralost pylu. U odebraného pylu byla prokázána ve většině případů alespoň částečná životnost. Životnost pylu závisela na celkové stavu rostlin a především na aktuálních vnějších podmínkách při sběru pylu. V případě lokalit Pouzdřany a kultur z Troubska bylo pro sběr pylu nevhodné počasí – deštivo; což se objektivně projevilo na radikálně snížené životnosti pylu, v některých případech i nulové. Co se týče stanovení vývojového stádia pylu, výsledky úzce souvisely se stanovenou životností pylu. V několika případech byla detekována sterilní pylová zrna, což je běžný jev: stejně jako na životnost pylu má i na jeho sterilitu vliv řada vnitřních i vnějších faktorů (sawhney & shukla 1994). Tato pylová zrna nejsou životná a v důsledku toho nejsou schopna opylení a následného oplození. Většina pylových zrn však byla fertilní (zralá – v případě A. pancicii trojjaderná, nezralá – jedno, či dvoujaderná). V tomto případě jsou trojjaderná pylová zrna schopna opylení, oplození a následné tvorby embrya. Problémem u A. pancicii na sledovaných lokalitách je častá absence tvorby vyvinutých nažek. Doposud bylo zaznamenáno pouze vegetativní rozmnožování rozrůstáním podzemních oddenků (holub & Grulich 1999). bulová (2002) ve své diplomové práci testovala klíčivost semen s nulovou úspěšností. Námi zaznamenaná klíčivost semen potvrzuje možnost generativní reprodukce. Artemisia pancicii kvete velmi zřídka, k tomu patří k dlouhodenním rostlinám, které kvetou v pozdním létě a na podzim. K dozrání nažek dochází pouze v případě dlouhé vegetační sezóny. Generativní reprodukce je tedy velmi limitována, což potvrzují i výsledky studii genetické variability českých a rakouských populací (kitner et al., in prep.). Životaschopná semena byla zjištěna i u rostlin, které byly zaizolovány, což potvrzuje limitovanou možnost autogamie u pelyňků (Parihar et al. 2009).
Genetická struktura populace A. pancicii na lokalitě liščí kopec i
Celková úroveň genetické variability v populaci A. pancicii na lokalitě Liščí kopec I je nízká (He = 0,031), což pravděpodobně souvisí s problémy druhu se sexuálním rozmnožováním a tvorbou nažek. Podobně nízké hodnoty byly zjištěny u ohroženého druhu Astragalus cremnophylax var. cremnophylax (He = 0,020–0,130; travis et al. 1996), zatímco střední hodnoty heterozygotnosti u populací Silene tatarica (He = 0,127; tero et al. 2003), Rumex rothschildianus (He = 0,170; rottenberG & Parker 2003), Eryngium alpinum (He = 0,198; GauDel et al. 2001). Jak bylo uvedeno výše, celkem bylo zjištěno 17 různých klonů, a pokud hodnotíme počet různých genotypů, pak v podsouborech F a c jsou téměř všichni jedinci odlišní. Ve zbytku studovaných podsouborů se mezi sesbíranými jedinci nachází většina jedinců s klonálním genotypem, což naznačuje převahu vegetativního rozmnožování v dolní části lokality. Jedinci z podsouboru c vytvářejí samostatný diskrétní shluk (kromě jedince c13K shlukujícího se s více variabilními vzorky s podsouboru F), vykazující vyšší hodnoty indexů genetické variability. V 2D grafu z PCoA analýzy jsou patrné tři zřetelně oddělené shluky (viz obr. 6). Dva výrazněji se vyčleňující shluky s vyšší mírou variability jsou lokalizovány v pravé horní části lokality (při pohledu shora) – podsoubory c a F. V této části lokality jsme pozorovali častější výskyt kvetoucích rostlin. Pravděpodobně tedy v této části lokality existují lepší podmínky pro průběh sexuálního rozmnožování, a tudíž tato část lokality může být zdrojem variability populace. Důvodem může být mikrolokální

29
Příroda, Praha, 31: 11–31, 2011
zvýhodnění díky blízkosti akátového lesíku, který poskytuje částečné zastínění právě v místech podsouborů c a F. V těchto místech je patrně i vyšší vlhkost. Zde tedy existuje vhodnější mikroklima pro kvetení, opylování a popř. tvorbu semen. Přítomnost jedinců z podsouboru F ve všech třech shlucích v PCoA grafu ukazuje přítomnost „F“ genotypu na celé lokalitě.
Je nutné si uvědomit rozdíl ve schopnosti druhu přežít v současných podmínkách při zjištěné nízké genetické variabilitě populace a výhledově v „evolučním potenciálu“ druhu. Vyšší hodnoty genetické variability vypovídají o přítomnosti různých alel, které mohou umožnit adaptaci druhu na změnu klimatu či výskyt nových patogenů (beebee & rowe 2005), a tedy přežití druhu v dlouhodobém měřítku. Výsledky této části práce navazují na studii genetické variability českých a rakouských populací A. pancicii, které jsou v současné době připravovány k publikování (kitner et al., in prep.). V této práci byla zjištěna vysoká úroveň genetické variability mezi populacemi a vysoký stupeň genetické homogenity uvnitř populací, s omezeným tokem alel mezi populacemi (gene flow) a s převažujícím vegetativním způsobem rozmnožování. U lokality Liščí kopec I byly zjištěny druhé nejvyšší hodnoty indexů genetické variability (He = 0,142) po rakouské lokalitě Nickelsdorf (kitner et al., in prep.). Z výsledků obou studií lze usuzovat, že na lokalitě Liščí kopec I k pohlavnímu rozmnožování v (sub)recentní době docházelo a v případě příznivých podmínek i nadále dochází. Při stávajícím managementu lze předpokládat úspěšné přežití druhu na lokalitě.
summary
The population sizes vary a lot – the largest is the Liščí kopec I population, while the Prostřední Špidlák population is very weak. The plant fertility is very low; mowing in the Liščí kopec I locality in 2006 and grazing in the Pouzdřany locality in 2006 and 2007 had a positive effect on the number of flowering rametes. The plants in the Prostřední Špidlák and Liščí kopec II localities remained sterile for a long time. The viability of pollen grains and pollen development stages were tested on sixteen plants. Viable pollen grains appeared in the tested samples (maximum mean viability was 51.3%) and there were also unviable pollen grains. The pollen development stages were closely related to viability – sterile pollen grains were unviable, and fertile pollen grains (one to three nucleuses) were mostly viable. Only 0.72% of the seeds germinated. The possibility of limited autogamy was confirmed. Despite the generally low level of heterozygosity of the population (the area is quite small), it was possible to identify seventeen different genotypes among 82 plants analysed by the AFLP method. More variable individuals are localised in the upper and more shaded part of the locality.
Poděkování
Výsledků bylo dosaženo v rámci řešení následujících projektů a výzkumných záměrů: MŠMT VaV 2B06178, MSM 6198959215, MSM 2629608001, IGA Prf-2010-001. Rádi bychom také poděkovali Simonovi Gillovi za korekturu anglických textů a všem kolegům, kteří nám pomáhali při náročné práci v terénu.

30
Gillová et al.: Nové poznatky o Artemisia pancicii
literatura
beebee t. j. c. & rowe G. (2005): An introduction to molecular ecology. – Oxford University press, New York, USA.
bellinG j. (1962): The iron-acetocarmin method of fixing and staining chromosomes. – Biol. Bull. 50: 160–162.
boža P. (1999): Artemisia pancicii (Janka) Ronniger subsp. pancicii. – In: stevanović v. [ed.], Crvena knjiga flore Srbije 1., pp. 141–142, Ministarstvo za životnu sredinu R Srbije, Biološki fakultet Univ. u Beogradu i Zavod za zaštitu prirode R Srbije, Beograd.
bulová t. (2002): Biologie, ekologie a rozšíření Cardamine parviflora L., Artemisia pancicii (Janka) Ronn. a Gladiolus palustris Gaud. na Moravě. – Ms. [Dipl. pr.; depon. in: Knih. Přírod. Fak. MU, Brno.]
Danihelka j. (1995): O druhu Artemisia pancicii v České republice a příbuzných taxonech. – Zpr. Čes. Bot. Společ. 30: 81–90.
Danihelka j. & MarholD k. (2003): Validation of the name Artemisia pancicii (Asteraceae). – Willdenowia 33: 251–254.
ehrich D. (2006): AFLPDAT: a collection of R functions for convenient handling of AFLP data. – Mol. Ecol. Notes 6: 603–604.
GauDeul M., taberlet P. & till-bottrauD i. (2001): Genetic diversity in an endangered alpine plant, Eryngium alpinum L. (Apiaceae), inferred from amplified fragment length polymorphism markers. – Mol. Ecol. 9: 1625–1637.
Grulich v. (1987): Kriticky ohrožené druhy rostlin ČSR. Pelyněk Pančičův. Artemisia pancicii (Janka) Ronn. – Památ. a Přír. 12/5: 3. str. obálky.
Grulich v. (2004): Artemisia pancicii Danihelka et Marhold. – In: slavík b. & ŠtěPánková j. [eds], Květena České republiky 7, p. 172, Academia, Praha.
hennekens s. M. & schaMinée j. h. j. (2001): TURBOVEG, a comprehensive database managment systém for vegetation data. – J. Veg. Sci. 12: 589–591.
hintze j. l. (2007): NCSS Quick start & Self help manual. – NCSS, Kaysville.holub j. & Grulich v. (1999): Artemisia pancicii (Janka) Ronn. – In: Čeřovský j., feráková v., holub j.,
MaGlocký Š. & Procházka f. [eds], Červená kniha ohrožených a vzácných druhů rostlin a živočichů ČR a SR. Vol. 5. Vyšší rostliny, pp. 43, Příroda, Bratislava.
chytrý M. [ed.] (2007): Vegetace České republiky 1. Travinná a keříčková vegetace. – Academia, Praha.
kitner M., lebeDa a., Doležalová i., Maras M., křístková e., nevo e., PavlíČek t., MeGlic v. & beharav a. (2008): AFLP analysis of Lactuca saligna germplasm collections from four European and three Middle Eastern countries. – Israel J. Plant Sci. 56: 185–193.
kitner M., Majeský Ľ., Gillová l., vyMyslický t. & naGler M. (in prep.): Genetic structure of Artemisia pancicii populations inferred from AFLP data. – Preslia.
kuMP b. & javornik b. (1996): Evaluation of genetic variability among common buckwheat (Fagopyrum esculentum Moench) populations by RAPD markers. – Plant Sci. 114: 149–158.
larkin P. j. (1976): Purification and viability determinations of plant protoplasts. – Planta 128: 213–216.lichter r. (1981): Anther culture of Brassica napus in a liquid culture medium. – Z. Pfl. Physiol. 103:
229–237.Moravec j., blažková D., hejný s., husová M., jeník j., kolbek j., krahulec f., kreČMer v., kroPáČ z.,
neuhäusl r., neuhäuslová-novotná z., rybníČek k., rybníČková e., saMek v. & ŠtěPán j. (1994): Fytocenologie. Nauka o vegetaci. – Academia, Praha.
MünzberGová z. (2001): Obnova druhově bohatých xerotermních trávníků na příkladu rezervací Stráně u splavu a Stráně. – Příroda, Praha, 19: 101–121.
naGler M. (2010): Standorte und Vergesellschaftung von Artemisia pancicii in Österreich. – Ms. [Diploma thesis, depon. in: Faculty of Life Sciences, University of Vienna, Vienna.]
Parihar j., haMal i. a., chhibber n. & sharMa n. (2009): Polination mechanism and indirect pollen presentation in Artemisia maritima L. – Int. J. Plant. Rep. Biol. 1: 191–193.
Peakall r. & sMouse P. e. (2006): GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. – Molec. Ecol. Notes 6: 288–295.
rabinowitz D. (1981): Seven forms of rarity. – In: synGe h. [ed.], The biological aspects of rare plant conservation, pp. 205–217, John Wiley & Sons, New York.

31
Příroda, Praha, 31: 11–31, 2011
rohlf f. j. (1998): NTSYS-pc. Numerical taxonomy and multivariate analysis system. – Applied Biostatatistics, New York.
rottenberG a. & Parker j. s. (2003): Conservation of the critically endangered Rumex rothschildianus as implied from AFLP diversity. – Biol. Conserv. 114: 299–303.
sawhney v. k. & shukla a. (1994): Male sterility in flowering plants: Are plant growth substances involved? – Am. J. Bot. 81(12): 1640–1647.
tero n., asPi j., siikaMäki P., jäkälänieMi a. & tuoMi j. (2003), Genetic structure and gene flow in a metapopulation of an endangered plant species, Silene tatarica. – Mol. Ecol 12: 2073–2085.
tichý l. (2002): JUICE, software for vegetation classification. – J. Veg. Sci. 13: 451–453.travis s. e., Maschinski j. & keiM P. (1996), An analysis of genetic variation in Astragalus cremnophylax
var. cremnophylax, a critically endangered plant, using AFLP markers. – Mol. Ecol. 5: 735–745.vos P., hoGers r., bleeker M., reijans M., vanDelee t., hornes M., frijters a., Pot j., PeleMan j.,
kuiPer M. & zabeau M. (1995): AFLP - a new technique for DNAfingerprinting. – Nucl. Acid Res. 23: 4407–4414.
wenDelberGer G. (1959): Die mitteleuropäischen Reliktvorkommen der Artemisia-Arten aus der Sektion Heterophyllae. – Verh. Zool.-Bot. Ver. Wien 98/99: 57–95.

32
Gillová et al.: Nové poznatky o Artemisia pancicii

Příroda, Praha, 31: 33–52, 2011
33
dynamika dvou vzácných druhů kapradin (Asplenium adulterinum a A. cuneifolium) na lokální a krajinné úrovnidynamics of two rare fern species (Asplenium adulterinum and A. cunei fo li um) on local and regional scale
anna bucharová1, 2, *, PřeMysL táJek3 & ZuZana MünZbergová1, 2
1Botanický ústav Akademie věd České republiky, Zámek 1, CZ-252 43 Průhonice2Katedra botaniky Přírodovědecké fakulty Univerzity Karlovy, Benátská 2, CZ-128 01 Praha 23Správa chráněné krajinné oblasti Slavskovský les, Hlavní 504, CZ-353 01 Mariánské Lázně* autor pro korespondenci: [email protected]
Abstract: Asplenium adulterinum and A. cuneifolium are rare species in the entire Europe. They are restricted to serpentine rocks. To identify factors responsible for their rarity, we studied their population and metapopulation dynamics. Specifically, we aimed at dynamics of the two fern species both on local and regional scale in the model region of Slavkovský les (the Slavkov Forest). To study the local dynamics, we employed matrix models using data from a field survey, both in forested and unforested habitats.
The results at the local scale indicate that populations of both species are slowly growing. The current stage distribution corresponds to the stable stage distribution, suggesting long time stability of the population dynamics. Even small populations have a reasonable chance to survive; however, there are differences between the two species. Individuals of both species are long lived – mean life span for A. adulterinum was estimated 34 year, A. cuneifo-lium 44 years.
We studied the dynamics on the regional scale by employing analysis of habitat occupancy by the species. First, we identified the characteristics of both species that are of key importance for habitat suitability. Subsequently, we identified localities which are suitable for the species but currently unoccupied. A high number of such localities in both species suggests dispersal limitation.
The results of our study both on local and regional scale suggest that both species are rare, but they are not threatened. Rareness of the species is given by rareness of the habitat as well as by the ability of the species to disperse to these habitats. Suitable unoccupied habitats exist, which suggests the species form metapopulations. The dynamics of the metapopulations is, however, extremely slow. Conservation of these species should concentrate on habitat protection. Protected should be not only occupied habitats, but also unoccupied serpentine rocks, since they are potential habitats for the two species. In the habitat protection, the most important is to avoid spruce plantation on serpentine rocks, because rocks in dense canopy of spruce are highly unfavourable for the two species.Keywords: fern, matrix model, metapopulation, naturally rare species, population dynamics, suitable unoccupied habitats
Abstrakt: Sleziník nepravý (Asplenium adulterinum) a sleziník hadcový (Asplenium cuneifolium) jsou celoevropsky vzácné druhy kapradin (C1, resp. C2 Červeného seznamu

34
Bucharová et al.: Dynamika Asplenium adulterinum a A. cuneifolium na lokální a krajinné úrovni
České republiky). Jsou striktně vázány na hadce. Pro takto vzácné druhy je důležité vědět, jak jejich populace prosperují a co je limituje v jejich dalším šíření.
V této studii jsme se zabývali dynamikou obou druhů jak na populační, tak na krajinné úrovni v modelovém území Slavkovského lesa. Lokální dynamiku jsme studovali pomocí maticových modelů, na šesti, respektive čtyř populacích obou druhů jak v otevřené krajině, tak v lese. Populace obou druhů zde dlouhodobě mírně rostou. Velikostní složení populací odpovídá stabilnímu velikostnímu složení, což vypovídá o dlouhodobé stabilitě populační dynamiky. I malé populace mají relativně vysokou pravděpodobnost přežití, jsou zde ale mírné odlišnosti mezi druhy. Rostliny jsou dlouhověké – průměrná délka života jedince je u A. adulterinum 34 let, u A. cuneifolium 44 let.
Dynamiku na úrovni krajiny jsme studovali pomocí analýzy obsazenosti jednotlivých skalek. S pomocí široké škály abiotických i biotických charakteristik skalek jsme identifikovali stanovištní nároky obou druhů. Za jejich pomoci jsme pak identifikovali skalky, které jsou vhodné pro růst jednotlivých druhů sleziníků, ale sleziníky na nich nerostou. Vysoký počet takovýchto skalek ukazuje, že oba druhy jsou, alespoň do jisté míry, limitovány šířením.
Výsledky studia jak na lokálním měřítku, tak v měřítku krajinném ukazují, že oba druhy jsou vzácné, ale nejsou ohrožené. Vzácnost druhů je dána vzácností jejich stanovišť a omezenou schopností druhů se na tato stanoviště rozšířit. Existují volné lokality, což naznačuje, že druhy tvoří metapopulaci. Dynamika jejich metapopulací je ale extrémně pomalá.
Ochrana obou sleziníků by se měla soustředit na ochranu jejich stanovišť. A to nejen stanovišť obsazených, ale i neobsazených, protože se zde druhy mohou přirozeně vyskytnout a vytvořit tak další populaci. Zejména by mělo být zamezeno osazování hadcových skalek smrkem, protože skalky v hustém zapojeném smrkovém porostu jsou pro zájmové druhy neobyvatelné.Klíčová slova: kapradiny, maticové modely, metapopulace, neobsazená vhodná stanoviště, populační dynamika, přirozeně vzácné druhy
Úvod
Sleziník nepravý (Asplenium adulterinum) a sleziník hadcový (A. cuneifolium) jsou velmi vzácné druhy kapradin. V Červeném seznamu České republiky (Procházka & holub 2000) náleží do C1, respektive C2 kategorie. U takto vzácných druhů jednoznačně vyvstává otázka, proč jsou vzácné, co je omezuje v jejich dalším šíření.
Asplenium adulterinum a A. cuneifolium jsou striktně vázány na hadce, nejsou schopny růst na jiném než tomto substrátu, až na vzácné výjimky (ŠPryňar & schläGelová 2005). Vzácnost těchto druhů je proto do jisté míry dána dostupností hadce. Ne na všech hadcích však druhy najdeme. Na některých hadcích neroste ani jeden druh těchto kapradin, někde roste pouze relativně hojnější A. cuneifolium a pouze v několika regionech najdeme samostatný výskyt i A. adulterinum. Vyvstává tedy otázka, co je určující pro rozšíření obou druhů, proč jsou vlastně tak vzácné.
Abychom si na danou otázku zodpověděli, je nutné pochopit faktory ovlivňující přežívání dru hu na více prostorových škálách. Nejprve na jedné lokalitě, kde je důležité, jak se po-pu laci druhu daří, zda je stabilní nebo zda roste či se zmenšuje, jaká je pravděpodobnost vymření populace atd. Pokud se druhu na lokalitě daří špatně, je to jistě důležitý faktor jeho vzácnosti. K zhodnocení dynamiky jednotlivých populací je nutné zjistit detaily o životním cyklu druhu – tj. růst, přežívání, reprodukce a mortalita a ty pak propojit do modelu celého životního cyklu (caswell 2001, leslie 1945, scheMske et al. 1994). Tento přístup se běžně a úspěš ně využívá pro analýzu životaschopnosti semenných druhů rostlin (Garcia 2003, Garcia et al. 2008, Griffith & forseth 2005, jonGejans et al. 2008, MenGes 1990). Pro kapradiny byl poprvé použit právě pro Asplenium adulterinum a A. cuneifolium (bucharová et al. 2010).

Příroda, Praha, 31: 33–52, 2011
35
Pokud se mu ale na lokalitách daří dobře, populace prospívá, je problém někde jinde. Proto je nutné se podívat na problematiku na větším měřítku a zkoumat, proč má druh tak málo populací. Tento pohled je u mnoha druhů velmi důležitý, protože jednotlivé populace často netvoří izolované jednotky, ale nějakým způsobem spolu komunikují. V dlouhodobějším časovém horizontu může docházet k vymírání populací a vlivem šíření druhů zase ke vzniku populací nových. Takový systém se nazývá metapopulace (hanski 1994). Na úrovni metapopulace jsou druhy omezeny ve svém rozšíření dvěma základními faktory, a to dostupností vhodných stanovišť a schopností se na tato stanoviště rozšířit (clark et al. 2007, turnbull et al. 2000). Oba faktory se samozřejmě mohou kombinovat (MünzberGová & herben 2005). V poslední době se poměrně hodně studií věnovalo právě otázce relativního významu těchto dvou faktorů. Valná většina těchto studií se věnovala semenným rostlinám. Podstatně méně je však známo o kapradinách.
Známou vlastností kapradin je produkce velkého množství velmi malých spor. Tyto spory jsou často považovány za všudypřítomné a v minulosti bylo časté tvrzení, že šíření neznamená pro kapradiny velký problém (Guo et al. 2003, tryon 1970, 1986). Novější studie, zejména ty, co využívají genetické markery, ukazují, že i kapradiny jsou ve svém šíření do jisté míry omezeny, a to v měřítku od několika set metrů do několika kilometrů (holDereGGer & schneller 1994, reisch et al. 2007, staPulionyte et al. 2006). Navíc bylo dokázáno, že valná většina spor kapradin dopadá do těsné blízkosti mateřské rostliny (Peck et al. 1990, PenroD & MccorMick 1996).
Z pohledu šíření se kapradiny od semenných rostlin liší v jednom důležitém faktu. U se menných rostlin dochází k šíření semen, tedy diaspor po oplodnění. Ze semene mů že přímo vyrůst nová rostlina. U kapradin se šíří haploidní spory, které po dopadu na příznivý materiál vyklíčí v gametofyt. Teprve na gametofytu dochází k oplodnění. Pokud je gametofyt oboupohlavný a schopný samooplození, může dát jedna jediná spora vznik novému sporofytu. Pokud je ale gametofyt jednopohlavný, nebo je z nějakého důvodu omezena možnost samooplození, je nutné, aby na vhodný substrát do těsné blízkosti dopadly nejméně dvě spory. Teprve potom může vzniknout sporofyt. Jednotlivé druhy kapradin se samozřejmě liší co do schopnosti samooplození gametofytu. Řada studií ukázala (flinn 2006, lanDe & scheMske 1985, voGel et al. 1999), že polyploidní druhy jsou spíše schopné gametofytického samooplození nežli druhy diploidní a tudíž se i lépe šíří.
Je tedy jasné, že šíření kapradin není vůbec triviální záležitost, ale naopak se jedná o velmi zajímavý a u různých druhů často důmyslný systém samo- či cizosprášení. Zohlednění způsobu šíření kapradin je z ochranářského hlediska velmi důležité. To platí i u námi studovaných druhů. Asplenium adulterinum je totiž allotetraploid (rodičovské druhy A. viride L. a A. trichomanes Huds. subsp. trichomanes, lovis & lovis 1955) a A. cune ifo-lium je diploid (Křísa in hejný & slavík 1988).
Pokud je rozšíření druhu omezeno schopností šíření, lze předpokládat, že v krajině budou existovat lokality, které jsou pro druh vhodné, ale neobsazené. Kdyby nebyl omezen šířením, dostal by se na veškerá vhodná stanoviště, a tudíž by žádná nebyla volná a druh by byl limitován pouze dostupností těchto stanovišť. K odhadu míry limitace šířením je tudíž klíčové si nadefinovat, jak takové vhodné volné stanoviště vypadá (aneb jak jej poznat, když tam druh neroste). S rozvojem metapopulační teorie, která identifikaci vhodných neobsazených stanovišť přímo vyžaduje, bylo vyvinuto několik metod jak taková stanoviště poznat – od vysévacích pokusů (MünzberGová & herben 2005) po růz-né distribuční modely (Moore & elMenDorf 2006) založené na abiotických proměnných prostředí (husbanD & barrett 1996, ouborG 1993, Quintana-ascencio & MenGes 1996, wilD & GaGnon 2005) nebo na druhovém složení vegetace daných stanovišť (např. MilDen et al. 2006, MünzberGová & herben 2004, roMerMann et al. 2007, treMlová & MünzberGová 2007).

36
Bucharová et al.: Dynamika Asplenium adulterinum a A. cuneifolium na lokální a krajinné úrovni
Tyto přístupy se ukázaly být vcelku spolehlivé pro identifikaci vhodných neobsazených stanovišť (MünzberGová & herben 2004).
Pokud zjistíme, že existují vhodná neobsazená stanoviště, a tudíž potvrdíme omezenou schopnost šíření druhu, je důležité vědět, na čem je schopnost druhu šířit se závislá. U větrem šířených druhů je pravděpodobné, že druh se bude hůře šířit na větší vzdálenosti. V této práci si klademe otázku, zda toto platí také pro kapradiny, jejichž spory se větrem šíří extrémně dobře.
Konkrétním cílem této práce je tak 1) ohodnotit dynamiku druhů Asplenium adulterinum a A. cuneifolium na úrovni populací a identifikovat, zda jsou populace stabilní, klesají či rostou, a 2) zjistit, co limituje druhy Asplenium adulterinum a A. cuneifolium v jejich šíření na úrovni krajiny.
Metodika
studované druhy
Asplenium adulterinum Milde a Asplenium cuneifolium Viv. (Aspleniaceae) jsou kapradiny striktně vázané na evropské hadce (viane et al. 1993). Jedna populace A. adulterinum se vyskytuje i na Vancouver Island v Kanadě (oGilvie & ceska 1984). Rozšíření obou druhů je velmi nesouvislé, na hadcových substrátech od Středomoří po Norsko a od Řecka po Španělsko (A. cuneifoliumi, A. adulterinum roste jen od Rakouska po Francii). V ČR se druhy vyskytují především v západních Čechách, s několika lokalitami v severovýchodních Čechách. Ve zbytku České republiky je jen několik málo relativně malých populací. Asplenium cuneifolium je obecně hojnější než A. adulterinum a má v ČR více lokalit (bucharová et al. 2010). Modelové území
Studie probíhala ve Slavkovském lese v západních Čechách. V této oblasti se vyskytuje velké množství hadcových skalek (http://www.geologicke-mapy.cz/). Centrum jejich výskytu je v oblasti fytochorionu 28 Tepelské vrchy, podokresu 28c Mnichovské hadce. Zde v regionu je přibližně na území 10 × 10 km rozseto 98 hadcových skalek. Hadcové skalky o rozloze 1 m2 až téměř 9 ha zde tvoří jakési ostrovy v kontextu okolní krajiny, která je tvořená jinými horninami a pokrytá zejména smrkovým lesem a kulturními kosenými nebo pasenými loukami. Převážná většina hadcových skalek se nachází v centrální oblasti hadcového hřebenu na ploše 7 × 4 km, několik málo jich je pak odlehlejších. Nejodlehlejší hadcová skalka je vzdálena 4 km od nejbližší jiné hadcové skalky.
Oba druhy hadcových kapradin jsou v modelovém území relativně časté, A. adulterinum je častější a obývá jak skalky v otevřené krajině, tak skalky v lese (převážně borovém a smrkovém). Z celkového počtu 98 skalek A. adulterinum roste na 66, v počtu od několika jedinců po téměř 2000 (tájek 2003). A. cuneifolium roste převážně na skalkách v lese, skalkám v otevřené krajině se vyhýbá a pokud zde již roste, populace jsou pouze nepočetné. Celkem A. cuneifolium obývá 48 skalek, populace jsou tvořeny několika jedinci až několika stovkami jedinců (tájek 2003).
Populační dynamika
Populační dynamika druhů byla studována na šesti lokalitách pro Asplenium adulterinum (tři v lese, tři v otevřené krajině) a čtyřech lokalitách pro A. cuneifolium (tři v lese, jedna

Příroda, Praha, 31: 33–52, 2011
37
v otevřené krajině), viz tabulka 1. Vzhledem k omezenému výskytu A. cuneifolium v otevřené krajině byla k dispozici pouze jedna lokalita, kde byl pro studii dostatečný počet jedinců.
Data byla sebrána v letech 2004–2007. Po důkladném prohlédnutí jedinců obou druhů byly definovány čtyři velikostní kategorie. Kategorie 1 zahrnovala nejmenší sporofyty, kategorie 4 plně vyvinuté velké dospělé jedince. Kategorie 2 a 3 vymezovaly postupný růst rostlin. Pro přesnou definici jednotlivých kategorií viz tabulka 2.
Přežívání a přechody mezi jednotlivými kategoriemiV každé populaci jsme označili 200–400 jedinců. Ke každému jedinci jsme připíchli barevný hřebík (barva značila příslušnost k velikostní kategorii). Tímto způsobem jsme označili všechny jedince v jasně vymezené oblasti (např. jednotlivá, jasně vymezená skalka). Každý rok v srpnu jsme rostliny zkontrolovali a přeznačili dle nové příslušnosti do velikostní kategorie. Rostliny kategorie 1 (nejmenší sporofyty) bez označení z minulého roku byly považovány za nově vzniklé jedince.
tab. 1. Lokality, na kterých probíhala studie populační dynamiky.table 1. Localities used for the study of population dynamics.
Kód/ code souřadnice/ coordinates Početnost populace/ Population size
A. adulterinum A. cuneifolium
Otevřená krajina/ Open landscape 1
50°04‘16‘‘ N; 12°47‘08‘‘ E
Několik stovek/ several hundreds
Několik stovek/ several hundreds
Otevřená krajina/ Open landscape 2
50°03‘41‘‘ N; 12°43‘54‘‘ E ~ 300 _________
Otevřená krajina/ Open landscape 3
50°03‘22‘‘ N; 12°44‘36‘‘ E ~ 1800 _________
Les/ Forest 1 50°04‘15‘‘ N; 12°47‘00‘‘ E
Několik stovek/ several hundreds
Několik stovek/ several hundreds
Les/ Forest 2 50°01‘43‘‘ N; 12°43‘34‘‘ E
Několik tisíc/ several thousands
Několik tisíc/ several thousands
Les/ Forest 3 50°03‘15‘‘ N; 12°46‘30‘‘ E
Několik tisíc/ several thousands
Několik tisíc/ several thousands
38
Bucharová et al.: Dynamika Asplenium adulterinum a A. cuneifolium na lokální a krajinné úrovni
Porovnání jednotlivých charakteristik životního cykluJednotlivé druhy a druhy na jednotlivých typech stanovišť (les/otevřená krajina) se mohou lišit v pravděpodobnosti růstu, mortalitě, reprodukci a přežívání ve stejné velikostní kategorii. Proto jsme tyto charakteristiky porovnali pomocí logistické regrese v programu R (r_DeveloPMent_core_teaM 2009). Pravděpodobnost těchto charakteristik životního cyklu se logicky liší v závislosti na počáteční velikosti rostliny, tato proměnná byla také do modelu zahrnuta, stejně jako rok sběru dat (data sbírána v několika letech). Detaily analýzy viz bucharová et al. (2010).
tab. 2. Kriteria, podle kterých byly rostliny rozděleny do velikostních kategorií (tzv. stage).table 2. Criteria used for categorisation of plants by size (stages).
Asplenium adulterinum Asplenium cuneifolium
Kategorie/stage 1
Nejmenší rostliny s vyvinutým listem, list kratší než 1,5 cm.
Nejmenší rostliny s vyvinutým listem, list kratší než 1,5 cm.
Kategorie/stage 2
Nejdelší list kratší než 3,5 cm. Žádný list nemá více než dva páry úkrojků.
Kategorie/stage 3
Maximálně pět listů, alespoň jeden list delší než 3,5 cm.
Žádný list nemá více než čtyři páry úkroků.
Kategorie/stage 4
Rostlina má alespoň šest listů delších než 3,5 cm. Pokud jsou listy delší než sedm cm, dostatečný počet listů jsou čtyři (dospělé rostliny rostoucí hluboko ve štěrbinách s několika málo dlouhými listy).
Alespoň jeden list má více než čtyři páry úkrojků.
Prosperita populacíK analýze demografických dat jsme použili maticový model (caswell 2001). Z terénních dat jsme získali pravděpodobnost přechodu mezi jednotlivými kategoriemi, pravděpodobnost úmrtí jedince dané velikostní kategorie atd. Tyto pravděpodobnosti tvoří tzv. přechodovou matici (A). Máme-li v čase t nějakou populaci o dané velikosti daného velikostního složení (tzv. populační vektor v), pak platí: v(t+1)= v(t)×A.
Výsledkem těchto modelů je populační růstová rychlost λ, která udává, kolikrát se populace zvětší za jeden časový cyklus, tedy rok. Pokud je λ = 1, populace zůstává stejná, pokud je λ < 1, populace se zmenšuje, pokud λ > 1, populace se zvětšuje.
Maticové modely skýtají hodně možností a výsledkem může být celá řada velmi užitečných charakteristik, jako je stabilní velikostní složení, průměrný věk jedince, pravděpodobnost vymření malých populací atd. Užitečným nástrojem je i tzv. analýza elasticity, která nám odhaluje kritické fáze životního cyklu. Ke všem charakteristikám lze spočítat intervaly spolehlivosti, případně testovat rozdíly mez jednotlivými druhy/stanovišti atd. Často se používá tzv. stochastická simulace, kdy se v každém časovém kroku populační vektor násobí

Příroda, Praha, 31: 33–52, 2011
39
náhodně vybranou maticí daného typu. To simuluje náhodnost (stochasticitu) prostředí. Tímto způsobem získáme populační růstovou rychlost např. pro lokality v bezlesí nebo celkovou pro daný druh. Dále lze přidat i demografickou stochasticitu.
V tomto článku je interpretační smysl jednotlivých charakteristik a čísel vždy popsán v textu výsledků nebo v popisku obrázků. Přesnější popis analýz viz bucharová et al. (2010), principy výpočtu pak caswell (2001).
dynamika na úrovni krajiny
Sběr datData byla sebrána v rámci diplomové práce (tájek 2003) v letech 2001 až 2003. Veškeré hadcové skalky byly identifikovány z geologických map a následně v terénu zmapovány pomocí GPS. Byly nalezeny i nové skalky, v geologických mapách neuvedené. Jednotlivé lokality často zahrnovaly jemnou mozaiku skalek a neskalnatého terénu. Jednotlivé skalky byly tedy označeny jako nespojité, pokud mezi skalkami byla mezera alespoň 20 m. Pro všechny skaly byly shromážděny údaje uvedené tabulce 3.
Velikost lokalit byla změřena v terénu pomocí pásma (do 100 m2) nebo v GIS. Na lokalitách byla přítomna i neskalnatá místa, pro potřeby analýz byla proto rozloha upravena pomocí kvalifikovaného odhadu podílu skalnatých biotopů na skalkách. Dále byla změřena výška nejvyšší vertikální skalky. Byla zaznamenána přítomnost mikrostanovišť: skalní štěrbiny, skalní terásky, vrcholové plošiny, suť a svahy.
Na všech lokalitách byly zaznamenány všechny přítomné druhy cévnatých rostlin a typ vegetace (bezlesí, smrkový les, borový les).
Analýza dat
Druhové složeníDruhové složení vstupovalo do dalších analýz ve formě tzv. Bealsova indexu (beals 1984, MünzberGová & herben 2004). Tento index odráží skutečnost, že některé druhy spolu rostou častěji než jiné druhy. Čím vyšší Bealsův index má daná lokalita, tím je společenstvo pro druh „vhodnější“, přesněji, druhy na daném místě byly v kalibračním souboru (v tomto případě stejný soubor hadcových skalek) častěji nalezeny ve společnosti cílového druhu. Bealsův index pro danou lokalitu je samozřejmě odlišný pro A. adulterinum a A. cuneifolium (detaily viz tájek et al. 2011).
Vhodnost lokality pro zájmové druhyCílem bylo co nejpřesněji charakterizovat lokality, které jsou pro daný druh (A. adulterinum nebo A. cuneifolium) vhodné. Proto jsme sestavili model, který zahrnoval důležité charakteristiky lokality a který co nejpřesněji vysvětloval presenci nebo absenci zájmového druhu. Lokality s výskytem druhu měly vysokou hodnotu modelu, lokality bez druhu nízkou. Mohly existovat i lokality, které měly vysokou hodnotu modelu, tj. měly stejné parametry jako lokality obsazené (a pro druh tedy vhodné), ale druh na nich nerostl. Tyto skalky byly potom označeny jako vhodné neobsazené lokality. Statistické výpočty jsme provedli v programu R (r DeveloPMent core teaM 2009).
Jako model jsme zvolili logistickou regresi. Závislá proměnná byla binární, přítomnost/nepřítomnost cílového druhu. Vysvětlující proměnné byly výška nejvyšší skalky na lokalitě, přítomnost jednotlivých typů mikrostanovišť, typ vegetace, a dále průměrné Ellenbergovy hodnoty pro světlo, teplotu, kontinentalitu, vlhkost, půdní reakci a úživnost (ellenberG et al. 1992), a Bealsův index. Model nezahrnoval rozlohu
40
Bucharová et al.: Dynamika Asplenium adulterinum a A. cuneifolium na lokální a krajinné úrovni
stanoviště. Je logické, že diaspora má větší šanci dopadnou na větší území než na menší. Nás ale v tuto chvíli zajímaly charakteristiky, které dělají stanoviště obyvatelným a zahrnutí rozlohy by výsledek znepřesnilo. Pro každý druh jsme vytvořili samostatný model, který jsme použili jak k odhalení charakteristik stanoviště, které toto stanoviště dělají obyvatelným, tak následně k určení vhodných ale neobsazených stanovišť (detaily modelu viz tájek et al. 2011).
Neobsazená stanoviště, která měla vyšší nebo stejnou hodnotu modelu než všechna obsazená stanoviště jsme považovali za vhodná neobsazená stanoviště. Všechna obsazená stanoviště byla považována za vhodná.
tab. 3. Charakteristiky lokalit použité pro identifikaci vhodných neobsazených lokalit.table 3. Characteristics of localities used for identification of suitable unoccupied localities.
nezávislá proměná/ independent variable Hodnoty/ values
Výška skalky 0,2–10,5 m (kontinuální)
Rozloha (log transformace) 1−89 288 m2 (kontinuální)
Počet typů mikrostanovišť 1−6; kontinuální
Bealsův index (pro každý druh jiný) 0−1, kontinuální
Typ vegetace
Otevřená krajina 0−1, kontinuální
Borový les 0−1, kontinuální
Smrkový les 0−1, kontinuální
mikrostanoviště
Svahy Presence/absence
Štěrbiny Presence/absence
Suť Presence/absence
Plateau Presence/absence
Terásky Presence/absence
Ellenbergovy indikační hodnoty
Světlo 4,8−7,3; kontinuální
Teplota 3−5,6; kontinuální
Kontinentalita 3,2−4,5; kontinuální
Vlhkost 3,9−6; kontinuální
Půdní reakce 2−6,1; kontinuální
Živiny 1−5,8; kontinuální

Příroda, Praha, 31: 33–52, 2011
41
Izolovanost stanovištěStudovali jsme, zda pravděpodobnost obsazenosti vhodného stanoviště závisí na jeho izolovanosti. Izolovanost stanoviště jsme vypočítali na základě jeho rozlohy a vzdálenosti ke všem ostatním stanovištím (opět váženo rozlohou každého stanoviště). Potom jsme sestavili model (logistikou regresi), kde byla presence/absence druhu na lokalitě vysvětlována pomocí izolovanosti stanoviště a jeho rozlohy (detaily modelu viz tájek et al. 2011).
výsledky
Populační dynamika
Populační růstová rychlostStochastická populační růstová rychlost všech populací obou druhů je vyšší než 1 (přibližně 1,01–1,10), tj. populace mírně rostou. Celková stochastická populační růstová rychlost pro A. cuneifolium je signifikantně vyšší (1,067, P= 0,037, permutační test) než pro A. adu lte-rinum (1,046). V rámci druhu se stochastická populační růstová rychlost nelišila mezi typy stanoviště (les/bezlesí).
Pravděpodobnost extinkce malých populacíV této analýze jsme využili jak environmentální, tak demografickou stochasticitu pro simulaci osudu velmi malé populace – deset dospělých jedinců největší velikostní kategorie. Takhle malé populace obou druhů se ve studovaném území vyskytují. Projekci přes 20 let a 50 let jsme zopakovali 1000× a podívali se, v kolika % došlo k vymření populace. Toto číslo vyjadřuje pravděpodobnost extinkce.
Pro A. adulterinum je pravděpodobnost extinkce takto malé populace v bezlesí 24 %, respektive 39 % (20 a 50 let), v lese 25 %, respektive 42 %. Pravděpodobnost extinkce v lese a bezlesí je pro tento druh tedy podobná. Zcela jiná je situace u A. cuneifolium: v bezlesí je pravděpodobnost extinkce podobná jako u A. adulterinum (22 %, respektive 40 %), v lese však mnohem menší – 12 %, respektive 22 %).
Pokud jsme počáteční populaci zvolili větší (50 velkých jedinců), byla již pravděpodobnost extinkce pro oba druhy velmi nízká – méně než 5 %.
Jednotlivé charakteristiky životního cykluV této analýze jsme zkoumali, zda se jednotlivé charakteristiky životního cyklu (pravdě podobnost růstu, přežívání ve stejné velikostní kategorii, reprodukce, retrogrese – „zmenšování“, a mortality) liší mezi druhy, případně mezi jednotlivými biotopy (les/bezlesí). Nejdůležitějším výsledkem je, že Asplenium adulterinum v obou biotopech a Asplenium cuneifolium v lese má relativně malou pravděpodobnost mortality. S tím příkře kontrastuje pravděpodobnost mortality jedince A. cuneifolium v bezlesí – zde je pravděpodobnost mortality několikanásobně vyšší (obr. 1).
Poté jsme se podívali (v tzv. analýze elasticity), které fáze životního cyklu jsou tzv. kritické, tj. mají největší vliv na růstovou rychlost populací. Přesněji, o kolik se změní populační růstová rychlost, pokud změníme pravděpodobnost daného přechodu mezi kategoriemi o jednotku. Ty přechody, pro které tato malá změna v jejich pravděpodobnosti způsobí, že relativně největší změnu v populační růstové rychlosti jsou pro populační dynamiku nejdůležitější. V případě obou našich druhů se ukázalo, že nejdůležitější je přežívání rostlin v takovém stavu, v jakém jsou, zejména pak takovéto přežívání velkých, dospělých rostlin nejvyšší velikostní kategorie. Podstatně menší důležitost má růst, zdaleka nejmenší pak reprodukce.
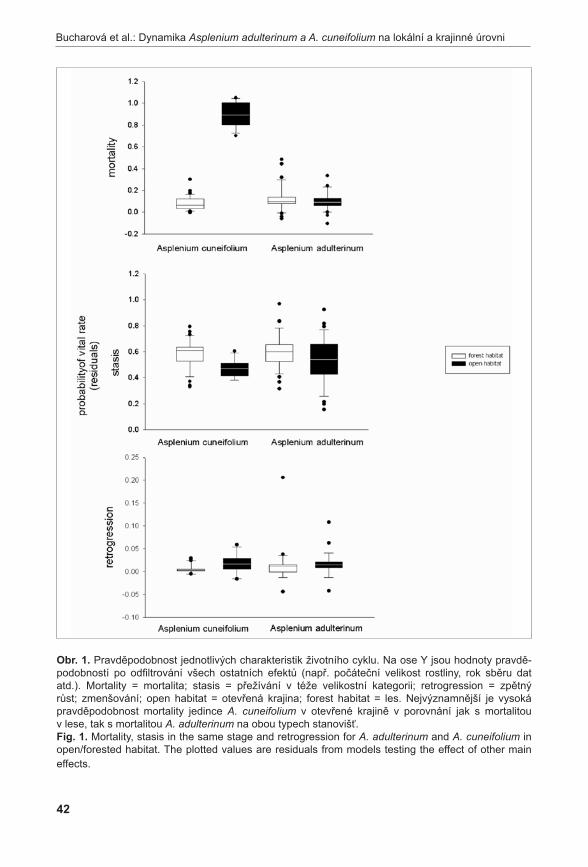
42
Bucharová et al.: Dynamika Asplenium adulterinum a A. cuneifolium na lokální a krajinné úrovni
obr. 1. Pravděpodobnost jednotlivých charakteristik životního cyklu. Na ose Y jsou hodnoty pravdě-podobností po odfiltrování všech ostatních efektů (např. počáteční velikost rostliny, rok sběru dat atd.). Mortality = mortalita; stasis = přežívání v téže velikostní kategorii; retrogression = zpětný růst; zmenšování; open habitat = otevřená krajina; forest habitat = les. Nejvýznamnější je vysoká pravděpodobnost mortality jedince A. cuneifolium v otevřené krajině v porovnání jak s mortalitou v lese, tak s mortalitou A. adulterinum na obou typech stanovišť.Fig. 1. Mortality, stasis in the same stage and retrogression for A. adulterinum and A. cuneifolium in open/forested habitat. The plotted values are residuals from models testing the effect of other main effects.
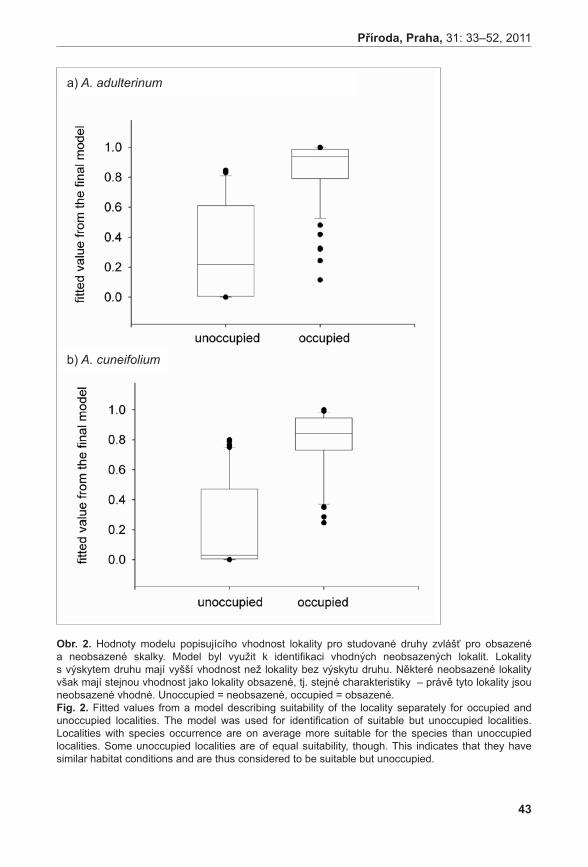
Příroda, Praha, 31: 33–52, 2011
43
a) A. adulterinum
b) A. cuneifolium
obr. 2. Hodnoty modelu popisujícího vhodnost lokality pro studované druhy zvlášť pro obsazené a neobsazené skalky. Model byl využit k identifikaci vhodných neobsazených lokalit. Lokality s výskytem druhu mají vyšší vhodnost než lokality bez výskytu druhu. Některé neobsazené lokality však mají stejnou vhodnost jako lokality obsazené, tj. stejné charakteristiky – právě tyto lokality jsou neobsazené vhodné. Unoccupied = neobsazené, occupied = obsazené.Fig. 2. Fitted values from a model describing suitability of the locality separately for occupied and unoccupied localities. The model was used for identification of suitable but unoccupied localities. Localities with species occurrence are on average more suitable for the species than unoccupied localities. Some unoccupied localities are of equal suitability, though. This indicates that they have similar habitat conditions and are thus considered to be suitable but unoccupied.

44
Bucharová et al.: Dynamika Asplenium adulterinum a A. cuneifolium na lokální a krajinné úrovni
Stabilní velikostní složeníPokud provedeme projekci populačního vektoru přes dostatečně dlouhou dobu, populační vektor, nehledě na celkový počet jedinců, se ustálí v určitém rozložení – např. 15 % jedinců kategorie 1, 10 % kategorie 2, 20 % kategorie 3 a 55 % kategorie 4. Získáme tzv. stabilní velikostní složení. Pokud současné rozložení populace odpovídá tomuto stabilnímu velikostnímu složení, tak dynamika, kterou jsme zachytili v době pokusu, odpovídá dlouhodobé dynamice. Pokud je zde velká odchylka, znamená to, že naše data nereprezentují dlouhodobou populační dynamiku.
V případě naší studie, stabilní velikostní složení všech populací obou druhů odpovídá aktuálnímu velikostnímu složení populace (až na jednu lesní populaci A. adulterinum, kde odchylka není příliš velká). Znamená to, že námi popsaná populační dynamika probíhá v populacích již poměrně dlouhou dobu.
Průměrný věk rostlinRostliny A. adulterinum se průměrně dožívají 34 let (v jednotlivých populacích od 26,4 do 56,4 let). Rostliny A. cuneifolium se dožívají průměrně více, 43,9 let (v jednotlivých populacích 24,8 až 67,6 let).
Spolehlivost modelůProtože si uvědomujeme, že jako všechny modely, i ten náš nemusí ve všem 100 % odpovídat realitě, ověřili jsme si jeho spolehlivost na reálných datech. Pro dvě studované populace A. adulterinum byl k dispozici celkový počet jedinců v populaci v roce 1997 a 2006 (tájek 2003, P. tájek nepublikováno). Pomocí našeho modelu jsme tedy na základě počtu jedinců na dané lokalitě v roce 1997 vypočítali počet jedinců v roce 2006 a získané číslo jsme porovnali s reálným počtem. Pro obě populace reálný počet jedinců v roce 2006 spadá do intervalu spolehlivosti počtu jedinců vypočítaného z modelu.
dynamika druhu na úrovni krajiny
Parametry vhodných stanovišťAsplenium adulterinum preferuje vyšší skalky. Potřebuje výskyt skalních štěrbin a preferuje lokality s přítomností skalní suti. Vhodnější jsou skalky s více kontinentálně laděnými mikro-stanovišti – většími extrémy teplot a srážek v rámci roku. Tento druh se naopak spíše vyhýbá více úživným stanovištím (lokality s vyšší průměrnou Ellenberovou hodnotou pro živiny) a kyselejším skalkám (průměrné Ellenbergovy hodnoty pro pH). Druhové složení stanovišť, vyjádřené jako Bealsův index, nemělo v našem modelu k výskytu A. adulterinum žádný vztah.
Asplenium cuneifolium také preferuje vyšší skalky a částečně je také vázáno na skalky se skalními štěrbinami a preferuje skalky s výskytem suti. Navíc, preferuje skalky s výskytem skalních terásek. Spíše se vyhýbá vlhkým skalkám (Ellebergovy hodnoty), skalkám v bezlesí, a kyselejšímu prostředí. Úživnost skalky neměla na obsazenost lokalit A. cuneifolium žádný vliv. Druhové složení, vyjádřené jako Bealsův index, mělo poměrně silnou vazbu na výskyt A. cuneifolium. Zajímavá je absence vlivu smrkového lesa na výskyt obou druhů kapradin. V tomto případě je to pravděpodobně dáno hrubým měřítkem sběru dat (tájek et al. 2011).
Na základě hodnot modelu pro každý druh jsme identifikovali vhodné neobsazené lokality – ty neobsazené, které měly hodnotu modelu vyšší nebo stejnou nežli poslední obsazená lokality s nejnižší hodnotou modelu (obr. 2). Pro A. adulterinum bylo z 32 neob sazených lokalit 15 pro druh vhodných a 17 nevhodných. Pro A. cuneifolium bylo z 50 neobsazených lokalit 18 vhodných a 32 nevhodných. Vhodné lokality (obsazené

Příroda, Praha, 31: 33–52, 2011
45
i neobsazené) pro A. cuneifolium se nachází převážně v centrální části studovaného území s jednou odlehlou obsazenou skalkou ca čtyři km severovýchodně od hlavního hadcového hřebene. Vhodné lokality pro A. adulterinum (obsazené i neobsazené) jsou jak v centrální části sudovaného území, tak na odlehlejších hadcových skalkách (obr. 3 a 4).
Efekt izolovanosti a velikosti stanovištěV této analýze jsme zkoumali, do jaké míry je obsazenost stanoviště dána jeho izolovaností a rozlohou. Pracovali jsme již pouze s vhodnými stanovišti (ať již obsazenými nebo neobsazenými). Rozloha stanoviště neměla u obou druhů signifikantní vliv na obsazenost stanoviště. Izolovanost stanoviště neměla žádný vliv na jeho obsazenost A. cuneifolium, ale měla vliv na obsazenost skalek druhem A. adulterinum (13 % variability, p = 0,001) – izolovanější stanoviště byla obsazena méně často.
diskusePopulační dynamika
Naše studie odhalila, že populace druhů Asplenium adulterinum a A. cuneifolium ve Slavkovském lese jsou v dobrém stavu, počet jedinců v populacích se dlouhodobě mírně zvyšuje. Dokonce i velmi malé populace obou druhů mají relativně vysokou šanci přežít.
Oba druhy se dle našich modelů dožívají průměrně několika desítek let – A. adulterinum 34 let, A. cuneifolium 43 let. Oba druhy se mohou v ojedinělých případech (1 % populace) dožívat i přes 100 let. Jedná se tedy o rostliny vpravdě dlouhověké. Zjištění z modelů v tomto směru odpovídají terénním pozorováním, kdy jsou zdokumentováni jedinci žijící minimálně 30, respektive 40–50 let (S. Jeβen, úst. sdělení). Dlouhověkost není příliš překvapivá, protože je typická pro rostliny adaptované na drsné prostředí (forbis & Doak 2004, Garcia 2003, Garcia et al. 2008), jímž skalní štěrbiny na hadcovém substrátu bezpochyby jsou. S dlouhověkostí souvisí i důležitost jednotlivých charakteristik životního cyklu a kritické fáze životního cyklu. Nejdůležitější je přežívání, zejména přežívání velkých rostlin. To je pro dlouhověké druhy typické (silvertown et al. 1993).
Ačkoli je populační dynamika obou druhů do značné míry podobná, najdeme zde důležité rozdíly. Nejdůležitější je rozdílná pravděpodobnost extinkce malých populací v otevřené krajině a v lese u jednotlivých druhů. Zatímco u Asplenium adulterinum je pravděpodobnost extinkce stejná u populací v otevřené krajině jako v lese (okolo 40 % za 50 let, populace deseti jedinců), u A. cuneifolium je extinkce nápadně nižší v lese (22 %) než v otevřené krajině (40 %). To je pravděpodobně důvodem, proč A. cuneifolium ve Slavkovském lese roste převážně v lese a populací v otevřené krajině je minimum. Populace tohoto druhu v lese mají nižší pravděpodobnost extinkce. Pokud k extinkci dojde, je populace obnovena kolonizací. Extinkce v otevřené krajině je však rychlejší, kolonizace tak nestačí lokality obnovovat a výsledkem je nižší počet populací. A to i přesto, že se populacím v otevřené krajině daří dobře (růstová rychlost vyšší než 1). Na druhou stranu A. adulterinum má populace jak v otevřené krajině, tak v lese, na obou typech stanovišť je podobná, relativně vysoká pravděpodobnost extinkce (40 % jako u A. cuneifolium v otevřené krajině). Druh však pravděpodobně stačí vymřelé populace obnovovat kolonizací. To naznačuje vyšší kolonizační potenciál A. adulterinum v porovnání s A. cuneifolium.
Vyšší kolonizační schopnosti A. adulterinum jsou očekávatelné. Jedná se o tetraploida a u kapradin je ploidie často spojená se systémem reprodukce – u polyploidů převažuje vnitogametofytické oplození a tudíž jsou schopni kolonizace jednou sporou. U diploidů
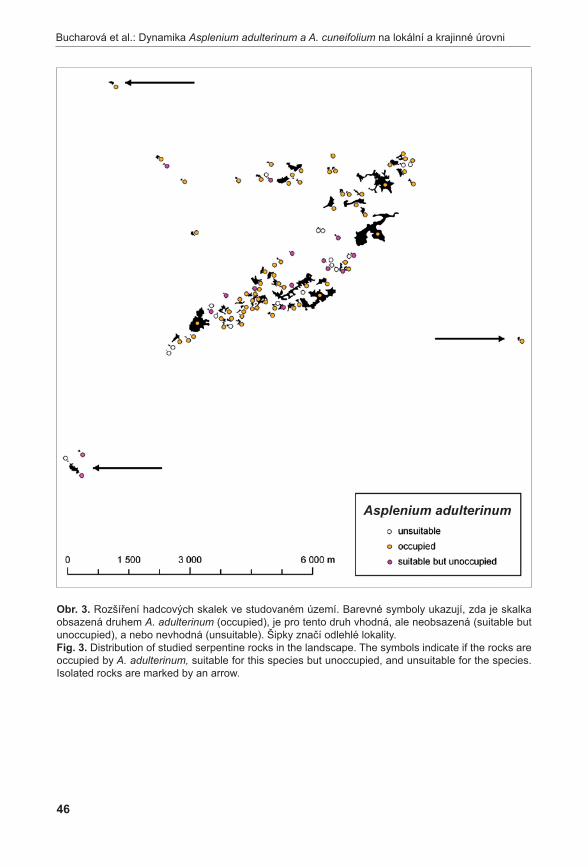
46
Bucharová et al.: Dynamika Asplenium adulterinum a A. cuneifolium na lokální a krajinné úrovni
Asplenium adulterinum
obr. 3. Rozšíření hadcových skalek ve studovaném území. Barevné symboly ukazují, zda je skalka obsazená druhem A. adulterinum (occupied), je pro tento druh vhodná, ale neobsazená (suitable but unoccupied), a nebo nevhodná (unsuitable). Šipky značí odlehlé lokality. Fig. 3. Distribution of studied serpentine rocks in the landscape. The symbols indicate if the rocks are occupied by A. adulterinum, suitable for this species but unoccupied, and unsuitable for the species. Isolated rocks are marked by an arrow.
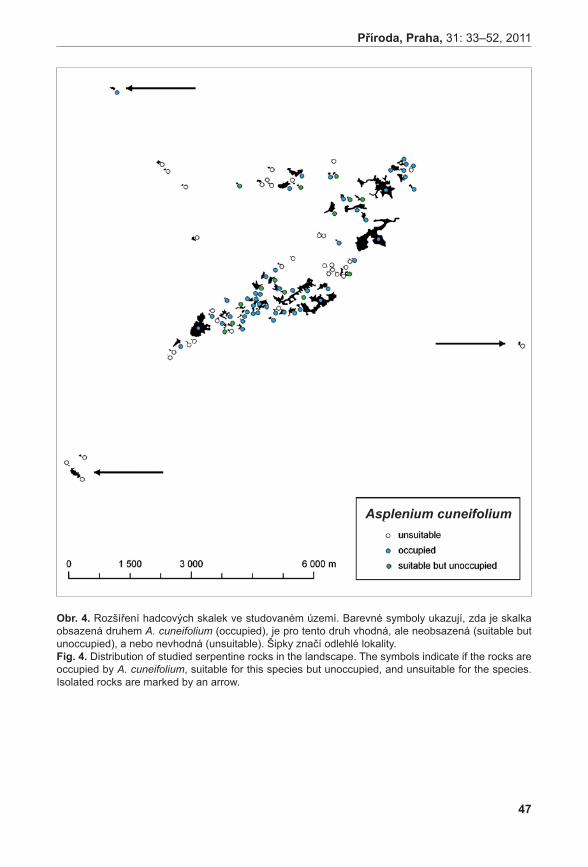
Příroda, Praha, 31: 33–52, 2011
47
Asplenium cuneifolium
obr. 4. Rozšíření hadcových skalek ve studovaném území. Barevné symboly ukazují, zda je skalka obsazená druhem A. cuneifolium (occupied), je pro tento druh vhodná, ale neobsazená (suitable but unoccupied), a nebo nevhodná (unsuitable). Šipky značí odlehlé lokality. Fig. 4. Distribution of studied serpentine rocks in the landscape. The symbols indicate if the rocks are occupied by A. cuneifolium, suitable for this species but unoccupied, and unsuitable for the species. Isolated rocks are marked by an arrow.

48
Bucharová et al.: Dynamika Asplenium adulterinum a A. cuneifolium na lokální a krajinné úrovni
(jako je A. cuneifolium) převažuje oplodnění mezi gametofyty a kolonizace jednou sporou je teoreticky obtížná (soltis & soltis 1992).
dynamika na úrovni krajiny
Charakteristika lokalit vhodných pro zájmové druhyOba druhy kapradin, Asplenium adulterinum a A. cuneifolium, jsou vázány na hadcové skalky. Ne všechny skalky v regionu jsou však pro druhy vhodné. Nejprve bylo nutné definovat, jak vhodné skalky vypadají.
Pro oba druhy je klíčové, zda se na skalkách vyskytují vhodná mikrostanoviště, zejména skalních štěrbiny. Asplenium cuneifolium se vyskytuje také na skalkách s malými skalními teráskami s mírně hlubší půdou, kde se již vyskytují i další druhy rostlin (např. Calamagrostis villosa). Oba druhy také preferují vyšší skalky, což souvisí s únikem před konkurencí ostatních druhů rostlin. Asplenium adulterinum se navíc vyhýbá skalkám bo-ha tým na živiny. Hadcové skalky jsou na živiny přirozeně chudé (chiarucci et al. 1999, turitzin 1982), jejich zvýšený obsah znamená eutrofizaci a degradaci biotopu (ricotta et al. 2004). Je proto logické, že druhy striktně vázané na hadce se budou takto degradovanými biotopům vyhýbat.
Přítomnost stromového patra má efekt na výskyt A. cuneifolium (roste na skalkách v lese, méně v otevřené krajině), ale ne na výskyt A. adulterinum. To souvisí s vyšší pravděpodobností vymření A. cuneifolium v otevřené krajině a celkově pomalejší metapopulační dynamikou (bucharová et al. 2010). Zajímavá je absence vlivu smrku na rozšíření obou druhů. Toto zjištění je v kontrastu s terénní zkušeností. Je to pravděpodobně dáno hrubým měřítkem sběru dat o vegetaci (tájek 2003). Výskyt smrkových výsadeb byl zaznamenáván na úrovni celých lokalit. Hadcové kapradiny se vyskytují ve skalních štěrbinách a i na skalkách pokrytých z velké části smrkovým lesem se najdou malé otevřené prostory, kde na skalku dopadá slunce, je zde minimum smrkového opadu a v malém měřítku se tak vyskytují podmínky vhodné pro hadcové kapradiny.
Spolehlivost identifikace vhodných neobsazených lokalitPoté, co jsme vytvořili model, který úspěšně charakterizoval obsazené a neobsazené skalky, jsme mohli identifikovat lokality pro druh vhodné, ale druhem neobsazené. Pro obě kapradiny jsme takové skalky našli, což znamená že oba druhy jsou alespoň do jisté míry limitovány schopností šíření. Uvědomujeme si, že toto tvrzení je klíčově závislé na kvalitě našeho modelu.
Náš model vysvětlil 52 % (A. adulterinum) a 47 % (A. cuneifolium) variability přítomné v datech. To je poměrně hodně ve srovnání s jinými studiemi podobného charakteru (0–46%, ehrlén et al. 2006). Čím více variability v datech model vysvětluje, tím lépe odráží skutečnost a lépe se dá použít k identifikaci vhodných neobsazených stanovišť (Moore & elMenDorf 2006). Námi identifikovaná vhodná a nevhodná stanoviště navíc odpovídala terénní zkušenosti Navíc na jedné lokalitě, definované jako vhodná neobsazená pro A. cuneifolium (na základě dat z roku 2003), byl druh v roce 2006 nalezen.
Izolovanost lokalitPravděpodobnost obsazenosti vhodných stanovišť A. adulterinum klesá se vzrůstající izolovaností daného stanoviště. Podobný trend byl mnohokráte nalezen u semenných rostlin (franzen & eriksson 2003, kolb & DiekMann 2005, ouborG 1993, Quintana-ascencio & MenGes 1996). U kapradin, které tvoří velké množství malých spor, je toto zjištění však méně očekávané.

Příroda, Praha, 31: 33–52, 2011
49
Průkazný efekt izolovanosti stanoviště znamená, že druh se šíří lépe na bližší stanoviště než na ta vzdálenější. Absence efektu izolovanosti u A. cuneifolium tak může mít dvě kontrastující vysvětlení: druh se buď šíří všude dobře, a nebo všude velmi špatně a jeho současné rozšíření v krajině je dáno jinými, pravděpodobně historickými faktory (brown & boutin 2009, chýlová & MünzberGová 2008, linDborG & eriksson 2004). Vzhledem k tomu že A. cuneifolium roste na menší proporci vhodných stanovišť (73 %) než A. adulterinum (84 %), velmi špatné šíření je pravděpodobnější. Tomu odpovídá i předpokládaný systém rozmnožování těchto druhů. Jak ukazují genetické analýzy provedené v populacích obou druhů (A. Bucharová & Z. Münzbergová, rukopis), u druhu A. adulterinum (tetraploid) převažuje pravděpodobně samooplození v rámci gametofytu a je tudíž dobře schopno kolonizace jedinou sporou. Na druhou stranu u druhu A. cuneifolium (diploid) pravděpodobně převažuje oplození mezi gametofyty. Tento předpoklad je ve shodě s jinými publikovanými pracemi věnujícími se rozmnožovacímu systému kapradin různých ploidií (flinn 2006, lanDe & scheMske 1985, voGel et al. 1999). Pro vznik sporofytu A. cuneifolium je tedy pravděpodobně nutné, aby dvě spory dopadly do těsné blízkosti do vhodných podmínek (Peck et al. 1990), vytvořily gametofyty a teprve pak může dojí k oplodnění. To je poměrně málo pravděpodobné a může tak vysvětlit pozorované špatné šíření tohoto druhu.
Závěr a doporučení pro druhovou ochranu
Velké populace Asplenium adulterinum a A. cuneifolium ve Slavkovském lese mírně rostou a tato populační dynamika je dlouhodobě stabilní. Kritickou fází životního cyklu je přežívání velkých rostlin. Rostliny žijí několik desítek let a i malé populace mají relativně vysokou šanci přežít. Nejedná se o druhy ohrožené, ale přirozeně vzácné. Vzácnost je do jisté míry dána specifickým substrátem – hadcem. Pro ochranu druhů je vhodné udržovat jejich stanoviště v optimálním stavu, žádné další zásahy typu kultivace ex situ nejsou nutné.
Při údržbě lokalit je vhodné vyhnout se eutrofizaci lokalit. Největším zdrojem živin byla v minulosti umělá hnojiva aplikovaná na přilehlá pole. To je na lokalitách stále patrné, protože vegetace se ze zvýšené aplikace živin vzpamatovává poměrně dlouho (norDin et al. 2005). V současnosti se na obohacování lokalit o živiny podílí atmosférická depozice (fottová 2003). Na vybraných skalkách je jako management prováděna pastva ovcí a koz. V rámci pastevního režimu nocuje dobytek mimo hadcové skalky, aby se zabránilo eutrofizaci.
Pravděpodobně nejsilnějším negativním vlivem působícím na Asplenium adulterinum a A. cuneifolium ve Slavkovském lese jsou smrkové výsadby. Smrk stíní skalky hustým korunovým zápojem, podporuje růst mechu a smrkový opad se špatně rozkládá a zaplňuje skalní štěrbiny, které jsou hlavním biotopem obou druhů. Pro zachování vhodných lokalit pro oba druhy je nutné vyhnout se smrkovým výsadbám nebo přirozenému zmlazení smrku. Ve stromovém patře je třeba podporovat borovici lesní, která stanoviště negativně neovlivňuje.
Na úrovni krajiny jsou Asplenium adulterinum a A. cuneifolium ve svém rozšíření omezeny jak schopností šířit se, tak dostatkem a hustotou vhodných stanovišť (hadcových skalek je omezené množství). Oba druhy kapradin mají ve studovaném území metapopulační dynamiku. Jejich dynamika je ale velmi pomalá. To znamená, že některé populace vlivem stochastických procesů vymírají, na jiných vhodných biotopech vznikají nové populace kolonizací. Kolonizace nových stanovišť je efektivnější u Asplenium adulterinum než u A. cu neifolium. V rámci péče o oba druhy je proto vhodné udržovat nejen skalky se současným výskytem druhu, ale i hadcové skalky bez těchto druhů, protože se může jednat o dočasně neobsazené, ale vhodné stanoviště druhu.

50
Bucharová et al.: Dynamika Asplenium adulterinum a A. cuneifolium na lokální a krajinné úrovni
summary
Large populations of Asplenium adulterinum and A. cuneifolium in the Slavkov Forest are slightly increasing and the population dynamics is stable over time. The critical phase of the life cycle is the survival of large plants. The plants live for decades and even small populations have a relatively high chance of surviving. Asplenium adulterinum and A. cuneifolium are thus not endangered species, but species which are naturally rare. Their rarity is to some extent given by their specific substrate – serpentine rocks. For the protection of species it is advisable to maintain their habitats in optimal condition, no further actions such as ex situ cultivation are necessary.
Protection of the localities should include avoiding eutrophication. The biggest source of nutrients in the past were fertilizers applied to adjacent fields. Previous fertilization is visible in the vegetation of the localities for several decades (norDin et al. 2005). Currently, the most important source of nutrient enrichment is probably atmospheric deposition (fottová 2003). Some of the rocks are managed by grazing of sheep and goats. It is important that the grazing animals spend the night outside the serpentine rock to prevent eutrophication.
Probably the strongest negative factor affecting survival of Asplenium adulterinum and A. cuneifolium in the Slavkov Forest are spruce plantations. Spruce trees shade the rocks by dense crown cover supporting the growth of moss. In addition, spruce litterfall decomposes very slowly and fills the crevices, which are the main habitat for both Asplenium species. To maintain suitable habitats for both species it is necessary to avoid spruce planting or natural regeneration of spruce at the localities. Spruce should be replaced by pine as pine trees do not have any negative effect on growth of Asplenium.
At the landscape level, Asplenium adulterinum and A. cuneifolium are limited in their expansion by their ability to spread as well as by availability of suitable serpentine rocks (number of serpentine rocks in the region is limited). Both species of ferns perform as metapopulations. This means that some populations are dying out due to stochastic processes, while other suitable habitats are being colonised. This process is, however, very slow. Colonization of new habitats is more efficient for Asplenium adulterinum than in A. cuneifolium. To protect both species it is therefore important to protect not only rocks with occurrence of the species, but even rocks without these species. These can represent temporarily unoccupied but suitable habitats.
Poděkování
Práce byla finančně podpořena granty MŠMT VaV 2B06178, FRVŠ 348/2004 a GAUK 176/2006, částečně MŠMT 0021620828 a AV0Z60050516.
literatura
beals e. w. (1984): Bray-Curtis ordination: an effective strategy for analysis of multivariate ecological data. – Adv. Ecol. Res. 14: 1–56.
brown c. D. & boutin c. (2009): Linking past land use, recent disturbance, and dispersal mechanism to forest composition. – Biol. Conserv. 142: 1647–1656.
bucharová a., MünzberGová z. & tájek P. (2010): Population biology of two rare fern species: long life and long-lasting stability. – Am. J. Bot. 97: 1260–1271.
caswell h. (2001): Matrix population models: construction, analysis, and interpretation. – Sinauer Associates, Sunderland, Massachusetts.

Příroda, Praha, 31: 33–52, 2011
51
clark c. j., Poulsen j. r., levey D. j. & osenberG c. w. (2007): Are plant populations seed limited? A critique and meta-analysis of seed addition experiments. – Am. Nat. 170: 128–142.
ehrlén j., MünzberGová z., DiekMann M. & eriksson o. (2006): Long-term assessment of seed limitation in plants: results from an 11-year experiment. – J. Ecol. 94: 1224–1232.
ellenberG h., weber h. e., Dull r., wirth v., werner w. & Paulissen D. (1992): Zeigerwerte von Pflanzen in Mitteleuropa. – Scr. Geobot. 18: 1–258.
flinn k. M. (2006): Reproductive biology of three fern species may contribute to differential colonization success in post-agricultural forests. – Am. J. Bot. 93: 1289–1294.
forbis t. a. & Doak D. f. (2004): Seedling establishment and life history trade-offs in alpine plants. – Am. J. Bot. 91: 1147–1153.
fottová D. (2003): Trends in sulphur and nitrogen deposition fluxes in the GEOMON network, Czech Republic, between 1994 and 2000. – Water Air Soil Poll. 150: 73–87.
franzen D. & eriksson o. (2003): Patch distribution and dispersal limitation of four plant species in Swedish semi-natural grasslands. – Plant Ecol. 166: 217–225.
Garcia M. b. (2003): Demographic viability of a relict population of the critically endangered plant Borderea chouardii. – Conserv. Biol. 17: 1672–1680.
Garcia M. b., Pico f. X. & ehrlén j. (2008): Life span correlates with population dynamics in perennial herbaceous plants. – Am. J. Bot. 95: 258–262.
Griffith a. b. & forseth i. n. (2005): Population matrix models of Aeschynomene virginica, a rare annual plant: implications for conservation. – Ecol. Appl. 15: 222–233.
Guo Q. f., kato M. & ricklefs r. e. (2003): Life history, diversity and distribution: a study of Japanese pteridophytes. – Ecography 26: 129–138.
hanski i. (1994): A practical model of metapopulation dynamics. – J. Anim. Ecol. 63: 151–162.holDereGGer r. & schneller j. j. (1994): Are small isolated populations of Asplenium septentrionale
variable? – Biol. J. Linn. Soc. 51: 377–385.husbanD b. c. & barrett s. c. h. (1996): A metapopulation perspective in plant population biology. –
J. Ecol. 84: 461–469.chiarucci a., Maccherini s., bonini i. & De DoMinicis v. (1999): Effects of nutrient addition on community
productivity and structure of serpentine vegetation. – Plant Biol. 1: 121–126.chýlová t. & MünzberGová z. (2008): Past land use co-determines the present distribution of dry
grassland plant species. – Preslia 80: 183–198.jonGejans e., De vere n. & De kroon h. (2008): Demographic vulnerability of the clonal and endangered
meadow thistle. – Plant Ecol.198: 225–240.kolb a. & DiekMann M. (2005): Effects of life-history traits on responses of plant species to forest
fragmentation. – Conserv. Biol. 19: 929–938.lanDe r. & scheMske D. w. (1985): The evolution of self-fertilization and inbreeding depression in
plants. 1. Genetic models. – Evolution 39: 24–40.leslie P. h. (1945): On the use of matrices in certain population mathematics. – Biometrika 33:
183–212.linDborG r. & eriksson o. (2004): Historical landscape connectivity affects present plant species
diversity. – Ecology 85: 1840–1845.lovis j. D. & lovis j. v. (1955): Asplenium adulterinum and its probable parents. – Proc. Bot. Soc. Brit.
Isles 1: 389–390.MenGes e. s. (1990): Population viability analysis for an endangered plant. – Conserv. Biol. 4: 52–62.MilDen M., MünzberGová z., herben t. & ehrlén j. (2006): Metapopulation dynamics of a perennial
plant, Succisa pratensis, in an agricultural landscape. – Ecol. Model. 199: 464–475.Moore k. a. & elMenDorf s. c. (2006): Propagule vs. niche limitation: untangling the mechanisms
behind plant species‘ distributions. – Ecol. Lett. 9: 797–804.MünzberGová z. & herben T. (2004): Identification of suitable unoccupied habitats in metapopulation
studies using co-occurrence of species. – Oikos 105: 408–414.MünzberGová z. & herben T. (2005): Seed, dispersal, microsite, habitat and recruitment limitation:
identification of terms and concepts in studies of limitations. – Oecologia 145: 1–8.

52
Bucharová et al.: Dynamika Asplenium adulterinum a A. cuneifolium na lokální a krajinné úrovni
norDin a., strenGboM j., witzell j., nasholM t. & ericson l. (2005): Nitrogen deposition and the biodiversity of boreal forests: implications for the nitrogen critical load. – Ambio 34: 20–24.
oGilvie r. t. & ceska a. (1984): Alpine plants of phytogeographic interest on Northwestern Vancouver-Island. – Can. J. Bot. 62: 2356–2362.
ouborG n. j. (1993): Isolation, population-size and extinction - the classical and metapopulation approaches applied to vascular plants along the dutch rhine-system. – Oikos 66: 298–308.
Peck j. h., Peck c. j. & farrar D. r. (1990): Influences of life-history attributes on formation of local and distant fern populations. – Am. Fern J. 80: 126–142.
PenroD k. a. & MccorMick l. h. (1996): Abundance of viable hay-scented fern spores germinated from hardwood forest soils at various distances from a source. – Am. Fern J. 86: 69–79.
holub j. & Procházka f. (2000): Red list of vascular plants of Czech Republic – 2000. – Preslia 72: 187–230.
Quintana-ascencio r. f. & MenGes e. s. (1996): Inferring metapopulation dynamics from patch-level incidence of Florida scrub plants. – Conserv. Biol. 10: 1210–1219.
r_DeveloPMent_core_teaM (2009): R: A language and environment for statistical computing. – Vienna.reisch c., Mayer f., ruther c. & nelle o. (2007): Forest history affects genetic diversity - molecular
variation of Dryopteris dilatata (Dryopteridaceae) in ancient and recent forests. – Nord. J. Bot. 25: 366–371.
ricotta c., chiarucci a. & avena G. (2004): Quantifying the effects of nutrient addition on community diversity of serpentine vegetation using parametric entropy of type alpha. – Acta Oecol. 25: 61–65.
roMerMann c., tackenberG o., scheuerer M., May r. & PoscholD P. (2007): Predicting habitat distribution and frequency from plant species co-occurrence data. – J. Biogeogr. 34: 1041–1052.
scheMske D. w., husbanD b. c., ruckelshaus M. h., GooDwillie c., Parker i. M. & bishoP j. G. (1994): Evaluating approaches to the conservation of rare and endangered plants. – Ecology 75: 584–606.
silvertown j., franco M., Pisanty i. & MenDoza a. (1993): Comparative plant demography - relative importance of life-cycle components to the finite rate of increase in woody and herbaceous perennials. – J. Ecol. 81: 233–244.
soltis D. e. & soltis P. s. (1992): The distribution of selfing rates in homosporous ferns. – Am. J. Bot. 79: 97–100.
staPulionyte a., lazutka j. r., naujalis j. r. & oDlanD a. (2006): Assessment of genetic polymorphism in natural populations of ostrich fern (Matteuccia struthiopteris L., Dryopteridaceae) in Lithuania by using RAPD markers. – Biologija 2006/1: 53–59
ŠPryňar P. & schläGelová j. (2005): Jarní botanicko-geologická vycházka za květenou nevápencových hornin Českého krasu. – Český kras 31: 56–60.
tájek P. (2003): Hadcové výchozy ve Slavkovském lese z pohledu ostrovní biogeografie. – Ms. [Dipl. pr.; depon. in: Knihovna katedry botaniky PřF UK, Praha.].
tájek P., bucharová a. & MünzberGová z. (2011): Limitation of distribution of two rare ferns in fragmented landscape. – Acta Oecol. 37: 495–502.
treMlová k. & MünzberGová z. (2007): Importance of species traits for species distribution in fragmented landscapes. – Ecology 88: 965–977.
tryon r. (1970): Development and evolution of fern floras of oceanic islands. – Biotropica 2: 76–84.tryon r. (1986): The biogeography of species, with special reference to ferns. – Bot. Rev. 52: 117–156.turitzin s. n. (1982): Nutrient limitations to plant-growth in a California serpentine grassland. –
American Midland Naturalist 107: 95–99.turnbull l. a., crawley M. j. & rees M. (2000): Are plant populations seed-limited? A review of seed
sowing experiments. – Oikos 88: 225–238.voGel j. c., ruMsey f. j., schneller j. j., barrett j. a. & Gibby M. (1999): Where are the glacial refugia
in Europe? Evidence from pteridophytes. – Biol. J. Linn. Soc. 66: 23– 37.wilD M. & GaGnon D. (2005): Does lack of available suitable habitat explain the patchy distributions
of rare calcicole fern species? – Ecography 28: 191–196.viane r., jerMy a. c. & lovis j. D. (1993): Asplenium. – In: tutin t. G., burGes n. a., chater a. o.,
eDMonDson j. r., heywooD v. h., Moore D. M., valentine D. h., walters s. M. & webb D. a., Flora Europaea, Vol. 1, 2nd ed., pp. 18–23, Cambridge University Press.

53
Příroda, Praha, 31: 53–83, 2011
Populační dynamika a genetická diverzita kriticky ohroženého druhu Dracocephalum austriacum
Population dynamics and genetic diversity of critically endangered Dracocephalum austriacum
toMáš dostáLek1, 2 & ZuZana MünZbergová1, 2
1Botanický ústav Akademie věd ČR, Zámek 1, CZ-252 43 Průhonice; [email protected] botaniky, Přírodovědecká fakulta Univerzity Karlovy, Benátská 2, CZ-128 01 Praha 2
Abstract: The aim of our study was to compare the population dynamics and distribution of genetic diversity in the populations of critically endangered species Dracocephalum austriacum L. in two distant regions (Bohemian and Slovak Karst). We used transition matrix models to examine the population dynamics of the species and computed population growth rates, elasticities, stochastic population growth rates and extinction probabilities. The genetic diversity, its importance for plant fitness, and selection of the most valuable populations for conservation of genetic diversity within the species in the studied regions were assessed performing allozyme variation of 12 populations. Furthermore we wanted to understand the effects of changes in the genetic diversity on the population dynamics. Using the relationship between the expected heterozygosity and seed production, we estimated the effects of changes in heterozygosity on the long-term population growth rate and extinction probabilities of the populations.
The results showed that the populations in the Slovak Karst are overall doing better than populations in the Bohemian Karst. The transitions most contributing to the population growth rate were transitions of stasis in the stage of seeds and in stages of small and large adult plants. These transitions are comparable between the regions and we thus suggest that the knowledge of the most important life cycle transitions can be transferred between populations in different regions. The results of genetic diversity analyses showed high genetic diversity within populations (80.14%) and relatively low differentiation among populations within regions (9.42%) and between regions (10.45%). Seed production was significantly higher in larger, genetically more diverse and less inbred populations. The results also show large variation in genetic diversity between populations and indicate which populations should get a priority in attempts to conserve all the genetic diversity within the region. We also determined a strong effect of reduced expected heterozygosity on a decrease in seed production. However, only severe reduced heterozygosity caused significant decrease in the long term population growth rate. Reduction in heterozygosity increased extinction probability of the populations but this was visible only in a simulation based on a 100-year period due to a long life span of the species. The effects of reduction in heterozygosity were consistent between the two karst regions but they were stronger in the Slovak Karst.Keywords: Natura 2000, perennial herb, small population size
Abstrakt: Cílem naší práce bylo porovnat populační dynamiku a rozložení genetické diverzity v populacích kriticky ohroženého druhu, včelníku rakouského (Dracocephalum austriacum L.), ve dvou vzdálených územích (Český a Slovenský kras). Populační dynamiku jsme analyzovali s použitím maticových modelů, s jejichž pomocí jsme spočítali populační růstové rychlosti, elasticity, stochastické populační růstové rychlosti a pravděpodobnosti

54
Dostálek & Münzbergová: Populační dynamika a genetická diverzita Dracocephalum austriacum
vymření populací D. austriacum. Rozložení genetické diverzity, její význam pro fitness a výběr nejcennějších populací pro ochranu genetické diverzity v rámci sledovaných území byly studovány pomocí analýzy variability izoenzymů ve 12 populacích D. austriacum. Navíc jsme se snažili porozumět vlivu změn genetické diverzity na populační dynamiku druhu. Pomocí vztahu mezi očekávanou heterozygozitou a produkcí semen jsme odhadovali vliv změn heterozygozity na dlouhodobou populační růstovou rychlost a pravděpodobnosti vymření populací.
Výsledky ukazují, že populace ve Slovenském krasu mají vyšší populační růstovou rychlost a prosperují tak lépe než populace z Českého krasu. Přechody, které nejvíce ovlivňují populační růstovou rychlost, jsou přechody přežívání v životní fázi semen a přežívání v životních fázích malých a velkých dospělých rostlin. Tyto přechody jsou srovnatelné mezi studovanými územími, a proto předpokládáme, že znalosti o významných fázích životního cyklu mohou být využity i v dalších populacích druhu ze vzdálených územích. Výsledky analýzy genetické diverzity ukázaly vysokou míru genetické diverzity v rámci populací (80,14 %) a relativně nízkou odlišnost mezi populacemi v rámci regionu (9,42 %) a mezi regiony (10,45 %). Zaznamenali jsme také průkazně vyšší produkci semen ve větších, geneticky variabilnějších a méně inbredních populacích. Naše analýza také ukazuje na velkou variabilitu genetické diverzity mezi populacemi. V této práci jsme se snažili proto navrhnout, které populace by měly být upřednostněny v případě, že chceme zachovat maximální míru genetické diverzity v daném území. Objevili jsme také silný vliv snížení očekávané heterozygozity na produkci semen D. austriacum. Avšak pouze velký pokles heterozygozity má za následek průkazné snížení dlouhodobé populační růstové rychlosti. Snížení heterozygozity také průkazně zvýšilo pravděpodobnost vymření jednotlivých populací, ale tento vliv je více patrný až při simulacích přes 100 let kvůli dlouhému životu studovaného druhu. Vlivy snížení heterozygozity byly podobné v obou krasových územích, ale silnější trendy jsme pozorovali ve Slovenském krasu.Klíčová slova: malá velikost populace, Natura 2000, vytrvalá bylina
Úvod
V rámci procesu sjednocování Evropy a přistupování České republiky k evropským legislativním požadavkům je nutné zabývat se také sbližováním postupů v ochraně přírody. V rámci evropského konceptu NATURA 2000 je stanoven promyšlený ucelený přístup řešící ochranu stanovišť i jednotlivých ohrožených taxonů (euroPean coMMission 1992). Přibližně v roce 2000 byl započat projekt zajišťující bazální monitoring těchto vybraných druhů v České republice. V rámci tohoto monitoringu je sčítán počet kvetoucích (někdy i sterilních) jedinců na lokalitách a jsou zaznamenávány základní charakteristiky prostředí těchto lokalit (http://biodiversity.eionet.europa.eu/activities/Natura_2000/pdfs/MONITORING.pdf). Tyto informace umožní identifikovat trendy ve vývoji populací, neumožní však identifikovat faktory za tyto změny zodpovědné. V případě výrazného poklesu velikosti populace tak není jasné, která část životního cyklu druhu byla dotčena a zaslouží si tedy zvýšenou pozornost při další snaze o ochranu druhu. K plánování účinných ochranářských opatření potřebujeme získat informace o populační biologii, genetické diverzitě i ekologii těchto vzácných druhů a získat tak podrobnější náhled na mechanismy zodpovědné za změny v jejich populačních velikostech. Přestože v poslední době vzrůstá počet studií, které se zabývají těmito různými aspekty (např. GléMin et al. 2005, lóPez-Pujol et al. 2007, Pino et al. 2007, colas et al. 2008), máme stále velice málo studií spojujících tyto aspekty do celkového obrazu. Častou nevýhodou většiny těchto prací je však to, že probíhají pouze na jednom omezeném území, často dokonce jen na jedné populaci druhu. Není proto známo, jaká je míra variability demografie druhu

55
Příroda, Praha, 31: 53–83, 2011
na větším území a tedy nakolik je tyto závěry možno přenášet mezi různými populacemi či dokonce územími.
Ve snaze získat podrobnější náhled na mechanismy zodpovědné za změny v po-pulační velikosti vzácných druhů se v současné době objevilo několik prací snažících se o podrobnější sledování dynamiky jejich populací (např. van buren & harPer 2003, Pino et al. 2007, eckstein et al. 2009, bucharová et al. 2010). Výsledky těchto studií umožňují identifikovat kritické fáze životního cyklu a tedy rozhodnout, která fáze cyklu má být přednostně podporována v případě poklesu velikosti populace druhu.
Znalost demografie druhu ale není sama o sobě schopna poskytnout kompletní informaci potřebnou k efektivní ochraně, neboť důležitým aspektem při ochraně vzácných druhů je zachování nejen dostatečného počtu jedinců, ale i jejich genetické diverzity (lóPez-Pujol et al. 2003, oosterMeijer et al. 2003). Data o genetické diverzitě však v Evropě chybí i pro mnoho prioritních druhů (euroPean coMMission 1992), které jsou považovány za druhy nejvíce celoevropsky ohrožené (výjimkami jsou práce colas et al. 1997, GauDeul et al. 2000, brzosko et al. 2002, GléMin et al. 2005). Význam studia genetické diverzity je také podpořen tím, že populace vzácných a ohrožených druhů jsou často malé a izolované. Genetická diverzita v takovýchto populacích je ohrožena náhodnými změnami v alelických frekvencích, tzv. genetickým driftem. V extrémním případě to může vést až ke ztrátě alel z populace, a tak ke ztrátě polymorfismu, tj. lokus obsahuje pouze jedinou variantu alel (lowe et al. 2004). V tom případě pak populace ztrácí schopnost se adaptovat na měnící se podmínky prostředí a stává se náchylnější k vymření i po menších změnách okolního prostředí (newMan & Pilson 1997, frankhaM & ralls 1998, saccheri et al. 1998). Podobný pokles genetické diverzity může být dále způsoben zvýšeným křížením blízce příbuzných jedinců, což má za následek zvýšení homozygozity (lowe et al. 2004). Křížení mezi blízce příbuznými jedinci může také vést ke snížení fitness populace, tzv. inbrední depresi (De Mauro 1993, anDerson & walDMann 2002, ishihaMa et al. 2005). Dva hlavní následky inbrední deprese jsou 1) větší pravděpodobnost projevení škodlivých alel a 2) snížená schopnost přizpůsobit se měnícím se podmínkám prostředí (reeD & frankhaM 2003). Genetický drift i inbreeding tak mohou vést ke snížení fitness rostlin. Pokud se nám ale podaří zjistit vztah mezi genetickou diverzitou a fitness rostlin, tak nám včasné odhalení výše zmíněných negativních vlivů může velice pomoci snížit riziko vymření populace vzácného druhu.
I přes možný významný vliv genetické diverzity na fitness rostlin oosterMeijer et al. (2003) zjistili, že mezi prácemi zabývajícími se ochranářskou biologií publikovanými v letech 1979 až 2000 je pouze několik prací, které zkoumají vztah demografie a genetické diverzity rostlin. Studie, které se zabývají demografií i genetickou diverzitou, nám mohou pomoci vytvořit mnohem jasnější představu o budoucím osudu populací vzácných druhů (oosterMeijer 2000, Picó et al. 2004, kirchner et al. 2006).
V této práci srovnáváme populační dynamiku kriticky ohroženého druhu Dracocephalum austriacum L. (včelník rakouský) ve dvou vzdálených územích. Také jsme se zabývali rozložením genetické diverzity v populacích tohoto druhu a pokusili se zhodnotit význam genetické diverzity pro jeho populační dynamiku. V práci jsme si kladli následující otázky: 1) Jaké jsou kritické fáze životního cyklu? 2) Jaké je rozložení genetické diverzity mezi jednotlivými populacemi a územími? 3) Jaký je vliv změn genetické diverzity na populační dynamiku druhu? 4) Liší se populační dynamika, rozložení genetické diverzity a vliv změn genetické diverzity na populační dynamiku mezi dvěmi vzdálenými územími?
Abychom mohli odpovědět na tyto otázky, studovali jsme kompletní životní cyklus druhu D. austriacum ve třech populacích v Českém krasu a třech populacích ve Slovenském krasu v letech 2003–2006. Dále jsme pomocí analýzy izoenzymů odhadli genetickou

56
Dostálek & Münzbergová: Populační dynamika a genetická diverzita Dracocephalum austriacum
diverzitu ve všech populacích v ČR a ve třech populacích na Slovensku. Pomocí vztahu mezi genetickou diverzitou a produkcí semen jsme nakonec sledovali vliv změn genetické diverzity na populační dynamiku druhu.
Metodika
studovaný druh
Dracocephalum austriacum L. (včelník rakouský, Lamiaceae) je vytrvalá bylina se vzpřímenými nebo vystoupavými lodyhami až 60 cm vysokými. Druh je diploid (2n = 14) (heywooD 1980). Kvete od druhé poloviny května do poloviny června. V České a Slovenské republice roste na skalních stepích a kamenitých výslunných svazích většinou na vápenci (Čeřovský 1999). Druh je entomogamní (hrouDa 2000) a podle předběžných výsledků je schopen i samoopylení, přestože produkuje mnohem více semen při cizosprášení (Dostálek 2005, Castro et al., nepublikováno).
V České republice patří tento druh do kategorie kriticky ohrožených (holub & Procházka 2000) a je také na seznamu celoevropsky ohrožených druhů (euroPean coMMission 1992).
Celkový areál druhu je tvořen jednotlivými arelami a zasahuje od východních Pyrenejí, přes Francii, Itálii, Švýcarsko, Rakousko, Českou republiku (severní okraj areálu), Slovensko, Maďarsko, Rumunsko až na Ukrajinu, nejvýchodněji až do Předkavkazí (Meusel et al. 1978).
obr. 1. Mapa lokalit D. austriacum v Českém krasu.Fig. 1. Map of D. austriacum localities in the Bohemian Karst.
Historická data o výskytu D. austriacum v České republice a na Slovensku nejsou moc podrobná. heufler (1851) píše o prvním nálezu D. austriacum na území dnešní ČR v roce 1792. nevole (1910) a valouŠek (1928) uvádějí ve svých pracích seznam lokalit

57
Příroda, Praha, 31: 53–83, 2011
v Českém krasu a na Moravě. nevole (1910) také zmiňuje lokalitu v Českém středohoří na vrchu Deblík. Tato lokalita však už nebyla potvrzena od roku 1996 (haMerský 2000). Moucha (1990) uvádí osm populací v Českém krasu na místech, kde jsou populace D. austriacum dodnes (viz obr. 1). Dodává také, že některé historické populace byly zničeny těžbou. Čeřovský (1999) také udává devět populací v České republice a zmiňuje se o třech dalších populacích, které už vymřely. V České republice tedy bylo v minulosti minimálně 12 populací D. austriacum. Na Slovensku jsou data o historickém rozšíření druhu ještě stručnější. Několikrát se objevuje zmínka o populacích na území Slovenského krasu a Slovenského ráje (Moucha 1990, Čeřovský 1999). Populace D. austriacum ve Slovenském krasu v posledních 50 letech početně klesly a některé z nich úplně zmizely. V současné době je na území Slovenského krasu minimálně šest populací (E. Karasová, osobní sdělení).
D. austriacum se doposud více zabývalo i několik dalších studií. Moucha (1986) se zabýval metodou množení D. austriacum a možnostmi kultivace pro jeho ochranu ex-situ nebo posilování menších populací. Zjistil, že D. austriacum jde poměrně snadno pěstovat jak ze semen, tak řízkováním. kubíková (1993) sledovala počet rostlin v populaci D. austriacum v Radotínském údolí. V Maďarsku se druhem zabývali zejména virok (2006) a Puska (2006). Ve Francii se demografií a genetikou tohoto druhu zabývala ve své disertační práci nicolè (2005). bonin et al. (2007) vytvořili nový způsob pro výběr populací, které zahrnují maximum genetické diverzity. nicolè et al. (2011) publikovali studii, kde ukázali rozdílný vliv klimatu na populační dynamiku druhu v různých podmínkách prostředí. Zjistili, že populace na strmějších svazích mají nižší pravděpodobnosti přežívání a nižší populační růstové rychlosti než populace na pozvolnějších svazích. Přežívání a populační růstová rychlost byla také nižší v letech s nízkými jarními a vysokými letními teplotami. Navíc negativní vliv vysokých letních teplot byl do značné míry závislý na podmínkách prostředí, kdy byl silnější na strmějších svazích.
studované populace
V této práci jsme studovali všech devět současných populací D. austriacum v ČR a tři největší populace na Slovensku (tab. 1). Osm českých populací je na území CHKO Český kras (jednotlivé populace vzdáleny max. 20 km, viz obr. 1) a devátá populace je v PR Zázmoníky na jižní Moravě. Studované slovenské populace jsou všechny ze Slovenského krasu (vzdáleny od sebe max. 30 km). Vzdálenost mezi populacemi v Českém a Slovenském krasu je ca 500 km. Populace v Zázmoníkách je vzdálena zhruba 200 km od Českého a 300 km od Slovenského krasu. Populace jsou umístěny ve třech geograficky vymezených územích (Český kras - C1–C8, Morava - M1, Slovenský kras - S1–S3). Lokality D. austriacum v Českém i ve Slovenském krasu jsou na podobných stanovištích. V obou územích rostliny D. austriacum rostou na vápencových svazích, kde kompetují s křovinami a trávami. Lokality ve Slovenském krasu jsou však na hlubších půdách s vyšším obsahem oxidovatelného uhlíku, nižším pH a nižším obsahem celkového dusíku (nebyl ale zjištěn průkazný rozdíl v podílu C:N, Dostálek 2005). Obě krasová území se také liší druhovým složením na lokalitách. Druhy, které lokality nejvíce odlišují, jsou Thesium linophyllon, Festuca rupicola, Fragaria viridis a Vincetoxicum hirundinaria, které se vyskytují více ve Slovenském krasu, a Galium glaucum, Sedum sexangulare, Stachys recta a Stipa pennata vyskytující se častěji v Českém krasu (Dostálek 2005). Lokalita Zázmoníky v moravském regionu se liší od obou krasových území. Rostliny zde rostou na spraši, která je mnohem hlubší než půda na vápencových skalách a obsahuje méně oxidovatelného uhlíku a celkového dusíku.
58
Dostálek & Münzbergová: Populační dynamika a genetická diverzita Dracocephalum austriacum
tab.
1. P
očty
vzo
rků
a úd
aje
o m
írách
gen
etic
ké d
iver
zity
pro
des
et lo
kusů
ve
12 s
ledo
vaný
ch p
opul
acíc
h D
raco
ceph
alum
aus
triac
um.
tab.
1. S
ampl
e si
zes
and
gene
tic d
iver
sity
sta
tistic
s fo
r ten
loci
in 1
2 po
pula
tions
of D
raco
ceph
alum
aus
triac
um.
Pop
ulac
eN
TAP
AP
HE
HO
F ST
F ISU
Prů
m.
pořa
díVe
l pop
PK
PV
Čes
ký k
ras
142
210,
62,
830,
119
0,06
20,
143
0,34
03
1787
--
C1
- Cís
ařsk
á ro
kle
2214
50,
332,
336
0,05
5110,
0466
0,21
320,
1684
024,
921
115
10,
139
C2
- Hak
nove
c22
163
0,42
2,50
40,
1286
0,11
430,
1519
0,10
3202
3,9
640
500
0,13
3
C3
– K
arlic
ké ú
dolí
713
60,
242,
504
0,07
990,
0298
0,17
760,
63811
026,
37
60,
023
C4
- Kod
ská
stěn
a22
163
0,51
2,20
70,
1325
0,08
240,
1776
0,37
8602
4,3
181
130
0,18
8
C5
- Koz
elsk
á ro
kle*
310
70,
05-
0,00
0120,
0009
0,41
41-
026
43
0,21
3
C6
- Rad
otín
ské
údol
í22
163
0,42
2,50
40,
1107
0,04
170,
14910
0,63
01002
5,6
6031
0,05
9
C7
– Va
novi
ce*
2217
20,
512,
405
0,07
5100,
0417
0,15
880,
4569
025,
554
330,
118
C8
- Vel
ká h
ora
2219
10,
423,
252
0,10
480,
0645
0,12
2110,
3847
024,
863
051
50,
322
Mor
ava
(M1
- Záz
mon
íky)
415
40,
243,
501
0,13
640,
1252
0,17
450,
0791
022,
94
2-
Slo
vens
ký k
ras
6620
0,4
3,50
0,18
90,
123
0,08
80,
347
345
0-
-
S1
- Zád
iels
ký k
ameň
2217
20,
422,
753
0,18
310,
1124
0,18
340,
3858
023,
310
053
0,17
5
S2
- Dom
ické
škr
apy
2216
30,
422,
504
0,17
120,
1143
0,18
730,
3365
112,
920
016
60,
289
S3
- Žel
ezná
vra
ta22
172
0,42
2,75
30,
1673
0,14
110,
1657
0,15
4311
2,8
150
670,
278

59
Příroda, Praha, 31: 53–83, 2011
Seřadili jsme všechny populace podle hodnot jejich TA, P, AP, HE and HO, FST, FIS a podle počtu unikátních alel. Populace s celkovým nejnižším průměrným pořadím byly považovány za ty, které jsou nejhodnotnější z hlediska zachování celkové genetické diverzity v rámci každého regionu (neel & ellstranD 2003). Tři nejlepší hodnoty každé proměnné jsou tučně zvýrazněny. (N = počet odebraných vzorků, TA = celkový počet alel, P = podíl polymorfních lokusů, AP = počet alel na polymorfní lokus, HE = očekávaná heterozygozita, HO = pozorovaná heterozygozita, FST = průměrná míra genetické diferenciace od ostatních populací, FIS = koeficient inbreedingu, U = počet unikátních alel, Vel pop = velikost populace v roce 2005, PK = počet kvetoucích rostlin, PV = podíl vyvinutých semen).* populace, které byly znovuobnoveny vysazením v osmdesátých letech 20. století byly vyloučeny z některých analýz (pro detaily viz Metodika).We ranked the populations TA, P, AP, HE and HO, FST, FIS, and all occurrences of unique alleles. Populations with the lowest mean rank were considered to have higher priority for conservation of genetic diversity within the species in each region (neel & ellstranD 2003). Three best values for each variable are in bold. (N = number of samples, TA = total number of alleles, P = proportion of polymorphic loci, AP = number of alleles per polymorphic locus, HE = expected heterozygosity, HO = observed heterozygosity, FST = mean genetic distance to other populations, FIS = inbreeding coefficient, U = number of unique alleles, Vel pop = population size in 2005, PK = number of flowering plants, PV = proportion of developed seeds).
Populační dynamika
Data o jednotlivých přechodech mezi fázemi životního cyklu
Pro sledování demografických parametrů byly vybrány tři populace v CHKO Český kras (Císařská rokle - C1, Haknovec - C2, Kodská stěna - C4) a tři nejpočetnější populace v NP Slovenský kras (Zadielský kameň - S1, Domické škrapy - S2, Železná vrata - S3). Byly vybrány populace s nejméně 100 jedinci, abychom mohli získat dobrý odhad demografických parametrů. Sledováni byli většinou všichni jedinci na lokalitách (s výjimkou rostlin na těžko přístupných strmých místech). Rostliny byly značeny a sledovány po čtyři roky (2003–2006) a po jejich odkvetení (červenec) u nich byl zaznamenáván počet kvetoucích a nekvetoucích lodyh. Produkce semen byla každý rok sledována na vzorku 20 náhodně vybraných kvetoucích rostlin. Byl sčítán počet černých tvrdých semen u každé vybrané rostliny. Získaná semena byla po spočítání ponechána na lokalitě.
Rostliny byly rozděleny do čtyř fází, ze kterých se skládá životní cyklus D. austriacum: semena, semenáčky, malé dospělé rostliny a velké dospělé rostliny. Semenáčky jsou rostliny pouze s jednou nekvetoucí tenkou lodyhou, nejvýše deset cm vysokou. Jsou to rostliny, které vyklíčily v roce, kdy byly poprvé zaznamenány nebo v roce předcházejícím. Žádný semenáček nemůže být starší než dva roky. Za dva roky semenáček vyroste do fáze malé dospělé rostliny, která může už vykvést. Malé dospělé rostliny jsou rostliny s jednou lodyhou větší než deset cm nebo se dvěmi až pěti lodyhami. Velké dospělé rostliny mají šest a více lodyh. Nenašli jsme žádný vztah mezi počtem lodyh a pravděpodobností kvetení rostliny, a tak byly malé a velké dospělé rostliny rozlišeny pouze podle počtu lodyh. Počet lodyh pro rozdělení mezi malé a velké dospělé rostliny byl stanoven tak, aby byl v obou kategoriích dostatečně velký počet rostlin a aby měly rostliny v obou kategoriích podobné biologické vlastnosti. U rostlin se šesti a více lodyhami jsme nalezli průkazně vyšší pravděpodobnosti přežívání a kvetení v druhém roce přechodu. V letech, kdy jsou pro druh špatné podmínky, nebo při stárnutí může docházet k přechodu z fáze velkých dospělých rostlin do fáze malých dospělých rostlin. Tento přechod nazýváme retrogrese.
Semena D. austriacum mají tvrdý obal a očekáváme, že mohou přežívat v půdě po mnoho let. Abychom mohli odhadnout, jak dlouho jsou semena schopná v půdě přežívat,

60
Dostálek & Münzbergová: Populační dynamika a genetická diverzita Dracocephalum austriacum
udělali jsme následující pokus na podzim 2006. Na každé ze tří sledovaných lokalit v Českém krasu jsme na transektech přes nejdelší průměr lokality odebrali 30–35 vzorků půdy (100 cm3, dva metry od sebe). Vzorky byly odebírány z povrchu půdy do hloubky pěti cm. V každém půdním vzorku jsme pak spočetli živá semena a jejich počet použili k výpočtu všech živých semen na lokalitě. Za živá semena byla považována ta, která měla pevné a bílé embryo. Že jsou tato semena opravdu živá, bylo potvrzeno také tetrazoliovým barvícím testem (Delouche et al. 1962). Přežívání v semenné bance pak bylo počítáno jako Si = 1 - ai/bi; kde Si – pravděpodobnost přežití do dalšího roku v semenné bance na lokalitě i, ai – průměrný počet živých semen vyprodukovaných za rok na lokalitě (průměrná data z let 2003–2006), bi – počet všech živých semen nalezených v půdě na lokalitě i. Tento pokus byl proveden pouze na lokalitách v Českém krasu. Pro slovenské populace jsme použili průměrnou hodnotu přežívání v semenné bance ze všech tří českých populací. Abychom pak mohli odhadnout klíčení ze semenné banky, potřebovali jsme znát ještě počet živých semen v semenné bance i ve slovenských populacích. To jsme spočítali pomocí dat o průměrném přežívání v semenné bance v populacích v Českém krasu a dat o průměrném počtu vyprodukovaných semen v populacích ve Slovenském krasu.
Abychom mohli odhadnout klíčení semen, počítali jsme každý rok všechny nové semenáčky na lokalitě (na většině lokalit na úplně celé lokalitě). Klíčení semen pak bylo vypočítáno jako podíl počtu nově nalezených semenáčků a všech živých semen na lokalitě (odhadnuto z experimentu popsaném výše).
Modelování populační dynamiky
Demografická data byla analyzována pomocí přechodových maticových modelů (caswell 2001). Pomocí analýzy přechodových matic můžeme vypočítat populační růstovou rychlost (λ). Z projekce přechodových matic také můžeme získat informaci o změně populační růstové rychlosti, δλ, která je následkem malé změny jednoho z přechodů aij, tzv. sensitivitě. Poměrná sensitivita, tzv. elasticita, je obvykle používána jako míra, jakou přispívá určitý prvek v matici k fitness (De kroon et al. 2000). Pravý vlastní vektor matice pak koresponduje se stabilním rozložením fází životního cyklu v populaci (caswell 2001).
V této práci jsme spočítali populační růstovou rychlost pro každou populaci a každý přechodový interval. Pomocí stochastických simulací (caswell 2001) jsme navíc kombinací všech matic jednoho typu (rok, populace, region) získali odhad celkové populační růstové rychlosti pro každý tento typ. Podobně jsme získali pro každý typ také hodnoty elasticity a stabilního rozložení fází životního cyklu (detaily viz Dostálek & MünzberGová 2009).
Dále jsme spočítali 95 % intervaly spolehlivosti pro všechny výše popsané charakteristiky populací (alvarez-buylla & slatin 1994). Populační růstové rychlosti a elasticity byly pova žovány za průkazně odlišné, pokud nedocházelo k překryvu jejich 95 % intervalů spolehlivosti. Průkaznost rozdílů populační růstové rychlosti mezi Českým a Slovenským krasem jsme navíc počítali ještě permutačním testem (detaily viz Dostálek & MünzberGová 2009).
Dále jsme počítali pravděpodobnosti vymření českých a slovenských populací tak, že jsme násobili populační vektor každé populace vždy jednou z náhodně vybraných matic z této populace. Populaci jsme považovali za vymřelou, pokud v ní už nebyly žád né semenáčky, ani malé nebo velké dospělé rostliny (tj. stále tam mohla být semena v se-menné bance). Pravděpodobnost vymření byla spočítána jako podíl vymřelých populací z 500 opa kování. Pravděpodobnost vymření byla počítána do 20, 50 a 100 let.
Pravděpodobnost vymření populace jsme počítali rovněž pro modelové populace z Českého a Slovenského krasu, abychom mohli obě území porovnat. Jako populační

61
Příroda, Praha, 31: 53–83, 2011
vektor jsme použili 10 000, 5 000 a 1 000 jedinců, kteří byly rozloženi do jednotlivých fází životního cyklu tak, aby to odpovídalo průměrnému stabilnímu rozložení fází životního cyklu, které bylo spočítáno z matic u obou regionů. Vysoké počty jedinců byly použity proto, že 99,1 % všech jedinců tvoří semena v semenné bance. (Např. 10 000 jedinců odpovídá 10 semenáčkům, 42 malým a 40 velkým dospělým rostlinám.) Pro výpočet pravděpodobností vymření modelových populací z Českého a Slovenského krasu jsme použili všechny dostupné matice z populací z daného regionu.
Některé populace D. austriacum jsou ohrožovány vykopáváním rostlin zahrádkáři. Tento problém je zejména na území Slovenského krasu (E. Karasová, osobní sdělení), ale z tohoto důvodu zanikla pravděpodobně i jediná lokalita v Českém středohoří na vrchu Deblík (R. Hamerský, osobní sdělení). Proto jsme počítali také pravděpodobnosti vymření populací, kde zahrádkáři každý rok vykopou jednu, tři, pět nebo deset velkých dospělých rostlin. Počet vykopávaných rostlin nebyl vždy stejný, ale byl nahrazen hodnotou odvozenou z Poissonova rozdělení s daným průměrem. Takto bylo vykopávání rostlin částečně náhodné.
Všechny analýzy byly provedeny v programu Matlab verze 7.3.0.267 (The MathWorks, Inc., Natick, Massachusetts, USA).
Analýza genetické diverzity
Genetická diverzita populací D. austriacum byla stanovena pomocí analýzy izoenzymů. Izoenzymy jsou považovány za neutrální markery (soltis & soltis 1989, skibinski et al. 1993) a byly opakovaně použity ke stanovování genetické diverzity u vzácných druhů (např. Petit et al. 1998, neel & ellstranD 2003, neel & cuMMinGs 2003, GoDt et al. 2005). Některé studie nesouhlasí s použitím genetické diverzity stanovené pomocí analýzy izoenzymů nebo jiných neutrálních markerů u vzácných druhů pro potřeby ochrany jejich populací a poukazují na možnou slabou korelaci mezi genetickou variabilitou a adaptivní genotypickou variabilitou, která je důležitá pro přežití druhu (např. haMrick et al. 1991, storfer 1996). Na druhou stranu neutrální genetické markery nám umožňují zjistit potenciální variabilitu, která nemusí mít adaptivní výhodu teď, ale může ji získat později, když dojde ke změně podmínek prostředí.
Izoenzymová analýza byla provedena celkem u 212 rostlin D. austriacum z 12 po-pu lací ze třech geografických regionů (Český kras a Morava v ČR, Slovenský kras na Slovensku). V každé populaci byly odebrány vzorky listů z 22 rostlin (v menších populacích byly odebrány vzorky ze všech rostlin, tab. 1). Rostliny byly v populaci vybrány náhodně (z výběru byly vyloučeny pouze semenáčky, tj. rostliny pouze s jednou lodyhou nejvýše deset cm vysokou). Rozhodli jsme se neodebírat vzorky ze semenáčků, protože ty mají obvykle vyšší genetickou diverzitu než dospělé rostliny (ManDák et al. 2006, honnay et al. 2008) a kdybychom brali vzorky v nestejném poměru mezi semenáčky a dospělými rostlinami, mohli bychom získat zavádějící odhad genetické diverzity. Navíc by odběrem mohlo dojít k poškození semenáčků.
Celkem bylo analyzováno deset enzymatických systémů (LAP, SOD, AAT, 6-PGDH, ADH, SKDH, PGM, ME, EST, IDH). Pro odhad genetické diverzity byly použity pouze čtyři enzymatické systémy (deset lokusů), které vykazovaly variabilitu a umožňovaly alelické vyhodnocení – LAP (tři), SOD (tři), 6-PGDH (dva), AAT (dva). Detaily ohledně analýz jsou v práci Dostálek et al. (2010).
Spočítali jsme celkový počet alel (TA), podíl polymorfních lokusů (P), počet alel na jeden polymorfní lokus (AP), pozorovanou (HO) a očekávanou heterozygozitu (HE) pro každý region (Český kras, Morava, Slovenský kras) a každou populaci v programu Popgene 1.32

62
Dostálek & Münzbergová: Populační dynamika a genetická diverzita Dracocephalum austriacum
(yeh & boyle 1997). Koeficient inbreedingu (FIS) byl spočítán pro každou populaci jako FIS = (HE – HO)/HE, kde HE je průměrná očekávaná heterozygozita populace a HO je průměrná pozorovaná heterozygozita populace. Pomocí programu Popgene 1.32 jsme dále spočítali i míru genetické diferenciace populací (FST) v Českém a Slovenském krasu. Program ARLEQUIN 3.11 (eXcoffier et al. 2005) byl použit k určení rozložení genetické variability v rámci populací, mezi populacemi a mezi regiony pomocí analýzy molekulární variance (AMOVA, eXcoffier et al. 2005). Populace v analýze molekulární variance byly seskupeny podle jejich geografické blízkosti do dvou skupin – Český kras a Slovenský kras. Moravský region byl z analýzy vyloučen, protože obsahoval pouze jednu populaci se čtyřmi rostlinami. V programu ARLEQUIN 3.11 jsme dále také spočítali míru genetické diferenciace všech párů jednotlivých populací (FST).
Zjišťovali jsme také počet unikátních alel (alely nalezené pouze v jedné populaci či regionu). Pomocí Mantelova testu (Mantel 1967) jsme testovali, zda je matice geografických vzdáleností korelována s maticí genetických vzdáleností (ARLEQUIN 3.11, eXcoffier et al. 2005, 10 000 permutací). Korelace genetických a geografických vzdáleností byla rovněž spočítána samostatně pro populace v Českém a ve Slovenském krasu.
Testovali jsme také korelaci mezi mírami genetické diverzity a různými mírami fitness (lineární regrese, S-PLUS 2000). Fitness rostlin byla vyjádřena jako počet vyprodukovaných semen na kvetoucí lodyhu, počet vyprodukovaných semen na kvetoucí rostlinu a podíl vyvinutých semen z celkového počtu semen, které mohou být v jednom květu vyprodukovány. Abychom mohli odhadnout podíl vyvinutých semen, počítali jsme na stejném vzorku 20 rostlin na populaci, u kterých byl počítán také počet vyvinutých semen, také počet kalichů na kvetoucí lodyhu. Každý kalich může u tohoto druhu obsahovat až čtyři vyvinutá semena, a tak je v ideálním případě poměr semen a kalichů 4:1. Fitness celé populace byla vyjádřena jako logaritmus populační velikosti (celkový počet rostlin na lokalitě) a podíl kvetoucích rostlin v populaci. Tyto míry fitness byly zaznamenávány v roce 2005.
Lokality C5 a C7 z Českého krasu byly vyloučeny z analýzy molekulární variance, Mantelova testu, z počítání celkového FST a FST počítaného samostatně pro Český kras a z počítání korelací mezi mírami genetické diverzity a fitness, protože byly nově vysazeny v osmdesátých letech 20. století (ložek 2003).
Abychom mohli stanovit nejhodnotnější populace z hlediska parametrů genetické diverzity, seřadili jsme všechny populace podle hodnot jejich TA, P, AP, HE, HO, FST, FIS a podle počtu unikátních alel. Nejvyšší hodnoty parametrů genetické diverzity dostaly nejnižší (nejlepší) hodnoty pořadí (až na FIS, kde byla za nejlepší považována nejnižší hodnota). Populace s celkovým nejnižším průměrným pořadím byly považovány za ty, které jsou nejhodnotnější z hlediska zachování celkové genetické diverzity v rámci každého regionu (neel & ellstranD 2003).
Kombinace dat o genetické diverzitě a populační dynamice
Poté, co jsme nalezli průkaznou korelaci mezi genetickou diverzitou (vyjádřenou jako oče kávaná heterozygozita) a produkcí semen na kvetoucí lodyhu (v letech 2003–2006), použili jsme tento vztah k tomu, abychom odhadli produkci semen při různých hodnotách genetické diverzity.
Očekávaná heterozygozita je heterozygozita, kterou bychom očekávali za podmínek, kdy dochází v populaci k náhodnému křížení při daných alelických frekvencích podle Hardy-Weinbergovy rovnováhy. Dává nám informaci o genetické diverzitě populace a kombinuje informaci o počtu alel a jejich frekvenci v populaci. V jednom lokusu se dvěmi alelami může nabývat hodnot od 0 do 0,5 a v populaci, která obsahuje mnoho alel o malých

63
Příroda, Praha, 31: 53–83, 2011
frekvencích se očekávaná heterozygozita může blížit jedné (frankhaM et al. 2002). Očekávaná heterozygozita je standardně používaná míra pro genetickou diverzitu po-pu lace (např. haMrick & GoDt 1996, frankhaM et al. 2002) a odráží dlouhodobé změny v genetické struktuře populace. Ze vztahu mezi heterozygozitou a produkcí semen jsme počítali produkci semen na kvetoucí lodyhu pro hodnoty očekávané heterozygozity mezi 0 a 0,3, což je rozpětí, kterého obvykle dosahují dlouhověké vytrvalé rostliny se smíšeným reprodukčním systémem (haMrick & GoDt 1989, haMrick & GoDt 1996).
Abychom mohli předpovědět následky změn produkce semen, které nastaly v důsledku změn v genetické diverzitě, na populační dynamiku D. austriacum, použili jsme nově získané hodnoty produkce semen v populačních přechodových maticích, které byly vytvořeny v části této práce o populační dynamice. Upravili jsme prvky matic, které se týkají produkce malých a velkých dospělých rostlin s použitím hodnot pro vypočítanou produkci semen za jednotlivých hladin očekávané heterozygozity. Počítali jsme změnu produkce semen s měnící se očekávanou heterozygozitou v každé populaci a v každém roce. Pro očekávanou heterozygozitu HE = 0 (tj. situace, kdy je v každém lokusu pouze jediná zbývající alela), byla vypočítaná produkce semen pro roky 2003 a 2004 záporná. Záporné hodnoty jsme nahradili 0, abychom zachovali biologický význam. Ze získaných matic jsme pak spočítali populační růstové rychlosti s použitím stochastických maticových modelů (viz kapitola Populační dynamika). Abychom se dozvěděli o vlivu změn očekávané heterozygozity v každé populaci, kombinovali jsme při výpočtech populační růstové rychlosti a elasticit matice ze třech přechodových intervalů (2003–2006). Abychom se dozvěděli o vlivu změn v Českém a ve Slovenském krasu, kombinovali jsme matice ze třech přechodových intervalů a ze třech populací v rámci každého krasového regionu. Pro hodnoty populační růstové rychlosti jsme také spočítali 95 % intervaly spolehlivosti, které nám určily průkazné rozdíly mezi nimi (viz kapitola Populační dynamika).
Dále jsme také počítali pravděpodobnosti vymření modelových populací v Českém a Slovenském krasu za různých hodnot očekávané heterozygozity (viz výše). Modelové populace byly použity stejné jako v části Populační dynamika o velikosti 10 000, 5 000 a 1 000 jedinců rozložených ve fázích životního cyklu podle stabilního rozložení životních fází. Počítali jsme pravděpodobnost vymření populací do 20, 50 a 100 let. Pravděpodobnosti byly založeny na 500 opakováních.
Všechny analýzy byly provedeny v programu Matlab verze 7.3.0.267 (The MathWorks, Inc., Natick, Massachusetts, USA).
výsledky
Populační dynamika
Rostliny ze Slovenského krasu mají v průměru o 9,9 % vyšší pravděpodobnost přežívání, o 39,4 % vyšší kvetení a 2,4× vyšší produkci semen než rostliny z Českého krasu. Nenašli jsme průkazné rozdíly v pravděpodobnostech růstu rostlin a retrogrese. Nalezli jsme ale mnoho rozdílů mezi jednotlivými roky a populacemi (obr. 2). Podrobné výsledky statistických analýz jsou k dispozici v práci Dostálek & MünzberGová (2009).
Mortalita rostlin klesala s jejich velikostí, přičemž semenáčky měly větší mortalitu než malé dospělé rostliny 4,8× v Českém a 13,0× ve Slovenském krasu a ve srovnání s velkými dospělými rostlinami to bylo 15,3× více v Českém a 52,6× více ve Slovenském krasu (příloha A). Velké dospělé rostliny také produkovaly více semen než malé dospělé rostliny v průměru 5,5× v Českém a 3,2× více ve Slovenském krasu. Semena klíčila s pravděpodobností 0 až 0,04 % (příloha A).

64
Dostálek & Münzbergová: Populační dynamika a genetická diverzita Dracocephalum austriacum
Jednotlivé přechodové matice měly hodnoty populační růstové rychlosti od 0,960 do 1,050. Všechny populační růstové rychlosti nebyly průkazně odlišné od 1 (indikující stabilní populace) s výjimkou přechodu v letech 2005–2006, kdy se populace C4 průkazně zmenšovala a v roce 2003–2004, kdy se populace S3 průkazně zvětšovala. Podobné výsledky jsme dostali také po zkombinování dat z let 2003–2006 pro každou populaci (obr. 3). V tomto případě byly populační růstové rychlosti od 0,986 do 1,039. Populace S1 a S3 ve Slovenském krasu se průkazně zvětšovaly a populace C4 v Českém krasu se průkazně zmenšovala. Ostatní populace byly stabilní (populační růstová rychlost se průkazně nelišila od 1). Výsledky byly velice podobné, i když jsme zkombinovali všechny matice v rámci každého regionu. Populační růstová rychlost byla průkazně větší ve Slovenském krasu ve všech letech (obr. 3).
Ve všech populacích byly největší hodnoty elasticity pro prvky matice, které vyjadřují přežívání ve stejné fázi životního cyklu. Celkový součet prvků v matici, které vyjadřují přežívání, byl od 0,78 do 0,88. Jednotlivé nejdůležitější prvky v matici byly přežívání ve fázi semen v semenné bance pro populace C2, C4 a S3 (elasticita 0,39, 0,53 a 0,31). V populaci S2 to bylo přežívání ve fázi malých dospělých rostlin (elasticita 0,36) a v populacích C1 a S1 to bylo přežívání ve fázi velkých dospělých rostlin (elasticita 0,36 a 0,45). Když byly zkombinovány všechny matice v jednom regionu, tak byly nejdůležitější přechody v obou regionech rovněž přežívání semen v semenné bance a ve fázi malých a velkých dospělých rostlin.
Pravděpodobnost vymření všech studovaných populací byla velice nízká i v horizontu 100 let. Nejvyšší pravděpodobnost vymření do 100 let byla 23 % a vykazovala ji populace C4. Velká populace C1 a všechny slovenské populace měly dokonce 0 % pravděpodobnost vymření do 100 let. Obě modelové populace s počáteční velikostí 10 000 jedinců a populace s počáteční velikostí 5 000 jedinců ve Slovenském krasu měly také téměř 0 % pravděpodobnost vymření. Modelová populace s 5 000 jedinci z Českého krasu měla pravděpodobnost vymření do 100 let 12 %. Modelové populace s 1 000 jedinců měly mnohem větší pravděpodobnosti vymření – 31 a 26 % do 20 let, 61 a 40 % do 50 let a 78 a 43 % do 100 let (Český a Slovenský kras), viz tab. 2.
Vykopávání velkých rostlin zahrádkáři významně zvýšilo pravděpodobnosti vymření populací. Vykopávání menšího počtu (jedna až tři) velkých dospělých rostlin by však neměl být vážný problém pro velké populace jako je C2 nebo pro populace ve Slovenském krasu. Vykopání deseti jedinců každý rok však vede k vysokým pravděpodobnostem vymření v delším horizontu i ve velkých vitálních populacích (až na největší populaci C2). Když porovnáme populace z Českého a Slovenského krasu, tak pravděpodobnosti vymření jsou vyšší u českých populací. Slovenské populace jsou odolnější k menšímu počtu vykopávaných jedinců. V malých populacích s přibližně deseti rostlinami však vykopávání velkých dospělých rostlin vede k vymření všech populací do 20 let (tab. 2).
tab. 2. Pravděpodobnosti vymření (v % z 500 simulací) studovaných populací v Českém krasu (CK), na Moravě a ve Slovenském (SK) krasu a pravděpodobnosti vymření modelových populací z Českého a Slovenského krasu s počáteční velikostí 10 000, 5 000 a 1 000 jedinců. Populační vektor je počet semen, semenáčků, malých a velkých dospělých rostlin, který byl použit jako počáteční velikost populace.tab. 2. Extinction probabilities (in % of 500 simulations) of studied populations in the Bohemian Karst (CK), Moravian and Slovak (SK) Karst and extinction probabilities of model populations from the Bohemian and Slovak Karst with 10 000, 5 000, and 1 000 individuals as the initial population size. The population vector is a number of seeds, seedlings, and small and large adult plants used as an initial population vector.
65
Příroda, Praha, 31: 53–83, 2011
Pra
vděp
odob
nost
vym
ření
[%]
do 2
0 le
t
do 5
0 le
t
do 1
00 le
t
P
očet
vyk
opan
ých
velk
ých
rost
lin z
a ro
k0
13
510
0
13
510
0
13
510
Pop
ulac
eP
opul
ační
vek
tor
C1
2142
9-27
-103
-81
00
00
00
00,
86
170,
611
7697
99
C2
5799
5-13
2-27
6-18
10
00
00
00
00
00
00
04.
8
C3
664-
0-2-
538
8291
9594
7199
100
100
100
8710
010
010
010
0
C4
6562
-23-
116-
690
00
00,
20,
60,
89
2438
2357
9910
010
0
C5
828-
3-1-
826
6685
9090
5897
100
100
100
7910
010
010
010
0
C6
5152
-23-
45-1
10
0,2
23,
87,
60,
414
6278
858,
874
100
100
100
C7
4416
-5-4
0-8
00,
44.
48,
811
231
7688
9116
8810
010
010
0
C8
5060
0-12
3-25
7-16
80
00
00
00
00
00
00
1,4
21
M1
736-
0-4-
439
7591
9093
7198
100
100
100
8610
010
010
010
0
S1
1565
7-26
-31-
560
00
00
00
0,2
2,6
210
03,
834
98
S2
4613
2-32
-93-
350
00
00,
20
00,
40,
41,
80
06,
228
66
S3
2805
1-53
-63-
410
00
00
0
00
00,
4
00,
20,
45
36
Mod
elov
á po
pula
ceC
K –
10
000
9908
-10-
42-4
00
00
0,4
0,6
01,
215
3553
124
9510
010
0
SK
– 1
0 00
099
08-1
0-42
-40
00
00,
81,
40
03,
613
330
024
7199
CK
– 5
000
4954
-5-2
1-20
0,2
06,
610
141,
828
7889
8912
8010
010
010
0
SK
– 5
000
4954
-5-2
1-20
00,
45,
87,
614
05,
845
7183
018
8910
010
0
CK
– 1
000
991-
1-4-
431
7489
9293
6199
100
100
100
7810
010
010
010
0
SK
– 1
000
991-
1-4-
426
6584
8683
4090
100
100
100
4394
100
100
100
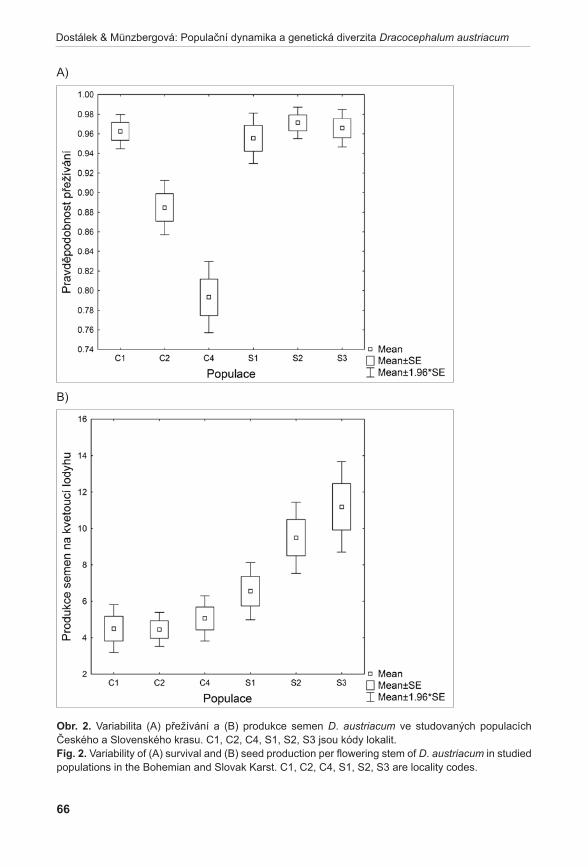
66
Dostálek & Münzbergová: Populační dynamika a genetická diverzita Dracocephalum austriacum
A)
B)
obr. 2. Variabilita (A) přežívání a (B) produkce semen D. austriacum ve studovaných populacích Českého a Slovenského krasu. C1, C2, C4, S1, S2, S3 jsou kódy lokalit.Fig. 2. Variability of (A) survival and (B) seed production per flowering stem of D. austriacum in studied populations in the Bohemian and Slovak Karst. C1, C2, C4, S1, S2, S3 are locality codes.

67
Příroda, Praha, 31: 53–83, 2011
Genetická diverzita
Celkem šest z deseti lokusů hodnocených o enzymatických systémů bylo polymorfních (variabilních) a deset hodnocených lokusů obsahovalo celkem 24 alel. Na každý lokus tak připadlo v průměru 2,40 (SO = 2,12) alel a 3,33 (SO = 2,13) alel na polymorfní pokus. Očekávaná heterozygozita (HE) pro celý datový soubor (druh) činila 0,15 (SO = 0,19) a pozorovaná heterozygozita (HO) byla 0,08 (SO = 0,11).
Podíl polymorfních lokusů ve 12 sledovaných populacích činil od 0 (C5) do 0,5 (C4 a C7). Počet alel na populaci byl 10 (C5) až 19 (C8). Počet alel na polymorfní lokus pro jednotlivé populace byl od 2,2 (C4) do 3,5 (M1). HE v rámci populace dosahovala od 0,000 (C5) do 0,189 (S1). HO v rámci populace dosahovala od 0 (C5) do 0,141 (S3), viz tab. 1.
Jedna unikátní alela byla nalezena v populaci S2 a jedna v populaci S3. Tři unikátní alely byly nalezeny pro Český kras a tři pro Slovenský kras (tab. 1). Koeficient inbreedingu (FIS) sahal od 0,079 (M1) do 0,638 (C3). Ve všech populacích byly hodnoty pozorované heterozygozity menší než hodnoty očekávané heterozygozity (tab. 1).
Nejnižší průměrné pořadí bylo zaznamenáno u moravské populace (M1) a u všech slovenských populací (S1, S2, S3). V Českém krasu se jako nejhodnotnější populace z hlediska genetické diverzity jevily populace C2, C3 a C8 (tab. 1).
Analýza molekulární variance (AMOVA) ukázala, že největší podíl genetické variability je v rámci populací (80,14 %), zatímco mezi regiony je to pouze 10,45 %. Variabilita mezi populacemi v rámci regionů byla 9,42 %. Hodnota FST = 0,177 (variabilita mezi populacemi mezi regiony) ukazuje na nízký genový tok mezi populacemi. Hodnoty FST byly 0,143 pro Český kras a 0,088 pro Slovenský kras, což znamená, že populace v Českém krasu jsou geneticky více odděleny.
Hodnoty FST pro všechny páry populací sahaly od -0,022 do 0,637 (54 ze 66 párů bylo průkazně geneticky odlišných). Genetické vzdálenosti mezi populacemi byly průkazně korelovány s geografickými vzdálenostmi pro všechny populace dohromady (r = 0,44, P = 0,002), avšak samostatně pro populace z Českého a Slovenského krasu tato korelace průkazná nebyla (P = 0,58 pro Český kras; P = 0,66 pro Slovenský kras).
Mezi jednotlivými mírami genetické diverzity byly následující průkazné korelace: pozitivní mezi HE a HO (r = 0,83, P = 0,003), TA a P (r = 0,66, P = 0,036), TA a AP (r = 0,73, P = 0,024) a negativní mezi FIS a HO (r = -0,70, P = 0,025).
Nalezli jsme pozitivní průkazné korelace mezi produkcí semen na kvetoucí lodyhu a mírami genetické diverzity – očekávanou heterozygozitou (r = 0,70, P = 0,036, obr. 4) a pozorovanou heterozygozitou (r = 0,80, P = 0,009). Produkce semen na kvetoucí rostlinu byla pozitivně průkazně korelována s pozorovanou heterozygozitou (r = 0,78, P = 0,012) a negativně korelována s koeficientem inbreedingu (r = -0,71, P = 0,033). Průkazná korelace byla také mezi celkovým počtem alel a produkcí semen na kvetoucí rostlinu (r = 0,75, P = 0,034). Podíl vyvinutých semen byl průkazně korelován s celkovým počtem alel (r = 0,75, P = 0,020).
Nenalezli jsme žádnou průkaznou korelaci mezi mírou genetické diverzity a velikostí populace, počtem kvetoucích rostlin nebo podílu kvetoucích rostlin v populaci, až na průkaznou pozitivní korelaci mezi počtem polymorfních lokusů a velikostí populace (r = 0,78, P = 0,008).
Dále jsme našli průkaznou pozitivní korelaci mezi velikostí populace a podílem vyvinutých semen (r = 0,67, P = 0,049) a produkcí semen na kvetoucí rostlinu (r = 0,73, P = 0,026), ale ne mezi velikostí populace a produkcí semen na kvetoucí lodyhu (P = 0,368).
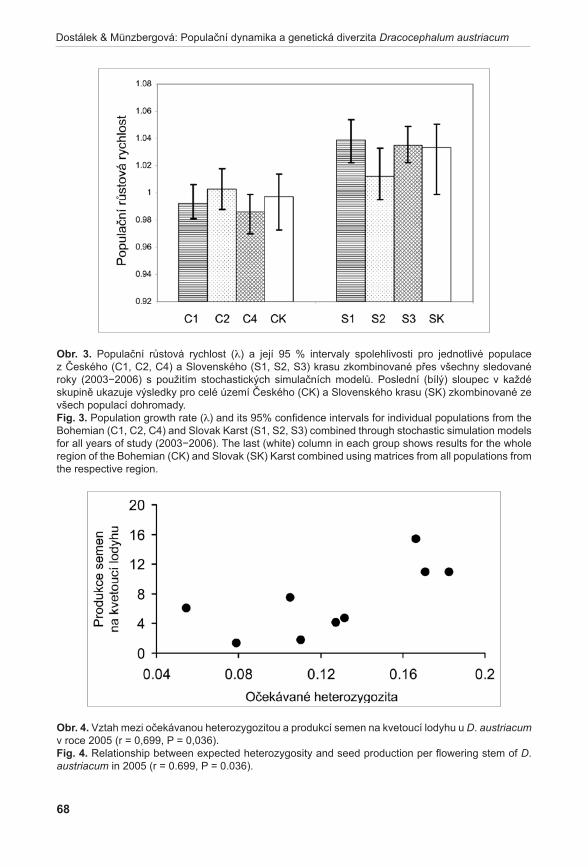
68
Dostálek & Münzbergová: Populační dynamika a genetická diverzita Dracocephalum austriacum
obr. 3. Populační růstová rychlost (l) a její 95 % intervaly spolehlivosti pro jednotlivé populace z Českého (C1, C2, C4) a Slovenského (S1, S2, S3) krasu zkombinované přes všechny sledované roky (2003−2006) s použitím stochastických simulačních modelů. Poslední (bílý) sloupec v každé skupině ukazuje výsledky pro celé území Českého (CK) a Slovenského krasu (SK) zkombinované ze všech populací dohromady.Fig. 3. Population growth rate (l) and its 95% confidence intervals for individual populations from the Bohemian (C1, C2, C4) and Slovak Karst (S1, S2, S3) combined through stochastic simulation models for all years of study (2003−2006). The last (white) column in each group shows results for the whole region of the Bohemian (CK) and Slovak (SK) Karst combined using matrices from all populations from the respective region.
obr. 4. Vztah mezi očekávanou heterozygozitou a produkcí semen na kvetoucí lodyhu u D. austriacum v roce 2005 (r = 0,699, P = 0,036).Fig. 4. Relationship between expected heterozygosity and seed production per flowering stem of D. austriacum in 2005 (r = 0.699, P = 0.036).
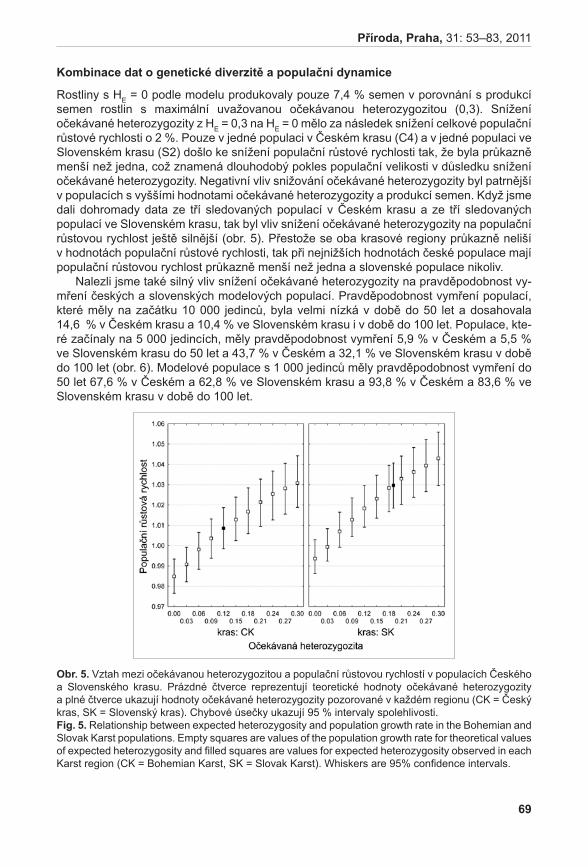
69
Příroda, Praha, 31: 53–83, 2011
Kombinace dat o genetické diverzitě a populační dynamice
Rostliny s HE = 0 podle modelu produkovaly pouze 7,4 % semen v porovnání s produkcí semen rostlin s maximální uvažovanou očekávanou heterozygozitou (0,3). Snížení očekávané heterozygozity z HE = 0,3 na HE = 0 mělo za následek snížení celkové populační růstové rychlosti o 2 %. Pouze v jedné populaci v Českém krasu (C4) a v jedné populaci ve Slovenském krasu (S2) došlo ke snížení populační růstové rychlosti tak, že byla průkazně menší než jedna, což znamená dlouhodobý pokles populační velikosti v důsledku snížení očekávané heterozygozity. Negativní vliv snižování očekávané heterozygozity byl patrnější v populacích s vyššími hodnotami očekávané heterozygozity a produkcí semen. Když jsme dali dohromady data ze tří sledovaných populací v Českém krasu a ze tří sledovaných populací ve Slovenském krasu, tak byl vliv snížení očekávané heterozygozity na populační růstovou rychlost ještě silnější (obr. 5). Přestože se oba krasové regiony průkazně neliší v hodnotách populační růstové rychlosti, tak při nejnižších hodnotách české populace mají populační růstovou rychlost průkazně menší než jedna a slovenské populace nikoliv.
Nalezli jsme také silný vliv snížení očekávané heterozygozity na pravděpodobnost vy-mření českých a slovenských modelových populací. Pravděpodobnost vymření populací, kte ré měly na začátku 10 000 jedinců, byla velmi nízká v době do 50 let a dosahovala 14,6 % v Českém krasu a 10,4 % ve Slovenském krasu i v době do 100 let. Populace, kte-ré začínaly na 5 000 jedincích, měly pravděpodobnost vymření 5,9 % v Českém a 5,5 % ve Slovenském krasu do 50 let a 43,7 % v Českém a 32,1 % ve Slovenském krasu v době do 100 let (obr. 6). Modelové populace s 1 000 jedinců měly pravděpodobnost vymření do 50 let 67,6 % v Českém a 62,8 % ve Slovenském krasu a 93,8 % v Českém a 83,6 % ve Slo ven ském krasu v době do 100 let.
obr. 5. Vztah mezi očekávanou heterozygozitou a populační růstovou rychlostí v populacích Českého a Slovenského krasu. Prázdné čtverce reprezentují teoretické hodnoty očekávané heterozygozity a plné čtverce ukazují hodnoty očekávané heterozygozity pozorované v každém regionu (CK = Český kras, SK = Slovenský kras). Chybové úsečky ukazují 95 % intervaly spolehlivosti.Fig. 5. Relationship between expected heterozygosity and population growth rate in the Bohemian and Slovak Karst populations. Empty squares are values of the population growth rate for theoretical values of expected heterozygosity and filled squares are values for expected heterozygosity observed in each Karst region (CK = Bohemian Karst, SK = Slovak Karst). Whiskers are 95% confidence intervals.
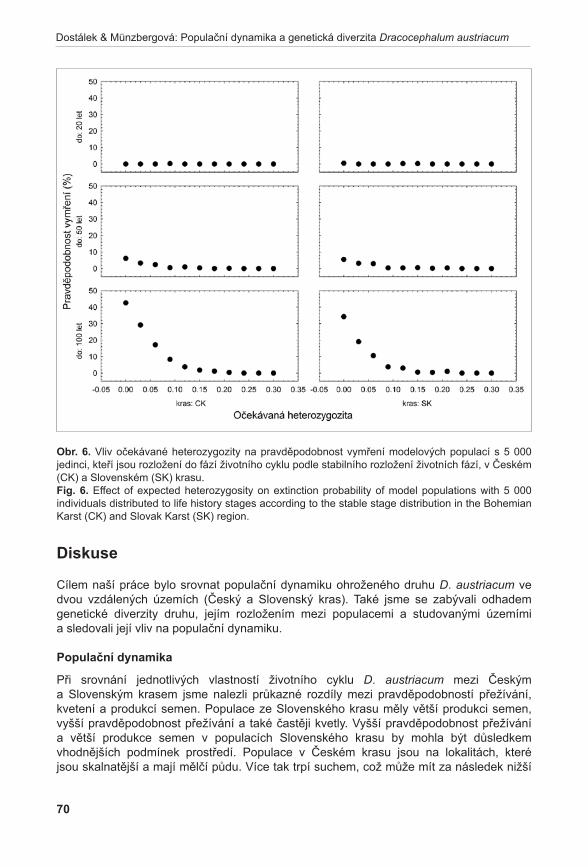
70
Dostálek & Münzbergová: Populační dynamika a genetická diverzita Dracocephalum austriacum
obr. 6. Vliv očekávané heterozygozity na pravděpodobnost vymření modelových populací s 5 000 jedinci, kteří jsou rozložení do fází životního cyklu podle stabilního rozložení životních fází, v Českém (CK) a Slovenském (SK) krasu.Fig. 6. Effect of expected heterozygosity on extinction probability of model populations with 5 000 individuals distributed to life history stages according to the stable stage distribution in the Bohemian Karst (CK) and Slovak Karst (SK) region.
diskuse
Cílem naší práce bylo srovnat populační dynamiku ohroženého druhu D. austriacum ve dvou vzdálených územích (Český a Slovenský kras). Také jsme se zabývali odhadem genetické diverzity druhu, jejím rozložením mezi populacemi a studovanými územími a sledovali její vliv na populační dynamiku.
Populační dynamika
Při srovnání jednotlivých vlastností životního cyklu D. austriacum mezi Českým a Slovenským krasem jsme nalezli průkazné rozdíly mezi pravděpodobností přežívání, kvetení a produkcí semen. Populace ze Slovenského krasu měly větší produkci semen, vyšší pravděpodobnost přežívání a také častěji kvetly. Vyšší pravděpodobnost přežívání a větší produkce semen v populacích Slovenského krasu by mohla být důsledkem vhodnějších podmínek prostředí. Populace v Českém krasu jsou na lokalitách, které jsou skalnatější a mají mělčí půdu. Více tak trpí suchem, což může mít za následek nižší

71
Příroda, Praha, 31: 53–83, 2011
pravděpodobnosti přežívání rostlin v Českém krasu a jejich nižší produkci semen. willeMs & ellers (1996) nalezli rozdíly v populačních charakteristikách vytrvalého druhu Orchis simia v jeho severozápadní a jihovýchodní části areálu. V jihovýchodní části areálu nalezli vyšší hodnoty fitness populací způsobené lepšími klimatickými podmínkami a příznivější strukturou vegetace. Navíc García et al. (2000) ukázali rozdíly v produkci semen v rámci evropské části areálu Juniperus communis. Domnívají se, že by to mohlo být způsobeno zhoršenými klimatickými podmínkami nebo sníženou genetickou diverzitou některých po-pu lací.
Rozdíly mezi jednotlivými přechody v životním cyklu měly také vliv na celkovou populační dynamiku. Populační růstová rychlost spočítaná z dat z let 2003–2006 byla průkazně vyšší v populacích Slovenského krasu. Vyšší produkce semen v populacích Slovenského krasu, které jsou více v centru areálu rozšíření, je pravděpodobně jedním z hlavních důvodů rozdílů v populační dynamice nalezených mezi jednotlivými regiony. Průkazné rozdíly v populační dynamice mezi centrem a okrajem ukázali ve své práci také nantel & GaGnon (1999). Navrhli také, že za tyto rozdíly je zodpovědná variabilita v podmínkách prostředí. V jiné naší studii (Dostálek & MünzberGová 2009) se však nepotvrdily vlivy podmínek prostředí nebo srážek či teplot na populační dynamiku. To však mohlo být způsobeno relativně krátkou dobou sledování nebo malým počtem sledovaných populací.
Populace D. austriacum také velmi často zarůstají křovinami a nálety stromů, což ovlivňuje jednotlivé přechody v životním cyklu. Nicméně vliv tohoto zarůstání zatím nebyl studován, třebaže vyřezávání křovin a náletových stromů je jedním z převažujících typů managementu v obou územích. Další studie by se proto měly určitě zabývat právě vlivem zarůstání lokalit.
Největší vliv na změnu populační růstové rychlosti měly změny v přežívání semen v semenné bance a změny v přežívání ve fázi malých a velkých dospělých rostlin. Podle klasifikace silvertown et al. (1993) je D. austriacum podle hodnot elasticity řazen mezi iteroparní byliny otevřených stanovišť, což odpovídá námi pozorované skutečnosti. Malé rozdíly v hodnotách elasticity mezi regiony jsou pak ve shodě s očekáváním, že přechody mezi fázemi životního cyklu, které mají největší vliv na populační růstovou rychlost, jsou značně konzervativní (např. ehrlén et al. 2005).
Semena D. austriacum mají tvrdý obal, a tak předpokládáme, že jejich přežívání v semenné bance je vysoké. Dlouho vytrvávající semenná banka může hrát velkou roli zejména u malých populací ohrožených snižující se genetickou diverzitou. Význam semenné banky pro znovuobnovení rostlinných populací ukazují ve své studii např. aleXanDer & schraG (2003). Výsledky některých dalších prací však ukazují (např. thoMPson et al. 1997, baskin & baskin 1998, hanDlová & MünzberGová 2006), že semenná banka obvykle nestačí ke znovuobnovení populace, která dříve na lokalitě už vymřela. Přežívání semen v semenné bance bylo v této práci odhadováno nepřímo z množství živých semen přítomných v půdě na lokalitách v Českém krasu. Přímý odhad by vyžadoval pokus, ve kterém bychom sledovali přežívání semen zakopaných v půdě na lokalitách. Tento přístup už byl opakovaně úspěšně použit pro odhad přežívání v semenné bance (např. warDle 2003, MünzberGová 2004, 2005, treMlová & MünzberGová 2007). Zakopávací pokus však nebylo možno v případě D. austriacum provést z důvodu malého množství semen přítomného v populacích. Tento pokus také obvykle trvá dva až tři roky, ale naše nepřímá data ukazují, že semena mohou v půdě přežívat i více než 12 let, a tento krátkodobý pokus by tak v případě D. austriacum nebyl moc užitečný. Když jsme však zkoušeli počítat populační růstové rychlosti i se sníženou pravděpodobností přežívání semen v semenné bance, zjistili jsme relativně malý vliv na snížení populační růstové rychlosti.

72
Dostálek & Münzbergová: Populační dynamika a genetická diverzita Dracocephalum austriacum
Genetická diverzita
Naše studie ukazuje na velké množství genetické diverzity, která je soustředěna v populacích D. austriacum, což ukazuje na to, že populace D. austriacum mají v rámci obou regionů poměrně vysokou genetickou diverzitu. Navzdory našemu očekávání však byla genetická odlišnost obou regionů relativně nízká. Podobně malé rozdíly byly pozorovány u populací druhů s dobře šiřitelným pylem a semeny (např. brzosko et al. 2002). To však není případ D. austriacum (viz níže).
I přes nízkou odlišnost studovaných regionů jsme nalezli poměrně velké rozdíly v míře genetické diferenciace mezi studovanými populacemi (FST = 0,177). Tyto rozdíly odpovídají hodnotám udávaným pro druhy opylované zvířaty (hmyzem), které uvádějí haMrick & GoDt (1989) ve své studii, která se zabývala přehledem studií genetické diverzity pomocí analýzy izoenzymů. Podobné výsledky ukazují i další studie (např. ayres & ryan 1999, Martínez-Palacios et al. 1999).
Vysoká genetická izolovanost studovaných populací je také v souladu se skutečností, že D. austriacum je opylován většinou čmeláky a včelami (S. Castro, nepublikováno) a je tak dost nepravděpodobné, že by byli tito opylovači schopni přenést pyl z jedné lokality na jinou (Darvill et al. 2004). Šíření semen mezi jednotlivými populacemi je také dost nepravděpodobné (wiDén & wiDén 1990), protože semena D. austriacum jsou velká a těžká bez jakýchkoliv adaptací k šíření. Podobně vysoká míra genetické izolovanosti byla nalezena i pro další hmyzem opylované druhy (např. colas et al. 1997, Martínez-Palacios et al. 1999). Naproti tomu ben faDhel & boussaïD (2004) nalezli v populacích Mentha pulegium, která je ze stejné čeledi jako D. austriacum, dost vysokou vnitropopulační genetickou diverzitu, ale nižší mezipopulační variabilitu. Předpokládali, že by to mohlo být způsobeno relativně nedávným původem sledovaných populací.
Hodnoty očekávané heterozygozity pro jednotlivé populace D. austriacum jsou poměrně vysoké ve srovnání s hodnotami, které udávají haMrick & GoDt (1989) pro hmyzem opylované druhy se smíšeným opylovacím systémem (mixed mating). Nalezli jsme také nižší genetickou diverzitu v populacích Českého krasu než v populacích Slovenského krasu. Jedním z vysvětlení může být pozice Českého krasu na severním okraji areálu druhu. Nižší genetickou diverzitu na okraji areálu uvádějí také další studie (laMMi et al. 1999, fauGeron et al. 2004).
Produkce semen byla průkazně vyšší v geneticky variabilnějších a méně inbredních populacích. Podobné výsledky byly zjištěny i v dalších studiích (např. husbanD & scheMske 1996, fischer & Matthies 1998, schMiDt & jensen 2000, hensen & oberPrieler 2005). Ve shodě s těmito studiemi jsme také nalezli průkaznou korelaci mezi produkcí semen na kvetoucí rostlinu a velikostí populace. Tato korelace je často udávána pro druhy, jejichž populace se zmenšily vlivem fragmentace nebo úplným zničením jejich stanovišť (např. luijten et al. 2000). Toto je pravděpodobně také případ D. austriacum. Některé populace se za posledních 50 let zmenšily (E. Karasová, osobní sdělení) a jiné byly zničeny těžbou (Moucha 1990). Nižší produkce semen v malých a geneticky méně variabilních populacích D. austriacum může být způsobena inbreedingem, který může hrát důležitou roli v populacích těchto druhů se smíšeným rozmnožovacím systémem (kePhart et al. 1999). Inbreeding může být důsledkem zvýšené míry křížení blízce příbuzných rostlin v malých populacích a/nebo křížením s geneticky blízce příbuznými sousedícími rostlinami (např. prostorově oddělené skupiny v rámci velké populace). Dalším důvodem pro vztah mezi velikostí populace a produkcí semen by mohla být zvýšená frekvence geitonogamie v malých populacích (lloyD & schoen 1992). Toto vysvětlení je velice pravděpodobné, protože rostliny D. austriacum tvoří velké trsy s mnoha květy a je mnohem pravděpodobnější,

73
Příroda, Praha, 31: 53–83, 2011
že opylovači létají mezi květy téže rostliny. Dalším vysvětlením, které je pravděpodobné na základě prozatímních opylovacích pokusů (S. Castro, nepublikováno), pak může být jednoduše nedostatek opylovačů, který vede ke spontánnímu samoopylování.
vztah mezi genetickou diverzitou a populační dynamikou
V této části jsme se snažili porozumět vlivu změn genetické diverzity na dlouhodobou populační růstovou rychlost D. austriacum. Nalezli jsme pozitivní korelaci mezi očekávanou heterozygozitou a produkcí semen na kvetoucí lodyhu a s použitím tohoto vztahu jsme odhadovali vliv změn očekávané heterozygozity na dlouhodobou populační růstovou rychlost. Přestože vliv snižující se očekávané heterozygozity na snížení produkce semen byl relativně silný, pouze velký pokles očekávané heterozygozity měl za následek průkazné snížení dlouhodobé populační růstové rychlosti. Snížení očekávané heterozygozity také ovlivnilo nárůst pravděpodobnosti vymření populací D. austriacum. Všechny tyto vlivy byly podobné v obou studovaných krasových územích, ale těsnější závislosti byly v populacích Slovenského krasu.
Na rozdíl od většiny předchozích studií jsme se nesoustředili na vztah mezi genetickou diverzitou a fitness pouze jedné části životního cyklu, ale zajímaly nás také důsledky tohoto vztahu na celkovou populační růstovou rychlost (fischer & Matthies 1998, luijten et al. 2002, Picó et al. 2004). Jediné studie, které se snažily přímo propojit genetickou diverzitu s populační růstovou rychlostí byly Picó et al. (2004), kirchner et al. (2006) a oosterMeijer (2000). Zjistily silný negativní vliv inbreedingu (Picó et al. 2004) a alogamických (self-incom-patible) alel (kirchner et al. 2006) na populační růstovou rychlost. oosterMeijer (2000) ve své práci také nalezl silný vliv inbrední deprese na velikost populace, pravděpodobnost vymření populace a populační růstovou rychlost. Podobně jako předcházející studie jsme i my nalezli silný negativní vliv klesající očekávané heterozygozity na populační růstovou rychlost. Na rozdíl od předcházejících studií jsme však tyto vlivy sledovali v šesti populacích ve dvou vzdálených územích a zahrnuli jsme celé spektrum hodnot očekávané heterozygozity. Mohli jsme tak porovnat změny ve tvaru závislosti mezi očekávanou heterozygozitou a populační růstovou rychlostí ve dvou odlišných vzdálených územích. Také jsme získali informace o vlivech změn očekávané heterozygozity na pravděpodobnosti vymření sledovaných populací.
Ve Slovenském krasu i při velkém snížení očekávané heterozygozity populace početně průkazně neklesají. Naproti tomu v populacích Českého krasu vede snížení očekávané heterozygozity k průkaznému snižování velikosti populací a jejich postupnému vymírání.
Při analýze jednotlivých populací jsme zaznamenali u všech populací (až na jednu) průkazný vliv na snížení populační růstové rychlosti vlivem snížené očekávané heterozygozity. Jedinou výjimkou byla populace C1 v Českém krasu, kde ani snížení očekávané heterozygozity na HE = 0 nemělo za následek průkazné snížení populační růstové rychlosti. V této populaci jsme zjistili velmi nízkou klíčivost semen ze semenné banky a relativně nízkou produkci semen v porovnání s ostatními populacemi. Pouze ve dvou populacích (C4 a S2) způsobila snížená očekávaná heterozygozita průkazný dlouhodobý pokles populací, což ukazuje na to, že snížení produkce semen nemusí vést k přímému ohrožení populací. Je to také pravděpodobně způsobeno tím, že D. austriacum je dlouhověká vytrvalá rostlina, pro kterou je přežívání mnohem důležitější než produkce semen (silvertown et al. 1993). Nicméně jakýkoliv pokles populační růstové rychlosti, i když populace ještě průkazně neklesá, by neměl zůstat bez povšimnutí a měli bychom podniknout opatření, která by vedla k nápravě situace předtím, než bude příliš pozdě (např. inGvarsson 2001).

74
Dostálek & Münzbergová: Populační dynamika a genetická diverzita Dracocephalum austriacum
Nalezli jsme také silný vliv snížení očekávané heterozygozity na pravděpodobnost vymření modelových populací z Českého a Slovenského krasu. Velikosti modelových populací (82, 41 a 8 dospělých rostlin) jsou ve skutečnosti opravdové velikosti většiny populací D. austriacum ve sledovaných územích. Hlavně v malých populacích může mít snížení očekávané heterozygozity a následné snížení produkce semen fatální důsledky pro jejich další existenci. Vliv snížení očekávané heterozygozity je více patrný až v delším časovém horizontu, což je způsobeno zejména dlouhým životním cyklem druhu (až 40 let, nepublikovaný odhad založený na demografických datech). Zvýšenou pravděpodobnost vymření populací následkem sníženého fitness udávají i další studie (newMan & Pilson 1997, brook et al. 2002, frankhaM 2003, Picó et al. 2004).
Na závěr je důležité zmínit, že výsledky modelů populační dynamiky je potřeba interpretovat s opatrností, protože to jsou pouze modely (beissinGer & westPhal 1998, Marrero-GóMez et al. 2007). Je proto potřeba si být vědom jejich omezení, zvláště když jsou jejich výsledky používány k rozhodnutím týkajících se ochrany druhů a jejich managementu. V případě našeho modelu je důležité zdůraznit, že mohou existovat také další faktory zodpovědné za variabilitu v produkci semen, které nebyly brány v úvahu. Bylo by také užitečné mít více informací o příčinách snížené heterozygozity v populacích. Proto by se těmito tématy měly zabývat další studie tohoto druhu.
Závěry pro ochranu druhu
Populace D. austriacum jsou dnes ohroženy zejména snižováním plochy jejich původních stanovišť, která zarůstají křovinami a stromy (Machová & kubát 2004, E. Karasová, ústí sdělení, T. Dostálek, osobní pozorování). Ve studovaných populacích Českého a Slovenského krasu téměř nikdy neklesly stochastické populační růstové rychlosti pod jednu, což znamená, že populace v současné době početně neklesají. Výhledy pro přežívání populací D. austriacum jsou tedy dobré. Je však nutné udržet alespoň současný stav jejich stanovišť a omezit zarůstání lokalit křovinami a stromy. Kromě úbytku původních stanovišť jsou populace ohrožovány také vykopáváním rostlin skalničkáři, což představuje závažný problém zejména v malých populacích. Několik populací ve sledovaném území už z tohoto důvodu vymřelo.
Malé populace D. austriacum jsou také ohroženy demografickou a environmentální stochasticitou. I populace s 50 dospělými rostlinami však nejsou bezprostředně ohroženy vymřením, pokud se nezmění stávající podmínky na lokalitách. Ale populace s deseti a méně dospělými rostlinami mají velkou pravděpodobnost vymření už jenom kvůli náhodným procesům, které v populacích a na lokalitách probíhají.
Přežívání ve fázi semen, malých a velkých dospělých rostlin má největší teoretický vliv na populační růstovou rychlost. Tyto přechody jsou však stabilní mezi jednotlivými populacemi a roky, což ukazuje na to, že nejsou moc citlivé ke změnám na lokalitách. Přechody, jejichž variabilita nejvíce přispívá ke skutečné variabilitě populační růstové rychlosti a které by měly být cílem managementových opatření, jsou produkce semen a vzcházení semenáčků (Dostálek & MünzberGová 2009). Tyto přechody se zdají být nejvíce ovlivněny fluktuací klimatu a expanzí dřevin na lokalitách. Snížená genetická diverzita a následný inbreeding jsou také pravděpodobně částečně zodpovědné za tuto variabilitu.
Ze studia genetické diverzity druhu vyplývá, že všechny studované slovenské populace mají vysokou míru genetické diverzity a každá z populací nese unikátní genetickou informaci v rámci Slovenského krasu. Z toho důvodu by všechny tři slovenské populace měly být prioritně chráněny. V České republice jsou z hlediska genetické diverzity tři

75
Příroda, Praha, 31: 53–83, 2011
nejhodnotnější populace C2, C4 a C8. Pokud bychom však chtěli zachovat všechnu genetickou diverzitu v rámci Českého krasu, bylo by potřeba přidat ještě populaci C6, která obsahuje alely, které nejsou přítomny v žádné jiné české populaci (vyskytují se však ve slovenských populacích). Obnovení degradovaných částí lokalit by mohlo být rovněž užitečné, ale hlavní úsilí by mělo být zaměřeno na péči o současné populace, kde je soustředěno největší množství genetické diverzity. Podle naší analýzy většina malých populací nepřispívá významně ke genetické diverzitě v rámci studovaného území (s výjimkou malé izolované populace M1).
Přestože vliv snižování genetické diverzity na snížení produkce semen byl silný, pouze velké snížení genetické diverzity mělo průkazný vliv na snížení dlouhodobé populační růstové rychlosti. Snížení genetické diverzity také mělo průkazný vliv na důležitost jednotlivých přechodů mezi fázemi životního cyklu na růst populací, měřený jejich elasticitou. Nejdůležitějšími přechody však vždy zůstalo ve všech případech přežívání. Snížení genetické diverzity také zvýšilo pravděpodobnost vymření populací. Tento vliv je však pozorovatelný až při simulacích přes 100 let kvůli dlouhé délce života D. austriacum. Všechny vlivy byly srovnatelné v Českém i Slovenském krasu, ale ve Slovenském krasu byly silnější.
Ideální management druhu by tak měl spočívat v odstraňování dřevin zarůstajících lokality a posílení genového toku mezi populacemi. Posílení genového toku je však ještě potřeba ověřit s ohledem na možné negativní účinky outbrední deprese. Ochranářská opatření by také měla spočívat v aktivní ochraně druhu a vzdělávání společnosti, aby se zamezilo vykopávání rostlin zahrádkáři.
summary
Populations of D. austriacum are nowadays threatened by reduction of their original habitats and overgrowing with grasses, shrubs and trees (Machová & kubát 2004, E. Karasová, pers. comm., T. Dostálek, pers. obs.). In the studied populations in the Bohemian and Slovak Karst, the stochastic population growth rates were never significantly below one and so populations are not currently decreasing. The prospect of survival of these populations is thus good. It is, however, necessary to maintain at least the present state of the habitats and thus limit the ongoing expansion of shrubs and trees. Apart from the changing habitat conditions, alpine gardeners collecting plants may represent serious danger particularly to small populations. In fact, populations on several localities in the studied regions are nowadays extinct due to this problem.
Small populations of D. austriacum are also endangered by demographic and environmental stochasticity. But even populations of about 50 plants are not expected to become extinct in the future if current conditions on localities do not change. Populations of ten and fewer plants show high probability of becoming extinct due to demographic and environmental stochasticity and the probability will even increase if they continue losing their genetic diversity.
Survival in stages of seeds and small and large adult plants has the highest theoretical effect on the population growth rate. These transitions are, however, stable between populations and years, suggesting that they are not very sensitive to the ongoing changes. Seed and seedling production and growth from seedlings to small plants are transitions, whose variations nowadays contribute most to variation in the population growth rate and should thus be a target of management actions (Dostálek & MünzberGová 2009). These seem to be affected by climate fluctuation, especially severe droughts and by shrub expansion. Reduced genetic diversity and consequent inbreeding also seems to be an important factor causing this variation.

76
Dostálek & Münzbergová: Populační dynamika a genetická diverzita Dracocephalum austriacum
The results of the genetic part of the study imply that all the studied Slovak populations have high genetic diversity indicating that all three examined Slovak populations should be of high conservation priority. All three Slovak populations should be maintained to ensure that all alleles identified in our analyses could be preserved. Each of them has some unique genetic information within the Slovak Karst region. In the Czech Republic, three populations (C2, C4 and C8) seem to be the most valuable populations for conserving the genetic diversity within D. austriacum. However, if we wont to preserve all the existing genetic diversity within the Bohemian Karst we should also protect the C6 population. The C6 population contains alleles, which are not present in other populations in the Czech Republic (but are included in the Slovak populations). Restoration of degraded sites could be useful but should be of a lower priority than protecting existing populations where the largest amount of genetic diversity is concentrated. According to our analysis, most of the small populations do not substantially contribute to the genetic variability of the species in each of the studied regions (with the exception of the isolated small population M1).
Although the effect of the reduced heterozygosity on decrease in seed production was strong, only severe decrease in heterozygosity caused significant decrease in the long term population growth rate. Changes in heterozygosity also had a significant effect on the importance of single life history traits for population growth measured by their elasticity; proportions among survival, growth and fecundity did not change, giving the highest importance to survival in all cases. The reduction in heterozygosity increased extinction probability of the populations but this effect was visible only in a simulation based on a period of 100 years due to a long life span of the species. All these effects were consistent between two Karst regions but they were stronger in the Slovak Karst.
The optimal management action should thus include combination of removal of expanding shrubs and support of gene flow between populations. The latter step, however, has to be preceded with exploration of possible negative consequences of outbreeding depression. The management actions should also involve active protection of the species and education of the public with the aim of minimising damage caused by certain alpine gardeners.
Poděkování
Tato práce vznikla za podpory projektů GAUK 198/2005, MŠMT VaV 2B06178, GAČR 206/08/H049, MŠMT 0021620828 a AV0Z60050516. Autoři děkují E. Karasové ze Správy NP Slovenský kras a pracovníkům Správy CHKO Český kras za poskytnutí všech informací o historii a současnosti studovaných území. Dále I. Plačkové a dalším pracovníkům z isozymové laboratoře BÚ AV ČR v Průhonicích za pomoc s analýzou isozymů.

77
Příroda, Praha, 31: 53–83, 2011
literatura
aleXanDer h. M. & schraG a. M. (2003): Role of soil seed banks and newly dispersed seeds in population dynamics of the Annual Sunflower, Helianthus annuus. − J. Ecol. 91: 987−998.
alvarez-buylla e. r. & slatkin M. (1994): Finding confidence limits on population growth rates: three real examples revised. − Ecology 75: 255−260.
anDersson s. & walDMann P. (2002): Inbreeding depression in a rare plant, Scabiosa canescens (Dipsacaceae). − Hereditas 136: 207−211.
ayres D. R. & ryan F. J. (1999): Genetic diversity and structure of the narrow endemic Wyethia reticulata and its congener W. bolanderi (Asteraceae) using RAPD and allozyme techniques. − Am. J. Bot. 86: 344–353.
baskin c. c. & baskin j. M. (1998): Seeds: ecology, biogeography, and evolution of dormancy and germination. − Academic Press, San Diego, California, USA.
beissinGer s. r. & westPhal M. i. (1998): On the use of demographic models of population viability in endangered species management. − J. Wildlife Manage. 62: 821−841.
ben faDhel n. & boussaïD M. (2004): Genetic diversity in wild Tunisian populations of Mentha pulegium L. (Lamiaceae). − Genet. Resour. Crop Ev. 51: 309–321.
bonin a., nicole f., PoMPanon f., MiauD c. & taberlet P. (2007): Population adaptive index: a new method to help measure intraspecific genetic diversity and prioritize populations for conservation. − Conserv. Biol. 21: 697−708.
brook b. w., tonkyn D. w., o‘GraDy j. j. & frankhaM r. (2002): Contribution of inbreeding to extinction risk in threatened species. − Conserv. Ecol. 6: 16.
brzosko e., wróblewska a. & ratkiewicz M. (2002): Spatial genetic structure and clonal diversity of island populations of lady’s slipper (Cypripedium calceolus) from the Biebrza National Park (northeast Poland). − Mol. Ecol. 11: 2499−2509.
bucharová a., MünzberGová z. & tájek P. (2010): Population biology of two rare fern species: long life and long-lasting stability. − Am. J. Bot. 97: 1260−1271.
caswell h. (2001): Matrix population models: construction, analysis, and interpretation. − Sinauer Associates, Sunderland, Massachusetts, USA.
colas b., olivieri i. & riba M. (1997): Centaurea corymbosa, a cliff-dwelling species tottering on the brink of extinction: a demographic and genetic study. − Proc. Natl. Acad. Sci. USA 94: 3471−3476.
colas b., kirchner f., riba M., olivieri i., MiGnot a., iMbert e., beltraMe c., carbonell D. & fréville h. (2008): Restoration demography: a 10-year demographic comparison between introduced and natural populations of endemic Centaurea corymbosa (Asteraceae). − J. Appl. Ecol. 45: 1468–1476.
Čeřovský J. (1999): Dracocephalum austriacum L. − In: Čeřovský J., feráková V., holub J., MaGlocký Š. & Procházka F. [eds], Červená kniha ohrožených a vzácných druhů rostlin a živočichů ČR a SR. 5. díl. Vyšší rostliny, p. 138, Příroda, Bratislava.
Darvill b., kniGht M. & Goulson D. (2004): Use of genetic markers to quantify bumblebee foraging range and nest density. − Oikos 107: 471−478.
De kroon h., GroenenDael j. v. & ehrlen j. (2000): Elasticities: a review of methods and model limitations. − Ecology 81: 607−618.
De Mauro M. M. (1993): Relationship of breeding system to rarity in the lakeside daisy (Hymenoxys acaulis var. glabra). − Conserv. Biol. 7: 542−550.
Delouche J. C., still T. W., rasPet M. & lienharD M. (1962): The tetrazolium test for seed viability. − Miss. Agric. Exp. Sta. Tech. Bull. 51: 1−63.
Dostálek t. (2005): Identification of critical life history stages in the life cycle of endangered species, Dracocephalum austriacum L. − Ms. [Master thesis; depon. in: Department of Botany, Faculty of science, Charles University in Prague.]
Dostálek t., MünzberGová z. & PlaČková i. (2010): Genetic diversity and its effect on fitness in an endangered plant species, Dracocephalum austriacum L. − Conserv. Genet. 11: 773−783.
Dostálek t. & MünzberGová z. (2009): Comparative population biology of critically endangered Dracocephalum austriacum L. (Lamiaceae) in two distant regions. − In: Dostálek t., Population biology and genetics of rare plant species, Ms. [Doctoral thesis; depon. in: Department of Botany, Faculty of science, Charles University in Prague.]

78
Dostálek & Münzbergová: Populační dynamika a genetická diverzita Dracocephalum austriacum
eckstein r., Danihelka j. & otte a. (2009): Variation in life-cycle between three rare and endangered floodplain violets in two regions: implications for population viability and conservation. − Biologia 64: 69−80.
ehrlén j., syrjaenen k., leiMu r., Garcia M. b. & lehtilae k. (2005): Land use and population growth of Primula veris: an experimental demographic approach. − Ecology 42: 317–326.
euroPean coMMission (1992): Directive 92/43/EEC of the European Parliament and of the Council of 21st May 1992: conservation of natural habitats and of wild fauna and flora. − Official Journal of the European Communities L206: 7–50.
eXcoffier l., laval G. & schneiDer s. (2005): Arlequin (version 3.0): an integrated software package for population genetics data analysis. − Evol. Bioinform. Online 1: 47−50.
fauGeron s., Martínez e. a., correa j. a., carDenas l., DestoMbe c. & valero M. (2004): Reduced genetic diversity and increased population differentiation in peripheral and overharvested populations of Gigartina skottsbergii (Rhodophyta, Gigartinales) in southern Chile. − J. Phycol. 40: 454–462.
fischer M. & Matthies D. (1998): RAPD variation in relation to population size and plant fitness in the rare Gentianella germanica (Gentianaceae). − Am. J. Bot. 85: 811−819.
frankhaM r. & ralls k. (1998): Conservation biology: inbreeding leads to extinction. − Nature 392: 441–442.
frankhaM r., ballou j. D., briscoe D. a. & Mcinness k. h. (2002): Introduction to conservation genetics. – Cambridge University Press.
frankhaM r. (2003): Genetics and conservation biology. – C. R. Biol. 326: 22–29. García D., zaMora r., GoMez j. M., jorDano P. & hoDar j. a. (2000): Geographical variation in seed
production, predation and abortion in Juniperus communis throughout its range in Europe. − J. Ecol. 88: 436−446.
GauDeul M., taberlet P. & till-bottrauD i. (2000): Genetic diversity in an endangered alpine plant, Eryngium alpinum L. (Apiaceae), inferred from amplified fragment length polymorphism markers. − Mol. Ecol. 9: 1625−1637.
GléMin s., GauDe t., GuilleMin M., lourMas M., olivieri i. & MiGnot a. (2005): Balancing selection in the wild: testing population genetics theory of self-incompatibility in the rare species Brassica insularis. − Genetics 171: 279−289.
GoDt M. j. w., caPlow f. & haMrick j. (2005): Allozyme diversity in the federally threatened golden paintbrush, Castilleja levisecta (Scrophulariaceae). − Conserv. Genet. 6: 87−99.
haMerský R. (2000): Dracocephalum austriacum L. − In: Natura 2000 − mapování a botanický průzkum lokalit ohrožených druhů vyšších rostlin (České středohoří). Ms. [Depon. in: Agentura ochrany přírody a krajiny ČR, Praha.]
haMrick j. l. & GoDt M. j. w. (1989): Allozyme diversity in plant species. − In: brown a. h. D., cleGG M. t., kahler a. l. & weir b. s. [eds], Plant population genetics, breeding and genetic resources, pp. 43–63, Sinauer Associates, Sunderland, Massachusetts, USA.
haMrick j. l., Murawski D. a. & loveless M. D. (1991): Correlations between species traits and allozyme diversity: implications for conservation biology. − In: falk D. a. & holsinGer k. e. [eds], Genetics and conservation of rare plants, pp. 75–86, Oxford University Press, New York.
haMrick j. l. & GoDt M. j. w. (1996): Effects of life history traits on genetic diversity in plant species. − Phil. Trans. R. Soc. Lond. B. 351: 1291−1298.
hanDlová v. & MünzberGová z. (2006): Seed banks of managed and degraded grasslands in the Krkonoše Mts., Czech Republic. − Folia Geobot. 41: 275−288.
hensen i. & oberPrieler c. (2005): Effects of population size on genetic diversity and seed production in the rare Dictamnus albus (Rutaceae) in central Germany. − Conserv. Genet. 6: 63−73.
heufler L. (1851): Trattnick’s briefwechsel. − Oesterr. Bot. Wochenbl. 1: 157–167.heywooD v. h. (1980): Dracocephalum austriacum L. − In: tutin t. G., chater a. o., heywooD v. h.,
burGes n. a., richarDson i. b. k., valentine D. h., walters s. M. & webb D. a. [eds], Flora Europaea, Vol. 3., p. 161, Cambridge University Press, London, UK.
holub j. & Procházka f. (2000): Red List of vascular plants of the Czech Republic − 2000. − Preslia 72: 187–230.
honnay o., bossuyt b., jacQueMyn h., shiMono a. & uchiyaMa k. (2008): Can a seed bank maintain the genetic variation in the above ground plant population? − Oikos 117: 1−5.

79
Příroda, Praha, 31: 53–83, 2011
hrouDa l. (2000): Dracocephalum austriacum L. − In: slavík b. [ed.], Květena České republiky 6, pp. 638–639, Academia, Praha.
hrouDa l. (2002): Dracocephalum austriacum L. − In: kubát k., hrouDa l., chrtek j., kaPlan z., kirschner j. & ŠtěPánek j. [eds], Klíč ke květeně České republiky, p. 592, Academia, Praha.
husbanD b. & scheMske D. (1996): Evolution of the magnitude and timing of inbreeding depression in plants. − Evolution 50: 54–70.
inGvarsson P. k. (2001): Restoration of genetic variation lost – the genetic rescue hypothesis. − Trends Ecol. Evol. 16: 62−63.
ishihaMa f., ueno s., tsuMura y. & washitani i. (2005): Gene flow and inbreeding depression inferred from fine-scale genetic structure in an endangered heterostylous perennial, Primula sieboldii. − Mol. Ecol. 14: 983−990.
kePhart s., brown e. & hall j. (1999): Inbreeding depression and partial selfing: evolutionary implications of mixed-mating in a coastal endemic, Silene douglasii var. oraria (Caryophyllaceae). − Heredity 82: 543–554.
kirchner f., robert a. & colas b. (2006): Modelling the dynamics of introduced populations in the narrow-endemic Centaurea corymbosa: a demo-genetic integration. − J. Appl. Ecol. 43: 1011−1021.
kubíková j. (1993): Survival of two endangered plant species (Dracocephalum austriacum and Gagea bohemica) in the surroundings of the City of Prague. − Novit. Bot. Univ. Carol. 7 (1991–1992): 75–86.
laMMi a., siikaMaki P. & Mustajarvi k. (1999): Genetic diversity, population size, and fitness in central and peripheral populations of a rare plant Lychnis viscaria. − Conserv. Biol. 13: 1069−1078.
lóPez-Pujol j., bosch M., siMon j. & blanché c. (2003): Population genetics and conservation priorities for the critically endangered island endemic Delphinium pentagynum subsp. formenteranum (Ranunculaceae). − Biodivers. Conserv. 12: 1937−1951.
lóPez-Pujol j., orellana M. r., bosch M., siMon j. & blanché c. (2007): Low genetic diversity and allozymic evidence for autopolyploidy in the tetraploid Pyrenean endemic larkspur Delphinium montanum (Ranunculaceae). − Bot. J. Linn. Soc. 155: 211−222.
lowe D. a., harris s., harris s. e. & ashton D. P. (2004): Ecological genetics: design, analysis, and application. − Blackwell Publishing, Oxford.
lloyD D. G. & schoen D. j. (1992): Self-and cross-fertilization in plants. I. Functional dimensions. − Int. J. Plant Sci. 153: 358–369.
ložek v. (2003): Lokality včelníku rakouského (Dracocephalum austriacum L.) v Českém krasu v letech 1983–2003. − Ms. [Depon. in: Správa Chráněné krajinné oblasti Český kras, Karlštejn.]
luijten s. h., Dierick a., oosterMeijer j. G. b., raijMann l. e. l. & Den nijs j. c. M. (2000): Population size, genetic variation, and reproductive success in a rapidly declining, self-incompatible perennial (Arnica montana) in the Netherlands. − Conserv. Biol. 14: 1776–1787.
luijten s. h., kery M., oosterMeijer j. G. b. & Den nijs h. c. M. (2002): Demographic consequences of inbreeding and outbreeding in Arnica montana: a field experiment. − J. Ecol. 90: 593−603.
Machová i. & kubát k. (2004): Zvláště chráněné a ohrožené druhy rostlin Ústecka. – Academia. Praha.ManDák b., bíMová k., Mahelka v. & PlaČková i. (2006): How much genetic variation is stored in the
seed bank? A study of Atriplex tatarica (Chenopodiaceae). − Mol. Ecol. 15: 2653−2663.Mantel n. (1967): The detection of disease clustering and a generalized regression approach. −
Cancer Res. 27: 209.Marrero-GóMez M. v., oosterMeijer j. G. b., carQué-álaMo e. & bañares-bauDet á. (2007): Population
viability of the narrow endemic Helianthemum juliae (Cistaceae) in relation to climate variability. − Biol. Conserv. 136: 552−562.
Martínez-Palacios a., eGuiarte l. e. & furnier G. r. (1999): Genetic diversity of the endangered endemic Agave victoriae-reginae (Agavaceae) in the Chihuahuan Desert. − Am. J. Bot. 86: 1093−1098.
Meusel h., jäGer e., rauschert s. & weinert e. (1978): Vergleichende Chorologie der zentraleuropäischen Flora. II. Karten. – Gustav Fischer Verlag, Jena, Germany.
Moucha P. (1986): Záchrana včelníku rakouského a zvonovce liliolistého. – In: anonyMus, Problematika záchrany ohrožených druhů rostlin, pp. 90–92, Praha.
Moucha P. (1990): Včelník rakouský. − Nika, Praha, 11: 34.

80
Dostálek & Münzbergová: Populační dynamika a genetická diverzita Dracocephalum austriacum
MünzberGová z. (2004): Effect of spatial scale on factors limiting species distributions in dry grassland fragments. − J. Ecol. 92: 854−867.
MünzberGová z. (2005): Determinants of species rarity: population growth rates of species sharing the same habitat. − Am. J. Bot. 92: 1987−1994.
nantel P. & GaGnon D. (1999): Variability in the dynamics of northern peripheral versus southern populations of two clonal plant species, Helianthus divaricatus and Rhus aromatica. − J. Ecol. 87: 748−760.
neel M. c. & cuMMinGs M. P. (2003): Effectiveness of conservation targets in capturing genetic diversity. − Conserv. Biol. 17: 219−229.
neel M. c. & ellstranD n. c. (2003): Conservation of genetic diversity in the endangered plant Eriogonum ovalifolium var. vineum (Polygonaceae). − Conserv. Genet. 4: 337−352.
nevole j. (1910): Studien über die Verbreitung von sechs südeuropäischen Pflanzenarten. − Mitteilungen des naturwissenschaftlichen Vereines für Steiermark 46: 3–25.
newMan D. & Pilson D. (1997): Increased probability of extinction due to decreased genetic effective population size: experimental populations of Clarkia pulchella. − Evolution 51: 354−362.
nicolè f. (2005): Biologie de la conservation appliquée aux plantes menacées des Alpes. – Ms. [Doctoral thesis, depon in: de l’Université Joseph Fourier, Grenoble I.]
nicolè f., DahlGren j. P., vivat a., till-bottrauD i. & ehrlén j. (2011): Interdependent effects of habitat quality and climate on population growth of an endangered plant. − J. Ecol. 99: 1211–1218.
oosterMeijer j. G. b. (2000): Population viability analysis of the rare Gentiana pneumonanthe: the importance of genetics, demography and reproductive biology. − In: younG a. G. & clarke G. M. [eds], Genetics, demography and viability of fragmented populations, pp. 313–334, Cambridge University Press, UK.
oosterMeijer j. G. b., luijten s. h. & Den nijs j. c. M. (2003): Integrating demographic and genetic approaches in plant conservation. − Biol. Conserv. 113: 389−398.
Petit r. j., MousaDik a. e. & Pons o. (1998): Identifying populations for conservation on the basis of genetic markers. − Conserv. Biol. 12: 844−855.
Picó f. X., jonGejans e., Quintana-ascencio P. f. & De kroon h. (2004): Inbreeding depression significantly influences population dynamics: a population-based model for a long-lived perennial herb. − In: jonGejans e., Life history strategies and biomass allocation. Ms. [Doctoral thesis, Wageningen University, The Netherlands.]
Pino j., Picó f. X. & De roa e. (2007): Population dynamics of the rare plant Kosteletzkya pentacarpos (Malvaceae): a nine-year study. − Bot. J. Linn. Soc. 153: 455−462.
Puska V. (2006): Az osztrák sárkányfű (Dracocephalum austriacum L.) Vizsgálata. − Ms. [Diploma thesis, Debreceni Egyetem Természettudományi Kar Növénytani Tanszék, Debrecen.]
reeD D. h. & frankhaM r. (2003): Correlation between fitness and genetic diversity. − Conserv. Biol. 17: 230−237.
saccheri i., kuussaari M., kankare M., vikMan P., fortelius w. & hanski i. (1998): Inbreeding and extinction in a butterfly metapopulation. − Nature 392: 491–494.
schMiDt k. & jensen k. (2000): Genetic structure and AFLP variation of remnant populations in the rare plant Pedicularis palustris (Scrophulariaceae) and its relation to population size and reproductive components 1. − Am. J. Bot. 87: 678–689.
silvertown j., franco M., Pisanty i. & MenDoza a. (1993): Comparative plant demography−relative importance of life-cycle components to the finite rate of increase in woody and herbaceous perennials. − J. Ecol. 81: 465−476.
skibinski D., wooDwark M. & warD r. D. (1993): A quantitative test of the neutral theory using pooled allozyme data. − Genetics 135: 233−248.
soltis D. e. & soltis o. s. (1989): Isozymes in plant biology. − Dioscorides Press, Portland.s−Plus (2000): Professional edition for Windows, release 2. − MathSoft, Inc., Massachusetts.storfer a. (1996): Quantitative genetics: a promising approach for the assessment of genetic variation
in endangered species. − Trends Ecol. Evol. 11: 343–348.thoMPson k., bakker j. P. & bekker r. M. (1997): The soil seed banks of north west Europe:
methodology, density and longevity. − Cambridge University Press, UK.

81
Příroda, Praha, 31: 53–83, 2011
treMlová k. & MünzberGová z. (2007): Importance of species traits for species distribution in fragmented landscapes. − Ecology 88: 965–977.
valouŠek B. (1928): Včelník rakouský (Dracocephalum austriacum L.) − nová rostlina na Moravě. − Příroda 21: 258−260.
van buren r. & harPer k. t. (2003): Demographic and environmental relations of two rare Astragalus species endemic to Washington County, Utah: Astragalus holmgreniorum and A. ampullarioides. − West. N. Am. Naturalist 63: 236–243.
warDle G. M. (2003): Experimental determination of seed emergence and carry-over in the soil seed bank of the herbaceous perennial, Trachymene incisa (Apiaceae). − Austral Ecol. 28: 161−172.
wiDén b. & wiDén M. (1990): Pollen limitation and distance-dependent fecundity in females of the clonal gynodioecious herb Glechoma hederacea (Lamiaceae). − Oecologia 83: 191−196.
willeMs j. & ellers j. (1996): Plant performance and population characteristics of Orchis simia (Orchidaceae) in two extremes of its distribution area. − Flora 191: 41−48.
virók V. (2006): Osztrák sárkányfű „Dracocephalum austriacum” – KvVm Természetvédelmi Hivatal Fajmegőrzési Tervek – Környezetvédelmi és Vízügyi Minisztérium, Természetvédelmi Hivatal Kiadója.
yeh f. c. & boyle t. j. b. (1997): Population genetic analysis of co-dominant and dominant markers and quantitative traits. − Belg. J. Bot. 129: 157.
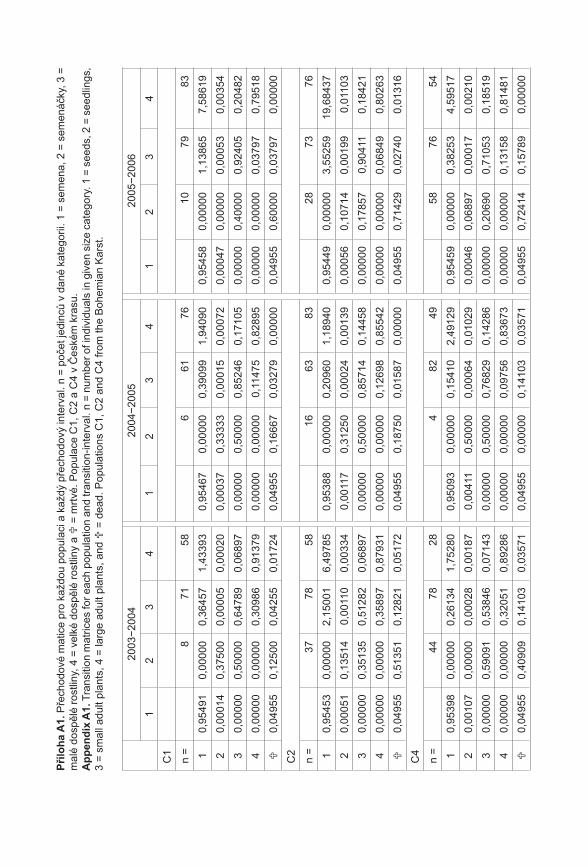
82
Dostálek & Münzbergová: Populační dynamika a genetická diverzita Dracocephalum austriacum
Přílo
ha A
1. P
řech
odov
é m
atic
e pr
o ka
ždou
pop
ulac
i a k
aždý
pře
chod
ový
inte
rval
. n =
poč
et je
dinc
ů v
dané
kat
egor
ii. 1
= s
emen
a, 2
= s
emen
áčky
, 3 =
m
alé
dosp
ělé
rost
liny,
4 =
vel
ké d
ospě
lé ro
stlin
y a U
= m
rtvé.
Pop
ulac
e C
1, C
2 a
C4
v Č
eské
m k
rasu
.A
ppen
dix
A1.
Tra
nsiti
on m
atric
es fo
r eac
h po
pula
tion
and
trans
ition
-inte
rval
. n =
num
ber o
f ind
ivid
uals
in g
iven
siz
e ca
tego
ry. 1
= s
eeds
, 2 =
see
dlin
gs,
3 =
smal
l adu
lt pl
ants
, 4 =
larg
e ad
ult p
lant
s, a
nd U
= d
ead.
Pop
ulat
ions
C1,
C2
and
C4
from
the
Boh
emia
n K
arst
.
2003
−200
420
04−2
005
2005
−200
6
12
34
12
34
12
34
C1
n =
871
586
6176
1079
83
10,
9549
10,
0000
00,
3645
71,
4339
30,
9546
70,
0000
00,
3909
91,
9409
00,
9545
80,
0000
01,
1386
57,
5861
9
20,
0001
40,
3750
00,
0000
50,
0002
00,
0003
70,
3333
30,
0001
50,
0007
20,
0004
70,
0000
00,
0005
30,
0035
4
30,
0000
00,
5000
00,
6478
90,
0689
70,
0000
00,
5000
00,
8524
60,
1710
50,
0000
00,
4000
00,
9240
50,
2048
2
40,
0000
00,
0000
00,
3098
60,
9137
90,
0000
00,
0000
00,
1147
50,
8289
50,
0000
00,
0000
00,
0379
70,
7951
8
U0,
0495
50,
1250
00,
0425
50,
0172
40,
0495
50,
1666
70,
0327
90,
0000
00,
0495
50,
6000
00,
0379
70,
0000
0
C2
n =
3778
5816
6383
2873
76
10,
9545
30,
0000
02,
1500
16,
4978
50,
9538
80,
0000
00,
2096
01,
1894
00,
9544
90,
0000
03,
5525
919
,684
37
20,
0005
10,
1351
40,
0011
00,
0033
40,
0011
70,
3125
00,
0002
40,
0013
90,
0005
60,
1071
40,
0019
90,
0110
3
30,
0000
00,
3513
50,
5128
20,
0689
70,
0000
00,
5000
00,
8571
40,
1445
80,
0000
00,
1785
70,
9041
10,
1842
1
40,
0000
00,
0000
00,
3589
70,
8793
10,
0000
00,
0000
00,
1269
80,
8554
20,
0000
00,
0000
00,
0684
90,
8026
3
U0,
0495
50,
5135
10,
1282
10,
0517
20,
0495
50,
1875
00,
0158
70,
0000
00,
0495
50,
7142
90,
0274
00,
0131
6
C4
n =
4478
284
8249
5876
54
10,
9539
80,
0000
00,
2613
41,
7528
00,
9509
30,
0000
00,
1541
02,
4912
90,
9545
90,
0000
00,
3825
34,
5951
7
20,
0010
70,
0000
00,
0002
80,
0018
70,
0041
10,
5000
00,
0006
40,
0102
90,
0004
60,
0689
70,
0001
70,
0021
0
30,
0000
00,
5909
10,
5384
60,
0714
30,
0000
00,
5000
00,
7682
90,
1428
60,
0000
00,
2069
00,
7105
30,
1851
9
40,
0000
00,
0000
00,
3205
10,
8928
60,
0000
00,
0000
00,
0975
60,
8367
30,
0000
00,
0000
00,
1315
80,
8148
1
U0,
0495
50,
4090
90,
1410
30,
0357
10,
0495
50,
0000
00,
1410
30,
0357
10,
0495
50,
7241
40,
1578
90,
0000
0
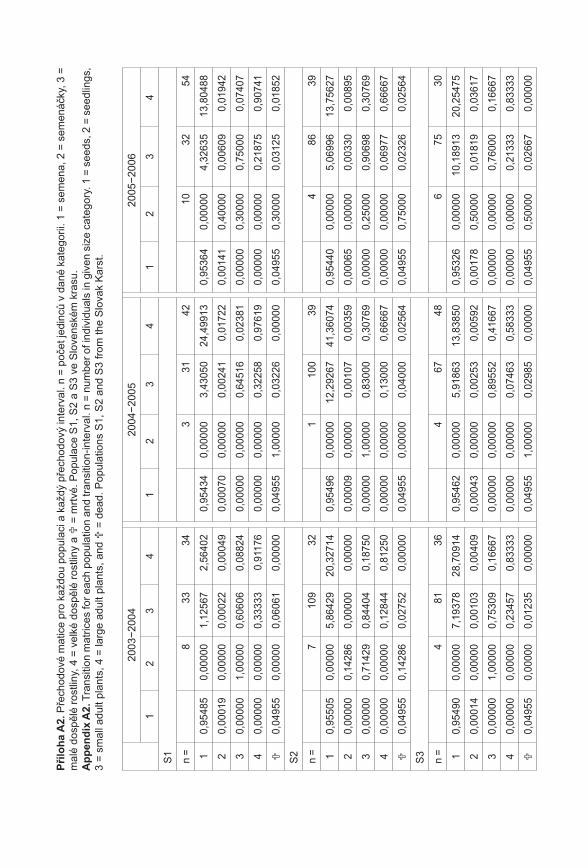
83
Příroda, Praha, 31: 53–83, 2011Př
íloha
A2.
Pře
chod
ové
mat
ice
pro
každ
ou p
opul
aci a
kaž
dý p
řech
odov
ý in
terv
al. n
= p
očet
jedi
nců
v da
né k
ateg
orii.
1 =
sem
ena,
2 =
sem
enáč
ky, 3
=
mal
é do
spěl
é ro
stlin
y, 4
= v
elké
dos
pělé
rost
liny
a U
= m
rtvé.
Pop
ulac
e S
1, S
2 a
S3
ve S
love
nské
m k
rasu
.A
ppen
dix
A2.
Tra
nsiti
on m
atric
es fo
r eac
h po
pula
tion
and
trans
ition
-inte
rval
. n =
num
ber o
f ind
ivid
uals
in g
iven
siz
e ca
tego
ry. 1
= s
eeds
, 2 =
see
dlin
gs,
3 =
smal
l adu
lt pl
ants
, 4 =
larg
e ad
ult p
lant
s, a
nd U
= d
ead.
Pop
ulat
ions
S1,
S2
and
S3
from
the
Slo
vak
Kar
st.
2003
−200
420
04−2
005
2005
−200
6
12
34
12
34
12
34
S1 n =
833
343
3142
1032
54
10,
9548
50,
0000
01,
1256
72,
5640
20,
9543
40,
0000
03,
4305
024
,499
130,
9536
40,
0000
04,
3263
513
,804
88
20,
0001
90,
0000
00,
0002
20,
0004
90,
0007
00,
0000
00,
0024
10,
0172
20,
0014
10,
4000
00,
0060
90,
0194
2
30,
0000
01,
0000
00,
6060
60,
0882
40,
0000
00,
0000
00,
6451
60,
0238
10,
0000
00,
3000
00,
7500
00,
0740
7
40,
0000
00,
0000
00,
3333
30,
9117
60,
0000
00,
0000
00,
3225
80,
9761
90,
0000
00,
0000
00,
2187
50,
9074
1
U0,
0495
50,
0000
00,
0606
10,
0000
00,
0495
51,
0000
00,
0322
60,
0000
00,
0495
50,
3000
00,
0312
50,
0185
2
S2 n =
710
932
110
039
486
39
10,
9550
50,
0000
05,
8642
920
,327
140,
9549
60,
0000
012
,292
6741
,360
740,
9544
00,
0000
05,
0699
613
,756
27
20,
0000
00,
1428
60,
0000
00,
0000
00,
0000
90,
0000
00,
0010
70,
0035
90,
0006
50,
0000
00,
0033
00,
0089
5
30,
0000
00,
7142
90,
8440
40,
1875
00,
0000
01,
0000
00,
8300
00,
3076
90,
0000
00,
2500
00,
9069
80,
3076
9
40,
0000
00,
0000
00,
1284
40,
8125
00,
0000
00,
0000
00,
1300
00,
6666
70,
0000
00,
0000
00,
0697
70,
6666
7
U0,
0495
50,
1428
60,
0275
20,
0000
00,
0495
50,
0000
00,
0400
00,
0256
40,
0495
50,
7500
00,
0232
60,
0256
4
S3 n =
481
364
6748
675
30
10,
9549
00,
0000
07,
1937
828
,709
140,
9546
20,
0000
05,
9186
313
,838
500,
9532
60,
0000
010
,189
1320
,254
75
20,
0001
40,
0000
00,
0010
30,
0040
90,
0004
30,
0000
00,
0025
30,
0059
20,
0017
80,
5000
00,
0181
90,
0361
7
30,
0000
01,
0000
00,
7530
90,
1666
70,
0000
00,
0000
00,
8955
20,
4166
70,
0000
00,
0000
00,
7600
00,
1666
7
40,
0000
00,
0000
00,
2345
70,
8333
30,
0000
00,
0000
00,
0746
30,
5833
30,
0000
00,
0000
00,
2133
30,
8333
3
U0,
0495
50,
0000
00,
0123
50,
0000
00,
0495
51,
0000
00,
0298
50,
0000
00,
0495
50,
5000
00,
0266
70,
0000
0

84
Dostálek & Münzbergová: Populační dynamika a genetická diverzita Dracocephalum austriacum

85
Příroda, Praha, 31: 85–109, 2011
vliv obhospodařování na životní cyklus hořečku mnohotvarého českého (Gentianella praecox subsp. bohemica)
Effect of management on life cycle of Gentianella praecox subsp. bohemica
Jiří brabec1, anna bucharová2, 3 & MichaL štefánek3, 4
1Muzeum Cheb, příspěvková organizace Karlovarského kraje, Krále Jiřího z Poděbrad 493/4, CZ-350 11 Cheb, [email protected]ý ústav Akademie věd České republiky, v. v. i., Zámek 1, CZ-252 43 Průhonice, [email protected] botaniky Přírodovědecké fakulty Univerzity Karlovy, Benátská 2, CZ-128 01 Praha 24Herbářové sbírky Univerzity Karlovy, Benátská 2, CZ-128 01 Praha 2, [email protected]
Abstract: Through the evaluation of a detailed experimental description of the population biology of Gentianella praecox subsp. bohemica on four localities from the centre of the species area, we were able: (i) to examine the effects of different management regimes on the survival of the biennial G. praecox subsp. bohemica; (ii) to determine factors influencing the individual phases of the plant’s life cycle; (iii) to quantify the size and germination of the plant’s seed bank; (iv) to suggest appropriate management for the localities with G. praecox subsp. bohemica. Populations of G. praecox subsp. bohemica clearly show the best viability at the experimental sites managed by the combination of grass mowing and disturbance of vegetation by raking, lower viability at the sites managed by grass mowing only, and the worst viability on the control sites without any management. Seedling emergence is proven to be the phase of the plant’s life cycle most affected by the type of management. The first summer period of the biennial plant’s life cycle, the seedlings at the experimental sites managed by mowing survive best; plants at sites managed by mowing and disturbances slightly worse and those on the control sites show the worst survival rate. This trend could be affected by extreme conditions like the 2003 droughts in our case. There is evidence of a long-lasting viable seed bank of G. praecox subsp. bohemica. Each year, the amount of viable seeds buried in the seed bank dropped by one half, however, we proved that the seeds are able to emerge even after eight years. Under optimal conditions (experimental site 15 × 15 cm, grass mowed and vegetation disturbed by raking before sowing) the mean number of seeds emerging during the first year is around 15.7%. Among the rest of the seeds remaining in the seed bank, around 1.6% emerge over the next three years.Keywords: biennial, conservation action plan, critical phases of the life cycle, grassland management, population biology, seedbank.
Abstrakt: Vyhodnocení podrobného experimentálního popisu populační biologie hořečku mnohotvarého českého (Gentianella praecox subsp. bohemica) na čtyřech lokalitách v centru areálu taxonu umožnilo (a) stanovit vliv různých typů obhospodařování na přežívání těchto dvouletých rostlin, (b) určit faktory ovlivňující jednotlivé fáze životního cyklu, (c) kvantifikovat velikost a klíčivost semenné banky a (d) stanovit doporučení pro další obhospodařování lokalit. Populace hořečků jednoznačně nejlépe přežívají na plochách

86
Brabec et al.: Vliv obhospodařování na cyklus Gentianella praecox subsp. bohemica
obhospodařovaných sečí spojenou s následnou disturbancí zápoje vegetace, o něco hůře v plochách pouze sečených a nejhůře v neobhospodařovaných kontrolních plochách. Fází životního cyklu, která tyto rozdíly způsobuje, je prokazatelně vzcházení. První letní období dvouletého životního cyklu přežívají nejlépe semenáčky v sečených plochách, o něco hůře v sečených s disturbancí a nejhůře v kontrolních. Tento mírný trend však narušují následky extrémních událostí, v našem případě sucha v roce 2003. Experimentálně byla u G. praecox subsp. bohemica prokázána dlouhodobě přetrvávající semenná banka. Počet zachovalých semen v pohřbené semenné bance klesá každoročně zhruba na polovinu předchozího roku, nicméně ještě po osmi letech se podařilo prokázat klíčení. Za optimálních podmínek na lokalitě (experimentální ploška 15 × 15 cm ošetřená sečí a disturbancí před vysetím semen) klíčí v následujícím roce průměrně ca 15,7 % semen. Ze semen, která první rok nevzejdou, vyklíčí následující tři roky (tj. ze semenné banky) ca 1,6 % semen.Klíčová slova: dvouletý druh, kritické fáze životního cyklu, management travinných porostů, populační biologie, semenná banka, záchranný program.nomenklatura: kubát et al. (2002) - vyšší rostliny, Moravec et al. (1995) – rostlinná společenstva
Úvod
U mnoha rostlin evropských pastvin a luk bylo v posledních desetiletích zaznamenáno radikální snížení počtu a velikosti lokalit (fischer & stöcklin 1997, Procházka 2001). Tyto změny jsou tradičně přičítány celkovému úbytku a fragmentaci ploch pastevních a lučních enkláv, změně agrotechnických postupů a fragmentaci areálů populací jednotlivých druhů (PoschloD & wallis De vries 2002, eriksson et al. 2002, luoto et al. 2003, foley et al. 2005). Zásadní otázkou pro ochranu přírody je, jak se zachovat ke zbylým populacím, zda a jak provádět cílený management k jejich ochraně. Převedeno do pragmatické řeči peněz, kdy ještě investovat do jejich záchrany.
Taxony rodu hořeček (Gentianella) patří v Evropě k typickým příkladům krátkověkých (jednoletých či dvouletých) druhů travinných společenstev, které se z běžně rozšířených rostlin staly v mnoha oblastech extrémně vzácnými. Na území České republiky bylo historicky věrohodně zaznamenáno celkem 12 taxonů rodu Gentianella (kirschner & kirschnerová 2000). Z toho je v současné době pět taxonů vyhynulých nebo nezvěstných. Zbylých sedm patří mezi kriticky ohrožené taxony (Procházka 2001), u kterých dochází v posledních desetiletích k výraznému snížení počtu lokalit (kirschner & kirschnerová 2000, 2003, brabec 2005, 2010a, b, c, d, kirschnerová & kirschner 1997, kolbek et al. 1994, PoPelářová et al. 2008, soMol et al. 1999, zahraDníková 1995). Z těchto sedmi taxonů zůstávají pouze dva, u kterých je možné vzhledem k počtu lokalit a velikosti populací provádět rozsáhlejší experimentální studium. Jsou jimi hořeček nahořklý pravý (Gentianella amarella subsp. amarella) mající velký euroasijský areál a endemit Českého masivu hořeček mnohotvarý český (Gentianella praecox subsp. bohemica). Ten je velmi vhodným modelem i pro další blízce příbuzné druhy (např. alpsko-západoevropský taxon G. germanica, východoalpský G. austriaca, alpsko-subatlantský G. obtusifolia, karpatsko-balkánský G. lutescens).
Stanovení vhodného obhospodařování lokalit s výskytem vzácných druhů rostlin je většinou prováděno subjektivně. Způsob a intenzita obhospodařování (seč, pastva, vláčení) jsou nejčastěji určovány na základě typu porostu, znalosti tradičních zemědělských postupů v daném regionu a příležitostném či pravidelném monitoringu populací vzácných druhů. To je u druhů s podobnou biologií a ekologií jako Gentianella praecox subsp. bohemica spojeno s celou řadou obtíží:

87
Příroda, Praha, 31: 85–109, 2011
1)Studovaný druh (podobně jako většina ostatních druhů rodu) není příliš vázán na určité typy vegetace (brabec 2005, 2010d, J. Brabec, Z. Křenová & S. Rösler, nepublikované údaje). Roste zejména na pastvinách, podhorských a horských smilkových loukách (tzn. společenstva svazu Cynosurion, Violion caninae a Nardion); dále roste v některých typech sečených mezických a vysýchavých mokřadních luk (společenstva svazu Arrhenatherion a Molinion) a v některých porostech širolistých suchých trávníků a pastvin (společenstva svazů Bromion erecti a Koelerio-Phleion phleoidis). Roste také na mnoha typech narušovaných stanovišť (například na okrajích cest, při hranách lomů a v lesních lemech). Druh nevykazuje ani žádné výraznější nároky ve vazbě na substrát a živiny. Roste od velmi kyselých půd po půdy vápnité, včetně půd s vysokým obsahem iontů Mg2+ (brabec 2010d, J. Brabec, Z. Křenová & S. Rösler, nepublikované údaje).
2)Přestože jde o dvouletý druh, jsou v porostech viditelné a monitorovatelné pouze exempláře, které dospěly až do konce životního cyklu, tedy do stádia kvetení a zrání. Vliv obhospodařování na populaci druhu tak nemusí být na první pohled vůbec patrný.
3)Dvouleté taxony rodu Gentianella (G. praecox subsp. bohemica nevyjímaje) se vyznačují značnou meziroční fluktuací v počtu kvetoucích exemplářů (např. brabec 2010a). Sezóny, kdy na lokalitách v celém areálu kvete několikanásobně více hořečků než je průměr posledních let, nazýváme tzv. „hořečkovými roky“. Za posledních deset let jsme u G. praecox subsp. bohemica zaznamenali tři „hořečkové roky“ – 2002, 2005 a 2009 (brabec 2010c, d). U taxonu tak zjevně hraje obrovskou roli semenná banka (bucharová et al. 2011).
Z výše uvedeného vyplývá, že základní faktory ovlivňující přežívání populací G. praecox subsp. bohemica je nutno hledat ve struktuře porostů, nikoli v druhovém složení vegetace. Zároveň je ke stanovení dlouhodobě optimálního a zároveň co nejméně nákladného obhospodařování lokalit potřeba kvantifikovat vliv různých opatření na jednotlivé fáze dvouletého životního cyklu.
Cílem této práce je doplnit a stručně popsat životní cyklus G. praecox subsp. bohemica, určit faktory ovlivňující jednotlivé fáze jeho životního cyklu, odhadnout velikost a klíčivost semenné banky a stanovit vliv různých typů obhospodařování lokalit na přežívání rostlin tohoto taxonu. S ohledem na výsledky práce jsou také stanovena a diskutována doporučení pro obhospodařování lokalit hořečku mnohotvarého českého.
Materiál a metodika
taxon Gentianella praecox subsp. bohemica
Taxon byl popsán v České republice v roce 1969 (skalický 1969) jako druh Gentianella bohemica skalický rozlišený od taxonů G. germanica (willD.) börner a G. austriaca (a. et j. kern.) holub. Typová položka uložená v herbáři Univerzity Karlovy v Praze (PRC) pochází z Pošumaví od Hor Matky Boží (skalický 1969: „na pastvinách mezi městečkem Hory Matky Boží blízko Sušice a vrchem Křížovka, ca 700 m, 18. 8. 1966, leg. V. Skalický et A. Skalická“). Na této lokalitě však nebyl hořeček již mnoho let zaznamenán (K. Čížek, úst. sdělení, M. Štech, úst. sdělení, poslední údaj zřejmě skalický & sofron 1990). V současné době je taxon zcela akceptován současnými zpracovateli rodu Gentianella ve střední Evropě, a to jak na úrovni druhové (wisskirchen & haeuPler 1998, haeuPler

88
Brabec et al.: Vliv obhospodařování na cyklus Gentianella praecox subsp. bohemica
& Muer 2000, jäGer & werner 2002, zając & zając 2001), tak na úrovni poddruhové (kirschner & kirschnerová 2000, kirschnerová 2002, fischer et al. 2008).
Hořeček mnohotvarý český je dvouletá bylina s přímou, jednoduchou nebo častěji již od báze vrcholičnatě větvenou lodyhou (tvar a větvení se mění po poškození rostliny, tj. např. po posečení, ukousnutí apod.). Lodyžních článků je obvykle (4–)6–12(–16), ± stejně dlouhých. Přízemní listy (za květu často již odumřelé) jsou až 4,5 cm dlouhé. Květní obaly jsou zpravidla pětičetné, zřídka čtyřčetné. Zářezy mezi kališními cípy jsou proměnlivého tvaru, alespoň některé ve tvaru širokého U, jiné však někdy ve tvaru V. Velikost rostliny i počet květů v květenství je velmi variabilní podle sezóny a typu stanoviště. Výška rostlin kolísá v rozmezí (2–)10–25(–75) cm, počet květů 1–50(–360) (podle kirschner & kirschnerová 2000 a vlastních měření).
Gentianella praecox subsp. bohemica je endemitem Českého masivu a subendemitem České republiky. Historický areál zahrnuje Českou republiku (většinu území kromě západních a severozápadních Čech a jihovýchodní a východní Moravy), severní Rakousko (sever Horních Rakous a severozápad Dolních Rakous), Bavorsko (jen západ Dolního Bavorska) a jižní Polsko (rozšíření není dokonale známé, mapa rozšíření taxonu v zając & zając 2001 je nepřesná, A. Zając, úst. sdělení). Celkový areál taxonu zhruba odpovídá dnešnímu, výrazně se však snížil počet lokalit a velikost populací (brabec 2010d). Na území České republiky druh v minulosti rostl roztroušeně až hojně od nížin (hist. min. – Štětí, ca 170 m n. m.) až do hor (max. historické i recentní – Kvilda, Vilémov, ca 1030 m n. m.) s těžištěm v suprakolinním až submontánním stupni (kirschner & kirschnerová 2000). Několik desítek historických lokalit je známo i z Rakouska, Polska a Bavorska. Od roku 2000 byl (i přes intenzivní pátrání) v celém areálu zaznamenán pouze na 113 lokalitách: Česká republika – 70, Rakousko – 30, Německo – 9, Polsko – 4. Z těchto 113 lokalit se však na 23 lokalitách v posledních pěti letech (tj. 2006 až 2010) neobjevil žádný kvetoucí exemplář (brabec 2010c, enGleDer 2006, 2010, Gerža & ČePa 2007, Gołąb 2004, rösler 2005, ziPP 2006, T. Engleder, úst. sdělení, M. Smoczyk, úst. sdělení).
Experimentální lokality
Kompletní několikaleté sady experimentálních ploch se podařilo založit a vyhodnotit na čtyřech lokalitách v centru historického areálu taxonu v jihozápadních a jižních Čechách. Dvě lokality (Chvalšiny a Hroby) jsou mezické s porosty svazu Arrhenatherion, druhé dvě (Onšovice, Dobročkov) suché s porosty širolistých trávníků svazu Bromion erecti. Lokalizaci a podrobnou charakteristiku lokalit a populací sledovaného druhu přináší tab. 1 a 2.
Experimentální design
Přežívání rostlin v různých typech obhospodařování (experiment 1)
Experiment byl založen na čtyřech lokalitách, a to čtyřikrát až pětkrát během let 2000 až 2005 (viz tab. 3). Původní záměr založit experiment několik let v řadě po sobě se podařil pouze na jedné lokalitě (Chvalšiny). Důvodem byl extrémně suchý rok 2003, během kterého uhynula většina dvouletých i jednoletých rostlin na většině lokalit, což zapříčinilo dramatický propad v počtu kvetoucích a plodících exemplářů v celém areálu taxonu v letech 2003 a 2004 (brabec 2004). Na lokalitách tak nebyl v roce 2003, popř. i 2004 dostatek semen pro experimentální výsevy.
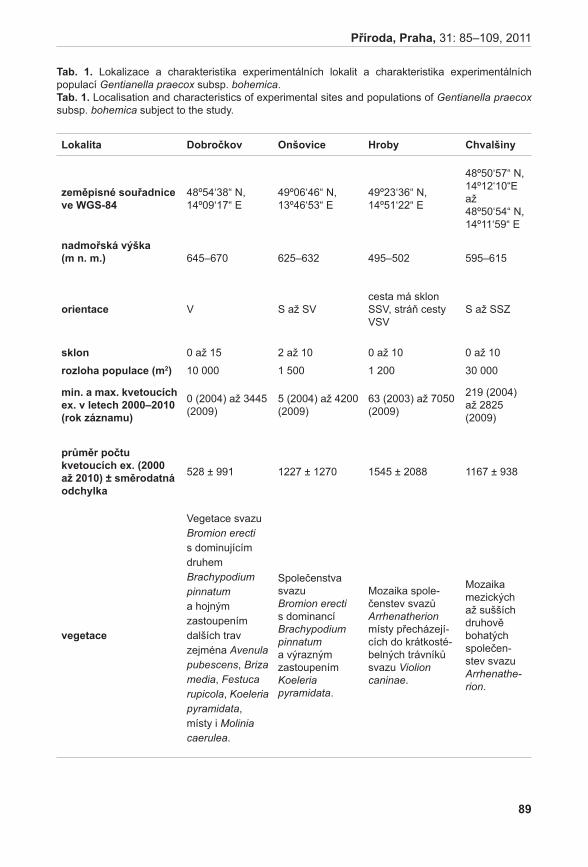
89
Příroda, Praha, 31: 85–109, 2011
tab. 1. Lokalizace a charakteristika experimentálních lokalit a charakteristika experimen tálních populací Gentianella praecox subsp. bohemica.tab. 1. Localisation and characteristics of experimental sites and populations of Gentianella praecox subsp. bohemica subject to the study.
lokalita dobročkov onšovice Hroby chvalšiny
zeměpisné souřadnice ve WGs-84
48º54‘38“ N, 14º09‘17“ E
49º06‘46“ N,13º46‘53“ E
49º23‘36“ N, 14º51‘22“ E
48º50‘57“ N, 14º12‘10“E až48º50‘54“ N, 14º11‘59“ E
nadmořská výška (m n. m.) 645–670 625–632 495–502 595–615
orientace V S až SVcesta má sklon SSV, stráň cesty VSV
S až SSZ
sklon 0 až 15 2 až 10 0 až 10 0 až 10
rozloha populace (m2) 10 000 1 500 1 200 30 000
min. a max. kvetoucích ex. v letech 2000–2010 (rok záznamu)
0 (2004) až 3445 (2009)
5 (2004) až 4200 (2009)
63 (2003) až 7050 (2009)
219 (2004) až 2825 (2009)
průměr počtu kvetoucích ex. (2000 až 2010) ± směrodatná odchylka
528 ± 991 1227 ± 1270 1545 ± 2088 1167 ± 938
vegetace
Vegetace svazu Bromion erecti s dominujícím druhem Brachypodium pinnatum a hojným zastoupením dalších trav zejména Avenula pubescens, Briza media, Festuca rupicola, Koeleria pyramidata, místy i Molinia caerulea.
Společenstva svazu Bromion erecti s dominancí Brachypodium pinnatum a výrazným zastoupením Koeleria pyramidata.
Mozaika spole-čenstev svazů Arrhenatherion místy přecházejí-cích do krátkosté-belných trávníků svazu Violion caninae.
Mozaika mezických až sušších druhově bohatých společen-stev svazu Arrhenathe-rion.
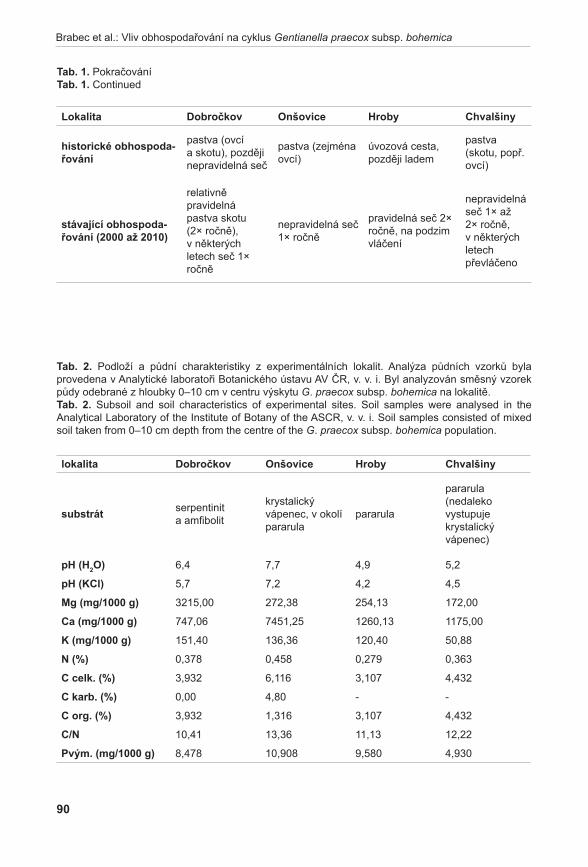
90
Brabec et al.: Vliv obhospodařování na cyklus Gentianella praecox subsp. bohemica
tab. 1. Pokračovánítab. 1. Continued
lokalita dobročkov onšovice Hroby chvalšiny
historické obhospoda-řování
pastva (ovcí a skotu), později nepravidelná seč
pastva (zejména ovcí)
úvozová cesta, později ladem
pastva (skotu, popř. ovcí)
stávající obhospoda-řování (2000 až 2010)
relativně pravidelná pastva skotu (2× ročně), v některých letech seč 1× ročně
nepravidelná seč 1× ročně
pravidelná seč 2× ročně, na podzim vláčení
nepravidelná seč 1× až 2× ročně, v některých letech převláčeno
tab. 2. Podloží a půdní charakteristiky z experimentálních lokalit. Analýza půdních vzorků byla provedena v Analytické laboratoři Botanického ústavu AV ČR, v. v. i. Byl analyzován směsný vzorek půdy odebrané z hloubky 0–10 cm v centru výskytu G. praecox subsp. bohemica na lokalitě.tab. 2. Subsoil and soil characteristics of experimental sites. Soil samples were analysed in the Analytical Laboratory of the Institute of Botany of the ASCR, v. v. i. Soil samples consisted of mixed soil taken from 0–10 cm depth from the centre of the G. praecox subsp. bohemica population.
lokalita dobročkov onšovice Hroby chvalšiny
substrát serpentinit a amfibolit
krystalický vápenec, v okolí pararula
pararula
pararula (nedaleko vystupuje krystalický vápenec)
pH (H2o) 6,4 7,7 4,9 5,2
pH (Kcl) 5,7 7,2 4,2 4,5
Mg (mg/1000 g) 3215,00 272,38 254,13 172,00
ca (mg/1000 g) 747,06 7451,25 1260,13 1175,00
K (mg/1000 g) 151,40 136,36 120,40 50,88
n (%) 0,378 0,458 0,279 0,363
c celk. (%) 3,932 6,116 3,107 4,432
c karb. (%) 0,00 4,80 - -
c org. (%) 3,932 1,316 3,107 4,432
c/n 10,41 13,36 11,13 12,22
Pvým. (mg/1000 g) 8,478 10,908 9,580 4,930

91
Příroda, Praha, 31: 85–109, 2011
Základem experimentálního designu na lokalitě je šest ploch o velikosti 60 cm × 40 cm roz dělených na šest podploch o rozměrech 20 cm × 20 cm (experimentální ploška 15 cm × 15 cm + 2,5 cm nárazníková zóna). Vždy na dvou podplochách byly provedeny tři typy obhospodařování: kontrola (bez managementu), seč (tj. ostříhání = simulace seče) a seč s disturbancí (tj. ostříhání a disturbance železnými hrabičkami = simulace seče a disturbance vláčením). Na ploškách sečených s disturbancí železnými hrabičkami byla kompletně odstraněna veškerá odmuřelá nerozložená stařina a téměř absolutně odstraněna nadzemní vrstva mechorostů. Celková pokryvnost mechového a bylinného patra činila těsně po zásahu 10 až 60 %.
Obhospodařování proběhlo v průběhu října a počátku listopadu po dozrání semen na lokalitě. Vždy na jednu ze shodně obhospodařovaných plošek (15 cm × 15 cm) v každé ploše bylo vyseto 600 zralých semen sebraných na shodné lokalitě. Semena byla vysévána téhož dne nebo následující den po odběru. Hustota 600 semen na plošku 15 cm × 15 cm vychází ze zjištění, že většina zralých semen „padá“ do blízkosti mateřské rostliny, přičemž jedna rostlina G. praecox subsp. bohemica vyprodukuje průměrně 613 semen (J. Brabec, nepublikovaná data). Druhá shodně obhospodařovaná ploška v každé ploše sloužila jako kontrolní plocha pro vzcházení z případné semenné banky. Každý rok výsevu (ROK 0) na lokalitě (viz tab. 3) bylo založeno šest nových experimentálních ploch (šest opakování), kromě výsevu na lokalitě Hroby v roce 2004, kdy byla z důvodu nedostatku semen založena pouze tři opakování (tři plochy). Obhospodařování experimentálních podploch bylo provedeno jednorázově, vždy před výsevem semen. Prvním rokem po vysetí (ROK I) byly podplochy (kromě kontrolních) ostříhány (imitace seče) v průběhu října, druhým rokem (ROK II) pak po odběru všech rostlin ke spočtení produkce semen.
Počet semenáčků a přežívajících rostlin v každé plošce byl po dobu dvou let (tj. po dobu celého životního cyklu dvouletého druhu) zaznamenán vždy třikrát ročně: počátek května (jaro I, jaro II), přelom června a července (léto I, léto II), přelom září a října (podzim I, podzim II). Druhým rokem byly zrající rostliny odebrány a spočtena produkce semen ve všech semenících. Celkem bylo takto hodnoceno sedm přechodů („a“ až „g“ viz obr. 2) v rámci jednotlivých životních fází.
Vliv podzimního obhospodařování na rostliny Gentianella praecox subsp. bohemica (experiment 2)
Experiment ke sledování vlivu podzimního obhospodařování na růžice G. praecox subsp. bohemica byl založen na čtyřech lokalitách, a to třikrát až čtyřikrát během let 2000 až 2005. K uskutečnění zásahu simulujícího obhospodařování však nedošlo na podzim 2003, kdy v důsledku extrémního sucha byla naprostá většina růžic již zcela suchá. Výsledky jsou tak k dispozici pro každou lokalitu ze dvou (Chvalšiny) až tří (ostatní tři lokality) experimentálních zásahů (viz tab. 4).
Základem experimentálního designu na každé lokalitě je šest ploch o velikosti 60 cm × 40 cm. Tyto plochy byly před započetím experimentu v říjnu posečeny a disturbovány železnými hráběmi. Následně bylo na každou plochu vyseto větší množství semen (v rozmezí ca jeden až tři tisíce) odebraných tentýž den na lokalitě. Následující rok od výsevu byla v říjnu každá plocha rozdělena na tři podplochy o rozměrech 20 cm × 40 cm. V rámci každé podplochy byl vyčleněn středový pás o šíři 15 cm a délce 35 cm a okrajová zóna 2,5 cm. V každém pásu byl zaznamenán počet semenáčků. V každé ploše pak byla jedna podplocha ponechána bez obhospodařování, druhá posečena srpem, třetí posečena srpem a vyhrabána železnými hrabičkami. Simulaci obhospodařování prováděla jiná osoba než sčitatel růžic a bylo provedeno bez ohledu na růžice hořečků. Počet přežívajících
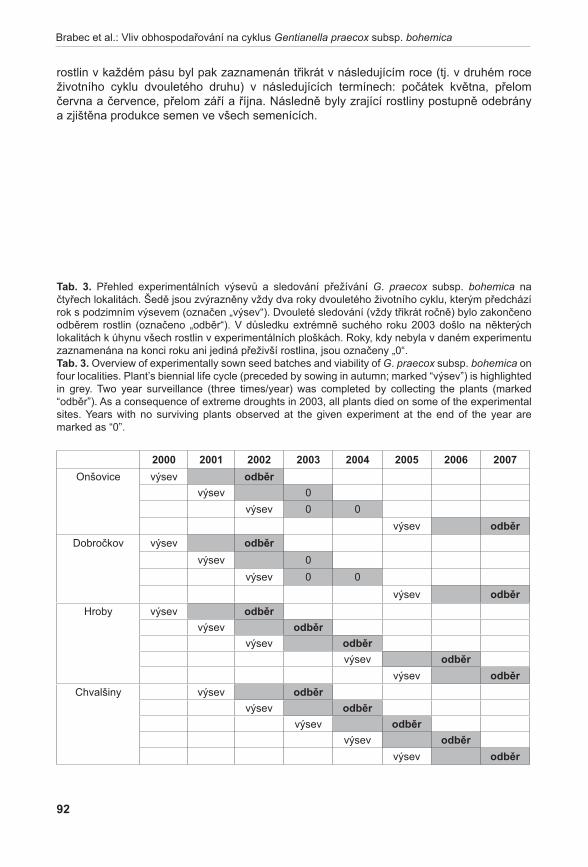
92
Brabec et al.: Vliv obhospodařování na cyklus Gentianella praecox subsp. bohemica
rostlin v každém pásu byl pak zaznamenán třikrát v následujícím roce (tj. v druhém roce životního cyklu dvouletého druhu) v následujících termínech: počátek května, přelom června a července, přelom září a října. Následně byly zrající rostliny postupně odebrány a zjištěna produkce semen ve všech semenících.
tab. 3. Přehled experimentálních výsevů a sledování přežívání G. praecox subsp. bohemica na čtyřech lokalitách. Šedě jsou zvýrazněny vždy dva roky dvouletého životního cyklu, kterým předchází rok s podzimním výsevem (označen „výsev“). Dvouleté sledování (vždy třikrát ročně) bylo zakončeno odběrem rostlin (označeno „odběr“). V důsledku extrémně suchého roku 2003 došlo na některých lokalitách k úhynu všech rostlin v experimentálních ploškách. Roky, kdy nebyla v daném experimentu zaznamenána na konci roku ani jediná přeživší rostlina, jsou označeny „0“.tab. 3. Overview of experimentally sown seed batches and viability of G. praecox subsp. bohemica on four localities. Plant’s biennial life cycle (preceded by sowing in autumn; marked “výsev”) is highlighted in grey. Two year surveillance (three times/year) was completed by collecting the plants (marked “odběr”). As a consequence of extreme droughts in 2003, all plants died on some of the experimental sites. Years with no surviving plants observed at the given experiment at the end of the year are marked as “0”.
2000 2001 2002 2003 2004 2005 2006 2007Onšovice výsev odběr
výsev 0 výsev 0 0 výsev odběr
Dobročkov výsev odběr výsev 0 výsev 0 0 výsev odběr
Hroby výsev odběr výsev odběr výsev odběr výsev odběr výsev odběr
Chvalšiny výsev odběr výsev odběr výsev odběr výsev odběr výsev odběr
93
Příroda, Praha, 31: 85–109, 2011
tab. 4. Přehled experimentálních výsevů a sledování přežívání G. praecox subsp. bohemica po podzimním managementu na čtyřech lokalitách. Šedě zvýrazněny jsou vždy dva roky dvouletého životního cyklu, kterým předchází rok s podzimním výsevem (označen „výsev“). Management byl proveden v říjnu prvního roku (označeno „zásah“). Následné sledování (třikrát ročně) bylo zakončeno odběrem rostlin (označeno „odběr“). V důsledku extrémně suchého roku 2003 došlo na některých lokalitách k úhynu všech rostlin v experimentálních ploškách. Roky, kdy nebyla v daném experimentu zaznamenána na konci roku ani jediná přeživší rostlina, jsou označeny „0“.tab. 4. Overview of experimentally sown seed batches and viability of G. praecox subsp. bohemica on four localities with management measures in autumn. Plant’s biennial life cycle (preceded by sowing in autumn; marked “výsev”) is highlighted in grey. Management measures were undertaken in the first October after sowing (marked “zásah”). Subsequent surveillance (three times/year) was completed by collection of the plants (marked “odběr”). As a consequence of extreme droughts in 2003 all plants died at some of the experimental sites. Years without any surviving plants observed at the given experiment at the end of the year marked as “0”.
2000 2001 2002 2003 2004 2005 2006 2007Onšovice výsev zásah odběr
výsev zásah 0 výsev zásah odběr
Dobročkov výsev zásah odběr výsev zásah 0 výsev zásah odběr
Hroby výsev zásah odběr výsev zásah odběr výsev zásah odběr
Chvalšiny výsev zásah odběr výsev zásah odběr
Semenná banka – pohřbívací experiment (experiment 3)
Odhad velikosti semenné banky byl stanoven metodou pohřbených semen uložených do sáčků vyrobených z monofilu (umělého textilního vlákna). Na každé ze čtyř experimentálních lokalit bylo na podzim 2000 nebo 2001 zakopáno 18 monofilových sáčků o velikosti ca 3 × 3 cm do hloubky jeden až pět cm. Každý sáček obsahoval 300 semen. Po dobu semi až osmi následujících let byly vždy na podzim vyndány dva (výjimečně jeden) sáčky. V sáčcích byl spočten počet nezničených (neshnilých, nerozpadlých) semen, ta byla označena jako „zachovalá“. Následně byl veškerý obsah sáčků vyset do středové plošky o rozměrech 15 × 15 cm v předem připravené ploše o velikosti 20 × 20 cm, která byla před výsevem ošetřena sečí s disturbancí (ve smyslu takto nazývaného typu obhospodařování v experimentu 1, tj. seč s vyhrabáním stařiny, mechorostů a obnažením volné půdy). Počet vyklíčených rostlin byl následně odečítán vždy na jaře, v létě a na podzim po dobu tří let.
Semenná banka – experiment s dokličováním (experiment 4)
Počítání dalších vzešlých semenáčků na ploškách z „experimentu ke zjištění vlivu obhospodařování na životní cyklus (experiment 1)“ bylo prováděno v rozmezí dvou až čtyř let od výsevu, tj. rok II, rok III a rok IV. Procento vyklíčených semen ze semenné banky

94
Brabec et al.: Vliv obhospodařování na cyklus Gentianella praecox subsp. bohemica
v daném roce od výsevu bylo stanoveno jako podíl počtu nalezených semenáčků k počtu semen zbylých v dané plošce (z vysetých 600 semen po odečtu vyklíčených v předchozím roce resp. v předchozích letech).
statistické vyhodnocení
Veškeré statistické vyhodnocení bylo provedeno v programu Statistica 8.0 (statsoft, inc. 1984–2007).
Přežívání rostlin v různých typech obhospodařování (experiment 1)
Ke zjištění vlivu obhospodařování na životní cyklus Gentianella praecox subsp. bohemica (tj. klíčení a přežívání rostlin v průběhu dvou sezón) byla použita analýza variance s opakovaným měřením: nezávislá proměnná (počty přežívajících exemplářů v jednotlivých časech měření, tj. jaro I, léto I, podzim I, jaro II, léto II a podzim II) byla zadána jako opakované měření (faktor ČAS) se šesti opakováními, vysvětlující kategoriální proměnné byly typ obhospodařování (MANAGEMENT) a lokalita (LOKALITA).
Ke zjištění vlivu obhospodařování (MANAGEMENT), lokality (LOKALITA) a sezóny (ROK) na klíčení, přežívání a produkci semen Gentianella praecox subsp. bohemica (viz jednotlivé přechody „a“ až „g“ na obr. 2) byla použita mnohorozměrná analýza varia-nce (bez interakcí). Stanovení vzájemných rozdílů v rámci jednotlivých vysvětlujících kategoriálních proměnných (MANAGEMENT, LOKALITA a ROK) bylo provedeno ná-sled ným mnohonásobným porovnáním Tukey testem. Jako nezávislá proměnná byly testovány: počty vzešlých semen (pro přechod „a“ = klíčení), procento přežívajících rostlin mezi jednotlivými po sobě jdoucími životními fázemi (pro přechody „b“ až „f“) a produkce semen na jednu rostlinu (pro přechod „g“ = produkce semen).
Vliv podzimního obhospodařování na rostliny Gentianella praecox subsp. bohemica (experiment 2)
Přežívání a produkce semen rostlin Gentianella praecox subsp. bohemica v závislosti na podzimním managementu bylo vyhodnoceno mnohorozměrnou analýzou variance (bez interakcí). Jako vysvětlující proměnné byly vždy použity: způsob ošetření (kontrola, seč, seč s disturbancí), lokalita (čtyři lokality) a rok zásahu (2001, 2002 a 2006). Jako nezávislá proměnná byl testován podíl přežívajících rostlin mezi jednotlivými po sobě jdoucími životními fázemi (podzim I až jaro II, jaro II až léto II, léto II až podzim II), podíl rostlin, které došly až do stádia kvetení z počtu rostlin před ošetřením (podzim I až podzim II) a produkce semen na jednu rostlinu. Stanovení vzájemných rozdílů v rámci jednotlivých typů managementu pro přechod podzim I až podzim II bylo provedeno následným mnohonásobným porovnáním Tukey testem.
Semenná banka – experiment s dokličováním (experiment 4)
Klíčení semen rostlin Gentianella praecox subsp. bohemica ze semenné banky v závislosti na managementu výsevových ploch bylo vyhodnoceno mnohorozměrnou analýzou va-ria nce. Jako vysvětlující proměnné byly vždy použity: způsob ošetření (kontrola, seč, seč s disturbancí) a lokalita (čtyři lokality). Jako nezávislá proměnná byl testován podíl vyklíčených semen ze semenné banky v daném roce od výsevu. Stanovení vzájemných rozdílů v rámci jednotlivých typů managementu bylo provedeno následným mnohonásobným porovnáním Tukey testem.

95
Příroda, Praha, 31: 85–109, 2011
výsledky
Životní cyklus a fenologie
Gentianella praecox subsp. bohemica patří mezi dvouleté taxony rodu Gentianella (viz schema životního cyklu na obr. 1). Experimentálně jsme doložili, že rostliny G. praecox subsp. bohemica jsou výlučně dvouleté. Některými botaniky uváděná příležitostná jednoletost nebyla jakkoli potvrzena. Z veškerých provedených experimentů je však zřejmé, že často nalézané malé (jedno- až málokvěté) rostliny (označované některými botaniky za jednoleté) mohou vyrůst z růžic sledovaných po dva roky.
obr. 1. Schéma životního cyklu dvouletého hořečku mnohotvarého českého (Gentianella praecox subsp. bohemica). Podle brabec (2010d).Fig. 1. Life cycle of biennial Gentianella praecox subsp. bohemica (after brabec 2010d).
Semena zpravidla klíčí na jaře, nejčastěji během dubna, přičemž ojediněle lze zastihnout klíční rostlinky i později až do července. Přibližně od počátku května do července tvoří klíční rostlinky první listy a děložní lístky postupně zanikají (tento proces je velmi různý i v rámci jedné lokality). Koncem první vegetační sezóny se rostliny nacházejí ve formě růžice přízemních listů (2–12 listů, nejčastěji 4–8) o průměru od několika milimetrů po ca čtyři cm (maximální průměr dosud nalezené růžice byl osm cm). Koncem podzimu růžice zatahují a zimu přežívají rostliny v přízemních pupenech. Nové listy se objevují v dalším roce již od března. Zpočátku se tvoří víceméně přízemní růžice, často však již s patrnými internodii. Internodia se postupně prodlužují a rostliny se větví. Rychlost a délka prodlužování a celková velikost rostlin je silně závislá na lokalitě a průběhu počasí.

96
Brabec et al.: Vliv obhospodařování na cyklus Gentianella praecox subsp. bohemica
Rostliny rozkvétají od druhé poloviny srpna, výjimečně již dříve, optimum kvetení však bývá od poloviny září do začátku října (na některých výše položených lokalitách ca o dva až tři týdny dříve). Jedna rostlina může mít 1–360 květů, přičemž průměrný počet květů na rostlinu se výrazně liší jak mezi jednotlivými lokalitami, tak mezi jednotlivými roky na téže lokalitě. Nejčastěji je však průměrný počet květů na rostlině mezi 5–30 květy. Hořečky jsou opylovány různým druhem hmyzu (blanokřídlí, dvoukřídlí). Hořeček mnohotvarý český je schopen autogamie, v autogamicky oplodněných semenících se však vyvíjí menší procento semen než při umělé či spontánní allogamii (klauDisová 2003, reitschläGer 2000).
Semena dozrávají koncem léta a na podzim (od konce srpna do poloviny listopadu podle průběhu počasí a polohy lokality). Jeden semeník obsahuje podle polohy na rostlině v průměru přibližně 40–60 (max. 130) zralých semen. Semena z velké části vypadávají z tobolek již během podzimu, zčásti pak během zimy a jara, kdy se postupně rozpadají i celé tobolky. Ojediněle je možné nalézt zbytky semen v tobolkách i během další vegetační sezóny. Semena přes zimu podléhají dormanci (přeléhají). Z části živých semen se vytváří přetrvávající semenná banka a část na podzim dozrátých semen klíčí počátkem následující vegetační sezóny.
Přežívání rostlin v různých typech obhospodařování (experiment 1)
Jak dokládá obr. 2, nejlépe prosperují rostliny na plochách upravených před vysetím se čí a narušením drnu. Analýza variance s opakovaným měřením počtu přežívajících e xem plářů v jednotlivých ploškách jednoznačně ukazuje rozdíly mezi jednotlivými typy ob hospodařování (viz tab. 5). Obr. 2 ukazuje, že rozdíly v přežívání rostlin mezi jednotlivými ty py obhospodařování jsou patrné pouze ve třech ze sedmi sledovaných přechodů, a to při vzchá zení (přechod „a“), přežívání letního období (přechod „c“) a produkci semen (přechod „g“).
Hořečky vzcházejí nejméně v neobhospodařovaných plochách, přibližně třikrát více v plochách sečených a ca desetinásobně v plochách obhospodařovaných sečí s disturbancí. Tento trend se v jednotlivých letech nemění. Analýza variance vzcházení (přechod „a“) vykazuje pro efekt obhospodařování průkazné rozdíly [F(2, 304) = 173,7, p < 0,01], přičemž tyto rozdíly jsou průkazné mezi všemi typy obhospodařování (Tukey test: MS = 1235,4, df = 304, vždy p < 0,01).
Jednoleté rostliny přežívají v průměru letní období (viz obr. 2, přechod „c“) nejlépe v sečených plochách (50,1 % ± 32,5 %), o něco hůře v neobhospodařovaných (45,9 % ± 38,6 %) a výrazně nejhůře v sečených s disturbancí (36,1 % ± 24,9 %). Efekt obhospodařování je při přežívání letního období (přechod „c“) průkazný [analýza variance: F (2, 230) = 5,4, p = 0,005]; průkazně se však od ostatních typů odlišuje pouze obhospodařování sečí s disturbancí: MS = 0,07, df = 230, p < 0,05 pro kontrolu a p < 0,01 pro seč). Přežívání letního období v různých typech obhospodařování (přechod „c“) však nemá v jednotlivých letech shodný trend (viz obr. 3). Efekt roku je pro přežívání letního velmi průkazný: mnohocestná analýza variance bez interakcí F (5, 230) = 15,5, p < 0,001; mnohonásobná porovnávání Tukey testem (MS = 0,07, df = 223) ukazují na hladině průkaznosti 0,001 výraznou odlišnost sezóny 2003 od všech ostatních sezón (žádné další statisticky výrazné rozdíly v rámci efektu roku nebyly zaznamenány).
Obr. 2 ukazuje kladný vliv obhospodařování na množství vyprodukovaných semen na rostlinu (přechod „g“). Tento efekt však není na jednoprocentní hladině významnosti průkazný [analýza variance: F (2, 437) = 4,1, p = 0,02]. V ostatních fázích životního cyklu se schopnost přežívání mezi jednotlivými typy obhospodařování víceméně neliší (viz obr. 2).
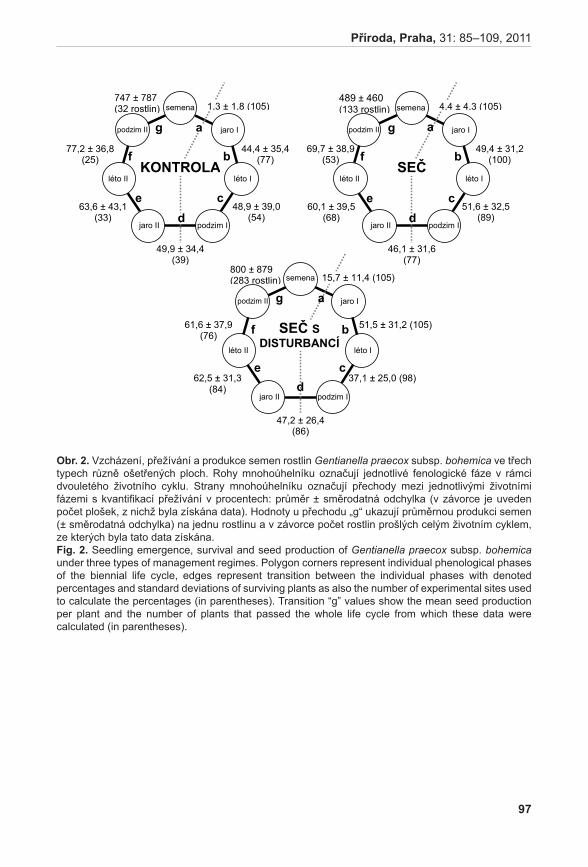
97
Příroda, Praha, 31: 85–109, 2011
jaro I
léto I léto II
podzim II
jaro II podzim I
1,3 ± 1,8 (105)
44,4 ± 35,4 (77)
48,9 ± 39,0 (54)
49,9 ± 34,4 (39)
63,6 ± 43,1 (33)
77,2 ± 36,8 (25)
747 ± 787 (32 rostlin)
KONTROLA
semena
jaro I
léto I léto II
podzim II
jaro II podzim I
4,4 ± 4,3 (105)
49,4 ± 31,2 (100)
51,6 ± 32,5 (89)
46,1 ± 31,6 (77)
60,1 ± 39,5 (68)
69,7 ± 38,9 (53)
489 ± 460 (133 rostlin)
SEČ
jaro I
léto I léto II
podzim II
jaro II podzim I
15,7 ± 11,4 (105)
51,5 ± 31,2 (105)
37,1 ± 25,0 (98)
47,2 ± 26,4 (86)
62,5 ± 31,3 (84)
61,6 ± 37,9 (76)
800 ± 879 (283 rostlin)
SEČ S DISTURBANCÍ
a
b
c d
e
f
g
semena
semena
a
b
c d
e
f
f
g
g
a
b
e c d
obr. 2. Vzcházení, přežívání a produkce semen rostlin Gentianella praecox subsp. bohemica ve třech typech různě ošetřených ploch. Rohy mnohoúhelníku označují jednotlivé fenologické fáze v rámci dvouletého životního cyklu. Strany mnohoúhelníku označují přechody mezi jednotlivými životními fázemi s kvantifikací přežívání v procentech: průměr ± směrodatná odchylka (v závorce je uveden počet plošek, z nichž byla získána data). Hodnoty u přechodu „g“ ukazují průměrnou produkci semen (± směrodatná odchylka) na jednu rostlinu a v závorce počet rostlin prošlých celým životním cyklem, ze kterých byla tato data získána.Fig. 2. Seedling emergence, survival and seed production of Gentianella praecox subsp. bohemica under three types of management regimes. Polygon corners represent individual phenological phases of the biennial life cycle, edges represent transition between the individual phases with denoted percentages and standard deviations of surviving plants as also the number of experimental sites used to calculate the percentages (in parentheses). Transition “g” values show the mean seed production per plant and the number of plants that passed the whole life cycle from which these data were calculated (in parentheses).
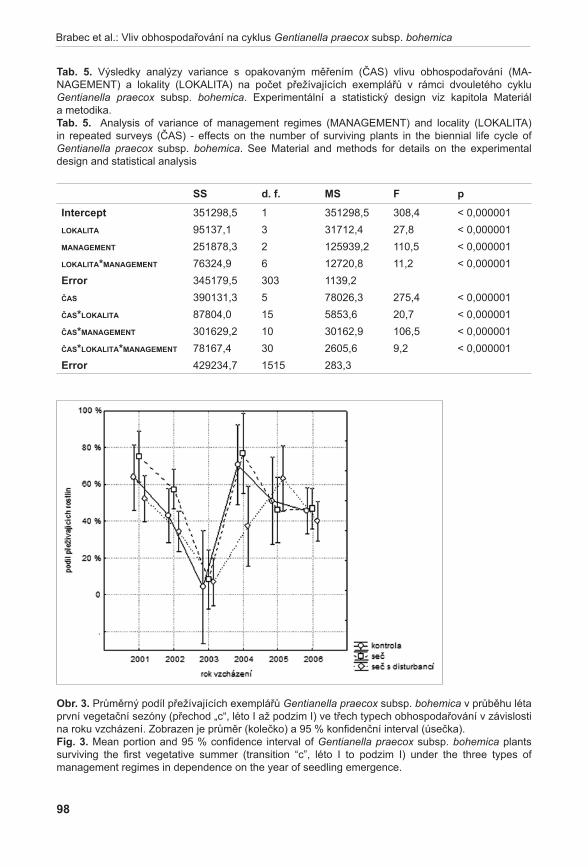
98
Brabec et al.: Vliv obhospodařování na cyklus Gentianella praecox subsp. bohemica
tab. 5. Výsledky analýzy variance s opakovaným měřením (ČAS) vlivu obhospodařování (MA-NAGEMENT) a lokality (LOKALITA) na počet přežívajících exemplářů v rámci dvouletého cyklu Gentianella praecox subsp. bohemica. Experimentální a statistický design viz kapitola Materiál a metodika.tab. 5. Analysis of variance of management regimes (MANAGEMENT) and locality (LOKALITA) in re peated surveys (ČAS) - effects on the number of surviving plants in the biennial life cycle of Gentianella praecox subsp. bohemica. See Material and methods for details on the experimental design and statistical analysis
ss d. f. Ms F p
intercept 351298,5 1 351298,5 308,4 < 0,000001LokaLita 95137,1 3 31712,4 27,8 < 0,000001ManageMent 251878,3 2 125939,2 110,5 < 0,000001LokaLita*ManageMent 76324,9 6 12720,8 11,2 < 0,000001Error 345179,5 303 1139,2Čas 390131,3 5 78026,3 275,4 < 0,000001Čas*LokaLita 87804,0 15 5853,6 20,7 < 0,000001Čas*ManageMent 301629,2 10 30162,9 106,5 < 0,000001Čas*LokaLita*ManageMent 78167,4 30 2605,6 9,2 < 0,000001Error 429234,7 1515 283,3
obr. 3. Průměrný podíl přežívajících exemplářů Gentianella praecox subsp. bohemica v průběhu léta první vegetační sezóny (přechod „c“, léto I až podzim I) ve třech typech obhospodařování v závislosti na roku vzcházení. Zobrazen je průměr (kolečko) a 95 % konfidenční interval (úsečka). Fig. 3. Mean portion and 95 % confidence interval of Gentianella praecox subsp. bohemica plants surviving the first vegetative summer (transition “c”, léto I to podzim I) under the three types of management regimes in dependence on the year of seedling emergence.

99
Příroda, Praha, 31: 85–109, 2011
Jak ukazuje tab. 5, existují významné rozdíly v přežívání některých životních fází ro-stlin G. praecox subsp. bohemica nejen mezi jednotlivými typy obhospodařování, ale i me zi jednotlivými roky a lokalitami. Z postupných analýz přechodů mezi jednotlivými sledovanými životními fázemi („a“ až „g“) vyplývá největší vliv faktorů ROK a LOKALITA na přežívání jednoletých semenáčů během léta (tj. přechod „c“ z léto I na podzim I). Např. extrémně suchá a teplá vegetační sezóna 2003 se výrazně odlišuje (mnohonásobná porovnávání Tukeyho testem na hladině významnosti 0,01) od ostatních sledovaných let v procentech přežívajících exemplářů během celé první vegetační sezóny (přechody „b“ a „c“, tj. jaro I – léto I – podzim I) a letního období druhé vegetační sezóny (přechod „f“, tj. léto II – podzim II).
vliv podzimního obhospodařování na rostliny Gentianella praecox subsp. bohemica (experiment 2)
V dalším experimentu jsme testovali vliv podzimního obhospodařování (shodně nača so-vaného i provedeného jako v experimentu 1) na přežívání jednoletých růžic G. praecox subsp. bohemica. Jak ukazuje tab. 6 na jaře po zásahu (jaro II v rámci životního cyklu) nejlépe přežívají růžice v ploškách ošetřených pouze sečí, hůře jak v plochách kontrolních, tak v plochách ošetřených sečí s disturbancí. Rozdíly však nejsou průkazné [F (2, 178) = 2,28, p = 0,11]. Podzimní obhospodařování má naopak kladný vliv na následné přežívání rostlin z jara II do léta II [viz tab. 6, rozdíly jsou průkazné F (2, 165) = 4,95, p = 0,008] a z léta II do podzimu II [viz tab. 6, rozdíly nejsou průkazné F (2, 110) = 1,78, p = 0,17]. Celkově je vliv podzimního managementu na přežívání do stádia kvetoucích rostlin (přechod podzim I až podzim II) příznivý [viz tab. 6, rozdíly jsou průkazné F (2, 178) = 5,76, p = 0,004], na hladině průkaznosti 0,01 se však odlišují pouze plochy sečené s disturbancí od ploch kontrolních. V průměru bylo nejvíce vyvinutých semen z jedné rostliny v plochách sečených s disturbancí, nejméně pak v kontrolních, tyto rozdíly však byly průkazné pouze na pětiprocentní hladině významnosti [viz tab. 6, F (2, 276) = 3,09, p = 0,047].
tab. 6. Přežívání jeden rok starých rostlin Gentianella praecox subsp. bohemica v závislosti na podzimním managementu (podzim I) a jejich produkce semen. Přežívání je uvedeno v procentech: průměr ± směrodatná odchylka (v závorce je uveden počet plošek, z nichž byla získána data). Hodnoty v posledním sloupci (počet semen na rostlinu) ukazují průměrnou produkci semen (± směrodatná odchylka) na jednu rostlinu a v závorce počet rostlin prošlých celým životním cyklem, ze kterých byla tato data získána.tab. 6. Surviving and seed production of one year old Gentianella praecox subsp. bohemica plants in dependence on the type of the management measures in autumn (podzim I). Surviving is denoted in percentages, with standard deviations and the number of experimental sites used to calculate the percentages in parentheses. The values in the last column show the mean number of seeds produced per plant as also the number of plants that passed the whole life cycle from which these data were calculated (in parentheses).
přežívání rostlin v experimentálních ploškách počet semen na rostlinupodzim I až
jaro IIjaro II až
léto IIléto II až podzim II
podzim I až podzim II podzim II
kontrola 58 ± 33 (61) 38 ± 42 (52) 47 ± 52 (29) 12 ± 27 (61) 307 ± 312 (82)
seč 64 ± 26 (62) 45 ± 44 (59) 57 ± 51 (39) 18 ± 29 (62) 466 ± 485 (74)
sečs disturbancí 56 ± 20 (63) 60 ± 38 (62) 67 ± 43 (50) 23 ± 25 (63) 620 ± 552 (128)
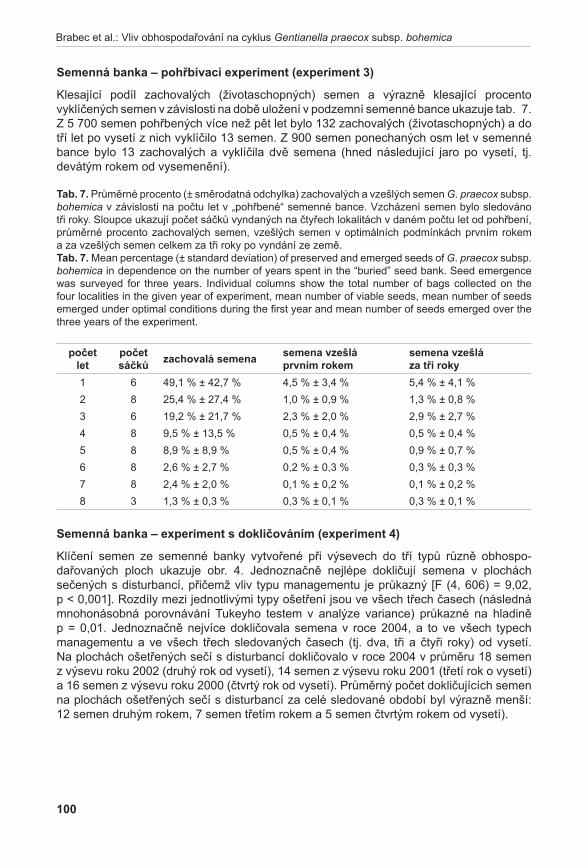
100
Brabec et al.: Vliv obhospodařování na cyklus Gentianella praecox subsp. bohemica
semenná banka – pohřbívací experiment (experiment 3)
Klesající podíl zachovalých (životaschopných) semen a výrazně klesající procento vyklíčených semen v závislosti na době uložení v podzemní semenné bance ukazuje tab. 7. Z 5 700 semen pohřbených více než pět let bylo 132 zachovalých (životaschopných) a do tří let po vysetí z nich vyklíčilo 13 semen. Z 900 semen ponechaných osm let v semenné bance bylo 13 zachovalých a vyklíčila dvě semena (hned následující jaro po vysetí, tj. devátým rokem od vysemenění).
tab. 7. Průměrné procento (± směrodatná odchylka) zachovalých a vzešlých semen G. praecox subsp. bohemica v závislosti na počtu let v „pohřbené“ semenné bance. Vzcházení semen bylo sledováno tři roky. Sloupce ukazují počet sáčků vyndaných na čtyřech lokalitách v daném počtu let od pohřbení, průměrné procento zachovalých semen, vzešlých semen v optimálních podmínkách prvním rokem a za vzešlých semen celkem za tři roky po vyndání ze země.tab. 7. Mean percentage (± standard deviation) of preserved and emerged seeds of G. praecox subsp. bohemica in dependence on the number of years spent in the “buried” seed bank. Seed emergence was surveyed for three years. Individual columns show the total number of bags collected on the four localities in the given year of experiment, mean number of viable seeds, mean number of seeds emerged under optimal conditions during the first year and mean number of seeds emerged over the three years of the experiment.
počet let
počet sáčků zachovalá semena semena vzešlá
prvním rokemsemena vzešlá za tři roky
1 6 49,1 % ± 42,7 % 4,5 % ± 3,4 % 5,4 % ± 4,1 %2 8 25,4 % ± 27,4 % 1,0 % ± 0,9 % 1,3 % ± 0,8 %3 6 19,2 % ± 21,7 % 2,3 % ± 2,0 % 2,9 % ± 2,7 %4 8 9,5 % ± 13,5 % 0,5 % ± 0,4 % 0,5 % ± 0,4 %5 8 8,9 % ± 8,9 % 0,5 % ± 0,4 % 0,9 % ± 0,7 %6 8 2,6 % ± 2,7 % 0,2 % ± 0,3 % 0,3 % ± 0,3 %7 8 2,4 % ± 2,0 % 0,1 % ± 0,2 % 0,1 % ± 0,2 %8 3 1,3 % ± 0,3 % 0,3 % ± 0,1 % 0,3 % ± 0,1 %
semenná banka – experiment s dokličováním (experiment 4)
Klíčení semen ze semenné banky vytvořené při výsevech do tří typů různě obhospo-dařovaných ploch ukazuje obr. 4. Jednoznačně nejlépe dokličují semena v plochách sečených s disturbancí, přičemž vliv typu managementu je průkazný [F (4, 606) = 9,02, p < 0,001]. Rozdíly mezi jednotlivými typy ošetření jsou ve všech třech časech (následná mnohonásobná porovnávání Tukeyho testem v analýze variance) průkazné na hladině p = 0,01. Jednoznačně nejvíce dokličovala semena v roce 2004, a to ve všech typech managementu a ve všech třech sledovaných časech (tj. dva, tři a čtyři roky) od vysetí. Na plochách ošetřených sečí s disturbancí dokličovalo v roce 2004 v průměru 18 semen z výsevu roku 2002 (druhý rok od vysetí), 14 semen z výsevu roku 2001 (třetí rok o vysetí) a 16 semen z výsevu roku 2000 (čtvrtý rok od vysetí). Průměrný počet dokličujících semen na plochách ošetřených sečí s disturbancí za celé sledované období byl výrazně menší: 12 semen druhým rokem, 7 semen třetím rokem a 5 semen čtvrtým rokem od vysetí).
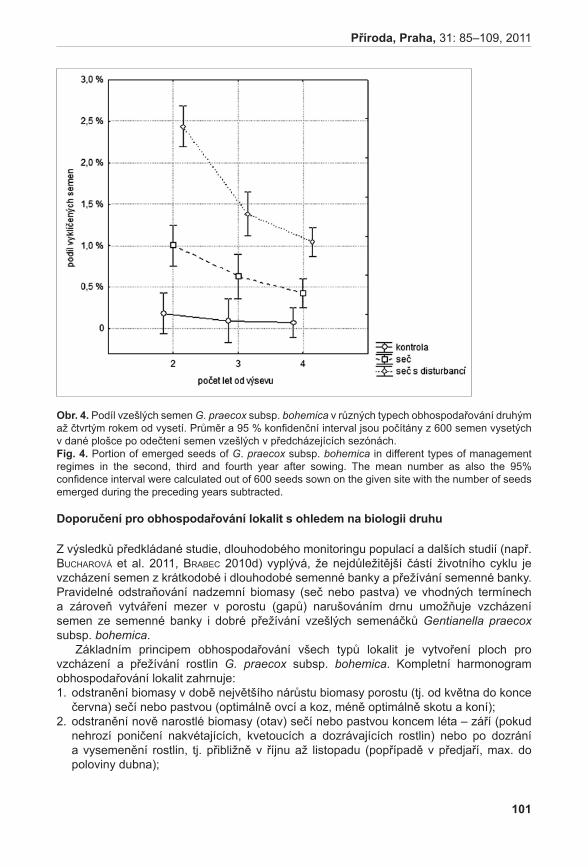
101
Příroda, Praha, 31: 85–109, 2011
obr. 4. Podíl vzešlých semen G. praecox subsp. bohemica v různých typech obhospodařování druhým až čtvrtým rokem od vysetí. Průměr a 95 % konfidenční interval jsou počítány z 600 semen vysetých v dané plošce po odečtení semen vzešlých v předcházejících sezónách.Fig. 4. Portion of emerged seeds of G. praecox subsp. bohemica in different types of management regimes in the second, third and fourth year after sowing. The mean number as also the 95% confidence interval were calculated out of 600 seeds sown on the given site with the number of seeds emerged during the preceding years subtracted.
doporučení pro obhospodařování lokalit s ohledem na biologii druhu
Z výsledků předkládané studie, dlouhodobého monitoringu populací a dalších studií (např. bucharová et al. 2011, brabec 2010d) vyplývá, že nejdůležitější částí životního cyklu je vzcházení semen z krátkodobé i dlouhodobé semenné banky a přežívání semenné banky. Pravidelné odstraňování nadzemní biomasy (seč nebo pastva) ve vhodných termínech a zároveň vytváření mezer v porostu (gapů) narušováním drnu umožňuje vzcházení semen ze semenné banky i dobré přežívání vzešlých semenáčků Gentianella praecox subsp. bohemica.
Základním principem obhospodařování všech typů lokalit je vytvoření ploch pro vzcházení a přežívání rostlin G. praecox subsp. bohemica. Kompletní harmonogram obhospodařování lokalit zahrnuje:1. odstranění biomasy v době největšího nárůstu biomasy porostu (tj. od května do konce
června) sečí nebo pastvou (optimálně ovcí a koz, méně optimálně skotu a koní);2. odstranění nově narostlé biomasy (otav) sečí nebo pastvou koncem léta – září (pokud
nehrozí poničení nakvétajících, kvetoucích a dozrávajících rostlin) nebo po dozrání a vysemenění rostlin, tj. přibližně v říjnu až listopadu (popřípadě v předjaří, max. do poloviny dubna);

102
Brabec et al.: Vliv obhospodařování na cyklus Gentianella praecox subsp. bohemica
3. vytvoření mezer pro klíčení semen v porostu po dozrání a vysemenění rostlin, tj. přibližně v říjnu až listopadu nebo v předjaří (max. do poloviny dubna, lépe dříve) narušením drnu (vláčení, vertikutace, výhrab) nebo pastvou. Narušení drnu lze teoreticky provést kdykoli během zimního období.Obr. 5 shrnuje životní fáze G. praecox subsp. bohemica a termíny komplexní péče
o lokality. Podle vegetačního typu, stavu lokality, polohy lokality, počtu kvetoucích exemplářů G. praecox subsp. bohemica a aktuálního průběhu počasí v sezóně lze základní postup různě modifikovat. Na sušších lokalitách (nejčastěji vegetace svazů Bromion erecti, Koelerio--Phleion phleoidis a sušší typy Violion caninae) stačí seč nebo pastva jedenkrát ročně. V tomto případě je ideální nepravidelné střídání letního a podzimního obhospodařování. Více mezické a vlhčí lokality (nejčastěji vegetace svazů Arrhenatherion, Molinion a vlhčí typy Violion caninae) vyžadují seč, nebo pastvu dvakrát ročně, tj. od května do konce června a na podzim po dozrání hořečků. Pouhé odstranění nadzemní biomasy (např. sečí) bez disturbance drnu a přítomnosti mezer v porostu s volnou půdou (gapů) v době vzcházení je na většině lokalit zcela nedostatečné. Některé lokality (zejména s vegetací s výrazným zápojem mechového patra nebo hustým přízemním bylinným patrem, což se vyskytuje nejčastěji na kyselejších substrátech) vyžadují disturbanci porostu (např. výhrabem, vláčením či vertikutací) každoročně. Na jiných lokalitách (zejména na sušších místech a na bazičtějších substrátech) postačuje provést disturbanci přibližně jedenkrát za tři roky. Volná místa v porostu (gapy) zde přetrvávají déle a jsou též častěji vytvářeny samovolně (např. suchem). Obecně platí, že radikálnější narušení drnu ve vhodném termínu může populaci G. praecox subsp. bohemica výrazně posílit. Prostorová rozrůzněnost obhospodařování v rámci jedné lokality je možná a vítaná.
Při plánování obhospodařování lokalit je potřeba mít na paměti, že i jednoleté vynechání obhospodařování nebo v dané sezóně nedostatečné vytvoření vhodných míst pro klíčení má výrazný vliv na celou populaci a vede k obrovským ztrátám životaschopných semen v semenné bance. Je potřeba zdůraznit, že mnohdy může být pro zachování populace důležitější odstranění biomasy a kvalitní disturbance drnu než dozrání a vysemenění jednoho či několika málo exemplářů. Je tedy možné provést zásah i v nevhodném termínu, např. v době květu či zrání hořečků. Takovéto rozhodnutí lze udělat na základě znalosti parametrů populace G. praecox subsp. bohemica a vegetace dané lokality. V případě známých lokalit v ČR jsou tyto údaje součástí pravidelného monitoringu populací (např. brabec 2010a).
diskuse
Přežívání rostlin v různých typech obhospodařování (experiment 1)
Jednoznačný kladný vliv obhospodařování (zejména pak disturbance) na vzcházení semenáčků hořečků není překvapivý. U „pastevních druhů“, mezi které hořečky je dno-značně patří, je efekt vlivu vhodného prostoru ke klíčení (tzv. gapu) známý a výrazný (DullinGer et al. 2003, ockinGer et al. 2006).
Další životní fází, která se ukázala jako silně ovlivněná stavem porostu (hospodařením), je přežívání letního období. Jak však ukazuje obr. 3, trend nejlepšího přežívání v pouze sečených porostech a nejhoršího v porostech sečených s disturbancí neplatí obecně. Výrazné odchylky od tohoto trendu byly zaznamenány v letech 2004 a 2005 (tj. z let následujících po extrémním suchu v roce 2003). Je potřeba mít na paměti, že údaje pro tyto roky jsou pouze z mezických lokalit Chvalšiny a Hroby (na ostatních byl naprostý nedostatek čerstvých semen). Na lokalitě Chvalšiny vyklíčilo

103
Příroda, Praha, 31: 85–109, 2011
1 2 3 4 5 6 7 8 9 10 11 121.
rok vzcházení
růst růžice
přízemní pupeny
2. ro
k
přízemní pupeny růst nadzemní části kvetení zrání semen
opat
ření
senoseč seč otav
nebo
pastva nebo pastva
výhrab výhrab asanační opatření
obr. 5. Fenologická tabulka dvouletého životního cyklu G. praecox subsp. bohemica s vyznačením fenofází a termínů pro obhospodařování. Sloupce (1 až 12) zobrazují jednotlivé měsíce v roce, řádky pak první a druhý rok růstu hořečku a opatření. Z obrázku je patrné, v jakých fenofázích jsou na lokalitě jedinci G. praecox subsp. bohemica při realizaci jednotlivých ošetření. Na lokalitách s realizovaným asanačním opatřením se předpokládá minimum rostlin G. praecox subsp. bohemica v jednotlivých zobrazených fenofázích. Rostliny zde však pravděpodobně perzistují v semenné bance. Podle brabec (2010d).Fig. 5. Phenological table of the biennial life cycle of G. praecox subsp. bohemica with individual phenological phases and outline of the management measures. The columns show the months of the year, the rows show the first and the second year of the life cycle and the management measures, respectively. From the figure it is evident which phenological phases are present at the site during each of the management measures. It is assumed that only a minimal number of G. praecox subsp. bohemica plants occur in the depicted phenological phases at the sites where rehabilitative management measures are carried into effect and that the plants rather persist in form of the seed bank at these sites. After brabec (2010d).
v roce 2004 na sečených plochách s disturbancí v průměru (± směrodatná odchylka) 261 ± 56 semenáčků na jednu plošku 15 × 15 cm (v ostatních experimentálních letech byl průměr vyklíčených semenáčků na plošku mezi 57 a 110). Zaznamenaný extrémně snížený podíl přežívání semenáčků v tomto roce v ploškách s disturbancí evidentně ukazuje výrazný samozřeďovací efekt. Ponecháme-li v analýzách data jen z let, kdy máme víceméně kompletní design (tj. klíčení z let 2000, 2001, 2002 a 2005) dostáváme opět jednoduchý trend (mírně nejlépe přežívají rostliny v pouze sečených plochách, hůře v kontrolních a nejhůře v sečených s disturbancí). Efekt obhospodařování však není průkazný [F (2,230) = 1,9, p = 0,15]. Při podrobnějším prozkoumání výsledků zjistíme banální věc, že přežívání semenáčků na mezických lokalitách (vegetace svazu Arrhenatherion – lokality Hroby a Chvalšiny) a na suchých lokalitách (vegetace svazu Bromion erecti – lokality Dobročkov a Onšovice) se výrazně liší v různě srážkově bohatých vegetačních sezónách a to i v závislosti na obhospodařování lokality. V suchých letech přežívají rostliny nejlépe na mezických lo kali tách v porostech obhospodařovaných pouze sečí, popř. sečí s disturbancí. Ve srážkově průměrných a nadprůměrných vegetačních sezónách lépe přežívají rostliny v sečených a disturbovaných poros tech na suchých lokalitách, na mezických lokalitách pak pouze v porostech obhos po dařovaných sečí s disturbancí (porosty pouze sečené se příliš zapojují).

104
Brabec et al.: Vliv obhospodařování na cyklus Gentianella praecox subsp. bohemica
Ovlivnění celkového množství semen na rostlinu obhospodařováním porostu není průkazné. Lze si představit, že rostliny v obhospodařovaných plochách jsou statnější, mají více plodů apod. v důsledku menší konkurence v porostu. Takový efekt však není patrný a celkové počty semen na jedné rostlině vykazují velmi velkou variabilitu v rámci všech typů obhospodařování.
To, že nebyly prokázány odlišnosti v přežívání rostlin v jiných fázích životního cyklu kromě vzcházení (pokud nenastanou extrémní klimatické podmínky), je poměrně pře-kvapivý výsledek. Z autopsie lze však říci, že ke vzejití semenáčku ve všech typech ošetření (včetně kontroly) dochází nejčastěji v místech, kde je na povrchu půdy nějaký volný prostor. Takový prostor („gap“) může vzniknout jak zásahem z venku (obhospodařování, disturbance zvěří apod.), tak přirozeně z dynamiky neobhospodařovaného porostu (např. místo po uhynulém listu z nějaké přízemní růžice apod.). Pokud je takový „gap“ obsazen semenáčkem hořečku, je stejně pravděpodobné, že v něm přežije bez ohledu na původ vzniku gapu.
vliv podzimního obhospodařování na rostliny Gentianella praecox subsp. bohemica (experiment 2)
Je velmi pravděpodobné, a naše údaje to potvrzují (viz experiment 2), že podzimní obhospodařování, zejména seč s disturbancí, zničí část jednoletých růžic hořečků. Jak se však ukazuje, celkový efekt pro populaci je jednoznačně kladný: 1) dojde k vy-tvoření vhodného prostoru pro klíčení hořečků následující rok; 2) celkový podíl rostlin v obhospodařovaných plochách, které dojdou až do stádia rozmnožování, se mezi typy ošetření (seč a seč s disturbancí) téměř neliší. Naopak je výrazně vyšší než v plochách kontrolních, tj. bez obhospodařování. Rostliny v ploškách ošetřených sečí s disturbancí měly nejvyšší počet plodů, což se odrazilo odlišnostmi v počtu vyprodukovaných semen na jednu rostlinu. Tyto rozdíly byly velmi pravděpodobně způsobeny podzimním obhospodařováním, které v plochách s disturbancí výrazně snížilo zápoj vegetace a uvolnilo rostlinám hořečků prostor pro růst.
semenná banka – pohřbívací experiment (experiment 3)
Správná interpretace výzkumů životního cyklu vzácných druhů rostlin s cílem jejich záchrany silně závisí na kvalitě údajů o životaschopnosti semenné banky. U druhů rodu Gentianella jsou udávány (sensu thoMson et al. 1997) tři typy semenné banky: přechodná, krátkodobě přetrvávající a dlouhodobě přetrvávající (Pons 1991, MilberG 1994, fischer & Matthies 1998b, kelly 1989, lennartsson & oosterMeijer 2001). Pokud budeme (stejně jako fischer & Matthies 1998b) považovat za životaschopná všechna zachovalá semena (viz tab. 7), je semenná banka průměrných populací pravděpodobně poměrně velká. Zachovalých krátkodobě pohřbených semen (tj. jeden až pět let) bylo v průměru 20 %, zachovalých dlouhodobě pohřbených semen (tj. šest až osm let) bylo v průměru 2 %. Poměrně překvapivá je velká míra klíčení dlouhodobě pohřbených semen (vyklíčilo 11 % ze zachovalých) ve srovnání s krátkodobě pohřbenými (vyklíčilo 9,5 % ze zachovalých). Všeobecně by se očekával spíše opačný trend (viz fischer & Matthies 1998b).
semenná banka – experiment s dokličováním (experiment 4)
Uměle (experimentálně) pohřbená semena samozřejmě neumožňují věrohodně od-had nout přirozenou dormanci a ztrátu klíčivosti semen. Z dokličování v optimálně ob-hospodařovaných plochách (seč s disturbancí) můžeme (byť velmi hrubě) odhadnout

105
Příroda, Praha, 31: 85–109, 2011
podíl semen klíčících ihned následující rok (přechodná semenná banka) a semen klíčích z krátkodobě přetrvávající semenné banky, v našem případě jeden až tři roky – dle definice (thoMson et al. 1997) jsou semena v krátkodobě přetrvávající semenné bance v půdě jeden až pět let. Ihned následující rok po vysetí vyklíčilo 15,7 % semen (viz obr. 1), následující tři roky pak vyklíčilo 1,6 % semen (z vysetých po odečtení vyklíčených první rok). Velmi hrubě lze odhadovat, že i za optimálních podmínek ke klíčení přibližně nejméně jedna desetina semen zůstává dormatních v tzv. krátkodobé přetrvávající semenné bance.
Nárůst vzcházení semenáčků různých druhů na jaře následujícím po suchém roce díky vytvoření gapů ve vegetaci vhodných pro klíčení je v literatuře dobře popsán (Gross et al. 1998, horvitz et al. 1998, sMith et al. 2005, töranG et al. 2010). Extrémně suchý rok 2003 takto ovlivnil populace Gentianella praecox subsp. bohemica v celém areálu (brabec 2010c). Důsledkem toho byl hořečkový rok 2005, kdy na většině lokalit vykvetlo nadprůměrné množství hořečků (brabec 2010c), které je v mnoha případech maximálním zaznamenaným počtem kvetoucích exemplářů za dobu celého pravidelného monitoringu (brabec 2010d).
doporučení pro obhospodařování lokalit s ohledem na biologii druhu
Pro ochranářskou praxi z výsledků vyplývá, že nejlépe kladně ovlivnitelnou fází životního cyklu je vzcházení. Jednoznačně nejlepším obhospodařováním se jeví seč s disturbancí, která vytvoří v porostu „gapy“ přetrvávající do doby klíčení hořečků (tj. duben až květen). Brát ohled na to, že v některých letech dochází v porostu ošetřeném sečí s disturbancí k výraznějšímu úhynu semenáčků během letního období, nemá příliš opodstatnění: 1) podíl rostlin, které projdou celým životním cyklem, je na plochách s disturbancí vždy výrazně vyšší než na plochách pouze sečených, a to i v sušších sezónách; 2) průběh počasí v sezóně nelze dopředu předpovědět. Extrémně suché roky, kdy dojde na lokalitě k úhynu všech nebo téměř všech vzešlých rostlin (a tím k výraznému zásahu do semenné banky), přicházejí s periodou přibližně jedenkrát za 20 let (bucharová et al. 2011). Tudíž nemá smysl s nimi v plánech obhospodařování kalkulovat.
Vyvláčení, vertikutaci, či razantní výhrab s narušením drunu je potřeba provádět v takové intenzitě, aby se zralá semena dostala do podmínek vhodných ke klíčení, tj. k volné zemi. Disturbance může být tak razantní, že odstraní většinu nadzemní biomasy rostlin. Podzemní části vegetačního krytu (kořenový systém, drn) mají být však pouze narušeny, nikoli plošně odstraněny. Není proto příliš vhodné porost zkopávat nebo rýt, v případě použití rýče, krumpáče, či motyky je lepší drn pouze při zemi „seříznout“, nevykopávat. Vyvláčenou (vyhrabanou apod.) biomasu je potřeba z lokality odstranit, neponechávat ji v ploše s výskytem hořečků. Co nejrazantnější disturbanci lze doporučit ideálně dva roky po sobě na obnovovaných lokalitách (tj. na lokalitách, s nepravidelným nebo nedostatečným obhospodařováním, kde se kvetoucí exempláře v posledních letech objevují jen zřídka a v malém množství, tj. dle rozlohy ca do 15 až 50 ex.). Na lokalitách stabilizovaných nebo s bohatší populací kvetoucích exemplářů v posledních letech lze velmi razantní disturbanci odstraňující velkou část nadzemní biomasy doporučit ca jedenkrát za tři roky (závisí na typu vegetace, nárůstu a zapojení nadzemní biomasy v dané sezóně, apod.). Razantnější disturbanci porostu je nanejvýš vhodné provést v období, kdy na lokalitě odkvetlo a vysemenilo nadprůměrné množství hořečků (např. v tzv. „hořečkovém roce“), a to buď ještě na podzim po dozrání většiny rostlin, tj. obvykle od druhé poloviny října do zámrazu, nebo po roztátí sněhu v předjaří následujícího roku, tj. v období do poloviny dubna.

106
Brabec et al.: Vliv obhospodařování na cyklus Gentianella praecox subsp. bohemica
Populace hořečků na lokalitě je tvořena semeny v semenné bance, jednoletými růžicemi a dvouletými kvetoucími rostlinami, které přináší čerstvá semena. V životaschopné populaci tvoří živá semena absolutně největší část populace taxonu – ve stabilizované populaci s více než 100 kvetoucích exemplářů ročně je to odhadem přes 99 %, v menších populacích je toto procento nižší, často je však odhadem přes 90 %. Jak ukazuje tab. 7 a obr. 4 klíčivost semen postupně klesá. Semena hořečků po dozrání vesměs volně pa-da jí k zemi, semena zachycená na vegetaci a stařině se splachováním srážkami po-stupně dostávají k hlíně, do půdního horizontu se dostávají nejčastěji v důsledku tlení a mineralizace nad nimi ležícího opadu. Čerstvá semena a semena z posledních let se tedy nachází v nejsvrchnějších částech půdního horizontu, semena starší pak mohou být i hlouběji v půdě. Z tohoto předpokladu je vhodné též vycházet při stanovení intenzity disturbance porostu na lokalitě. Chceme-li „aktivovat“ semena stará maximálně tři až pět let bohatě postačí důsledné vyvláčení s rozvolněním či větším odstraněním nadzemní biomasy bez výraznějšího zásahu do půdního horizontu. Naopak při snaze „dostat“ ze semenné banky semena stará pět a více let, je při tvorbě gapů vhodné nejvrchnější půdní horizont výrazněji narušit. Jinými slovy – lokality s nulovým či malým počtem kvetoucích exemplářů v posledních pěti letech vyžadují spíše razantnější zásahy s výrazným narušením drnu a půdního horizontu. U lokalit, kde byl alespoň jedenkrát v posledních ca třech až pěti letech větší počet zralých semenících hořečků, postačuje při tvorbě gapů odstranit mechorosty a stařinu, tj. nadzemní biomasu.
Výsledky ukazují, že dodržování zásad managementu, jeho dostatečný plošný rozsah a udržení jeho kvality po určitou dobu by mělo velmi rychle vést k růstu populace G. praecox subsp. bohemica na dané lokalitě. Maticové modelování životního cyklu druhu vytvořené na shodných datech (bucharová et al. 2011) predikuje při pravidelném obhospodařování sečí s disturbancí velkou pravděpodobnost přežití a rychlou stabilizaci i populacím s méně než deseti kvetoucími exempláři. Podobně velká pravděpodobnost přežívání při dodržovaní kvalitního a dobře načasovaného managementu byla prokázána i pro druh Gentianella campestris (lennartsson 2000). Je však potřeba mít na paměti, že experimenty byly provedeny na čtyřech velkých, víceméně stabilních lokalitách v centru historického areálu druhu. Podmínky v menších populacích a na okrajích areálu mohou být výrazně odlišné. Malé populace snáze podléhají vlivům extrémních událostí (beissinGer & westPhal 1998) a jsou více zranitelné demografickou a environmentální stochasticitou (např. nevhodným zásahem či nepovedeným obhospodařováním, cf. lennartsson 2000). U menších populací jsou také pravděpodobné genetické změny, např. imbreeding (ellstranD & elaM 1993, ouborG & vantreuren 1995) nebo změny způsobené nedostatkem sousedů stejného druhu, tzv. Allee effect (stePhens et al. 1999). fischer & Matthies (1997, 1998a) prokázali u Gentianella germanica v Juře závislost stávající velikosti populace (počítané jako počet kvetoucích exemplářů) a počtu semen na tobolku v jednom roce. Malé populace produkovaly výrazně menší počet semen na tobolku než populace velké. Jak již bylo uvedeno, G. praecox subsp. bohemica je schopna spontánní autogamie, která přináší relativně velké procento vyvinutých semen – v průměru 5 až 55 % oproti přírodním podmínkám (viz reitschläGer 2000, klauDisová 2003). Allogamie, opylování hmyzem, zejména čmeláky, však hraje u hořečků nezastupitelnou roli (reitschläGer 2000, klauDisová 2003, kruPa & jůzová 2007). Nelze tedy vyloučit i možnost snížení životaschopnosti populací v důsledku nedostatku opylovačů. Pro Gentianella praecox subsp. bohemica však dosud nebyli opylovači zkoumáni.

107
Příroda, Praha, 31: 85–109, 2011
summary
Gentianella praecox subsp. bohemica is a strictly biennial taxon inhabiting wide range of grassland communities. A detailed survey of population dynamics at four localities showed that population survival is mainly dependent on site management and size of seed bank. Seedling recruitments is a critical phase of G. praecox subsp. bohemica life cycle and can be positively influenced by hay mowing with subsequent sod disturbance. The resulting gaps in vegetation cover persist till seed germination starting in spring (April – May) after seed dissemination in a previous autumn (September – October). Sod disturbance can be carried out using harrows, verticutter or metal rakes either in autumn after seed ripening or in early spring after snow melting (latest till mid April). Disturbance must be strong enough for ripe seeds to reach conditions suitable for germination, i.e. soil surface.
Poděkování
Autoři jsou velmi zavázáni všem kolegům a přátelům, kteří pomáhali s prací na experimentálních plochách, zejména pak Honzovi Brabcovi. Díky patří všem, kteří přispěli diskusí, radou a nápadem při zpracování dat, psaní článku a konečných úpravách, zejména pak Lukáši Krinkemu, Františku Krahulcovi, Janu Brabcovi a recenzentům, jež si nepřáli býti anonymní, Lídě a Janu Kirschnerovým.
Výběr lokalit a založení experimentů bylo podpořeno projektem Grantové agentury Univerzity Karlovy „Rozšíření a ekologie taxonů rodu Gentianella Moench (hořeček) v České republice“ (268/1999/B-BIO/PřF). Vyhodnocení experimentů bylo součástí projektu MŠMT VaV 2B06178. Interpretace výsledků dílčích studií a jejich převedení do ochranářské praxe bylo usnadněno díky dlouhodobému pravidelnému monitoringu populací Gentianella praecox subsp. bohemica, kterého se v posledních deseti letech zúčastnilo více než 100 lidí a který je realizován za finanční podpory AOPK ČR, a díky podkladům shromážděným pro záchranný program druhu (brabec 2010d), který byl realizován za finanční podpory grantem z Islandu, Lichtenštejnska a Norska v rámci Finančního mechanismu EHP a Norského finančního mechanismu 003/2.
literatura
beissinGer s. r. & westPhal M. i. (1998): On the use of demographic models of population viability in endangered species management. – J. Wildl. Manage. 62: 821–841.
brabec j. (2004): Monitoring hořečku mnohotvarého českého (Gentianella praecox ssp. bohemica) v ČR (sezóna 2004) – Ms. [Depon in: ústřední pracoviště AOPK ČR, Praha.]
brabec j. (2005): Současný stav rozšíření hořečku mnohotvarého českého (Gentianella praecox subsp. bohemica) v ČR. – Zpr. Čes. Bot. Společ. 40: 1–44.
brabec j. (2010a): Monitoring hořečku mnohotvarého českého (Gentianella praecox subsp. bohemica) v ČR (sezóna 2010) – Ms. [Depon in: Agentura ochrany přírody a krajiny ČR, Praha.]
brabec j. (2010b): Hořeček drsný Sturmův – vymírající svědek pastvy vlhkých luk. – In: brabec j. [ed.], Přírodní fenomény a zajímavosti západních Čech, pp. 49–66, Mezi lesy, Prostiboř & Muzeum Cheb, p. o. Karlovarského kraje
brabec j. (2010c): Monitoring populací a stavu lokalit hořečku nahořklého (Gentianella amarella) a hořečku drsného (Gentianella obtusifolia). – Ms. [Depon in: Ústřední výkonná rada Českého svazu ochránců přírody, Praha.]
brabec j. (2010d): Záchranný program hořečku mnohotvarého českého (Gentianella praecox subsp. bohemica) v České republice. – Ms. [Depon in: Agentura ochrany přírody a krajiny ČR, Praha, URL:http://www.zachranneprogramy.cz.]
bucharová a., brabec j. & MünzberGová z. (2011, accepted): Effect of land use and climate change on future fate of populations of an endemic species of central Europe. – Biol. Conserv.
DullinGer s., Dirnbock t., GreiMler j. & Grabherr G. (2003): A resampling approach for evaluating effects of pasture abandonment on subalpine plant species diversity. – J. Veg. Sci. 14: 243–252.

108
Brabec et al.: Vliv obhospodařování na cyklus Gentianella praecox subsp. bohemica
ellstranD n. c. & elaM D. r. (1993): Population genetic consequences of small population size – implications for plant conservation. – Annu. Rev. Ecol. Syst. 24: 217–242.
enGleDer t. (2006): Der Böhmische Kranzenzian / Gentianella bohemica (Gentianaceae) im österreichischen Teil der Böhmischen Masse (Böhmerwald, Mühl- und Waldviertel). – Neilreichia 4: 215–220.
enGleDer t. (2010): Artenhilfsprojekt Böhmischer Enzian, Bericht 2009, Böhmerwald / Mühlviertel / Waldviertel. – Unterstützt von EU, Land OÖ, önj.
eriksson o., cousins s. a. o. & bruun h. h. (2002): Land-use history and fragmentation of traditionally managed grasslands in Scandinavia. – J. Veg. Sci. 13: 743–748.
fischer M. & Matthies D. (1997): Mating structure and inbreeding and outbreeding depression in the rare plant Gentianella germanica (Gentianaceae). – Am. J. Bot. 84: 1685–1692.
fischer M. & Matthies D. (1998a): Effects of population size on performance in the rare plant Gentianella germanica. – J. Ecol. 86: 195–204.
fischer M. & Matthies D. (1998b): Experimental demography of the rare Gentianella germanica: seed bank formation and microsite effects on seedling establishment. – Ecography 21: 269–278.
fischer M. & stöcklin j. (1997): Local extinctions of plants in remnants of extensively used calcareous grasslands 1950-85. – Conserv. Biol. 121–131.
fischer M. a., oswalD k. & aDler w. (2008): Exkursionsflora für Österreich, Liechtenstein und Südtirol. Ed. 3. – Land Oberösterreich, Biologiezentrum der OÖ Landesmuseen, Linz.
foley j. a., De fries r., asner G. P., barforD c., bonan G., carPenter s. r., chaPin f. s., coe M. t., Daily G. c., Gibbs h. k., helkowski j. h., holloway t., howarD e. a., kucharik c. j., MonfreDa c., Patz j. a., Prentice i. c., raMankutty n. & snyDer P. k. (2005): Global consequences of land use. – Science 309: 570–574.
Gerža M. & ČePa l. (2007): Nová lokalita Gentianella praecox subsp. bohemica v polské části Orlických hor. – Zpr. Čes. Bot. Společ. 42: 137–139.
Gołąb z. (2004): Gentianella bohemica Skalický, Goryczuszka czeska. – In: suDnik-wójcikowska b. & werblan-jakubiec h. [eds], Poradnik ochrony siedlisk i gatunków Natura 2000. Tom 9. Gatunki roślin, pp. 132–135, Ministerstwo Środowiska, Warszawa.
Gross k., lockwooD j. r., frost c. c. & Morris M. f. (1998): Modeling controlled burning and trampling reduction for conservation of Hudsonia montana. – Conserv. Biol. 12: 1291–1301.
haeuPler h. & Muer t. (2000): Bildatlas der Farn- und Blütenpflanzen Deutschlands. – Ulmer, Stuttgart.
horvitz c. c., Pascarella j. b., McMann s., freeDMan a. & hofstetter r. h. (1998): Functional roles of invasive non-indigenous plants in hurricane-affected subtropical hardwood forests. – Ecol. Appl. 8: 947–974.
jäGer e. j. & werner k. [eds] (2002): Rothmaler, Exkursionsflora von Deutschland. Gefäßpflanzen: Kritischer Band. – Ulmer, Stuttgart.
kelly D. (1989): Demography of short-lived plants in chalk grassland .1. Life-cycle variation in annuals and strict biennials. – J. Ecol. 77: 747–769.
kirschner j. & kirschnerová l. (2000): Gentianella Moench – hořeček. – In: slavík b. [ed.], Květena České republiky 6, pp. 82–98, Academia, Praha.
kirschner j. & kirschnerová l. (2003): Změny v rozšíření Gentianella lutescens subsp. lutescens a subsp. carpatica v České republice. – Zpr. Čes. Bot. Společ. 38: 205–216.
kirschnerová l. & kirschner j. (1997): Hořeček drsný Sturmův, ještě nevymřelý taxon české květeny. – Zpr. Čes. Bot. Společ. 32: 1–13.
kirschnerová l. (2002): Gentianaceae Juss. – hořcovité. – In: kubát k., hrouDa l., chrtek j. jun., kaPlan z., kirschner j. & ŠtěPánek j. [eds], Klíč ke květeně České republiky, pp. 500–506, Academia, Praha.
klauDisová M. (2003): Studium vybraných fází životního cyklu Gentianella praecox subsp. bohemica a G. ama rella subsp. amarella. – Ms. [Dipl. Práce, depon. in: knihovna Katedry botaniky PřF UK, Praha.]
kolbek j., Plesková e. & soMol v. (1994): Biomonitorování druhu Gentianella baltica na Křivoklátsku. – Příroda, Praha, 1: 197–205.
kruPa M. & jůzová b. (2007): Opylovači hořečků Gentianella lutescens (subsp. lutescens, carpatica) a Gentianella ciliata se zaměřením na čmeláky. – Ms. [Závěr. zpráva, depon in: Správa CHKO Bílé Karpaty.]

109
Příroda, Praha, 31: 85–109, 2011
kubát k., hrouDa l., chrtek j. jun., kaPlan z., kirschner j. & ŠtěPánek j. [eds.] (2002): Klíč ke květeně České republiky. – Academia, Praha.
lennartsson t. & oosterMeijer j. G. b. (2001): Demographic variation and population viability in Gentianella campestris: effects of grassland management and environmental stochasticity. – J. Ecol. 89: 451–463.
lennartsson t. (2000): Management and population viability of the pasture plant Gentianella campestris: the role of interactions between habitat factors. – Ecol. Bull. 48: 111–121.
luoto M., rekolainen s., aakkula j. & Pykala j. (2003): Loss of plant species richness and habitat connectivity in grasslands associated with agricultural change in Finland. – Ambio 32: 447–452.
MilberG P. (1994): Germination ecology of the endangered grassland biennial Gentianella campestris. – Biol. Conserv. 70: 287–290.
Moravec j. [ed.] (1995): Rostlinná společenstva ČR a jejich ohrožení. Ed. 2. – Severočes. Přír., Příl. 1995: 1–206.
ockinGer e., eriksson a. k. & sMith h. G. (2006): Effects of grassland abandonment, restoration and management on butterflies and vascular plants. – Biol. Conserv. 133: 291–300.
ouborG n. j. & vantreuren r. (1995): Variation in fitness-related characters among small and large populations of Salvia pratensis. – J. Ecol. 83: 369–380.
Pons t. l. (1991): Dormancy, germination and mortality of seeds in a chalk-grassland flora. – J. Ecol. 79: 765–780.
PoPelářová M., jonGePierová i. & kruPa M. (2008): Hořeček žlutavý – vymírající druh moravských Karpat. – In: Piro z. & wolfová j. [eds], Zachování biodiverzity karpatských luk, pp. 37–41, FOA, nadační fond pro ekologické zemědělství, Praha.
PoschloD P. & wallis De vries M. f. (2002): The historical and socioeconomic perspective of calcareous grasslands - lessons from the distant and recent past. – Biol. Conserv. 104: 361–376.
Procházka f. [ed.] (2001): Černý a červený seznam cévnatých rostlin České republiky (stav v roce 2000). – Příroda, Praha, 18: 1–166.
reitschläGer j. (2000): Ekologické vazby a opylovací strategie ohroženého druhu Gentianella bohemica (hořeček český). – Ms. [Magist. práce, depon. in: Biologická fakulta JU České Budějovice.]
rösler s. (2005): Böhmischer Enzian 2005. – Ms. [Unveröffentlichtes Gutachten im Auftrag des Bayerischen Landesamtes für Umwelt (LfU), Augsburg.]
skalický v. & sofron j. (1990): Zpráva o exkurzi Západočeské pobočky ČSBS na Sušicko. – Zprav. Západočes. Poboč. ČSBS 1: 6–9.
skalický v. (1969): Die Sammelart Gentianella germanica (Willd.) E. F. Warburg s. l. im Böhmischen Massiv. – Preslia 41: 140–147.
sMith M., caswell h. & Mettler-cherry P. (2005): Stochastic flood and precipitation regimes and the population dynamics of a threatened floodplain plant. – Ecol. Appl. 15: 1036–1052.
soMol v., Plesková e. & brabec j. (1999): Management lokality Gentianella baltica na Křivoklátsku. – Příroda, Praha, 15: 63–64.
statsoft, inc. (1984–2007): STATISTICA (data analysis software system), version 8.0. – URL: http://www.statsoft.com.
stePhens P. a., sutherlanD w. j. & freckleton r. P. (1999): What is the Allee effect? – Oikos 87: 185–190.thoMPson k., bakker j. P. & bekker r. M. (1997): The soil seed banks of North West Europe:
methodology, density and longevity. – Cambridge University Press.töranG P., ehrlén j. & ÅGren j. (2010): Linking environmental and demographic data to predict future
population viability of a perennial herb. – Oecologia 163: 99–109.wisskirchen r. & haeuPler h. (1998): Standardliste der Farn- und Blütenpflanzen Deutschlands. –
Ulmer, Stuttgart.zahraDníková j. (1995): Monitoring rodu hořeček v Krkonoších. – Opera Corcontica 32: 131–136.zając a. & zając M. [eds] (2001): Atlas rozmieszcezenia roślin naczyniowych w Polsce. – Instytut
Botaniki Uniwersytetu Jagiellońskiego, Kraków.ziPP t. (2006) Artenschutzprojekt „Maßnahmen zur Verbesserung der Bestandssituation von
Gentianella bohemica im Bayerischen Wald 2006.“ – Ms. [Unveröffentlichtes Gutachten im Auftrag des Bayerischen Landesamtes für Umwelt (LfU), Neureichenau.].

110
Brabec et al.: Vliv obhospodařování na cyklus Gentianella praecox subsp. bohemica

111
Příroda, Praha, 31: 111–126, 2011
Hrachor hrachovitý (Lathyrus pisiformis) v České a slovenské republiceLathyrus pisiformis in the czech and slovak Republics
MichaL štefánek
Herbářové sbírky a Katedra botaniky Přírodovědecké fakulty Univerzity Karlovy, Benátská 2, CZ-128 01 Praha 2, [email protected]
Abstract: Lathyrus pisiformis belongs to the most endangered plants in the Czech Republic. However, our knowledge of habitat requirements and population biology of this rare species is very insufficient. This work was focused on basic characteristics of two localities of this species in the Czech Republic: soil, phytosociology, shading at localities, and some basic parameters of population biology (number of stems, seeds etc.) between the years 2007 and 2011 were analyzed.Keywords: population biology, phytosociology, shading, soil analysis.
Abstrakt: Hrachor hrachovitý (Lathyrus pisiformis) patří v České republice mezi nejvzácnější rostliny vůbec. Nicméně o jeho nárocích na prostředí, ale také o jeho populační biologii máme v současnosti jenom velmi omezené informace. Analyzovány byly jak půdní nároky, tak i fytocenologické nároky hrachoru a též zastínění jeho lokalit a některé základní populačně biologické charakteristiky druhu.Klíčová slova: populační biologie, fytocenologie, zástin, půdní analýzy.nomenklatura: cévnaté rostliny – kubát et al. 2002, společenstva – Moravec et al. 1995
Úvod
Hrachor hrachovitý je jedním z nejvzácnějších a zároveň nejohroženějších druhů naší květeny. Recentně má v České republice pouhé dvě lokality a celkový počet jedinců se pohybuje v desítkách. Patří mezi druhy vyžadující dnes již víceméně vymizelý typ světlých teplomilných lesů (tzv. střední les), obdobně jako Festuca amethystina, Adenophora liliifolia a další jiné organismy, například některé teplomilné houby či druhy hmyzu (podrobnosti viz konviČka et al. 2004). Svou roli ve „vzácnosti“ tohoto taxonu u nás, ale také v okolních zemích (Slovensko, Polsko či Maďarsko) hraje také to, že ve střední Evropě má hrachor západní hranici svého areálu.
Navzdory výše uvedeným faktům nebyl tento druh u nás podrobněji zkoumán (až na dvě výjimky: Holub in Čeřovský 1999, kubát 2003) a stál na okraji zájmu botaniků věnujících se biologii vzácných druhů. Tento „dluh“ se snažil napravit projekt věnovaný kriticky ohroženým druhům rostlin (VaV) a studie hrachoru hrachovitého, co by jednoho z modelových druhů tohoto projektu.
Vzhledem k naprosté absenci znalostí o biologii druhu byla tedy pozornost věnována studiu základních parametrů biologie druhu (sledování kvetení, produkce semen, počtu jedinců) a prostředí, ve kterém taxon roste (půdní charakteristiky, fytocenologické nároky, sledování zástinu).

112
Štefánek: Lathyrus pisiformis v ČR a SR
Popis druhu
Hrachor hrachovitý (Lathyrus pisiformis L.) patří mezi hrachory sekce Orobus ser. Pisiformes (bässler 1973, kenicer et al. 2005).
Jde o druh vytrvalý se silným kořenem a krátkým, tlustým oddenkem, z něhož vyrůstají jednotlivé lodyhy. Ty jsou popínavé a až jeden metr dlouhé (hranaté, dvoukřídlé, rýhované a lysé). Listy jsou s 3–5 páry lístků přisedlých nebo krátce řapíčkatých, mladé či zakrslé rostliny ale mohou mít jen jeden až dva páry lístků. Vřeteno listu je zakončeno větvenou úponkou. Nápadné jsou, především u vyvinutých rostlin, velké palisty, které mohou být téměř tak velké jako lístky samotné; lístky 32–60 (80) mm dlouhé, 10–32 mm široké; palisty 22–50 mm dlouhé a 10–20 mm široké. Celé rostliny jsou lysé (kromě lusků, které jsou žlaznaté a krátce chlupaté). Květenství nijak nevybočují od řady jiných zástupců rodu Lathyrus (obr. 1), v květenství najdeme obvykle 5–14 květů, koruny jsou 1–1,5 cm dlouhé a špinavě růžové. V případě našich populací bývá jen menší část květů úspěšně opylena a obvykle se vyvíjejí jen maximálně tři lusky z květenství. Lusky bývají okolo pěti centimetrů dlouhé a obvykle obsahují 6 až 13 kulovitých semen (průměrně osm semen), které jsou ca tři mm velké a bývají hladké, tmavě hnědé nebo s tmavou kresbou na světlejším podkladě; Chrtková & Bělohlávková in slavík 1995)
obr. 1. Hrachor hrachovitý (Lathyrus pisiformis), Plešivská planina, červen 2010, foto M. Štefánek.Fig. 1. Lathyrus pisiformis, Plešivská plateau, Slovak Karst, June 2010, photo M. Štefánek.

113
Příroda, Praha, 31: 111–126, 2011
Celková produkce semen je tedy poměrně nízká. Dalším důležitým faktorem je, že při klíčení semen hraje podstatnou roli jejich dormance. Semena hrachoru klíčí velmi neochotně, pouze po mechanickém narušení osemení je situace lepší, pak klíčí poměrně spolehlivě (podrobnosti viz kubát 2003). Jak již bylo zmíněno, hrachor je rostlina vytrvalá, podle pozorování označených rostlin v terénu žijí konkrétní rostliny minimálně pět let, ale dá se předpokládat, že je to mnohem více.
Rozšíření druhu
Celkový areál druhu sahá od Dálného Východu přes Sibiř a okrajové části Mongolska a Číny do Pobaltských zemí a střední Evropy. České lokality tvoří (s výjimkou izolovaných lokalit ve Španělsku) západní okraj areálu druhu. Ve většině evropských zemí, kde se vyskytuje, patří hrachor hrachovitý mezi druhy velmi ohrožené, zařazené do červených seznamů jednotlivých regionů a nezřídka také do červených knih jednotlivých republik (například Čeřovský et al. 1999, Herbich in kaźMierczakowa & zarzycki 2001, baleviČius et al. 1992). Výjimkou je Rusko, zejména v centru areálu jde o druh roztroušený až hojný a potenciálně zajímavý z pícninářského hlediska.
Na Slovensku byl Lathyrus pisiformis poprvé nalezen ve Slovenském krasu v třicátých letech 20. století (Suza 1935 PR, PRC; krist 1936). Lokality nebyly dlouhou dobu potvrzeny (možná ani ověřovány, výjimkou je nález Klášterského z roku 1958), až R. Šuvada nalezl v letech 2004–2006 tři lokality (dvě na Plešivské planině a jednu na Dolném vrchu, podrobnosti viz ŠuvaDa 2005, 2007). Zároveň je také v Červené knize uváděna lokalita ve Slanských vrších (Holub in Čeřovský 1999), o této lokalitě se nepodařilo zjistit podrobnější informace.
V České republice jsou až dosud známy pouze tři lokality v Čechách, na Moravě a ve Slezsku tento hrachor nikdy nerostl. (obr. 2) Lokalita u Budeniček na Slánsku (též Budenická bažantnice) je s velkou pravděpodobností již dávnou minulostí, druh zde neověřil už F. A. Novák (novák 1923). Naopak v prostoru mezi Rožďalovicemi a Dymokury hrachor stále přežívá (viz Hadinec in ryDlo 2003) a nově byla objevena lokalita u Mšeného-lázní (viz kubát 2003). Další informace o hrachoru hrachovitém u nás a na Slovensku jsou uve-deny níže a také viz Štefánek (in press).
Přehled lokalit v České republice (údaje z herbářů PR, PRc, RoZ a z literatury)
Fyt. okr. 7a Libochovická tabule: Les Holínek u Mšeného-lázní kubát (2003): ...na severním okraji lesa Holínek nedaleko obce Mšené-lázně..., 30. V. 1992.
Fyt. okr. 7c Slánská tabule: Bažantnice u BudenicV bažantnici u Budenic, 19. VI. 1881 Fr. Bílek, PR. – V Budeničkách u Slaného v nízkém křoví bažantnice dost četně / V bažantnici v Budeničkách u Slaného, četně/, VI. 1881 K. Polák, PR, PRC. – Bažantnice u Budeniček blíž Zlonic, 18. VI. 1888 J. E. Kabát PR. – Nemus supra Budenice prope Zlonice, VI. 1894, Košťál, PR, PRC. – In dumetis sylvaticis ad Budeničky prope Slaný, planta rarissima, 29. VI. 1894 K. Polák, PRC. – V bažantnici v křoví Budenice, s. d., s. coll., PR. – V křoví bažantnice Budenice, s. d., s. coll., PR.Čelakovský (1882): In der Fasanerie bei Budenic unweit Zlonic, zwischen jungem Laubgebüsch an ziemlich umfangreicher Stelle recht zahlreich! (zuerst ganz zufällig von einem Schüller des Prof. Bílek in Schlan gesammelt). Čelakovský (1883): V bažantnici u Budeniček nedaleko Zlonic, v lupenatém mlází na prostoře dosti veliké v značném počtu! (nejprve od žáka prof. Bílka sbírán). bílek (1885): V lupenatém mlází Bažantnice u Budenic. krist (1936).

114
Štefánek: Lathyrus pisiformis v ČR a SR
Fyt. okr. 13a Rožďalovická tabule: Komárovský rybník Am Komarover Waldthal bei Dymokur (zwischen Nouzov und Rožďalovic), VII. 1871, E. Pospichal PR. – V lesním údolí u Nouzova blíž Rožďalovic, 1871, Ed. Pospíchal, PRC. – U Dymokur, 1876, Fr. Sitenský PR. – V údolí od Nouzova ke Komárovskému rybníku (u Dymokur), 23. VII. 1872, Čelákovský PR. – Rožďalovice: Komárovské údolí, 9. VII. 1896 J. Podpěra, PRC. – In dumetis prope Dymokury, 28. VI. 1901 K. Domin, PR. – Komárovské lesní údolí v Dymokursku, 29. VI. 1901, K. Domin, PRC. – Malý Nouzov u Dymokur, les u mlýna, 29. VI. 1901, Kašpar, PR. – U Dymokur, 29. VI. 1901, Kroulík, PRC. – Dymokury, VI. 1901 J. Židlický, ROZ. – Kraj lesa u Komárovských rybníků u Dymokur, 4. VI. 1909, E. Baudyš, PR. – M. Nouzov u Dymokur, s. d. Kašpar, PRC. – Dymokury, s. d. J. Pačes, ROZ. – Nouzov, podél pěšiny vedoucí lesem na pravém břehu přítoku Komárovského rybníka 1,3 km ZJZ od Malého Nouzova, asi 5 m od vody, na JV svahu, asi 40 m po proudu od místa, kde se potok přimyká ke svahu, 2. VI. 1988 J. Holub et J. Rydlo, ROZ. – Nouzov, podél pěšiny vedoucí lesem na pravém břehu přítoku Komárovského rybníka 1,3 km ZJZ od Malého Nouzova, asi 5 m od vody, na JV svahu, asi 40 m po proudu od místa, kde se potok přimyká ke svahu (asi 40 lodyh v deseti trsech, sterilní), 2. VII. 1988, J. Rydlo, ROZ. Čelakovský (1872): Unter einer Anzahl Pflanzen, die Herr Pospichal, Professor am Jiciner Gymnasium, ein sehr eifriger Jünger Florens, bei Kopidlno und Rozdalovic zwischen Podebrad und Jicin heuer gesammelt hat, sah ich auch den Lathyrus pisiformis L., der nach Mittheilung des Prof. Pospichal im Thale zwischen Nouzov und Rozdalovic in lichtem Waldgebüsch sehr häufig wächst. Čelakovský (1877): Na krajích lupenatých lesů v houští, a sice toliko u Dymokur v údolíčku mezi Nouzovem a Komárovským mlýnem dosti rozšířený, ačkoli nepříliš hojný! (nejprve nalezen od Pospíchala r. 1871). PosPíchal (1882): V Komárovském lesním údolí v západní bezrybničné polovici, hned pod mlýnem na louce šalvějová v houští kraje lesního počínajíc a v severním kraji až k rybníku se táhnouc, dosti četně. Před ní na krátce travnatých lučních okrajích květe Crepis praemorsa, Allium scorodoprassum a Euphorbia pilosa. Čelakovský (1897): Kraj lupenatého lesa u Dymokur; též v lesní pasece u Budenic blízko Zlonic. freiberG (1906): Na okraji lesních u Dymokur a Rožďalovic. bauDyŠ (1924): Rozšířen kolem všech rybníků komárovských tu a tam, ale i v rybničné části u Dymokur. krist (1936). PoDPěra (1938): V Komárovském lesním údolí hned pod mlýnem a v severním kraji až k rybníku dosti četně (Posp. 1872). Sr. Čelakovský, Uč. spol. 24. I. 1873. Rozšířen kolem všech rybníků komárovských, tu a tam i v rybničně části u Dymokur (Baudyš 1924). V Čechách roste ještě v bažantnici nedaleko Zlonic u Slaného (Bílek). Šachl (1970): Ojediněle v křovinatých okrajích lesa kolem Holského rybníka východně Rožďalovic, 205. Vyhnánek, 1964. V křovinách na severní straně Komárovského rybníka v místě zv. Na Bosně ojediněle ..., 204. Vyhnánek, 1965. Šachl (2000): V křovinách na severní straně Komárovského rybníka v místě zvaném Na Bosně, kolem Holského rybníka V Rožďalovic. Hadinec in ryDlo (2003): Svídnice, zbytky hájové květeny v kulturním smíšeném lese (výsadby modřínu, smrku a borovice v dubohabřině) na J svahu nad S břehem Komárovského rybníka v lesním smíšeném komplexu, 5. VII. 2002.
Metodika
sledování populačních parametrů na českých lokalitách
V průběhu trvání projektu byly sledovány základní charakteristiky velikosti a reprodukční schopnosti populace. Opakovaně (tj. červen v době kvetení, červenec v době dozrávání semen) byly každoročně (v průběhu let 2007–2011) navštíveny obě recentně známé lokality v ČR (jižně orientované svahy nad Komárovským rybníkem u Dymokur a les Holínek u Vrbice poblíž Mšeného-lázní).
Na obou lokalitách byly vždy dohledány na počátku sledování označené prýty (přesněji skupiny prýtů) hrachoru a zároveň byly označeny nově nalezené prýty (resp. skupiny prýtů).

115
Příroda, Praha, 31: 111–126, 2011
Jednotlivé skupiny prýtů (s velkou pravděpodobností geneticky totožné) byly označeny označeným hřebíkem v definované vzdálenosti. Počátkem června každého roku byl zaznamenán počet jednotlivých prýtů (a jejich velikost, přesněji délka), počet kvetoucích prýtů, květenství a počtu květů v květenství.
Obdobně byly v červenci (ca v polovině července, kdy se lusky otevírají a semena vypadávají) zaznamenány plodící prýty (tj. celkový počet plodných prýtů, počet lusků na prýtu a počet semen v lusku, jak plně vyvinutých, tak i semen abortovaných).
Je nutné zmínit, že jednotlivé skupinky prýtů jsou tvořeny s velkou pravděpodobností tvořeny geneticky totožnými prýty (tj. náleží k jednomu jedinci). Lathyrus pisiformis má pod zemí krátký oddenek, ze kterého vyrůstá obvykle jeden až čtyři prýty, v případě zastíněných jedinců to může být až devět či deset chabých prýtů o velikosti okolo 20 cm.
Zástin
Na všech (tj. obou) českých lokalitách a na dvou ze tří navštívených lokalit na Slovensku (lo ka litu na Dolném vrchu ve Slovenském krasu tvořilo netypické stanoviště v průseku hranice mezi Slovenskou republikou a Maďarskem), byla měřena hodnota celkového záření dopadající do bylinného patra. Tato hodnota (označeno dále jako zástin) byla zjištěna pomocí analýzy hemisférové fotografie (podrobnosti například vlasáková 2003, Černá 2008). Zástin byl vypo čítán v programu Gap Light Analyzer, GLA version 2 (frazer et al. 1999; www.ecostudies.org/gla/).
Fotografie byly pořízeny tzv. rybím okem (objektiv Sigma 8 mm F4 EX Fish-eye). Zmíněný objektiv je velmi širokoúhlý a schopný zachytit 180° zorného pole, tímto samozřejmě dochází díky projekci polokoule do půlkruhu k deformaci obrazu. Výše zmíněný program (Gap Light Analyzer, version 2) ovšem s touto deformací je schopen pracovat (dokáže projekci zpětně rekonstruovat do půlkruhu).
Fotografie byly pořízeny v místě nejhojnějšího výskytu hrachoru a zároveň v místě výskytu vitálních a plodných (kvetoucích) jedinců. Fotoaparát byl nastaven do vodorovné polohy (pomocí vodováhy), snímky byly orientovány k severu (nebo před analýzou byly tyto digitální fotografie upraveny otočením). Kvůli nežádoucím světelným efektům (světelné odrazy, přesvětlení některých míst atd.) by fotografování mělo být prováděno v době bez přímého slunečního záření (to v případě slovenských lokalit nebylo možné z časových důvodů). Nicméně GLA s těmito jevy počítá a lze v programu použít některé korekční prostředky (frazer et al. 1999).
Pedologické analýzy
Půdní vzorky byly odebrány standardním způsobem na dvou českých (les Holínek, u Ko márovského rybníka) a dvou slovenských lokalitách (Plešivská planina u obce Hámor a Plešivská planina nad obcí Vidová), po odstranění svrchního drnu (ca pět cm) byly odebrány bločky o velikosti přibližně 15 × 15 × 15 cm. Tyto vzorky byly následně usušeny při pokojové teplotě v laboratoři (ca 10 dní). Ze vzorků byly odebrány kamenité součásti a také zbytky rostlin (například kořeny). Vzorky byly pak rozemlety a analyzovány v pedologické laboratoři BÚ AV ČR Průhonice standardními pedologickými analýzami (viz například zbíral 1995). Zjišťovány byly následující hodnoty – obsah sušiny, aktuální pH, výměnné pH, obsah dusíku v sušině, celkový obsah uhlíku v sušině, obsah organického uhlíku v sušině, obsah vápníku, hořčíku, draslíku a fosforu.

116
Štefánek: Lathyrus pisiformis v ČR a SR
Fytocenologické snímky
Záznamy vegetace pomocí fytocenologických snímků zahrnovaly buď celou populaci Lathyrus pisiformis (v případě snímku u lesa Holínek) nebo podstatnou část vyskytujících se jedinců (Komárovský rybník). Fytocenologické snímky byly zaznamenány klasickou metodou pomocí odhadu pokryvnosti jednotlivých taxonu sedmičlennou Braun-Blanque to-vou stupnicí (Moravec et al. 1994).
výsledky
obr. 2. Současné rozšíření hrachoru hrachovitého (Lathyrus pisiformis) v České republice a na Slovensku.Fig. 2. Recent distribution of Lathyrus pisiformis in the Czech and Slovak Republics.
Popis recentních lokalit v Čechách
Svahy nad Komárovským rybníkem u DymokurLokalita u Komárovského rybníka leží v nadmořské výšce okolo 200 metrů na severním břehu Komárovského rybníka, asi 1,9 km SSV od železniční stanice Dymokury na pomezí katastrálních území obcí Břístev a Nouzov u Dymokur, souřadnice lokality jsou 50°16‘19,2‘‘N, 15°13‘28,5‘‘E. Území patří do fytogeografického podokresu 13a. Rožďalovická tabule.
Lokalita zahrnuje svažité (s relativním převýšením asi tři metry) břehy Komárovského rybníka (hrachor se vyskytuje i necelý metr od břehu rybníka) - menší světlinu (o rozloze několika metrů čtverečních) a okolními dubohabřiny v rozsáhlém lesním komplexu mezi Městcem Králové a Rožďalovicemi. (obr. 3). Vedle typických hájových druhů se zde vyskytují i některé prvky teplomilných doubrav, druhy teplých lesních lemů a také některé druhy patřící do širokolistých suchých trávníků svazu Bromion. Biotop charakterizuje následující fytocenologický snímek.
Po ověření výskytu hrachoru v roce 2002 bylo po dohodě se zástupcem majitele lesa (Lesy ČR) vybudována oplocenka, která má bránit přístupu zvěře a okusu hrachoru. Tato oplocenka o rozměru ca 10 × 30 m zahrnuje všechny nalezené exempláře hrachoru.

117
Příroda, Praha, 31: 111–126, 2011
V lesním komplexu v prostoru mezi Dymokury a Rožďalovicemi nelze vyloučit přítomnost dalších populací hrachoru hrachovitého, nicméně během průzkumu žádné nalezeny nebyly. Je ovšem nutné mít na vědomí, že zejména ve sterilním stavu je druh snadno přehlédnutelný a také to, že nezřídka bývá cílem potravního zájmu lesní zvěře.
Komárovský rybník u DymokurPlocha 10 × 15 m, expozice J–JV, sklon 5–20 %, 24. 6. 2007 E3, pokryvnost 85 %: Tilia cordata 4, Quercus robur 3, Ulmus glabra (mimo snímek), Carpinus betulus (mimo snímek)E2, pokryvnost 30 %: Tilia cordata 2, Ulmus glabra 1, Carpinus betulus +, Corylus avellana +, Cornus sanguinea +, Crataegus sp. rE1, pokryvnost 60 %: Astragalus glycyphyllos 2, Brachypodium sylvaticum 2, Campanula rapunculoides 2, Dactylis polygama 2, Galium odoratum 2, Hieracium murorum 2, Impatiens parviflora 2, Lathyrus vernus 2, Stellaria holostea 2, Anemone nemorosa 1, Brachypodium pinnatum 1, Calamagrostis arundinacea 1, Galium sylvaticum 1, Hieracium lachenalii 1, Lysimachia nummularia 1, Poa nemoralis 1, Vicia silvatica 1, Aegopodium podagraria +, Bromus benekenii +, Campanula persicifolia +, Carex muricata agg. +, Convallaria majalis +, Crataegus sp. juv. +, Festuca cf. rupicola +, Lathyrus niger +, Lathyrus pisiformis +, Lilium martagon +, Melica nutans +, Polygonatum odoratum +, Primula veris +, Pulmonaria obscura +, Tanacetum corymbosum +, Ulmus minor juv. +, Acer campestre juv. r, Carex digitata r, Fragaria vesca r, Galium boreale r, Geum urbanum r, Lapsana communis r, Ligustrum vulgare juv. r, Mellitis melissophyllum r, Mycelis muralis r, Rubus sp. juv. r, Sanicula europaea r, Cornus sanguinea juv. r, Veronica chamaedrys r, Vicia pisiformis r, Viola riviniana rE0 indet.: 20 %
obr. 3. Pohled na lokalitu hrachoru hrachovitého (Lathyrus pisiformis) u Komárovského rybníka nedaleko Dymokur.Fig. 3. Locality of Lathyrus pisiformis near the Komárovský pond (eastern part of Central Bohemia).

118
Štefánek: Lathyrus pisiformis v ČR a SR
Les Holínek u Vrbice-Podolí (Mšené-lázně)Tato lokalita leží při severním okraji lesa Holínek (na pomezí mezi lesem a loukou), přibližně 1,3 km JZ–JJZ od kaple v Mšeném-lázních, v katastrálním území obce Vrbice. Nadmořská výška se pohybuje okolo 245 m n. m. Souřadnice lokality jsou 50°21‘18,3‘‘N, 14°6‘38,5‘‘E.
Území náleží do termofytika, fytogeografický podokres 7a. Libochovická tabule. Les Holínek je izolovaným lesním celkem na severně orientovaných svazích údolí Mšenského potoka. Biotopem hrachoru je zde lesní plášť mezi lesem a občasně využívanou cestou vedoucí napříč loukou (Bromion). Tento lem je občasně kosen a zařadit se dá do lemových společenstev svazu Trifolion medii (obr. 4). Vegetaci zachycuje následující fytocenologický snímek (srovnej též kubát 2003).
Severní okraj lesa Holínek Plocha snímku 2,5 × 10 m, expozice S, sklon 1 %, 27. 6. 2007.E3 (stromy nerostou v ploše, ale plochu zastiňují), pokryvnost 40 %: Quercus robur 5, Fraxinus excelsior 2E2, pokryvnost 30 %: Carpinus betulus 2, Fraxinus excelsior 2, Prunus spinosa 2, Crataegus sp. 1, Li gu strum vulgare 1, Cornus sanguinea 1, Viburnum lantana 1, Acer campestre +E1, pokryvnost 85 % Arrhenatherum elatius 3, Melampyrum nemorosum 3, Securigera varia 3, Trifolium medium 3, Brachypodium pinnatum 2, Agrimonia eupatoria 1, Calamintha clinopodium 1, Carpinus betulus juv. 1, Festuca rubra 1, Fraxinus excelsior juv. 1, Prunus spinosa juv. 1, Trifolium repens 1, Vicia cracca 1, Viola cf. collina 1, Acer campestre juv. +, Bupleurum falcatum +, Crataegus sp. juv. +, Fragaria vesca +, Lathyrus pisiformis +, Lathyrus pratensis +, Plantago lanceolata +, Quercus robur juv. +, Rosa canina juv. +, Stellaria holostea +, Veronica chamaedrys +, Viola riviniana +, Achillea millefolium agg. r, Cornus sanguinea juv. r, Dactylis glomerata r, Galium sylvaticum r, Geum urbanum r, Ligustrum vulgare juv. r, Potentilla reptans r, Prunella vulgaris r, Ranunculus auricomus agg. r
obr. 4. Lokalita hrachoru hrachovitého (Lathyrus pisiformis) na okraji lesa Holínek u Mšeného-láz ní.Fig. 4. Locality of Lathyrus pisiformis at the edge of the wood called Holínek near the town Mšené-lázně, northern part of Central Bohemia.

119
Příroda, Praha, 31: 111–126, 2011
Poznámky ke slovenským lokalitám
Plešivská planina u obce HámorSouřadnice 48°34‘N, 20°24,3‘E, 489 m n. m.Tato lokalita leží při horní hraně Plešivské planiny, na jejím západně exponovaném svahu nad obcí Hámor ca 1,6 km SSZ od železniční stanice Plešivec. Hrachor zde roste především v teplomilných doubravách svazu Quercion pubescenti-petraeae a nehojně se rostliny objevují v lemových biotopech a navazujících trávnících svazu Bromo pannonici-Festucion pallentis (podrobnosti včetně fytocenologického snímku viz ŠuvaDa 2007). V roce 2010 zde bylo zaznamenáno ca 120 prýtů, ovšem pouze šest kvetoucích.
Plešivská planina nad obcí VidováSouřadnice 48°34,3‘N, 20°26,4‘E, 508 m n. m.Lokalitu tvoří J-JJV exponované svahy nad obcí Vidová ca 1 km SSZ od železniční zastávky Vidová. Jde o teplomilné doubravy svazu Quercion pubescenti-petraeae, na světlinách přecházející do společenstev Festucion valesiaceae (zčásti také v lemových společenstvech svazu Geranion sanguinei). Celkově zde bylo na několika mikrolokalitách zaznamenáno v roce 2010 několik stovek sterilních (ca 500) a přes osmdesát kvetoucích prýtů. Vzhledem k zvýšenému okusu zvěří byla část populace ohrazena provizorní ohrádkou. Podrobnější popis uvádí ŠuvaDa (2007).
Přibližně 100 m východně od předcházející populace se vyskytuje menší porost hrachoru (ca deset jedinců, statných a bohatě plodných) v lesním lemu v sedle poblíž silničky vedoucí od obce Vidová na planinu (označeno jako Vidová II.).
Dolný vrchSouřadnice 48°34,3‘N, 20°45‘E až 48°34,25‘N, 20°44,7‘E, ca 500 m n. m.Poslední lokalita hrachoru, která je v současnosti ze Slovenského krasu známa, je poněkud odlišná od předcházejících. Jde o přibližně 300 m dlouhý úsek průseku hraniční čáry mezi Slovenskem a Maďarskem, asi 4,2–4,5 km JJZ od železniční stanice Hrhov. Zaznamenáno zde bylo asi deset kvetoucích a několik desítek sterilních prýtů. Jde o biotop vytvořený pravidelným vysekáváním dřevin na státní hranici, většinou to jsou husté porosty ca 0,5–1 m vysokých zmlazujících habrů, méně pak fragmenty suchých trávníků svazu Bromion erecti nebo lemová společenstva.
Další historická mikrolokalita na Dolném vrchu (ŠuvaDa 2007), nacházející se ca 750 m západně od předcházející, v roce 2010 ověřena nebyla.
Analýza zástinuVýsledky analýzy zástinu lokalit jsou uvedeny v tabulce 1. Jak z této tabulky vyplývá, nejvíce zastíněná je lokalita u Komárovského rybníka, naopak nejméně zastíněné jsou lokality ve Slovenském krasu, přičemž nejméně zastíněná je malá populace na JV orientovaném okraji lesa na hraně Plešivské planině u Vidové. Množství dopadajícího světla ovlivňuje u hrachoru především kvetení a následné tvorby lusků, na více zastíněných místech hrachory přežívají ve sterilním stavu (nejméně kvetoucích jedinců bylo zaznamenáno na lokalitě u Komárovského rybníka, naopak na studovaných lokalitách ve Slovenském krasu bývají rostliny pravidelně kvetoucí a poměrně statné). Je ovšem nutné poznamenat, že roli hraje především zastínění konkrétní rostliny (světlost mikrostanoviště).
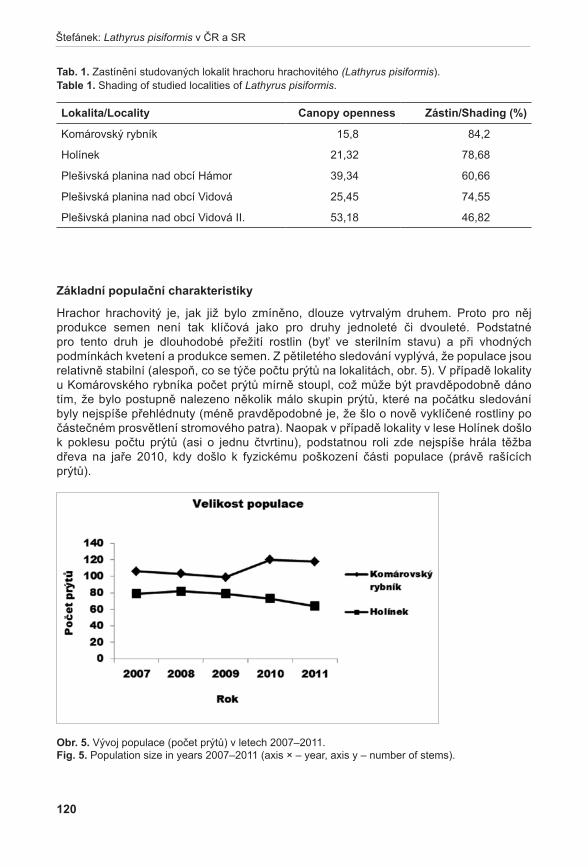
120
Štefánek: Lathyrus pisiformis v ČR a SR
tab. 1. Zastínění studovaných lokalit hrachoru hrachovitého (Lathyrus pisiformis).table 1. Shading of studied localities of Lathyrus pisiformis.
lokalita/locality canopy openness Zástin/shading (%)
Komárovský rybník 15,8 84,2
Holínek 21,32 78,68
Plešivská planina nad obcí Hámor 39,34 60,66
Plešivská planina nad obcí Vidová 25,45 74,55
Plešivská planina nad obcí Vidová II. 53,18 46,82
Základní populační charakteristiky
Hrachor hrachovitý je, jak již bylo zmíněno, dlouze vytrvalým druhem. Proto pro něj produkce semen není tak klíčová jako pro druhy jednoleté či dvouleté. Podstatné pro tento druh je dlouhodobé přežití rostlin (byť ve sterilním stavu) a při vhodných podmínkách kvetení a produkce semen. Z pětiletého sledování vyplývá, že populace jsou relativně stabilní (alespoň, co se týče počtu prýtů na lokalitách, obr. 5). V případě lokality u Komárovského rybníka počet prýtů mírně stoupl, což může být pravděpodobně dáno tím, že bylo postupně nalezeno několik málo skupin prýtů, které na počátku sledování byly nejspíše přehlédnuty (méně pravděpodobné je, že šlo o nově vyklíčené rostliny po částečném prosvětlení stromového patra). Naopak v případě lokality v lese Holínek došlo k poklesu počtu prýtů (asi o jednu čtvrtinu), podstatnou roli zde nejspíše hrála těžba dřeva na jaře 2010, kdy došlo k fyzickému poškození části populace (právě rašících prýtů).
obr. 5. Vývoj populace (počet prýtů) v letech 2007–2011.Fig. 5. Population size in years 2007–2011 (axis × – year, axis y – number of stems).
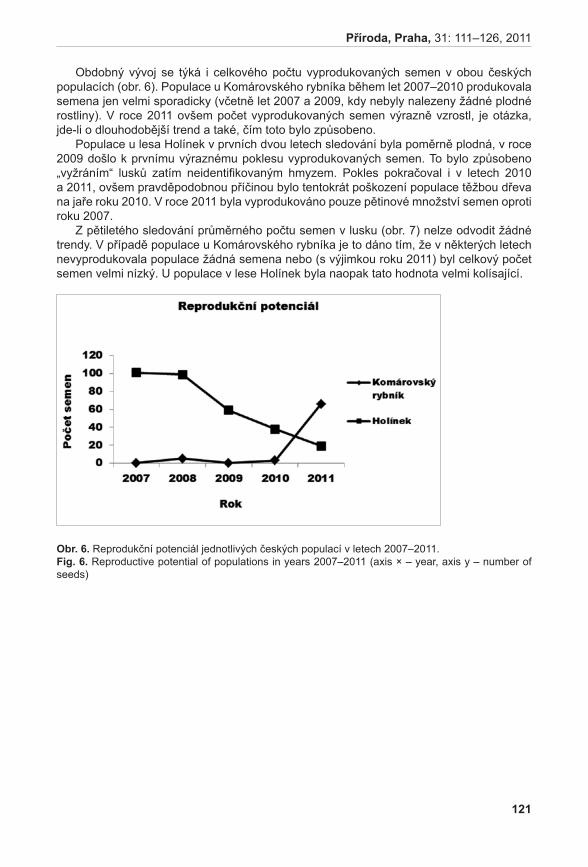
121
Příroda, Praha, 31: 111–126, 2011
Obdobný vývoj se týká i celkového počtu vyprodukovaných semen v obou českých populacích (obr. 6). Populace u Komárovského rybníka během let 2007–2010 produkovala semena jen velmi sporadicky (včetně let 2007 a 2009, kdy nebyly nalezeny žádné plodné rostliny). V roce 2011 ovšem počet vyprodukovaných semen výrazně vzrostl, je otázka, jde-li o dlouhodobější trend a také, čím toto bylo způsobeno.
Populace u lesa Holínek v prvních dvou letech sledování byla poměrně plodná, v roce 2009 došlo k prvnímu výraznému poklesu vyprodukovaných semen. To bylo způsobeno „vyžráním“ lusků zatím neidentifikovaným hmyzem. Pokles pokračoval i v letech 2010 a 2011, ovšem pravděpodobnou příčinou bylo tentokrát poškození populace těžbou dřeva na jaře roku 2010. V roce 2011 byla vyprodukováno pouze pětinové množství semen oproti roku 2007.
Z pětiletého sledování průměrného počtu semen v lusku (obr. 7) nelze odvodit žádné trendy. V případě populace u Komárovského rybníka je to dáno tím, že v některých letech nevyprodukovala populace žádná semena nebo (s výjimkou roku 2011) byl celkový počet semen velmi nízký. U populace v lese Holínek byla naopak tato hodnota velmi kolísající.
obr. 6. Reprodukční potenciál jednotlivých českých populací v letech 2007–2011.Fig. 6. Reproductive potential of populations in years 2007–2011 (axis × – year, axis y – number of seeds)
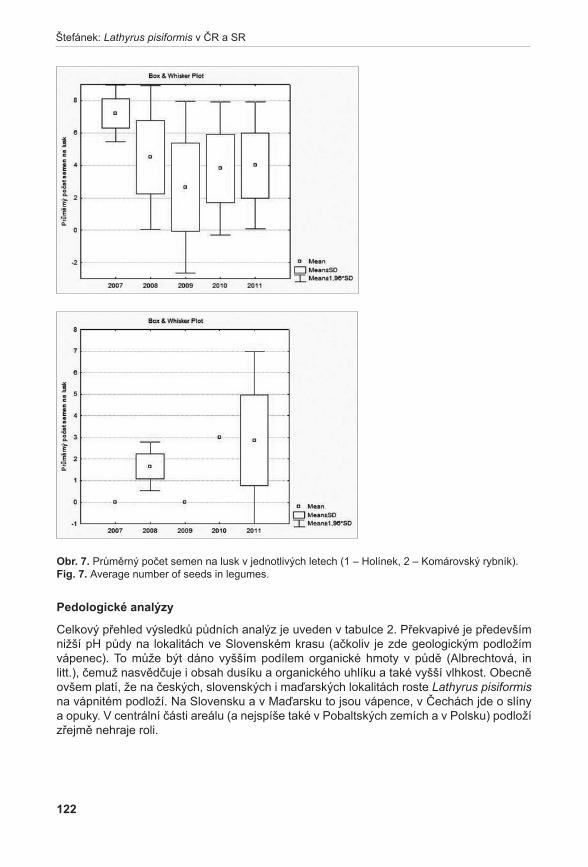
122
Štefánek: Lathyrus pisiformis v ČR a SR
1
2
obr. 7. Průměrný počet semen na lusk v jednotlivých letech (1 – Holínek, 2 – Komárovský rybník).Fig. 7. Average number of seeds in legumes.
Pedologické analýzy
Celkový přehled výsledků půdních analýz je uveden v tabulce 2. Překvapivé je především nižší pH půdy na lokalitách ve Slovenském krasu (ačkoliv je zde geologickým podložím vápenec). To může být dáno vyšším podílem organické hmoty v půdě (Albrechtová, in litt.), čemuž nasvědčuje i obsah dusíku a organického uhlíku a také vyšší vlhkost. Obecně ovšem platí, že na českých, slovenských i maďarských lokalitách roste Lathyrus pisiformis na vápnitém podloží. Na Slovensku a v Maďarsku to jsou vápence, v Čechách jde o slíny a opuky. V centrální části areálu (a nejspíše také v Pobaltských zemích a v Polsku) podloží zřejmě nehraje roli.
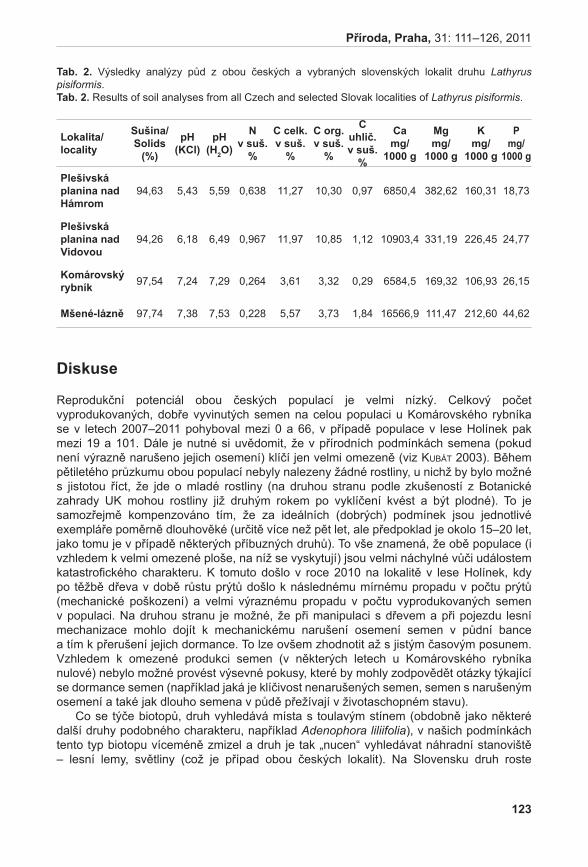
123
Příroda, Praha, 31: 111–126, 2011
tab. 2. Výsledky analýzy půd z obou českých a vybraných slovenských lokalit druhu Lathyrus pisiformis.tab. 2. Results of soil analyses from all Czech and selected Slovak localities of Lathyrus pisiformis.
lokalita/ locality
sušina/ solids
(%)
pH (Kcl)
pH (H2o)
nv suš.
%
c celk.v suš.
%
c org.v suš.
%
c uhlič.v suš.
%
camg/
1000 g
Mgmg/
1000 g
Kmg/
1000 g
Pmg/
1000 g
Plešivská planina nad Hámrom
94,63 5,43 5,59 0,638 11,27 10,30 0,97 6850,4 382,62 160,31 18,73
Plešivská planina nad vidovou
94,26 6,18 6,49 0,967 11,97 10,85 1,12 10903,4 331,19 226,45 24,77
Komárovský rybník 97,54 7,24 7,29 0,264 3,61 3,32 0,29 6584,5 169,32 106,93 26,15
Mšené-lázně 97,74 7,38 7,53 0,228 5,57 3,73 1,84 16566,9 111,47 212,60 44,62
diskuse
Reprodukční potenciál obou českých populací je velmi nízký. Celkový počet vyprodukovaných, dobře vyvinutých semen na celou populaci u Komárovského rybníka se v letech 2007–2011 pohyboval mezi 0 a 66, v případě populace v lese Holínek pak mezi 19 a 101. Dále je nutné si uvědomit, že v přírodních podmínkách semena (pokud není výrazně narušeno jejich osemení) klíčí jen velmi omezeně (viz kubát 2003). Během pětiletého průzkumu obou populací nebyly nalezeny žádné rostliny, u nichž by bylo možné s jistotou říct, že jde o mladé rostliny (na druhou stranu podle zkušeností z Botanické zahrady UK mohou rostliny již druhým rokem po vyklíčení kvést a být plodné). To je samozřejmě kompenzováno tím, že za ideálních (dobrých) podmínek jsou jednotlivé exempláře poměrně dlouhověké (určitě více než pět let, ale předpoklad je okolo 15–20 let, jako tomu je v případě některých příbuzných druhů). To vše znamená, že obě populace (i vzhledem k velmi omezené ploše, na níž se vyskytují) jsou velmi náchylné vůči událostem katastrofického charakteru. K tomuto došlo v roce 2010 na lokalitě v lese Holínek, kdy po těžbě dřeva v době růstu prýtů došlo k následnému mírnému propadu v počtu prýtů (mechanické poškození) a velmi výraznému propadu v počtu vyprodukovaných semen v populaci. Na druhou stranu je možné, že při manipulaci s dřevem a při pojezdu lesní mechanizace mohlo dojít k mechanickému narušení osemení semen v půdní bance a tím k přerušení jejich dormance. To lze ovšem zhodnotit až s jistým časovým posunem. Vzhledem k omezené produkci semen (v některých letech u Komárovského rybníka nulové) nebylo možné provést výsevné pokusy, které by mohly zodpovědět otázky týkající se dormance semen (například jaká je klíčivost nenarušených semen, semen s narušeným osemení a také jak dlouho semena v půdě přežívají v životaschopném stavu).
Co se týče biotopů, druh vyhledává místa s toulavým stínem (obdobně jako některé další druhy podobného charakteru, například Adenophora liliifolia), v našich podmínkách tento typ biotopu víceméně zmizel a druh je tak „nucen“ vyhledávat náhradní stanoviště – lesní lemy, světliny (což je případ obou českých lokalit). Na Slovensku druh roste

124
Štefánek: Lathyrus pisiformis v ČR a SR
v prosvětlených a zakrslých teplomilných doubravách. Druh tedy preferuje mírné až střední zastínění, v otevřených plochách hrachor potlačují konkurenčně silnější druhy (zejména trávy). Na okraji svého areálu (což je případ českých, slovenských a maďarských lokalit) Lathyrus pisiformis vyhledává vápnité podloží (což se nemusí nutně odrazit v půdní reakci).
Závěr
Pětileté sledování českých populací druhu Lathyrus pisiformis zaměřené zejména na základní charakteristiky populační biologie tohoto kriticky ohroženého druhu objasnilo některé základní otázky, týkající se produkce semen v populacích a velikost a dynamiku obou českých populací.
Zatímco obě populace druhu, alespoň co se množství vzešlých prýtů, se zdají být relativně stabilní (během pěti let nedošlo k výrazným výkyvům v počtu prýtů), některé události „katastrofického“ charakteru (zejména v případě lokality les Holínek) výrazně zasáhly do počtu vyprodukovaných semen.
summary
Five-year monitoring of Czech populations of Lathyrus pisiformis (localities: Holínek wood near the town Mšené-lázně in the northern part of Central Bohemia and wood nearby Komárovský pond in the eastern part of Central Bohemia), which focused on basic characteristics of population biology of this critically endangered species, provided answers to some questions regarding seed production, size and dynamics in both Czech populations.
While both populations seemed to be relatively stable (during five years there were no important differences in number of stems), some events of “catastrophic character” (such as timber harvesting or insects feeding on seeds) significantly affected number of produced seeds.
Poděkování
Studium hrachoru hrachovitého bylo financováno projektem MŠMT VaV 2B06178. Poděkování patří J. Hadincovi, J. Rydlovi, K. Kubátovi a M. Fialové za spolupráci v terénu či informace ohledně jednotlivých populací hrachoru, Z. Münzbergové za pomoc při zpracování dat, L. Černé za podklady a informace k zpracování dat o zástinu, O. Šídovi a M. Ducháčkovi za umožnění studia herbářových položek v botanickém oddělení NM, I. Štefánkové za pomoc při práci v terénu.
Velké díky patří R. Šuvadovi (Správa NP Slovenský kras) za množství informací o slovenských a některých maďarských lokalitách studovaného druhu a zejména za provedení po lokalitách ve Slovenském krasu, pracovníkům Správy NP Slovenský kras patří dík za pohostinnost.
Díky patří také M. Němečkovi (Lesní správa Nymburk) za vstřícný postoj a vybudování ohrádky na lokalitě u Komárovského rybníka.

125
Příroda, Praha, 31: 111–126, 2011
literatura
baleviČius k., laPelë M. & PaltanaviČius s. [eds] (1992): Lietuvos raudonoji knyga. [The Red Data Book of Lithuania]. – Lithuanian Department of Environmental Conservancy, Vilnius.
bässler M. (1973): Revision der eurasiatischen Arten von Lathyrus L. Sect. Orobus (L.) Gren. et Godr. – Feddes Repert. 84: 329–447.
bauDyŠ e. (1924): Příspěvek ke „Květeně poříčí Cidliny a Mrliny“. – Sborn. Klubu Přírod. Brno 6 (1923): 44–71.
bílek f. (1885): Soustavný přehled rostlin cévnatých v okolí Slaného samorostlých a obecně pěstovaných II. – Výroční zpráva obecného vyššího Gymnázia Slané 5: 3–36.
Čelakovský L. (1872): Correspondenzen. – Österr. Bot. Zeitschr. 22: 34–35. Čelakovský L. (1877): Prodromus květeny české. Vol. 3. – Arch. Přírod. Výzk. Čech, Praha.Čelakovský L. (1882): Resultate der botanischen Durchforschung Böhmens im Jahre 1881. – S.-B.
Königl. Böhm. Ges. Wiss. Prag 1881: 360–395.Čelakovský L. (1883): Prodromus květeny české. Vol. 4. – Arch. Přírod. Výzk. Čech, Praha.Čelakovský L. (1897): Analytická květena Čech, Moravy a rakouského Slezska. 3. ed. – Praha.Černá l. (2008): Srovnávací populační dynamika druhů Anthericum liliago L. a Anthericum ramosum
L. – Ms. [Dipl. práce, depon. in Knihovna Katedry botaniky PřF UK, Praha.]Čeřovský j. [ed.] (1999): Červená kniha ohrožených a vzácných druhů rostlin ČR a SR. Vol. 5. Vyšší
rostliny. – Príroda, Bratislava.frazer G. w., canhaM c. D. & lertzMan k. P. (1999): Gap Light Analyzer (GLA), Version 2.0: Imaging
software to extract canopy structure and gap light transmission indices from true-colour fisheye photographs, users manual and program documentation. – Simon Fraser University, Burnaby, British Columbia, and the Institute of Ecosystem Studies, Millbrook, New York; www.rem.sfu.ca/forestry/downloads/gap_light_analyzer.htm.
freiberG f. (1906): Květena. – In: brzák f. & ČeČetka f. j., Poděbradsko, pp. 76–100, Poděbrady.kaźMierczakowa r. & zarzycki k. (2001): Polish red data book of plants. – Polish Academy of Science,
Cracow.kenicer G. j., kajita t., PenninGton t. r. & Murata j. (2005): Systematics and biogeography of Lathyrus
(Leguminosae) based on internal transcribed spacer and cpDNA sequence data. – Amer. J. Bot. 92: 1199–1209.
konviČka M., Čížek l. & beneŠ j. (2004): Ohrožený hmyz nížinných lesů: ochrana a management. – Sagittaria, Olomouc.
krist v. (1936): Příspěvek k poznání květeny Československé republiky I. (Ad floram Čechoslovenicam additamentum I.) – Spisy Přírod. Fak. Masaryk. Univ. Brno 222: 1–13.
kubát k. (2003): Lathyrus pisiformis L. – Severočes. Přír. 35: 103–104.kubát k., hrouDa l., chrtek j. jun., kaPlan z., kirschner j. & ŠtěPánek j. [eds] (2002): Klíč ke květeně
České republiky. – Academia, Praha.Moravec j. [ed.] (1994): Fytocenologie (Nauka o vegetaci). – Academia, Praha.Moravec j., balátová-tuláČková e., blažková D., haDaČ e., hejný s., husák Š., jeník j., kolbek j.,
krahulec f., kroPáČ z., neuhäusl r., rybníČek k., řehořek v. & vicherek j. (1995): Rostlinná společenstva České republiky a jejich ohrožení. Ed. 2. – Severočes. Přír., Příl. 1995: 1–206.
novák f. a. (1923): Plantae Bohemicae rariores vel minus cognitae. – Sborn. Klubu Přírod. Praha 1921–1922: 45–51.
PoDPěra j. (1938): Poznámky ku geobotanickému výzkumu květeny na středním Pojizeří. – Pamět. Spis. 250 let gymn. Mladá Boleslav (1688–1938): 99–13.
PosPíchal e. (1882): Květena poříčí Cidliny a Mrliny. – Arch. Přírod. Pros. Čech díl IV. no. 5: Botan. odd.: 1–99.
ryDlo j. (ed.) (2003): Výsledky floristického kursu České botanické společnosti v Nymburce v roce 2002. – Muz. Současn., ser. natur., 18: 3–105.
slavík b. (ed.) (1995): Květena České republiky. Vol. 4. – Academia, Praha.Šachl j. (1970): Druhý příspěvek ke Květeně Poděbradska. – Čas. Nár. Muz., odd. přírodovědný, 137,
3/4:1–8.

126
Štefánek: Lathyrus pisiformis v ČR a SR
Šachl j. (2000): Liány u nás na Poděbradsku, Nymbursku, ve Středním Polabí. – Vlastiv. Zprav. Polabí 34: 170–189.
Štefánek M. (in press): Hrachor hrachovitý - nesnesitelná těžkost bytí. – Živa.ŠuvaDa r. (2005): Hrachor hrachovitý (Lathyrus pisiformis L.) na Slovensku. – Chránené územia
Slovenska, Banská Bystrica, 65: 22–23.ŠuvaDa r. (2007): Lathyrus pisiformis L. v teplomilných dubinách Slovenského krasu. – Bull. Slov. Bot.
Spoločn. 29: 106–110.vlasáková b. (2003): Ekologie Polytrichum commune a Polytrichum juniperinum na mraveništích
druhu Lasius flavus. – Ms. [Dipl. práce, depon. in Knihovna Katedry botaniky PřF UK, Praha.]zbíral j. (1995): Analýza půd I, Jednotné pracovní postupy. – Ústřední kontrolní a zkušení ústav
zemědělský, Brno.

127
Příroda, Praha, 31: 127–147, 2011
Faktory ovlivňující populační dynamiku kriticky ohroženého druhu Ligularia sibirica
Factors influencing the population dynamics of a critically endangered plant species Ligularia sibirica anna heinken-šMídová1, 2 & ZuZana MünZbergová1, 3
1Botanický ústav akademie věd České republiky, Zámek 1, CZ-252 43 Průhonice2Katedra botaniky, Přírodovědecká fakulta, Univerzita Palackého, Šlechtitelů 11,CZ-783 71 Olomouc; [email protected] botaniky, Přírodovědecká fakulta, Univerzita Karlova, Benátská 2, CZ-128 01 Praha; [email protected]
Abstract: Good knowledge of population biology, genetic diversity, and environmental factors is necessary for efficient conservation of rare species. Our study species is Ligularia sibirica, a critically endangered plant species protected under the EU Habitats Directive (Annex II).
We estimated the genetic diversity and the main habitat conditions of the localities, and assessed their importance for the population dynamics of L. sibirica in the Czech and Slovak Republic. The population dynamics was studied using population transition matrices. Most of the studied populations were performing well and only those growing in deteriorating habitats due to drainage showed a decreasing trend. Analyses of elasticity showed that transitions that most contribute to the population growth rate vary with the habitat condition. The genetic diversity was estimated using allozyme electrophoresis. Our findings suggest that the populations can retain sufficient genetic variability and that most of the variability is within populations (80.86%). The populations were significantly genetically differentiated. Those growing in nitrogen-poor habitats (nutrient-poor habitats) are performing better than populations growing in nitrogen-rich habitats (nutrient-rich habitats), even though the individuals are smaller and they have lower seed production in conditions poor in nutrients. Comparing the importance of genetic diversity, habitat conditions, and population size for the performance of the L. sibirica population, we identified the habitat conditions as the most important factor.
Our results indicate that the population size and the genetic diversity are not a major problem for L. sibirica and conservation of the species should focus on restoring habitat conditions of drained localities.Keywords: genetic diversity, habitat quality, Ligularia sibirica, population size, population viability
Abstrakt: Dobrá znalost populační biologie, genetické variability a stanovištních charakteristik je nezbytná pro efektivní ochranu vzácných druhů rostlin. Studovaly jsme kriticky ohrožený druh chráněný směrnicí o stanovištích (příloha II), Ligularia sibirica.
Stanovily jsme genetickou variabilitu, hlavní stanovištní charakteristiky lokalit a zhod-no tily jsme jejich význam pro populační dynamiku L. sibirica v České republice a na Slo-vensku. Populační dynamika byla studována pomocí populačních přechodových matic. Většina sledovaných populací prospívá dobře a pouze populace rostoucí na odvodněných stanovištích měly klesající trend. Analýzy elasticity ukázaly, že přechody, které nejvíce při-

128
Heinken-Šmídová & Münzbergová: Faktory ovlivňující populační dynamiku Ligularia sibirica
spívají k růstové rychlosti populací, se liší dle kvality stanoviště. Genetická variabilita byla stanovena pomocí metody isoenzymů. Naše zjištění ukazují, že populace jsou schopny uchovat dostatek genetické variability a že většina variability se nachází v uvnitř populací (80,86 %). Výsledky dále ukázaly, že populace rostoucí na stanovištích chudých na dusík (stanoviště chudé na živiny) prosperují lépe než populace rostoucí na stanovištích bohatých na dusík (stanoviště bohaté na živiny), a to i přesto, že je velikost jednotlivců menší a mají nižší produkci semen. Nakonec jsme regresní analýzou zjistily, který z faktorů (genetická variabilita, stanovištní podmínky nebo velikost populace) ovlivňují vitalitu populací L. sibirica. Ukázaly jsme, že tímto faktorem je kvalita stanoviště.
Naše výsledky ukazují, že velikost populace a genetická variabilita nejsou hlavní problém při ochraně druhu Ligularia sibirica. Ochrana tohoto vzácného druhu by se měla zaměřit hlavně na obnovení stanovištních podmínek odvodněných lokalit.Klíčová slova: genetická variabilita, Ligularia sibirica, kvalita stanoviště, velikost populací, životaschopnost populací
Úvod
V současné době jsou v Evropě tisíce rostlinných druhů považovány za ohrožené. Nicméně jen několik stovek těchto druhů má právní ochranu (ozinGa & schaMinée 2005), která je především na úrovni jednotlivých států. Od roku 1992 existuje také seznam Evropské unie na ochranu vzácných druhů rostlin tzv. Směrnice o stanovištích, příloha II (council of the euroPean coMMunity 1992). Většina ze zde uvedených druhů jsou endemity nebo relikty, jejichž rozšíření je v Evropě buď omezeno na malou oblast, nebo vyžadují specifické sta-no vištní podmínky.
Hlavním cílem ochrany výše zmiňovaných vzácných druhů rostlin ve smyslu jejich zachování a managementu je udržení životaschopných populací (soulé 1987). Jednou z nejčastěji používaných metod pro hodnocení životaschopnosti populací je analýza životaschopnosti populací (population viability analyses, PVA, MenGes 2000). PVA se zabývá především rychlostí růstu populací a rizikem jejich zániku. Kromě toho může být PVA také použita k identifikaci kritických fází života populací (raMula 2006).
K ohrožení nebo dokonce k vyhynutí celé populace či druhu může přispívat velké množství faktorů. Některé z těchto faktorů mohou být specifické pouze pro konkrétní druhy rostlin (např. specializované opylování a klíčení). Nicméně, na základě výsledků řady studií prováděných na vzácných druzích rostlin, můžeme identifikovat tři hlavní faktory, které ovlivňují životaschopnost populací většiny druhů (fischer & Matthies 1998, oosterMeijer et al. 1998, verGeer et al. 2003). Jsou to kvalita stanoviště, velikost populace a genetická variabilita. Tyto faktory mohou působit buďto jednotlivě nebo ruku v ruce.
Změny v kvalitě stanoviště jsou často zmiňovány jako nejdůležitější faktory ovlivňující životaschopnost populací (brys et al. 2005, schleuninG & Matthies 2009). Tyto změny souvisejí se změnami v dostupnosti živin a vody (collinG et al. 2002, brys et al. 2005), se sukcesí anebo se změnou obhospodařování (oosterMeijer et al. 1996). Negativní změny v kvalitě stanoviště mnohdy vedou k nižší životaschopnosti populací, což následně vede ke zmenšení velikosti populace (ouborG et al. 2006).
Změny velikosti populace mohou být buď důsledkem snížení fitness populace nebo zničením části populace (oosterMeijer et al. 2003, leiMu et al. 2006). Tyto změny často také souvisejí se změnami kvality stanoviště. Na druhé straně, malá velikost populace vede ke ztrátě genetické variability v důsledku náhodného genetického driftu nebo inbreedingu (oosterMeijer et al. 2003) či v důsledku narušení interakce mezi rostlinami a opylovači (ÅGren 1996).

Příroda, Praha, 31: 127–147, 2011
129
Snížení genetické variability je často důsledkem zmenšení velikosti populace (viz výše; oosterMeijer et al. 2003, leiMu et al. 2006). Krátkodobě je snížení genetické variability spojeno s nižší fitness (např. nízká produkce a klíčivost semen, vysoká úmrtnost a špatný růst semenáčků) (ellstranD & elaM 1993, oosterMeijer et al. 2003). Dlouhodobě může nižší genetická variabilita snížit schopnost druhů se přizpůsobit měnícím se podmínkám stanoviště a tím vést k zániku celé populace (ouborG et al. 2006).
V tomto článku jsme shrnuly poznatky, které jsme získaly v podrobné studii rostlinného reliktu chráněného dle směrnice o stanovištích, příloha II směrnice Rady Evropského společenství (1992), Ligularia sibirica (L.) Cass. Tento druh jsme sledovaly v průběhu čtyř let na 11 populacích v České a Slovenské republice. Studovaly jsme podrobnou populační dynamiku druhu, genetickou variabilitu a kvalitu stanoviště. V tomto článku bychom chtěly diskutovat o tom, jak studované populace prosperují a jaké jsou kritické fáze jejich životního cyklu. Poté se zmíníme, jaká je genetická variabilita a kvalita stanoviště sledovaných populací. A na konec se budeme zabývat tím, který z těchto tří faktorů (genetická variabilita, kvalita stanoviště nebo velikost populace) má vliv na životaschopnost populací L. sibirica.
Metodika
studovaný druh
Ligularia sibirica (L.) Cass. (Asteraceae) je vytrvalá bylina s přízemní růžicí listů a krátkým oddenkem. Během kvetení (polovina července – konec srpna) vytváří v průměru dvě kvetoucí lodyhy s 20 květy v květenství. Květy jsou oboupohlavní, opylované hmyzem. Na podzim dozrávají ochmýřené nažky (dále semena), které jsou na krátkou vzdálenost šířeny buď větrem, nebo gravitací (ŠMíDová et al. 2011). Semena jsou částečně požírána larvami hmyzího druhu z čeledi Gelechiidae (Lepidoptera), které se vyvíjejí uvnitř květu (Elsner & Patočka, ústní sdělení). Ligularia sibirica je diploidní rostlina (liu 2004) s převažujícím pohlavním rozmnožováním. Klonální růst je typu phalanx (sensu Doust 1981), tj. ve formě trsu s pomalu radiálně se šířícími rametami.
Ligularia sibirica je rostlinný druh, který preferuje plné oslunění. Vyskytuje se v různých typech mokřadů, jako jsou vlhké louky, vápnitá slatiniště, přechodné rašeliniště a lužní lesy s výškou hladiny spodní vody při povrchu půdy (slavík 2004). Jedná s o druh s rozsáhlým eurosibiřským areálem. V Evropě se vyskytuje na několika izolovaných lokalitách – Lotyšsko, Estonsko, Ukrajina, Rumunsko, Maďarsko (zanikla), Rakousko (druhotný výskyt), Chorvatsko, Česká republika, Slovensko a Polsko. Jediná populace v Bulharsku pravděpodobně zanikla (M. Hájek, osobní sdělení). Nejzápadnějším územím jejího výskytu jsou pohoří centrální a jižní Francie (Meusel & jäGer 1992). Ligularia sibirica je považována za glaciální (Mattauch 1936) nebo postglaciální relikt (henDrych 2003, ŠMíDová et al. 2011), tedy chladnomilný druh, který byl v pozdním glaciálu až preboreálu na místech připomínajících tajgovou mokřadní vegetaci hojnější a dnes je jeho rozšíření fragmentované. Dále je klasifikována jako kriticky ohrožený druh České republiky (Procházka 2001), ohrožený druh Slovenské republiky (feráková et al. 2001) a je chráněn směrnicí EU Habitat Directive, Annex II (council of the euroPean coMMunity 1992).
studované populace
Studovány byly populace v České republice a na Slovensku mající více jak 50 dospělých jedinců L. sibirica. V případě studia genetické variability a stanovištních podmínek jsme studovaly všechny populace v České a Slovenské republice, tj. 20 populací. Studium
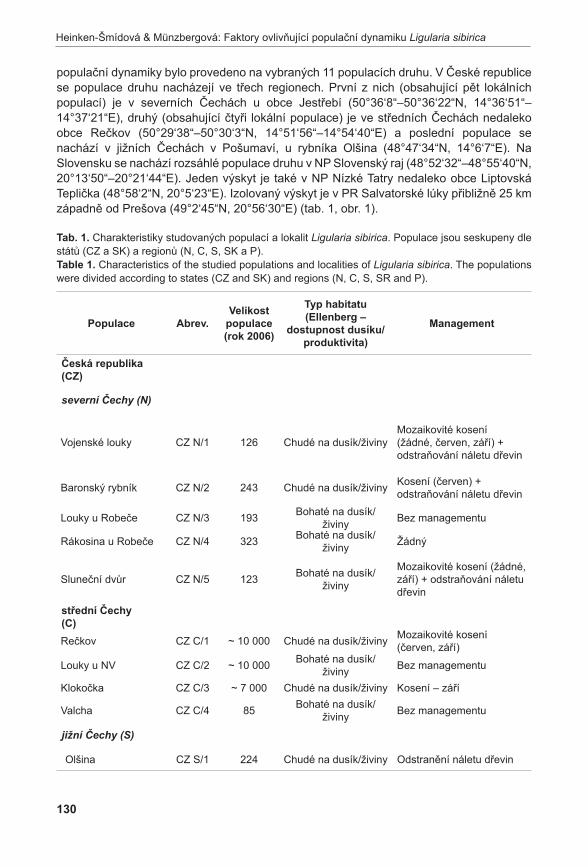
130
Heinken-Šmídová & Münzbergová: Faktory ovlivňující populační dynamiku Ligularia sibirica
populační dynamiky bylo provedeno na vybraných 11 populacích druhu. V České republice se populace druhu nacházejí ve třech regionech. První z nich (obsahující pět lokálních populací) je v severních Čechách u obce Jestřebí (50°36‘8“–50°36‘22“N, 14°36‘51“–14°37‘21“E), druhý (obsahující čtyři lokální populace) je ve středních Čechách nedaleko obce Rečkov (50°29‘38“–50°30‘3“N, 14°51‘56“–14°54‘40“E) a poslední populace se nachází v jižních Čechách v Pošumaví, u rybníka Olšina (48°47‘34“N, 14°6‘7“E). Na Slovensku se nachází rozsáhlé populace druhu v NP Slovenský raj (48°52‘32“–48°55‘40“N, 20°13‘50“–20°21‘44“E). Jeden výskyt je také v NP Nízké Tatry nedaleko obce Liptovská Teplička (48°58‘2“N, 20°5‘23“E). Izolovaný výskyt je v PR Salvatorské lúky přibližně 25 km západně od Prešova (49°2‘45“N, 20°56‘30“E) (tab. 1, obr. 1).
tab. 1. Charakteristiky studovaných populací a lokalit Ligularia sibirica. Populace jsou seskupeny dle států (CZ a SK) a regionů (N, C, S, SK a P).table 1. Characteristics of the studied populations and localities of Ligularia sibirica. The populations were divided according to states (CZ and SK) and regions (N, C, S, SR and P).
Populace Abrev.velikost
populace (rok 2006)
typ habitatu(Ellenberg –
dostupnost dusíku/produktivita)
Management
Česká republika (cZ)
severní Čechy (N)
Vojenské louky CZ N/1 126 Chudé na dusík/živinyMozaikovité kosení (žádné, červen, září) + odstraňování náletu dřevin
Baronský rybník CZ N/2 243 Chudé na dusík/živiny Kosení (červen) + odstraňování náletu dřevin
Louky u Robeče CZ N/3 193 Bohaté na dusík/živiny Bez managementu
Rákosina u Robeče CZ N/4 323 Bohaté na dusík/živiny Žádný
Sluneční dvůr CZ N/5 123 Bohaté na dusík/živiny
Mozaikovité kosení (žádné, září) + odstraňování náletu dřevin
střední Čechy (c)Rečkov CZ C/1 ~ 10 000 Chudé na dusík/živiny Mozaikovité kosení
(červen, září)
Louky u NV CZ C/2 ~ 10 000 Bohaté na dusík/živiny Bez managementu
Klokočka CZ C/3 ~ 7 000 Chudé na dusík/živiny Kosení – září
Valcha CZ C/4 85 Bohaté na dusík/živiny Bez managementu
jižní Čechy (S)
Olšina CZ S/1 224 Chudé na dusík/živiny Odstranění náletu dřevin
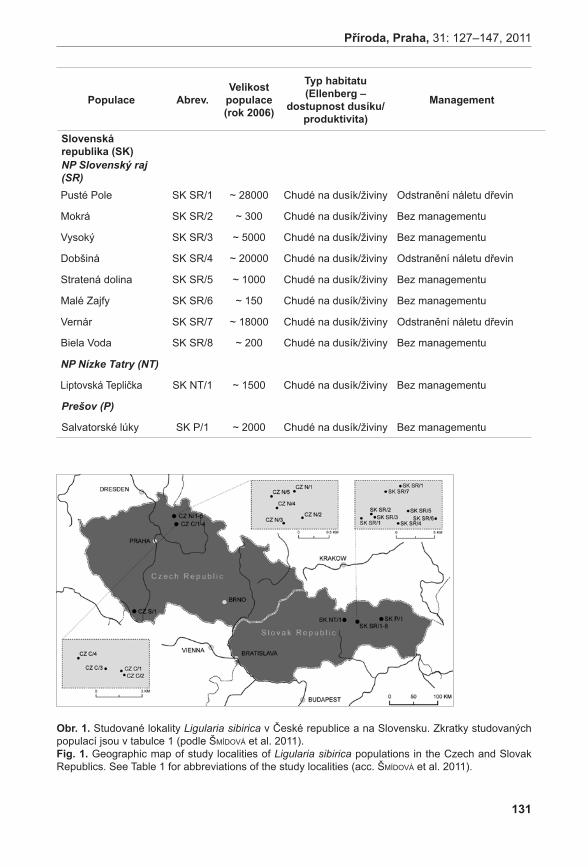
Příroda, Praha, 31: 127–147, 2011
131
Populace Abrev.velikost
populace (rok 2006)
typ habitatu(Ellenberg –
dostupnost dusíku/produktivita)
Management
slovenská republika (sK)NP Slovenský raj (SR)Pusté Pole SK SR/1 ~ 28000 Chudé na dusík/živiny Odstranění náletu dřevin
Mokrá SK SR/2 ~ 300 Chudé na dusík/živiny Bez managementu
Vysoký SK SR/3 ~ 5000 Chudé na dusík/živiny Bez managementu
Dobšiná SK SR/4 ~ 20000 Chudé na dusík/živiny Odstranění náletu dřevin
Stratená dolina SK SR/5 ~ 1000 Chudé na dusík/živiny Bez managementu
Malé Zajfy SK SR/6 ~ 150 Chudé na dusík/živiny Bez managementu
Vernár SK SR/7 ~ 18000 Chudé na dusík/živiny Odstranění náletu dřevin
Biela Voda SK SR/8 ~ 200 Chudé na dusík/živiny Bez managementu
NP Nízke Tatry (NT)
Liptovská Teplička SK NT/1 ~ 1500 Chudé na dusík/živiny Bez managementu
Prešov (P)
Salvatorské lúky SK P/1 ~ 2000 Chudé na dusík/živiny Bez managementu
obr. 1. Studované lokality Ligularia sibirica v České republice a na Slovensku. Zkratky studovaných populací jsou v tabulce 1 (podle ŠMíDová et al. 2011).Fig. 1. Geographic map of study localities of Ligularia sibirica populations in the Czech and Slovak Republics. See Table 1 for abbreviations of the study localities (acc. ŠMíDová et al. 2011).

132
Heinken-Šmídová & Münzbergová: Faktory ovlivňující populační dynamiku Ligularia sibirica
velikost populace
Velikost populace byla stanovena v roce 2006. V případě malých populací (< 500 dospělých jedinců), byl velikostí populace počet všech dospělých jedinců na dané lokalitě. V případě velkých populací (> 500 dospělých jedinců), byla přibližná velikost populace stanovena jako průměrný počet dospělých jedinců na deseti jeden m2 velkých plochách, vynásobeno rozlohou populace. Tyto plochy byly rozmístěny po celé lokalitě s cílem pokrýt rozdílnou hustotu dospělých jedinců.
sběr demografických dat a tvorba přechodových matic
Podrobné studium demografie bylo provedeno v průběhu čtyř let (2005–2008) na devíti populacích v České republice a na dvou populacích na Slovensku. Jedna ze slovenských populací se skládala ze tří dílčích populací. V každé studované populaci jsme označily a sledovaly pokud možno 50 jedinců v každé kategorii. Životní cyklus L. sibirica jsme rozdělily do sedmi kategorií: (1) semenná banka první rok, (2) semenná banka druhý rok, (3) semenáček, (4) juvenilní jedinec, (5) vegetativní dospělý jedinec, (6) kvetoucí dospělý jedinec a (7) dormantní dospělý jedinec (obr. 2).
„Semenná banka“ je tvořena živými semeny, která jsou v dormantním stavu. Tato semena se nacházejí buď na povrchu půdy, nebo přímo pod jejím povrchem.
„Semenáček“ je definován jako nově ze semene vytvořený jedinec, který není starší jednoho roku.
obr. 2. Životní cyklus druhu Ligularia sibirica: (1) semenná banka první rok, (2) semenná banka druhý rok (3), semenáček, (4) juvenilní jedinec (5), vegetativní dospělý jedinec (6) kvetoucí dospělý jedinec a (7) dormantní dospělý jedinec. Hodnoty a44, a55, a66, a77 představují přežívání jedinců v dané kategorii. Hodnoty a21, a31, a32, a43, a53, a54, a65, a75, a76 představují přechod jedinců do další kategorie (růst). Hodnoty a56, a57 představují návrat jedinců do předchozí kategorie (např. kvetoucí dospělý jedinec přejde do kategorie vegetativní dospělý jedinec). Hodnoty a16, a36 představují reprodukci (heinken-ŠMíDová & MünzberGová, v tisku).Fig. 2. Life-cycle diagram of the seven stages of Ligularia sibirica: (1) seed bank first year, (2) seed bank second year, (3) seedlings, (4) juveniles, (5) vegetative adults, (6) fertile adults, and (7) dormant adult individuals. Values a44, a55, a66, a77 represent survival of individuals in the stage. Values a21, a31, a32, a43, a53, a54, a65, a75, a76 represent transition of individuals into next stages (growth). Values a56, a57 represent retrogression of individuals. Values a16, a36 represent fecundity (heinken-ŠMíDová & MünzberGová, in press).

Příroda, Praha, 31: 127–147, 2011
133
„Juvenilní jedinec“ je jedinec velikosti semenáčku (mající max. dva listy o šířce do dvou cm) starší více než jeden rok. V kategorii juvenilní jedinec mohou jedinci zůstat až několik let. Větší jedinci byli umístěni do kategorie vegetativní dospělý jedinec.
„Vegetativní dospělý jedinec“ je nekvetoucí jedinec s 1–15 růžicemi v hustém trsu, které mohou být spojeny až deset cm dlouhými podzemní oddenky. U vegetativních dospělých jedinců jsme také zaznamenávaly počet listových růžic a počet lisů.
„Kvetoucí dospělý jedinec“ je jedinec tvořící kvetoucí lodyhy. U kvetoucích dospělých jedinců jsme dále zaznamenávaly počet listových růžic, počet lisů, počet kvetoucích lodyh a počet květů v květenství. Na podzim po dozrání semen jsme na vzorku 20 jedinců na populaci počítaly průměrnou produkci semen.
„Dormantní dospělý jedinec“ je jedinec, který přežívá pouze v podobě podzemních orgánů po dobu jednoho až dvou let. Jedinci, kteří nevytvořili nadzemní vegetační orgány po více než dva roky, byli považováni za mrtvé.
Demografická data byla zpracována pomocí populačních projekčních matic (caswell 2001). Naším cílem bylo stanovit populační rychlost růstu (λ) a elasticitu (hodnota elasticity ukazuje, které přechody nejvíce ovlivňují růstovou rychlost populace; De kroon et al. 2000) jednak spolu s 95 % intervalem spolehlivosti pro každou populaci a přechodový interval zvlášť, tak i pomocí stochastické simulace dohromady přes všechny populace a roky sledování. Dále jsme pomocí metody cochran & ellner (1992) vypočítaly průměrnou předpokládanou délku života L. sibirica a pravděpodobnost zániku populací. Všechny analýzy byly provedeny pomocí programu Matlab verze 7.3.0.267 (Mathworks inc. 2006). Detailní podrobnosti o konstrukci populačních projekčních matic a následných analýz popisují heinken-ŠMíDová & MünzberGová (v tisku).
Genetická variabilita
Genetická variabilita byla stanovena pomocí metody isoenzymů pro 20 populací L. sibirica v České a Slovenské republice (obr. 1). Z každé populace tohoto druhu byly z 20 náhodně vybraných dospělých jedinců odebrány vzorky listové plochy. Tyto vzorky byly po dobu převozu do laboratoře chlazeny. V laboratoři byly vzorky nejprve extrahovány a posléze analyzovány. Detailní postup elektroforézy je popsán v článku ŠMíDová et al. (2011). Vyhodnoceno bylo devět polymorfních lokusů z pěti enzymatických systémů: aspartate aminotransferase (AAT, E.C. 2.6.1.1), diaphorase (DIA, E.C. 1.6.99.3), leucyl aminopeptidase (LAP, E.C. 3.4.11.1), superoxide dismutase (SOD, E.C. 1.15.1.1) a 6-phosphogluconate dehydrogenase (6-PGDH, E.C. 1.1.1.44).
Statistické zpracování bylo provedeno v programu Arlequin (Arlequin 3.11; eXcoffier & schneiDer 2005) a Popgene (verze 1.32; yeh & boyle 1997). Zjišťovaly jsme základní parametry genetické variability tj. průměrný počet alel na lokus (A) a na polymorfní lokus (AP), procento polymorfních lokusů (P) a celkový počet alel (TA), průměrnou pozorovanou (HO) a očekávanou (HE) heterozygotnost. Pomocí AMOVA jsme stanovily rozložení genetické variability uvnitř a mezi populacemi a mezi regiony (Česká a Slovenská republika). Dále jsme vypočítaly index genetické diferenciace (FST) neboli genetické vzdálenosti, který se často používá pro porovnávání genetické rozdílnosti mezi populacemi. Pomocí Mantlova testu jsme následně testovaly vztah mezi geografickou vzdáleností a jí odpovídající genetickou vzdáleností (FST) mezi dvojicemi populací. Taktéž jsme pomocí lineární regrese testovaly vztah mezi velikostí populací a genetickou variabilitou (s-Plus 2000).

134
Heinken-Šmídová & Münzbergová: Faktory ovlivňující populační dynamiku Ligularia sibirica
stanovištní charakteristiky
Cílem této podkapitoly bylo popsat stanovištní podmínky, ve kterých L. sibirica roste. Pro popis stanovištních podmínek jsme vybraly několik faktorů, které mají pravděpodobně vliv na růst a distribuci studovaného druhu, tj. vegetační složení, chemické složení půdy a listové biomasy a management stanovišť studovaného druhu.
Vegetační složení je reprezentováno fytocenologickými snímky. Každému druhu cévnatých rostlin a mechorostů byly v každém fytocenologickém snímku přiděleny známé Ellenbergovy indikační hodnoty pro světlo, vlhkost, půdní reakci a dostupnost dusíku (ellenberG et al. 1992). Ellenbergova indikační hodnota dostupnosti dusíku může být také interpretována jako obecné zásobení živinami (ellenberG et al. 1992) neboli hodnota produktivity (schaffers & sýkora 2000). Z těchto hodnot jsme následně vypočítaly průměr. Ellenbergovy indikační hodnoty jsme k popisu stanoviště použily, protože odrážejí ekologické chování druhů. A proto jejich průměrné hodnoty nepopisují stanovištní podmínky v daném okamžiku, ale vzájemné působení stanovištních podmínek v průběhu času (ellenberG et al. 1992). Podobnost jednotlivých stanovišť na základě jejich druhového složení byla analyzovaná pomocí detrendované korespondenční analýzy (DCA) fytocenologických snímků provedené v programu Canoco 4.5 (ter braak & ŠMilauer 2002). Pro popsání gradientu druhového složení jsme také otestovaly vztah mezi skórami lokalit na první DCA ose a Ellenbergovými indikačními hodnotami pro světlo, vlhkost, půdní reakci, dostupnost dusíku (produktivita).
K chemickému rozboru půdy byl použit směsný půdní vzorek vytvořený z deseti odběrů z hloubky 20 cm v rámci každé lokality. Sledováno bylo % organického dusíku, % organického uhlíku, obsah vápníku (Ca2+), draslíku (K+), hořčíku (Mg2+), fosfátů (P2O5) a pH. Vlastní chemická analýza byla provedena dle standardní metody (králová 1990, zbíral 1995). K chemickému rozboru biomasy byl použit směsný vzorek z listové plochy 20 sterilních jedinců na lokalitu. Sledováno bylo % organického dusíku, % organického uhlíku, obsah vápníku (Ca2+), draslíku (K+), hořčíku (Mg2+) a fosfátů (P2O5) dle standardní metody (zbíral 1995).
V rámci hodnocení stanovištních nároků druhu byly provedeny dva typy analýz. První z nich byla provedena pro všechny studované populace druhu, tj. 20 populací. Pomocí korelační analýzy jsme hodnotily obsah prvků v půdě a listové biomase. Druhá analýza byla provedena pouze pro část populací, a sice pro ty, u kterých bylo provedeno studium populační dynamiky L. sibirica. U dospělých jedinců těchto populací byly měřeny vybrané parametry, konkrétně počet listových růžic, počet kvetoucích lodyh, počet květů v květenství a produkce semen na květ. Pomocí regresních analýz jsme zjistily vliv všech výše uvedených charakteristik stanovišť (tj. první DCA osa, Ellenbergovy indikační hodnoty, obsah prvků v půdě a listové biomase) na měřené morfologické parametry a velikost populace. Všechny statistické analýzy byly provedeny v programu s-Plus (2000).
vliv kvality stanoviště, velikosti populace a genetické variability na vitalitu populací L. sibirica
Vliv kvality stanoviště, velikosti populace a genetické variability byl zjišťován pomocí regresních analýz. Studovaly jsme celkový a čistý vliv faktorů. Celkový vliv studovaných faktorů je vliv jednoho faktoru, aniž by byly vlivy dalších dvou faktorů zahrnuty do analýzy. Čistý vliv je pak vliv každého faktoru za použití ostatních dvou jako kovariát. Tímto způsobem jsme zjistily vliv daného faktoru, aniž by byl smíchaný s vlivy ostatních faktorů.
Genetickou variabilitu reprezentovaly hodnoty pozorované heterozygotnosti (HO). Kvalitu stanoviště reprezentovaly skóry vegetačních snímků na první DCA ose popsané výše. Statistické analýzy byly provedeny v s-Plus (2000).
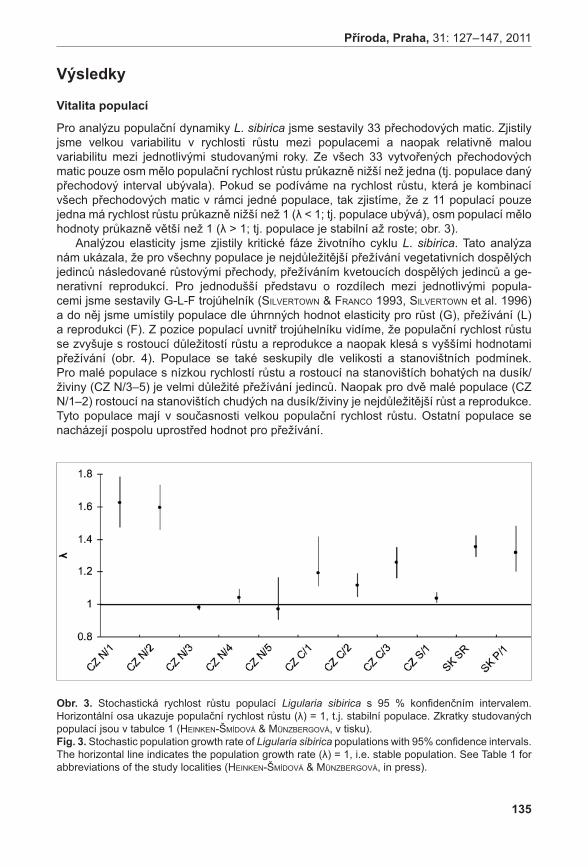
Příroda, Praha, 31: 127–147, 2011
135
výsledky
vitalita populací
Pro analýzu populační dynamiky L. sibirica jsme sestavily 33 přechodových matic. Zjistily jsme velkou variabilitu v rychlosti růstu mezi populacemi a naopak relativně malou variabilitu mezi jednotlivými studovanými roky. Ze všech 33 vytvořených přechodových matic pouze osm mělo populační rychlost růstu průkazně nižší než jedna (tj. populace daný přechodový interval ubývala). Pokud se podíváme na rychlost růstu, která je kombinací všech přechodových matic v rámci jedné populace, tak zjistíme, že z 11 populací pouze jedna má rychlost růstu průkazně nižší než 1 (λ < 1; tj. populace ubývá), osm populací mělo hodnoty průkazně větší než 1 (λ > 1; tj. populace je stabilní až roste; obr. 3).
Analýzou elasticity jsme zjistily kritické fáze životního cyklu L. sibirica. Tato analýza nám ukázala, že pro všechny populace je nejdůležitější přežívání vegetativních dospělých jedinců následované růstovými přechody, přežíváním kvetoucích dospělých jedinců a ge-ne rativní reprodukcí. Pro jednodušší představu o rozdílech mezi jednotlivými popu la-cemi jsme sestavily G-L-F trojúhelník (silvertown & franco 1993, silvertown et al. 1996) a do něj jsme umístily populace dle úhrnných hodnot elasticity pro růst (G), přežívání (L) a reprodukci (F). Z pozice populací uvnitř trojúhelníku vidíme, že populační rychlost růstu se zvyšuje s rostoucí důležitostí růstu a reprodukce a naopak klesá s vyššími hodnotami přežívání (obr. 4). Populace se také seskupily dle velikosti a stanovištních podmínek. Pro malé populace s nízkou rychlostí růstu a rostoucí na stanovištích bohatých na dusík/živiny (CZ N/3–5) je velmi důležité přežívání jedinců. Naopak pro dvě malé populace (CZ N/1–2) rostoucí na stanovištích chudých na dusík/živiny je nejdůležitější růst a reprodukce. Tyto populace mají v současnosti velkou populační rychlost růstu. Ostatní populace se nacházejí pospolu uprostřed hodnot pro přežívání.
obr. 3. Stochastická rychlost růstu populací Ligularia sibirica s 95 % konfidenčním intervalem. Horizontální osa ukazuje populační rychlost růstu (λ) = 1, t.j. stabilní populace. Zkratky studovaných populací jsou v tabulce 1 (heinken-ŠMíDová & MünzberGová, v tisku).Fig. 3. Stochastic population growth rate of Ligularia sibirica populations with 95% confidence intervals. The horizontal line indicates the population growth rate (λ) = 1, i.e. stable population. See Table 1 for abbreviations of the study localities (heinken-ŠMíDová & MünzberGová, in press).
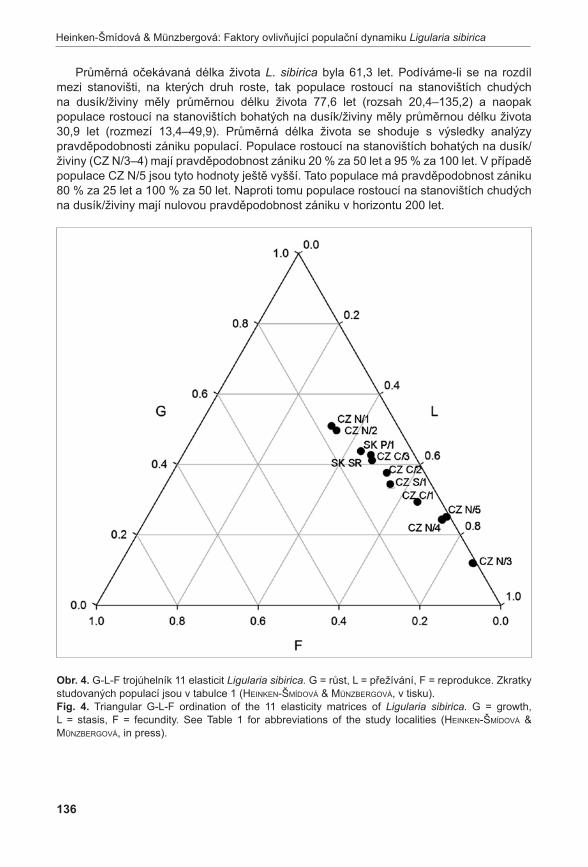
136
Heinken-Šmídová & Münzbergová: Faktory ovlivňující populační dynamiku Ligularia sibirica
Průměrná očekávaná délka života L. sibirica byla 61,3 let. Podíváme-li se na rozdíl mezi stanovišti, na kterých druh roste, tak populace rostoucí na stanovištích chudých na dusík/živiny měly průměrnou délku života 77,6 let (rozsah 20,4–135,2) a naopak populace rostoucí na stanovištích bohatých na dusík/živiny měly průměrnou délku života 30,9 let (rozmezí 13,4–49,9). Průměrná délka života se shoduje s výsledky analýzy pravděpodobnosti zániku populací. Populace rostoucí na stanovištích bohatých na dusík/živiny (CZ N/3–4) mají pravděpodobnost zániku 20 % za 50 let a 95 % za 100 let. V případě populace CZ N/5 jsou tyto hodnoty ještě vyšší. Tato populace má pravděpodobnost zániku 80 % za 25 let a 100 % za 50 let. Naproti tomu populace rostoucí na stanovištích chudých na dusík/živiny mají nulovou pravděpodobnost zániku v horizontu 200 let.
obr. 4. G-L-F trojúhelník 11 elasticit Ligularia sibirica. G = růst, L = přežívání, F = reprodukce. Zkratky studovaných populací jsou v tabulce 1 (heinken-ŠMíDová & MünzberGová, v tisku).Fig. 4. Triangular G-L-F ordination of the 11 elasticity matrices of Ligularia sibirica. G = growth, L = stasis, F = fecundity. See Table 1 for abbreviations of the study localities (heinken-ŠMíDová & MünzberGová, in press).

Příroda, Praha, 31: 127–147, 2011
137
Genetická variabilita
Devět z 11 lokusů (81,81 %) bylo polymorfních alespoň v jedné populaci. Celkově bylo v polymorfních lokusech zjištěno 32 různých alel. Průměrný počet alel na lokus (A) byl 2,25 (rozsah: 2,00–2,55), průměrný počet alel na polymorfní lokus (AP) byl 2,93 (rozsah: 2,43–3,17), procento polymorfních lokusů (P) bylo 65,91 % (rozmezí: 54,55–72,73 %) a celkový počet alel (TA) byl 24,55 v rozmezí 21 až 28. Průměrná očekávaná heterozygotnost populace (HE) byla 0,36 v rozmezí 0,24–0,37 a průměrná pozorovaná heterozygotnost (HO) byla 0,33 a pohybovala se v rozmezí 0,26–0,42.
Analýza molekulární variance (AMOVA) ukázala, že 80,81 % molekulární variance se nachází uvnitř populací, 12,65 % mezi populacemi a 6,54 % mezi Českou a Slovenskou republikou. Index genetické diferenciace (FST) byl 0,18.
Genetická vzdálenost byla statisticky průkazně korelována s geografickou vzdáleností (Mantel test; r = 0,40; P < 0,001). Tato korelace byla také průkazná zvlášť v rámci České republiky (Mantel test; r = 0,45; P = 0,002) a Slovenské republiky (Mantel test; r = 0,49; P < 0,001).
Zjistily jsme také průkaznou pozitivní korelaci mezi logaritmem velikosti populace a očekávanou heterozygotností (r = 0,56; P = 0,01), která se s velikostí populace zvyšuje. Podobné výsledky byly zjištěny i pro další parametry genetické variability, tj. průměrný počet alel na lokus (A; r = 0,63; P = 0,003), celkový počet alel (TA; r = 0,60; P = 0,005) a průměrná pozorovaná heterozygotnost (HO; r = 0,52; P = 0,02).
stanovištní charakteristiky
Ligularia sibirica roste v relativně široké škále společenstev. Většina slovenských populací roste na vápnitých slatinách ve společenstvu svazu Caricion davallianae a částečně také ve svazu Alno-Ulmion. V České republice se pak většinou jedná o společenstvo na přechodu mezi svazem Caricion davallianae a Molinion caeruleae. Některé stanoviště však nelze do této klasifikace zařadit. To je například lokalita CZ C/2 v údolní nivě říčky Rokytky ve středních Čechách, kterou lze klasifikovat jako živinami bohatý ostřicový porost náležející do svazu Magnocaricion elatae. Druhou výjimkou je lokalita v Pošumaví v jižních Čechách (CZ S/1), která je podobná slatinám chudým na živiny, ale narozdíl od nich není bohatá na vápník a je kyselá. Tuto lokalitu lze klasifikovat jako přechodové rašeliniště mozaiky společenstev svazů Caricion fuscae, Molinion caeruleae a Calthion palustris. Poslední výjimkou jsou lokality v severních Čechách (CZ N/3–5). Zde došlo v minulosti k opakovanému odvodnění slatin (henDrych 2003). Dnes lze tyto stanoviště jen obtížně fytocenologicky klasifikovat. Daly by se popsat jako netypická, na dusík/živiny bohatá Molinietalia s dominancí vysokých rostlin, například Molinia caerulea a Phragmites australis (A. Heinken-Šmídová et al., nepublikovaná data).
DCA analýza vegetačního složení na lokalitách ukázala, že hlavní gradient v datech je gradient od vysokých a povětšinou expanzivních druhů rostoucí na dusíkem/živinami bohatých stanovištích jako jsou například Calamagrostis epigeios, Carex acutiformis, Cirsium oleraceum, Eupatorium cannabinum a Phragmites australis po druhy vápnitých slatin jako jsou Carex davalliana, Parnassia palustris, Crepis paludosa a Succisa pratensis. Polohu jednotlivých lokalit na tomto gradientu dále považujeme za kvalitu stanoviště od lokalit degradovaných, a tedy ochranářsky bezcenných, po lokality zachovalé, a tedy ochranářsky cenné.
Rozdíly ve složení vegetace mezi jednotlivými lokalitami byly průkazně vysvětleny Ellenbergovými indikačními hodnotami pro dostupnost dusíku (produktivita) (r = -0,94; P < 0,001) a světla (r = 0,86, P < 0,001). Tyto dvě Ellenbergovy hodnoty byly navzájem

138
Heinken-Šmídová & Münzbergová: Faktory ovlivňující populační dynamiku Ligularia sibirica
silně korelovány. Kvalita stanoviště je tedy gradient od nízké kvality (stanoviště bohaté na dusík/živiny s dominancí vysokých druhů) po vysoce kvalitní (stanoviště chudé na dusík/živiny s dominancí druhů vápnitých slatin).
Chemické složení půdy ukazuje na vysoký obsah vápníku (Ca2+) a hořčíku (Mg2+) na lokalitách nacházejících se především v NP Slovenský raj, což je dáno především vápencovým a částečně dolomitickým podložím oblasti. Obsah ostatních prvků v půdě je širokého rozsahu. Pomocí korelační analýzy obsahu prvků v půdě a listové biomase bylo prokázáno, že se zvyšujícím se poměrem C/N v půdě se zvyšuje poměr C/N listové biomase. Podobně se zvyšujícím se obsahem hořčíku (Mg2+) a fosfátů (P2O5) v půdě se zvyšuje jejich koncentrace v listové biomase. Pouze v případě obsahu vápníku (Ca2+) je trend opačný, tedy se zvyšujícím se obsahem prvku v půdě se snižuje jeho obsah v listové biomase.
Vybrané morfologické parametry a velikost populace byly průkazně ovlivněny alespoň jednou ze stanovištních charakteristik. Počet listových růžic, jak vegetativních, tak i kvetoucích jedinců se průkazně snižuje se zvyšující se ochranářskou kvalitou stanoviště (skóry první DCA osy), Ellenbergovou indikační hodnotou pro světlo, C/N poměrem v půdě a hodnotou pH. Počet listových růžic se zvyšuje s rostoucím obsahem vápníku (Ca) v listové biomase. Dále se také počet listových růžic (pouze u vegetativních jedinců) výrazně zvyšuje s rostoucí Ellenbergovou indikační hodnotou pro dostupnost dusíku (produktivitou), obsah draslíku (K) v půdě, obsah dusíku (N) a uhlíku (C) v listové biomase. Počet listových růžic se pouze v případě kvetoucích jedinců průkazně snižuje se zvyšujícím se obsahem vápníku (Ca) a uhlíku (C) v půdě. Počet kvetoucích stvolů se průkazně snižuje se zvyšujícím se C/N poměrem v půdě a hodnotami pH. Počet květů v květenství se průkazně zvyšuje s rostoucí Ellenbergovou indikační hodnotou pro dostupnost dusíku (produktivitou) a naopak se snižuje s rostoucím obsahem vápníku (Ca) a uhlíku (C) v půdě. Počet semen na květ se průkazně zvyšuje s rostoucí Ellenbergovou indikační hodnotou pro dostupnost dusíku (produktivitou) a obsahem dusíku v listové biomase. Naopak průkazně klesá se zvyšující se kvalitou prostředí, Ellenbergovou indikační hodnotou pro světlo, C/N poměrem v půdě a listové biomase a hodnotou pH.
Výsledky regresní analýzy velikosti populace a charakteristik stanovišť se v porovnání s morfologickými parametry lišily. Velikost populace se průkazně snížila se zvyšujícím se obsahem draslíku (K) v půdě, dusíku (N) v půdě a v listové biomase. Naopak velikost populace se zvyšuje s rostoucím C/N poměrem v půdě a listové biomase, hodnotou pH a obsahem fosforu (P) v listové biomase.
vliv kvality stanoviště, velikosti populace a genetické variability na vitalitu populací L. sibirica
Výsledky regresních analýz ukázaly, že stochastická rychlost růstu populací průkazně roste s rostoucí kvalitou stanoviště (obr. 5) v případě celkového i čistého vlivu a je nezávislá na velikosti populace a genetické variabilitě. Elasticity byly nezávislé, jak na velikosti populace, tak i na genetické variabilitě. Průkazně však byly ovlivněny kvalitou stanoviště. Konkrétně, elasticity růstu a reprodukce byly vyšší na stanovištích s nízkou kvalitou stanoviště. A naopak elasticity přežívání byly na tomto typu stanoviště vyšší. Vliv kvality stanoviště byl průkazný pouze v případě celkového vlivu. Průměrná očekávaná délka života L. sibirica se průkazně zvyšuje s rostoucí kvalitou stanoviště a s rostoucí velikostí populace. Nicméně, v případě čistého vlivu, byl průkazný pouze vliv kvality stanoviště (A. Heinken-Šmídová a Z. Münzbergová, nepublikovaná data).
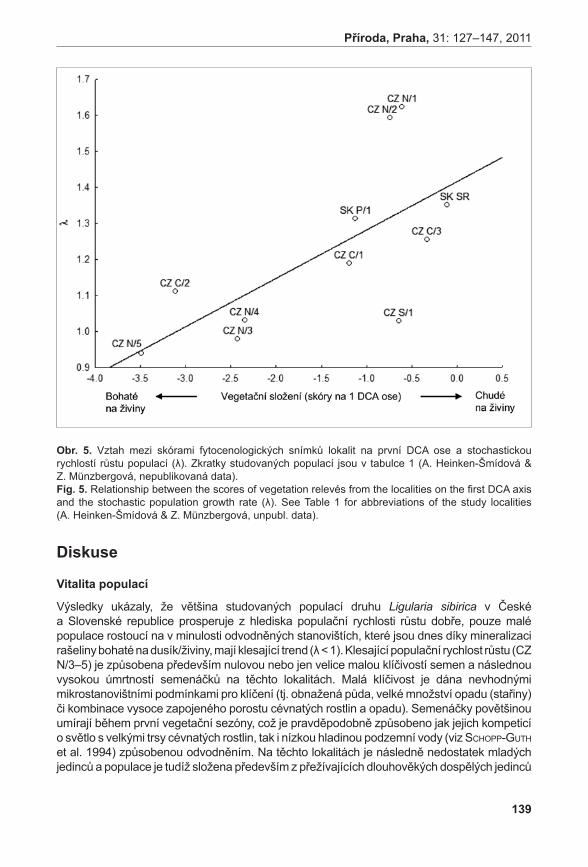
Příroda, Praha, 31: 127–147, 2011
139
obr. 5. Vztah mezi skórami fytocenologických snímků lokalit na první DCA ose a stochastickou rychlostí růstu populací (λ). Zkratky studovaných populací jsou v tabulce 1 (A. Heinken-Šmídová & Z. Münzbergová, nepublikovaná data).Fig. 5. Relationship between the scores of vegetation relevés from the localities on the first DCA axis and the stochastic population growth rate (λ). See Table 1 for abbreviations of the study localities (A. Heinken-Šmídová & Z. Münzbergová, unpubl. data).
diskuse
vitalita populací
Výsledky ukázaly, že většina studovaných populací druhu Ligularia sibirica v České a Slovenské republice prosperuje z hlediska populační rychlosti růstu dobře, pouze malé populace rostoucí na v minulosti odvodněných stanovištích, které jsou dnes díky mineralizaci rašeliny bohaté na dusík/živiny, mají klesající trend (λ < 1). Klesající populační rychlost růstu (CZ N/3–5) je způsobena především nulovou nebo jen velice malou klíčivostí semen a následnou vysokou úmrtností semenáčků na těchto lokalitách. Malá klíčivost je dána nevhodnými mikrostanovištními podmínkami pro klíčení (tj. obnažená půda, velké množství opadu (stařiny) či kombinace vysoce zapojeného porostu cévnatých rostlin a opadu). Semenáčky povětšinou umírají během první vegetační sezóny, což je pravděpodobně způsobeno jak jejich kompeticí o světlo s velkými trsy cévnatých rostlin, tak i nízkou hladinou podzemní vody (viz schoPP-Guth et al. 1994) způsobenou odvodněním. Na těchto lokalitách je následně nedostatek mladých jedinců a populace je tudíž složena především z přežívajících dlouhověkých dospělých jedinců

140
Heinken-Šmídová & Münzbergová: Faktory ovlivňující populační dynamiku Ligularia sibirica
(stárnoucí populace). Protikladem těchto populací jsou dvě malé, v současnosti expandující populace rostoucí na stanovišti chudém na dusík/živiny (CZ N/1–2). Tyto populace mají velké množství semenáčků díky dobře vyvinutému mechovému patru, které velice dobře udržuje mikroklimatické podmínky vhodné pro klíčení (overbeck et al. 2003).
Výsledky dále ukazují, že přežívání semen v semenné bance přispívá jen velice málo k variabilitě v rychlosti růstu populací. Semenná banka L. sibirica je v porovnání s produkcí semen velice malá. Dle práce thoMPson & GriMe (1979) se jedná o přechodnou semennou banku II. typu. Na stanovištích chudých na dusík/živiny buď většina semen vyklíčí, nebo se rozloží následující rok po svém vzniku, což vede k tomu, že se v semenné bance nenacházejí téměř žádná semena. Naproti tomu na stanovištích bohatých na dusík/živiny jsou semena v semenné bance schopna přežít po dva následující roky, což je dáno nízkou hladinou pozemní vody na těchto lokalitách. Příspěvek semenné banky k rychlosti růstu těchto populací je však velice nízký kvůli nízké klíčivosti semen.
Zatímco analýza elasticity jasně ukázala rozdíly mezi jednotlivými typy stanovišť (stanoviště chudé a bohaté na dusík/živiny), rozdíly mezi velikostmi populací byly nezřetelné. Vysoká hodnota elasticity naznačuje, že malá změna v těchto přechodech bude mít velmi výrazný vliv na rychlost růstu dané populace. U populací rostoucích na stanovištích chudých na dusík/živiny měly největší elasticitu: růst semenáčků, přežívání a růst juvenilních a vegetativních rostlin a také reprodukce. Populace rostoucí na stanovištích bohatých na dusík/živiny měly největší elasticitu v přežívání vegetativních a kvetoucích dospělých jedinců. Velký význam v přežívání dospělých jedinců byl již dříve zjištěn u řady vytrvalých druhů mokřadních rostlin, jako například v případě Gentiana pneumonanthe a Scorzonera humilis (oosterMeijer et al. 1996, collinG & Matthies 2006). Tento jev lze v případě těchto rostlin a také v případě L. sibirica vysvětlit schopností ukládat zásoby živin v krátkém oddenku, což může pomáhat rostlině překonat nepříznivé změny prostředí. Význam dormance dospělých rostlin mírně vzrostla u populací, které rostou na stanovištích bohatých na dusík/živiny. Dormance dospělých rostlin je známa především u zástupců čeledi Orchidaceae (viz shefferson 2009), a jen zřídka je kategorie dormance dospělých jedinců součástí studií populační dynamiky rostlin (např. ehrlén 1995, oosterMeijer et al. 1996). Stejně jako u jiných druhů, může být dormance dospělých rostlin L. sibirica reakcí na nepříznivé podmínky životního prostředí (Storage effect; hiGGins et al. 2000).
V případě populací rostoucích na stanovištích bohatých na dusík/živiny nám analýza elasticity identifikovala přežívání jako nejdůležitější přechod pro populační růst L. sibirica. Toto zjištění nám říká, že nejlepším managementovým zásahem pro podpoření populačního růstu tohoto typu populací, je takové opatření, které zvýší přežívání jedinců. Tento výsledek je však zavádějící, neboť, jak jsme se již dříve zmínily, na těchto lokalitách chybí semenáčci. Podporou přežívání dospělých jedinců nezískáme životaschopné populace, ale pouze oddálíme jejich zánik.
Pravděpodobnost zániku malých i velkých populací rostoucích na stanovištích chudých na dusík/živiny a i jedné velké populace rostoucí v prostředí bohatém na živiny (CZ C/2), byla nulová (počítáno pro 200 let). Tento výsledek může být jednak díky dlouhé délce života, tak i pomalému růstu a stabilitě těchto populací, což jsou vlastnosti typické pro mnoho druhů rostoucích na stanovištích s nepříznivými podmínkami pro růst rostlin (eriksson 1996). Malé populace rostoucí na stanovištích bohatých na dusík/živiny (CZ N/3–5) mají vysokou předpokládanou pravděpodobnost zániku. Tyto tři malé populace rostou po více než 50 let ve stále se zhoršujících podmínkách prostředí v důsledku opakovaných meliorací (henDrych 2003). Očekávaná délka života jednotlivců je proto velmi nízká.
Z výše uvedeného jasně vyplývá, že poslední tři zmíněné populace L. sibirica (CZN/3–5) vykazují charakteristiky zbytkových populací. Takovéto populace jsou schopny

Příroda, Praha, 31: 127–147, 2011
141
přežívat delší dobu v nepříznivých podmínkách prostředí (eriksson 1996, eriksson 2000) a to téměř bez reprodukce díky dlouhé životnosti a nízké úmrtnosti dospělých jedinců (oosterMeijer et al. 1994, collinG & Matthies 2006). V případě L. sibirica navíc také díky dormanci dospělých jedinců. Tyto vlastnosti druhu bohužel vedou k velkému časovému zpoždění mezi zhoršením stanovištních podmínek a poklesem velikosti populace (collinG & Matthies 2006).
Genetická variabilita
Hlavními vlastnostmi rostlin, které ovlivňují množství genetické variability, je životní forma druhu, geografické rozšíření a typ reprodukce. Vytrvalé, široce rozšířené a cizosprašné druhy mají obecně větší genetickou variabilitu než druhy krátkověké, samosprašné nebo druhy se smíšeným typem reprodukce a lokálním rozšířením (např. endemity; haMrick & GoDt 1989). Ligularia sibirica je vytrvalá rostlina se smíšeným typem reprodukce, v Evropě klasifikována jako relikt (henDrych 2003). S touto kombinací vlastností by genetická variabilita studovaného druhu na úrovni isozymů měla být někde mezi nízkou a vysokou hodnotou (haMrick & GoDt 1989). Počet alel na lokus (A) zjištěný u druhu L. sibirica(A = 2,25) je blízký hodnotám pozorovaným u endemických druhů (průměrný počet alel na lokus (A) = 3,00). Procento polymorfních lokusů a průměrná očekávaná heterozygosita však ukazují spíše na podobnost se široce rozšířenými druhy (procento polymorfních lokusů (P) = 59 %, L. sibirica: 65,91 % a průměrná očekávaná heterozygosita (HE) = 0,20, L. sibirica: 0,36). Podobný typ genetické variability byl nalezen i u dalších vzácných druhů rostlin např. Biscutella laevigata a Saxifraga aizoides (např. DanneMann 2000, lutz et al. 2000).
Analýza molekulární variance odhalila, že většina genetické variability se nachází uvnitř populací L. sibirica (80,81 %). Také studie jiných disjunktně rozšířených taxonů ukázaly podobně vysokou úroveň variability uvnitř populací (DanneMann 2000, lutz et al. 2000).
Velká genetická variabilita uvnitř populací spolu se střední genetickou diferenciací může naznačovat velký genový tok, nebo také může být výsledkem událostí probíhajících v minulosti (van rossuM & Prentice 2004). Jednou z hypotéz v případě L. sibirica může být, že by rozložení genetické variability mezi populacemi mohlo být odrazem genového toku z minulosti, tedy předtím než došlo ke fragmentaci v minulosti větších populací (hensen et al. 2005, lowe et al. 2004, Peterson et al. 2008). Druhou hypotézu, kterou je třeba zvážit, je, že současný tok genů spojuje účinněji populace, které jsou si geograficky blíže, než populace, které jsou vzdálenější (hensen et al. 2005, Peterson et al. 2008). Tato možnost je pravděpodobnější pro severní a střední Čechy. My předpokládáme, že obě výše uvedené hypotézy mohou být v případě L. sibirica pravdivé. Podobné výsledky byly zjištěny pro glaciální relikty (např. DanneMann 2000, lutz et al. 2000, reisch et al. 2003), tedy druhy, které byly v minulosti více rozšířené a jsou schopny si udržet vysokou genetickou variabilitu i přes ca 12 000 let izolace.
Podobně jako glaciální relikty byla pravděpodobně L. sibirica v minulosti více rozšířená a její populace mohly být pravděpodobně přirozeně fragmentovány přibližně po dobu 7 000–10 000 let. Nicméně v literatuře můžeme najít několik teorií o původu L. sibirica ve střední a západní Evropě. Jedna, v současnosti však již překonaná myšlenka, je dovoz semen v seně určeném pro koně Kozáků během napoleonských válek z Ruska (kneblová 1950). Další teorií je šíření semen na velké vzdálenosti prostřednictvím migrujících nebo přezimujících ptáků (walter & straka 1970, MarŠáková-něMejČková 1973). Takovéto vzácné případy by však musely být několikrát opakovány a několik efektů zakladatele (founder effect) by vedlo ke ztrátě alel a k homozygotnosti (hewitt 1999). Naše výsledky jsou tedy v rozporu s touto hypotézou. Běžně přijímané vysvětlení je migrace L. sibirica

142
Heinken-Šmídová & Münzbergová: Faktory ovlivňující populační dynamiku Ligularia sibirica
někdy během doby ledové (např. Mattauch 1936, kneblová 1950) nebo v pozdním glaciálu až preboreálu (např. Meusel & jäGer 1992, henDrych 2003). Názor na časové období migrace L. sibirica se neustále mění dle měnící se představy o průběhu doby ledové a jejím rozsahu (henDrych 2003). Ligularia sibirica pravděpodobně přišla do Evropy ze severovýchodu v době, kdy klimatické podmínky a vegetace v Evropě byly podobné podmínkám v současném centru rozdělení druhu, tedy na jihu centrální Sibiře (henDrych 2003); tj. vysoká vlhkost a teplota v létě, nízké teploty v zimě (Meusel & jäGer 1992). Pylově je L. sibirica doložena ze severních Čech z období starého holocénu (Svitavská et al, nepublikovaná data).
Vysoká úroveň genetické variability a střední genetická diferenciace pozorovaná u L. sibirica je charakteristická pro pomalou jednosměrnou postupnou migraci (izolace vzdáleností; hewitt 1999, GauDeul et al. 2000). Bohužel nemáme zcela jasnou představu o tom, jak byla L. sibirica v té době v Evropě rozšířena, určitě však byla přítomna rozsáhleji než dnes. Následkem čehož je většina současných populací pozůstatkem populacích větších, které přežívaly v klimaticky vhodných podmínkách (henDrych 2003). Tato hypotéza je také podporována pozitivním vztahem mezi genetickou a geografickou vzdáleností.
Z výše uvedeného vyplývá, že L. sibirica je druh, který je schopen udržovat dobrou úroveň genetické variability po delší časové období. V této studii jsme také nalezly průkazný vliv velikosti populace a genetické variability, což spíše ukazuje na charakteristiky typické pro druhy, u kterých došlo k výrazné redukci velikosti populace teprve nedávno. Naše zjištění je v souladu s hypotézou, že malé populace nejsou schopny udržet genetickou variabilitu tak vysokou jako populace velké (frankhaM et al. 2006). Tento jev lze obvykle vysvětlit genetickým driftem, buď náhodnou ztrátou alel, efektem zakladatele při kolonizaci, a nebo inbreedingem (soulé 1987). Genetický drift, konkrétně náhodná ztráta alel, může platit pro některé z malých populací L. sibirica v severních Čechách. U těchto populací došlo relativně nedávno ke zmenšení velikostí populací v důsledku zhoršení stanovištních podmínek, jako jsou meliorace a fragmentace slatinišť (henDrych 2003) a následná mineralizace povrchové vrstvy rašeliny.
stanovištní charakteristiky
Již výše, v kapitole Vitalita populací, výsledky ukázaly, že nejlépe prospívají populace rostoucí na zachovalých, na dusík (produkčně) chudých stanovištích společenstva svazu Caricion davallianae a populace rostoucí ve společenstvech klasifikovaných jako přechod mezi Caricion davallianae a Molinion caeruleae. Jedná se o společenstva, pro která je charakteristická vysoká hladina spodní nebo proudící vody, vysoký obsah uhličitanu vápenatého, nedostatek fosforu, neutrální až alkalické pH a nízká produkce biomasy (vysoký C/N poměr v půdě; rozbrojová & hájek 2008), což plně koresponduje s našimi výsledky. Nízká produkce biomasy slatinné vegetace je na českých lokalitách udržována pomocí kosení a s ním souvisejícím odstraňováním živin. Jiná situace je na Slovensku, konkrétně na vápenatých slatinách v NP Slovenský raj, kde je nízká produkce nadzemní biomasy udržována i bez pravidelného managementu. Toto může být vysvětleno vysokou a stabilní hladinou spodní nebo proudící vody (stanová 2000) a nízkou dostupností draslíku a fosforu v půdě, tedy prvků, které jsou důležité pro růst rostlin (rozbrojová & hájek 2008). Jedinci L. sibirica rostoucí na těchto stanovištích mají malý počet listových růžic, kvetoucích stvolů a květů v květenství a nízkou produkci semen na květ. Nutno podotknout, že na první pohled menší robustnost dospělých rostlin rostoucích na těchto stanovištích neznamená, že populace druhu je v ohrožení. Opak je pravdou, neboť díky

Příroda, Praha, 31: 127–147, 2011
143
nízké produkci biomasy, vysoké hladině spodní vody a vyvinutému mechovému patru jsou v populaci přítomna všechna životní stádia druhu včetně semenáčků.
Naproti tomu populace L. sibirica rostoucí na stanovištích bohatých na dusík/živiny prosperují hůře. Jak již víme, rychlost populačního růstu u těchto populací klesá. Jedná se zejména o populace rostoucí na v minulosti odvodněných stanovištích. Odvodněním rašelinišť dochází k provzdušnění povrchové vrstvy rašeliny a tím k její mineralizaci, čímž se uvolňují živiny (stanová 2000) a zvyšuje se produkce biomasy (nízký C/N poměr v půdě). Vysoká produkce biomasy na těchto lokalitách přímo souvisí s vyšším příjmem dusíku a fosforu vegetací. Dospělí jedinci L. sibirica rostoucí na těchto stanovištích jsou celkově robustnější, tj. mají větší počet listových růžic, kvetoucích stvolů, květů v květenství a vyšší produkci semen na květ. Bohužel tyto populace jsou složeny převážně ze starších dospělých rostlin a trpí nedostatkem semenáčků. Dlouhodobé přežívání těchto populací je tedy velmi pochybné. Snahy o obnovení životaschopnosti těchto populací by měly být zaměřeny především na podporu klíčení, přežívání a růstu semenáčků. K tomu je nutné nejprve zvýšení hladiny spodní vody. Nicméně samotné zvýšení hladiny spodní vody nebude mít vliv na omezení produkce biomasy. Následně je tedy nezbytné pokračovat každoročním kosením a odstraňováním biomasy.
Velké populace L. sibirica se nacházejí především na alkalických stanovištích chudých na dusík/živiny, s vysokým pH a nízkou produkcí biomasy (vysoký C/N poměr v půdě), což jsou charakteristiky společenstev vápnitých slatin. Malé populace se nacházejí naopak především na stanovištích s vysokým obsahem dusíku/živin a velkou produkcí biomasy, tedy stanovištích, kde došlo k mineralizaci rašeliny k uvolnění živin.
vliv kvality stanoviště, velikosti populace a genetické variability na vitalitu populací L. sibirica
Rychlost populačního růstu, příspěvek jednotlivých přechodů k rychlosti populačního růstu (elasticita) a očekávaná délka života L. sibirica byla silně ovlivněna kvalitou stanoviště, zatímco velikost populace a genetická variabilita nehrála žádnou roli v případě čistého vlivu těchto faktorů. Nicméně kvalita stanoviště a velikost populace měly smíšený vliv na očekávanou délku života. Rychlost populačního růstu výrazně vzrostla se zvyšující se kvalitou stanoviště. Se snižující se kvalitou stanoviště roste důležitost elasticity přežívání a zároveň sklesá elasticita růstu a reprodukce. Již publikované práce našly vztah mezi velikostí populace, stanovištními podmínkami a rychlostí populačního růstu. Například byl nalezen pozitivní vztah mezi velikostí populace a rychlostí populačního růstu u druhů Gentianella germanica (fischer & Matthies 1998) a Scorzonera hispanica (MünzberGová 2006). Naše výsledky jsou v souladu s výsledky několika dalších studií (např. brys et al. 2005, collinG & Matthies 2006, schleuninG & Matthies 2009), které zjistily, že zhoršení kvality stanoviště vedlo ke snížení rychlosti populačního růstu. Naše zjištění tak podporují závěr, že obohacení půdy dusíkem či vyšší produktivita půdy negativně ovlivňuje vitalitu populací vytrvalých bylin (brys et al. 2005, collinG & Matthies 2006). V kapitole Vitalita populací jsme ukázaly, že přechodem, který nejvíce přispěl k rychlosti populačního růstu, bylo přežívání jedinců. A skutečně jsme zjistily, že tento přechod byl silně ovlivněn kvalitou stanoviště. Toto spolu se skutečností, že očekávaná délka života kvetoucích dospělých jedinců je nižší v populacích rostoucích na stanovištích nižší kvality, naznačuje, že dospělé rostliny nemusí být již schopny dále vyrovnávat negativní vlivy degradace stanovišť na populace (brys et al. 2005). Časové zpoždění mezi zhoršením kvality stanovištních podmínek a zánikem celé populace tak může být výrazně zkráceno (brys et al. 2005, schleuninG & Matthies 2009).

144
Heinken-Šmídová & Münzbergová: Faktory ovlivňující populační dynamiku Ligularia sibirica
shrnutí
Výsledky naší studie ukazují, že většina studovaných populací L. sibirica, malých i velkých, prosperuje dobře a mají velmi nízkou pravděpodobnost zániku. Mezi studovanými populacemi je však jedna populace, která v současnosti neprosperuje dobře a jejíž početnost prokazatelně klesá, a další dvě populace, které mají tendenci k poklesu rychlosti populačního růstu. V další části naší studie jsme zjistily, že si populace L. sibirica zachovaly vysokou úroveň genetické variability a že genetická variabilita pozitivně koreluje s velikostí populace. Výjimkou jsou malé populace v severních Čechách, jejichž menší genetická variabilita je pravděpodobně způsobena zničením stanoviště meliorací v minulém století. Výsledky analýzy stanovištních poměrů ukázaly, že populace rostoucí na stanovištích chudých na dusík (vysoká hladina podzemní vody, neutrální až zásadité pH, malé množství živin, konkrétně dusíku v půdě a nízká produkce biomasy) prosperují lépe než populace rostoucí na stanovištích bohatých na dusík (nízká hladina podzemní vody, vysoký obsah dusíku/živin v půdě a vysoká produkce biomasy) a to i přes to, že jedinci jsou menšího vzrůstu a mají nižší produkci semen. Nakonec jsme regresní analýzou zjistily, který z faktorů (genetická variabilita, stanovištní podmínky nebo velikost populace) ovlivňují vitalitu populací L. sibirica. Výsledky ukázaly, že rychlost populačního růstu a význam jednotlivých přechodů pro růst populací (elasticita) jsou ovlivněny především kvalitou stanoviště. Zjistily jsme tedy, že velikost populace a genetická variabilita nejsou hlavním problém při ochraně L. sibirica. Ochrana tohoto vzácného druhu by se měla zaměřit hlavně na obnovení stanovištních podmínek degradovaných lokalit.
summary
The results of our study indicate that most of the studied populations of Ligularia sibirica, both small and large, are not declining and show very low extinction probabilities. There is only one population which is currently declining and other two populations that tend towards decline. In other parts of our study, we found that populations of L. sibirica still retained high levels of their genetic variability and that the genetic variability is positively correlated with population size. Small populations in northern Bohemia represent an exception. Their lower genetic variability is probably a result of deteriorated habitats caused by destructions of former fens by meliorations during the last century. The results of analyses of habitat quality showed that populations growing in nitrogen-poor habitats (high water table, neutral to alkaline pH, low content of nutrients in soil - namely nitrogen, low production of biomass), are performing better than populations growing in nitrogen-rich habitats (low water table, high content of nutrients in soil - namely nitrogen, and production of biomass), even though the individuals are smaller in size and the populations have lower seed production. We performed regression analyses to detect which of the factors (e.g. genetic diversity, habitat conditions, or population size) influenced the population performance of L. sibirica. Our findings indicate that the population growth rate of L. sibirica and the importance of single transitions to population growth rate (elasticity) are mainly affected by habitat conditions. Our results indicate that the population size and genetic diversity are not a major problem for the conservation of L. sibirica. The conservation activity should focus on restoring habitat conditions of the localities.

Příroda, Praha, 31: 127–147, 2011
145
Poděkování
Chtěly bychom poděkovat pracovníkům v chráněných územích České a Slovenské republiky za poskytnutí informací o lokalitách L. sibirica. Povolení pro manipulaci s tímto druhem a pro vstup na lokality jsme získaly od ministerstev životního prostředí České a Slovenské republiky. Tato studie byla podpořena grantem MŠMT VaV 2B06178 a částečně také MŠMT 0021620828 a AV0Z60050516.
literatura
ÅGren j. (1996): Population size, pollinator limitation, and seed set in the self-incompatible herb Lythrum salicaria. – Ecol. 77: 1779–1790.
brys r., jaQueMyn h., enDels P., De blust G. & herMy M. (2005): Effect of habitat deterioration on population dynamics and extinction risks in a previously common perennial. – Conserv. Biol. 19: 1633–1643.
caswell h. (2001): Matrix population models, construction, analysis, and interpretation. – Sinauer Associates, Sunderland, MA.
cochran M. e. & ellner s. (1992): Simple methods for calculating age-based life history parameters for stage-structured populations. – Ecol. Monographs 62: 345–364.
collinG G., Matthies D. & reckinGer c. (2002): Population structure and establishment of the threatened long-lived perennial Scorzonera humilis in relation to environment. – J. Appl. Ecol. 39: 310–320.
collinG G. & Matthies D. (2006): Effects of habitat deterioration on population dynamics and extinction risk of an endangered, long-lived perennial herb (Scorzonera humilis). – J. Ecol. 94: 959–972.
council of euroPean coMMunities (1992): Council Directive 92/43/EEC of 21 May on the conservation of natural habitats and of wild fauna and flora. – Official Journal of the European Communities 35: 7–50.
DanneMann a. (2000): Der Einfluss von Fragmentierung und Populationsgröße auf die genetische Variation und Fitness von seltenen Pflanzenarten am Beispiel von Biscutella laevigata (Brassicaceae). – Diss. Bot. 330: 1–151.
De kroon h., van GroenenDael j. & ehrlén j. (2000): Elasticities: a review of methods and model limitations. – Ecol. 81: 607–618.
Doust l. l. (1981): Population dynamics and local specialization in a clonal perennial (Ranunculus repens): I. The dynamics of ramets in contrasting habitats. – J. Ecol. 69: 743–755.
ehrlén j. (1995): Demography of the perennial herb Lathyrus vernus. II. Herbivory and population dynamics. – J. Ecol. 83: 297–308.
ellenberG h., weber h. e., Düll r., wirth v., werner w. & Paulissen D. (1992): Zeigerwerte von Pflanzen in Mitteleuropa. 2nd ed. – Scripta Geobot. 18: 1–258.
ellstranD n. c. & elaM D. c. (1993): Population genetic consequences of small population size: implication for plant conservation. – Annual Rev. Ecol. Syst. 24: 217–242.
eriksson o. (1996): Regional dynamics of plants: a review of evidence for remnant, source-sink and metapopulations. – Oikos 77: 248–258.
eriksson o. (2000): Functional roles of remnant plant populations in communities and ecosystems. – Glob. Ecol. Biogeogr. 9: 443–449.
eXcoffier l. G. l. & schneiDer s. (2005): Arlequin ver. 3.0: An integrated software package for population genetics data analysis. – Evol. Bioinformatics Online 1: 47–50.
feráková v., MaGlocký Š. & MarholD k. (2001): Červený zoznam papraďorastov a semenných rastlín Slovenska (december 2001). – In: baláž D., MarholD k. & urban P. [eds], Červený zoznam rastlín a živočíchov Slovenska, Ochr. Prír., Banská Bystrica, 20, suppl.: 48–81.
fischer M. & Matthies D. (1998): Effects of population size on performance in the rare plant Gentianella germanica. – J. Ecol. 86: 195–204.
frankhaM r., ballou j. D. & briscoe D. a. (2006): Introduction to conservation genetics. – Cambridge University Press, Cambridge.

146
Heinken-Šmídová & Münzbergová: Faktory ovlivňující populační dynamiku Ligularia sibirica
GauDeul M., taberlet P. & till-bottrauD i. (2000): Genetic diversity in an endangered alpine plant, Eryngium alpinum L. (Apiaceae), inferred from amplified fragment length polymorphism markers. – Mol. Ecol. 9: 1625–1637.
haMrick j. l. & GoDt M. j. w. (1989): Allozyme diversity in plant species. – In: brown a. h. D., cleGG M. t., kahler a. l. & weir b. s. [eds], Plant population genetics, breeding and genetic resources, pp. 43–63, Sinauer Associates, Sunderland, Massachusetts.
heinken-ŠMíDová a. & MünzberGová z. (v tisku): Population dynamics of the endangered, long-lived perennial species, Ligularia sibirica. – Folia Geobot.
henDrych r. (2003): Poznatky o druhu Ligularia sibirica v Čechách. – Preslia 75: 39–69.hensen i., oberPrieler c. & wesche k. (2005): Genetic structure, population size, and seed production
of Pulsatilla vulgaris Mill. (Ranunculaceae) in central Germany. – Flora 200: 3–14.hewitt G. M. (1999): Post-glacial re-colonization of European biota. – Bot. J. Linn. Soc. 68: 87–112.hiGGins s. i., Pickett s. t. a. & bonD w. j. (2000): Predicting extinction risk for plants: environmental
stochasticity can save declining populations. – Trends Ecol. Evol. 15: 516–520.kneblová v. (1950): Reservace u Rečkova a Ligularia sibirica (L.) Cass. – Čs. Bot. Listy 3: 17–21.králová M. (1990): Vybrané chemické analýzy půd a rostlin. – Studie ČSAV 12: 1–152.leiMu r., Mutikainen P., koricheva j. & fischer M. (2006): How general are positive relationships
between plant population size, fitness and genetic variation? – J. Ecol. 94: 942–952.liu j. Q. (2004): Uniformity of karyotypes in Ligularia (Asteraceae: Senecioneae), a highly diversified genus
of the eastern Qinghai-Tibet Plateau highlands and adjacent areas. – Bot. J. Linn. Soc. 144: 329–342.lowe a., harris s. & ashton P. (2004): Ecological genetics: design, analysis, and application. –
Blackwell publishing, Oxford.lutz e., schneller j. j. & holDereGGer r. (2000): Understanding population history for conservation
purposes: population genetic of Saxifraga aizoides (Saxifragaceae) in the lowlands and lower mountains north of the Alps. – Am. J. Bot. 87: 583–590.
MarŠáková-něMejČková M. (1973): Popelivka sibiřská. – Ochr. Přír. 28: 15–16.Mathworks inc. (2006): Matlab version 7.3.0.267. – Mathworks Inc., Natick, MA, USA.Mattauch f. (1936): Über die Ligularia sibirica (L.) Cass. – Natur Heimat, Aussig 7: 57–62.MenGes e. s. (2000): Population viability analyses in plants: challenges and opportunities. –Trends
Ecol. Evol. 15: 51–56.Meusel h. & jäGer j. (1992): Vergleichende Chorologie der zentraleuropäischen Flora. Vol. 3. –
G. Fischer, Jena.MünzberGová z. (2006): Effect of population size on prospect of species survival. – Folia Geobot. 41:
137–150.oosterMeijer j. G. b., van veer r. & Den nijs j. c. M. (1994): Population structure of the rare, long-lived
perennial Gentiana pneumonanthe in relation to vegetation and management in the Netherlands. – J. Appl. Ecol. 31: 428–438.
oosterMeijer j. G. b., bruGMan M. l., De boer e. r. & Den nijs h. c. M. (1996): Temporal and spatial variation in the demography of Gentiana pneumonanthe, a rare perennial herb. – J. Ecol. 84: 153–166.
oosterMeijer j. G. b., luijten s. h., křenová z. & Den nijs j. c. M. (1998): Relationship between population and habitat characteristics and reproduction of the rare Gentiana pneumonanthe L. – Conserv. Biol. 12: 1042–1053.
oosterMeijer j. G. b., luijten s. h. & Den nijs j. c. M. (2003): Integrating demographic and genetic approaches in plant conservation. – Biol. Conserv. 113: 389–398.
ouborG n. j., verGeer P. & MiX c. (2006): The rough edges of the conservation genetics paradigm for plants. – J. Ecol. 94: 1233–1248.
overbeck G., kiehl k. & abs c. (2003): Seedling recruitment of Succisella inflexa in fen meadows: Importance of seed and microsite availability. – Appl. Veg. Sci. 6: 97–104.
ozinGa w. a. & schaMinée j. h. j. [eds] (2005): Target species – Species of European concern. A database driven selection of plant and animal species for the implementation of the Pan European Ecological Network. – Alterra-report 1119, Wageningen, Alterra.
Peterson a., bartish i. v. & Peterson j. (2008): Effects of population size on genetic diversity, fitness and pollinator community composition in fragmented populations of Anthericum liliago L. – Plant Ecol. 198: 101–110.

Příroda, Praha, 31: 127–147, 2011
147
Procházka f. [ed.] (2001): Černý a červený seznam cévnatých rostlin České republiky (stav v roce 2000). – Příroda, Praha, 18: 1–166.
raMula s. (2006): Population viability analysis for plants: Practical recomendations and applications. – Ms. [PhD thesis; Department of Botany, Stockholm University & Jannes Kuvertproffset HB, Stockholm, Sweden.]
reisch ch., PoschloD P. & winGenDer r. (2003): Genetic variation of Saxifraga paniculata Mill. (Saxifragaceae): molecular evidence for glacial relict endemism in central Europe. – Biol. J. Linn. Soc. 80: 11–21.
rozbrojová z. & hájek M. (2008): Changes in nutrient limitation of spring fen vegetation across environmental gradients in the West Carpathians. – J. Veg. Sci. 19: 613–620.
schaffers a. P. & sýkora k. v. (2000): Reliability of Ellenberg indicator values for moisture, nitrogen and soil reaction: a comparison with field measurements. – J. Veg. Sci. 11: 225–244.
shefferson r. P. (2009): The evolutionary ecology of vegetative dormancy in mature herbaceous perennial plants. – J. Ecol. 97: 1000–1009.
schleuninG M. & Matthies D. (2009): Habitat change and plant demography: Assessing the extinction risk of a formerly common grassland perennial. – Conserv. Biol. 23: 174–183.
schoPP-Guth a., Maas D. & PfaDenhauer j. (1994): Influence of management on the seed production and seed bank of calcareous fen species. – J. Veg. Sci. 5: 569–578.
silvertown j. & franco M. (1993): Plant demography and habitat: a comparative approach. – Plant Spec. Biol. 8: 67–73.
silvertown j., franco M. & MenGes e. (1996): Interpretation of elasticity matrices as an aid to the management of plant populations for conservation. – Conserv. Biol. 10: 591–597.
slavík b. (2004): Ligularia Cass. - popelivka. – In: slavík b. & ŠtěPánková j. [eds], Květena České republiky 7, pp. 306–309, Academia, Praha.
soulé M. e. (1987): Viable populations for conservation. – Cambridge University Press, Cambridge.s-Plus (2000): Professional edition for Windows, release 2. (1999) – MathSoft Inc., Needham,
Massachusetts, USA.stanová v. [ed.] (2000): Rašeliniska Slovenska. – Daphne – Inštitút aplikovanej ekológie, Bratislava.ŠMíDová a., MünzberGová z. & PlaČková i. (2011): Genetic diversity of a relict plant species, Ligularia
sibirica (L.) Cass. (Asteraceae). – Flora 206: 151–157.ter braak c. j. f. & ŠMilauer P. (2002): CANOCO Reference manual and CanoDraw for Windows
User’s Guide: Software for canonical community ordination, Version 4.5. – Microcomputer Power, Ithaca, USA.
thoMPson k. & GriMe j. P. (1979): Seasonal variation in the seed banks of herbaceous species in ten contrasting habitats. – J. Ecol. 67: 893–921.
van rossuM f. & Prentice h. c. (2004): Structure of allozyme variation in Nordic Silene nutans (Caryophyllaceae): population size, geographical position and immigration history. – Biol. J. Linn. Soc. 81: 357–371.
verGeer P. v., renGelink r., coPal a. & ouborG n. j. (2003): The interacting effects of genetic variation, habitat quality and population size in performance of Succisa pratensis. – J. Ecol. 91: 18–26.
walter h. & straka h. (1970): Arealkunde - Floristisch-historische Geobotanik. Ed. 2. – Eugen Ulmer, Stuttgart.
yeh f. c. & boyle t. j. b. (1997): Population genetic analysis of co-dominant and dominant markers and quantitative traits. – Belg. J. Bot. 129: 157.
zbíral j. (1995): Analýza půd I, Jednotné pracovní postupy. – Státní a kontrolní a zkušební ústav zemědělský, Brno.

148
Heinken-Šmídová & Münzbergová: Faktory ovlivňující populační dynamiku Ligularia sibirica

Příroda, Praha, 31: 149–165, 2011
149
Populační biologie a stanovištní nároky druhů z okruhu Minuartia verna agg. Population biology and habitat requirements of species belonging to Minuartia verna agg.
hana Pánková1 & ZuZana MünZbergová1, 2
1Botanický ústav Akademie věd ČR, Zámek 1, CZ-252 43 Průhonice, [email protected] botaniky, Přírodovědecká fakulta Univerzity Karlovy, Benátská 2, CZ-128 01 Praha 2, [email protected]
Abstract: There are three plant species from Minuartia verna agg. growing in the Czech Republic: Minuartia smejkalii, M. corcontica and M. caespitosa. All the species are considered as critically endangered plant species, M. corcontica and M. smejkalii are Czech endemics. All these species occur in extreme habitats. In this study we tested whether these species behave as typical S strategists or whether they have some traits characteristic for R–strategist. We studied the population dynamics and the habitat preference of these species. The results showed that, in spite of the fact that all species occurs on different extreme habitats, their life cycle is quite similar. All species are long-lived, which is typical of stress tolerant plant species. On the other hand, they show high seed production and high natality and mortality, which is typical of R–strategists. For these reasons, Minuartia species could be considered as SR strategists. Nevertheless, each species has its specific life stages, which should be aimed at in the process of their conservation. Keywords: critically endangered species, endemic, Minuartia caespitosa, M. corcontica, M. smej kalii, practical conservation.
Abstrakt: V České republice se vyskytují tři druhy spadající do okruhu Minuartia verna agg.: Minuartia smejkalii, M. corcontica a M. caespitosa. Všechny tři druhy jsou v České republice zařazeny mezi kriticky ohrožené, druhy M. corcontica a M. smejkalii jsou českými endemity. Všechny druhy osidlují extrémní stanoviště. V této studii jsme pomocí studia populační dynamiky a stanovištních nároků druhů zjišťovali, do jaké míry se druhy chovají jako typičtí S stratégové a do jaké míry vykazují spíše charakteristiky ruderálních druhů (R stratégů). Výsledky ukázaly, že přestože všechny kuřičky osidlují velmi odlišná extrémní stanoviště, jejich životní cyklus je velmi podobný. Kuřičky odpovídají svoji dlouhověkostí charakteristikám stres tolerantních druhů, zatímco vysokou produkcí semen a vysokým obratem odpovídají spíše R stratégům. Studované druhy lze tedy považovat za SR stratégy. Nicméně růstová dynamika každého druhu má svá specifická stádia, na která je potřeba se při ochraně těchto druhů zaměřit. Klíčová slova: endemit, kriticky ohrožený druh, kuřička hadcová, k. hercynská, k. krkonošská, praktická ochrana druhu
ÚvodTaxonomie kuřiček spadajících do okruhu Minuartia verna (sekce Polymechana) je velmi komplikovaná a mezi autory nepanuje ani shoda, které druhy by v něm měly být zařazeny (hejný & slavík 1990, Dvořáková 2003, Dúbravková et al. 2010). Zatímco v České republice se jednotlivé taxony z okruhu M. verna vyčleňují v samostatné druhy (Dvořáková 1985, 1988, 1999, 2003), v zahraničí jsou obvykle popisovány jako M. verna (bauMbach 2005)

150
Pánková & Münzbergová: Populační biologie a stanovištní nároky Minuartia verna agg.
či jsou odděleny jako podruhy či ekotypy (např. neuMann et al. 1997). V České republice se vyskytují tři druhy spadající do okruhu Minuartia verna agg.: Minuartia smejkalii a M. corcontica, které by mohly být příbuzné se skupinou vysokohorských kuřiček, a M. caespitosa, která je nejspíše příbuzná severské kuřičce M. rubella (Dvořáková in hejný & slavík 1990). V minulosti byl ještě udáván výskyt druhu M. glaucina z Pavlovských vrchů, který má však jinou ploidii (Dvořáková 1985).
Druhy spadající do okruhu M. verna agg. jsou charakteristické tím, že osidlují stanoviště, u nichž lze alespoň jeden faktor považovat za extrémní. Většina těchto druhů osidluje holé skály či skalní štěrbiny ve vysokých nadmořských výškách (Alpy, Dolomity) či se vyskytují na substrátech s vysokými obsahy těžkých kovů (hadce, rudné výsypky). Rostliny extrémních stanovišť se obvykle řadí mezi S-stratégy. Pro S-stratégy je typické, že mají pomalou rychlost růstu, mají nízkou produkci, jsou to rostliny vytrvalé, květy a semena netvoří každým rokem, mají malé listy, často jehlice, nebo listy neopadavé, vždyzelené, někdy sukulentní, poměr roční produkce uložené v semenech je malý, mají pomalý obrat uhlíku, minerálních látek a vody, jeví nízkou fenotypickou plasticitu a často se šíří také vegetativně (slavíková 1986). Z těchto typických znaků by se dalo usoudit, že pro zachování dobré vitality populací S-stratégů, a tedy i kuřiček, bude klíčovým faktorem přežívání dospělých jedinců na stanovišti.
Důležitou charakteristikou všech výše zmíněných stanovišť je i to, že je na nich velmi nízká kompetice. Některé z těchto stanovišť jsou navíc relativně nestabilní (rudné výsypky, místa s obnaženým substrátem na hadcových skalkách). To naznačuje, že druhy okruhu M. verna agg. jsou velmi kompetičně slabé a mohly by vykazovat některé charakteristiky R-stratégů. Potom bychom očekávali, že tyto druhy budou relativně krátkověké a budou produkovat velké množství semen. To, do jaké míry lze charakterizovat druhy jako S-stratégy a do jaké míry se chovají spíše jako R-stratégové, bude významně určovat způsob managementu populací těchto druhů.
K stanovení optimálního managementu populací druhů jsme proto studovali populační dynamiky druhů a jejich stanovištní nároky. Konkrétně jsme chtěli odpovědět na tyto otázky: 1) jaká je populační dynamika tří druhů M. verna agg. rostoucích v České republice, 2) jak se dynamika liší mezi našimi druhy, 3) jaké jsou stanovištní nároky studovaných druhů, v čem jsou podobné a v čem se naopak liší a 4) jak závisí růstové charakteristiky druhů na stanovištních podmínkách.
studované druhy
Všechny tři druhy jsou v České republice zařazeny do kategorie kriticky ohrožené, druhy M. corcontica a M. smejkalii jsou českými endemity (Procházka 2001).
Jednotlivé typy kuřiček z okruhu M. verna se liší svým areálem a některými mor fo-lo gickými znaky (nejčastěji se uvádí povrchová struktura semene, foto 1). Na základě těchto znaků se kukuřičky vyčleňují do samostatných druhů a poddruhů či jsou vedeny jako ekotypy. Nicméně, i kdyby všechny takto vyčleněné typy byly pouze jedním druhem, jednalo by se o druh vzácný na úrovni celé EU.
Minuartia smejkalii (kuřička hadcová) je s největší pravděpodobností pozdně glaciálním nebo raně postglaciálním reliktem. Jako samostatný druh byla popsána teprve v roce 1988 Dvořákovou jako derivát vysokohorské M. gerardii (Dvořáková 1988). Kuřička hadcová je obligátním serpentinofytem vázaným na skalní plošiny, štěrbiny a mělké hadcové půdy nezapojených travinných porostů ve světlých perialpinských borech (svazy Asplenion serpentinii a Asplenio cuneifolii-Armerion serpentini). Kuřička hadcová je charakteristická velikostí květů 6–8 mm, nesouměrně okrouhle ledvinovitými semeny
Příroda, Praha, 31: 149–165, 2011
151
o velikosti 0,4–0,6 mm, rezavě až sytě hnědé barvy, na celém povrchu papilnatými. Papily jsou válcovité až úzce válcovité nebo kuželovité, alespoň na hřbetu zřetelně vyšší než širší (foto 1). V České republice se již vyskytuje pouze na dvou lokalitách (mapa 1).Centrem výskytu druhu je národní přírodní památka Hadce u Želivky (Dolnokralovické hadce, mapa 2). Tato oblast byla v minulosti značně narušena stavbou dálnice D1 a stavbou vodní nádrže Švihov, takže v současné době se zde kuřička hadcová nachází v sedmi oddělených populacích. V údolí bývalého Sedlického potoka se nachází celkem pět populací (Dolnokralovické hadce I–V), zbylé dvě se vyskytují v blízkosti bývalé říčky Želivky (Bernartice I a II). Populace se vyskytují jak na otevřených hadcových skalkách příkrých svahů v perialpinských borech s jižní orientací (populace DKI, DKII (Morava), DKV, BII), tak i na skalkách se severní orientací, které jsou vegetačně odlišné (DKIV). Kuřička hadcová osidluje v této oblasti i druhotně vzniklá stanoviště (opuštěný malý hadcový lom – BI, obnažené těleso bývalé protektorátní dálnice – DKIII). Druhou oblastí výskytu jsou Hadce u Hrnčíř, kde se kuřička hadcová nachází v hustém kulturním borovém lese. Bližší informace o jednotlivých populacích jsou uvedeny v samostatné publikaci (Pánková 2011). V minulosti se kuřička hadcová ještě vyskytovala v PP Borecká skalka u Chotěboře, kde však vyhynula dříve, než byl druh samostatně vyčleněn.
Minuartia caespitosa (kuřička hercynská) je druh skalnatých svahů, teras a štěrbin vápnitých pískovců, řidčeji se též vyskytuje na otevřených rozvoněných travnatých místech a písčinách ve světlých borech na sprašových vápnitých píscích nebo pískovcových říčních terasách. Celkový areál druhu je roztříštěn do několika arel, které jsou roztroušeny od V Belgie, přes SZ Německo až do S Čech (Dvořáková in hejný & slavík 1990). V minulosti se udával výskyt ještě v blízkosti Bělé pod Bezdězem. Od ostatních kuřiček z okruhu M. verna se odlišuje přítomností několikalistových svazečků v úžlabí kvetoucích i sterilních lodyh. Dalším znakem jsou semena, která mají na svém povrchu pouze jemně zrnité papily (foto 1). a) b) c)
Foto 1. Struktury na povrchu semen u kuřičky hadcové (a), kuřičky hercynské (b) a kuřičky krkonošské (c). Foto Jiří Machač, Botanický ústav AV ČR, Průhonice.Fig. 1. Structure of the seed cover in Minuartia smejkalii (a), M. caespitosa (b), and M. corcontica (c). Photo by Jiří Machač, Institute of Botany, Academy of Sciences of the Czech Republic, Průhonice
Kuřička hercynská se v České republice vyskytuje na několika skalkách v oblasti Hradčanských stěn u Mimoně (sáDlo et al. 2011), nicméně v době počátku výzkumu byly známy pouze dvě populace: Jelení vrch a Jelení skála. Populace se vyskytují na vápnitých píscích, které tvoří při úpatí skal menší písečné přesypy, které jsou v horní části poměrně nestabilní a dynamické, takže i vegetace je zde poměrně řídká. Spodní část písčiny je již více porostlá kostřavami a plynule přechází v kulturní borový les. Kuřička hercynská tedy
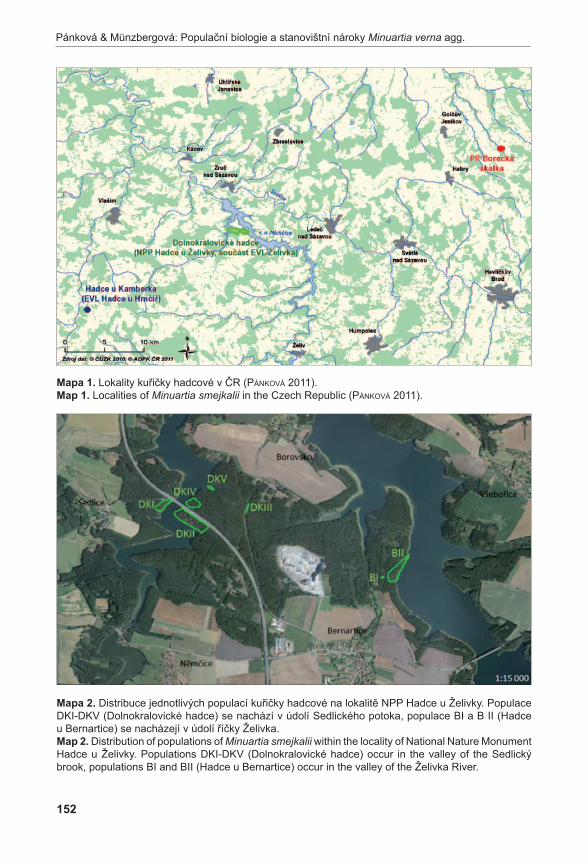
152
Pánková & Münzbergová: Populační biologie a stanovištní nároky Minuartia verna agg.
Mapa 1. Lokality kuřičky hadcové v ČR (Pánková 2011). Map 1. Localities of Minuartia smejkalii in the Czech Republic (Pánková 2011).
Mapa 2. Distribuce jednotlivých populací kuřičky hadcové na lokalitě NPP Hadce u Želivky. Populace DKI-DKV (Dolnokralovické hadce) se nachází v údolí Sedlického potoka, populace BI a B II (Hadce u Bernartice) se nacházejí v údolí říčky Želivka. Map 2. Distribution of populations of Minuartia smejkalii within the locality of National Nature Monument Hadce u Želivky. Populations DKI-DKV (Dolnokralovické hadce) occur in the valley of the Sedlický brook, populations BI and BII (Hadce u Bernartice) occur in the valley of the Želivka River.

153
Příroda, Praha, 31: 149–165, 2011
osidluje tři vegetačně odlišné typy – pískovcové skalky či písčiny se sporadickou vegetací (vrchol Jeleního vrchu – S2, skalky na úpatí Jelení skály – S1), travinný porost (spodní část Jelení skály – T1) a borový les (pod oběma lokalitami – L1).
Minuartia corcontica (kuřička krkonošská) patří obdobně jako k. hadcová mezi ne-oendemity České republiky. Jako samostatný druh byla vyčleněna v roce 1999 jako derivát alpské M. gerardii (Dvořáková 1999). Kuřička krkonošská roste ve společenstvech skalních štěrbin a terásek ledovcových karů vysokých pohoří Sudet (společenstvo Agrostion alpinae). Od kuřičky hadcové se odlišuje většími květy (8–12 mm) i většími semeny (0,7–1 mm). Papily semen jsou válcovité nebo kuželovité, zřetelně vyšší než široké (foto 1). V České republice je udáván výskyt pouze v Čertově zahrádce a v Čertově rokli, kde osidluje erlány, což jsou pyroxenové ruly vzniklé z karbonátových hornin s vysokým obsahem rudných minerálů a sulfidů.
Horní část Čertovy zahrádky (Velká Čertova zahrádka – VC) je strmá, obtížně prostupná skalnatá rokle na východním svahu Studniční hory, dolní část je tvořena suťovým kuželem spadajícím až na dno Obřího dolu, jen místy zpevněným a porostlým vegetací. Populace kuřičky krkonošské ve Velké Čertově zahrádce se nachází ve vrcholové části suťového kuželu. Kuřička zde osidluje štěrbiny kolmých skalek. Vegetace je zde díky dynamičnosti svahu velmi sporadická, takže kuřička téměř není vystavena kompetici. Výzkum probíhal pouze ve spodní části populace, nikoliv na vrcholu suťového kužele, protože vyšší část lokality je nepřístupná. Lokalita Malá Čertova zahrádka (MC) se nachází ca 100 m od suťového kužele směrem k Peci pod Sněžkou. Lokalita je méně strmá, vegetace je zde mnohem hustší a zarostlá mechem, takže kuřička je zde vystavena silnější kompetici.
Metodika
sledování populační dynamiky
Pro hodnocení populační dynamiky všech tří druhů se využila upravená metodika pro intenzivní monitoring kuřičky hadcové (Pánková 2011), který probíhá pod záštitou AOPK. Sledování populační dynamiky probíhalo od roku 2006, pro druh M. corcontica až od roku 2007. Z tohoto důvodu jsou prezentované výsledky založeny pouze na datech od prvního kompletního přechodu, tj. od roku 2008.
Na každé lokalitě bylo vytyčeno několik trvalých ploch o velikost 1 × 1 m či 0,5 × 0,5 m. V každé ploše byla zaznamenána poloha a velikost trsu na milimetrový papír za pomoci čtvercového rámu rozděleného vnitřní sítí s velikostí oka 10 × 10 cm. Každému jedinci bylo přiřazeno jedno číslo. Tyto mikromapy byly dále zpracovány v programu GIS a využity pro sledování vývoje velikosti a polohy trsů mezi jednotlivými roky.
Rostliny, které se vyskytují mimo čtverce, či se vyskytují na lokalitách, kde nebylo možné čtverce umístit, byly očíslovány pomocí kovových štítků s vyraženým číslem. Nově vzniklým jedincům bylo vždy přiřazeno nové číslo. Na každé lokalitě byl taktéž proveden schematický zákres s rozmístěním jednotlivých trsů.
Pomocí mikromap a zákresů bylo možno taktéž rozlišit, zda nový jedinec vznikl vegetativně (odnož původní rostliny) či generativně či se jedná o jedince regenerovaného po dormanci. Za jedince regenerovaného z dormance se považuje jedinec, který minimálně jednu vegetační sezónu nevyrostl, a poté došlo k jeho regeneraci. U každé rostliny jak ve čtvercích, tak mimo čtverec, byla každý rok změřena délka a šířka trsu, spočten počet kvetoucích i nekvetoucích lodyh a počet květů na jednu lodyhu. Za délku trsu je považován

154
Pánková & Münzbergová: Populační biologie a stanovištní nároky Minuartia verna agg.
nejdelší rozměr trsu. Šířka je rozměr kolmý na délku. Vynásobením délky a šířky byla spo čítána přibližná plocha trsu. V roce 2009 bylo sebráno od 20 jedinců v každé populaci deset zralých neotevřených tobolek a byla vyhodnocena průměrná produkce semen na jeden trs. U druhu M. corcontica byly odebírány pouze tobolky v Malé Čertově zahrádce, kde je populace stabilnější než ve Velké Čertově zahrádce. Na každém stanovišti byl spočítán i celkový počet jedinců.
U druhu M. smejkalii byly monitorovány všechny trsy na všech lokalitách (tj. celkem ca 400 trsů). U druhu M. corcontica byly vytyčeny dva pevné čtverce o velikosti 1 × 1 m ve Velké Čertově zahrádce (celkem ca 60 jedinců) a dva v Malé Čertově zahrádce (celkem ca 180 jedinců). U druhu M. caespitosa byly vytyčeny pevné čtverce na vrcholu Jeleního vrchu (celkem ca 130 trsů), v travinném porostu (celkem ca 150 trsů) a v lese (celkem ca 190 trsů), na skalkách na úpatí Jelení skály byly trsy značeny štítkem (celkem ca 300 jedinců).
Hodnocení klíčivosti semen, klíčivost ze semenné banky
Pro porovnání klíčivosti mezi jednotlivými druhy byly sebrány tobolky se zralými semeny z 20 jedinců. Semena byla spočítána a 50 ks od každého jednice bylo dáno klíčit na Petriho misky do klimaboxu. Pro druh M. smejkalii se podařilo získat větší množství semen, tak byla klíčivost testována i na standardní směsi substrátu s pískem v poměru 1:1 ve skleníku. Každý týden byl jak na Petriho miskách, tak na standardním substrátu spočítán počet vzniklých semenáčků.
Pro druh M. smejkalii byla klíčivost semen testována i v reálných podmínkách na jednotlivých lokalitách. V roce 2007 bylo 50 ks semen od každého jedince vyseto přímo do štěrbin hadcových skal, do vegetace či do strženého substrátu na lokalitách DKI a DKII, od každého ošetření bylo na každé lokalitě pět opakování. Na lokalitě Hrnčíře byla semena vyseta pouze do vegetace a do strženého substrátu, protože na této lokalitě se hadcové štěrbiny nenacházejí. Výsevový experiment byl proveden ještě v roce 2008 na lokalitě BI a DKIII, kde došlo díky managementovým zásahům k vytvoření obnažených ploch bez vegetace.
Pro hodnocení klíčivosti semen ze semenné banky bylo 3 × 50 ks semen od každého jedince uloženo do monofylového pytlíku a zakopáno na původní lokalitu. Semena byla vyjmuta po prvním a druhém roce, byl spočten počet zachovalých semen a tato semena byla dána klíčit na Petriho misky do skleníku.
stanovení ekologických nároků druhů
Pro stanovení abiotických charakteristik stanoviště byly z každé lokality v roce 2006 odebrány půdní vzorky pro stanovení obsahu základních živin i obsah těžkých kovů v půdě. Půdy byly analyzovány v analytické laboratoři BÚ AV ČR v Průhonicích. Hodnoty pH byly měřeny v roztocích destilované vody či v 0,1 M roztoku KCl. Ca2+ a K+ byly měřeny pomocí atomové emisní spektrometrie, Mg2+ byly měřeny pomocí atomové absorpční spektrometrie (Moore & chaPMan 1986, DěDina 1987). Dostupný P byl měřen pomocí fotometrické metody (olsen et al. 1954, olsen & soMMers 1982). Celkový obsah C a N by určen pomocí CHN katalyzátoru (Carlo Erba NC 2500, ehrenberGer & Gorbach 1973). Obsah organického C and obsah karbonátů byl měřen pomocí volumetrické metody (ISO-Standard 10693 1995). Obsah Cu, Zn, Mn, Fe, Cd, Pb, Ni, Co, Cr byl měřen v půdním extraktu roztokem DTPA-TEA metodou AAS v režimu plamenné nebo elektrotermické atomizace (zbíral 1996, linDsay & norwell 1978).
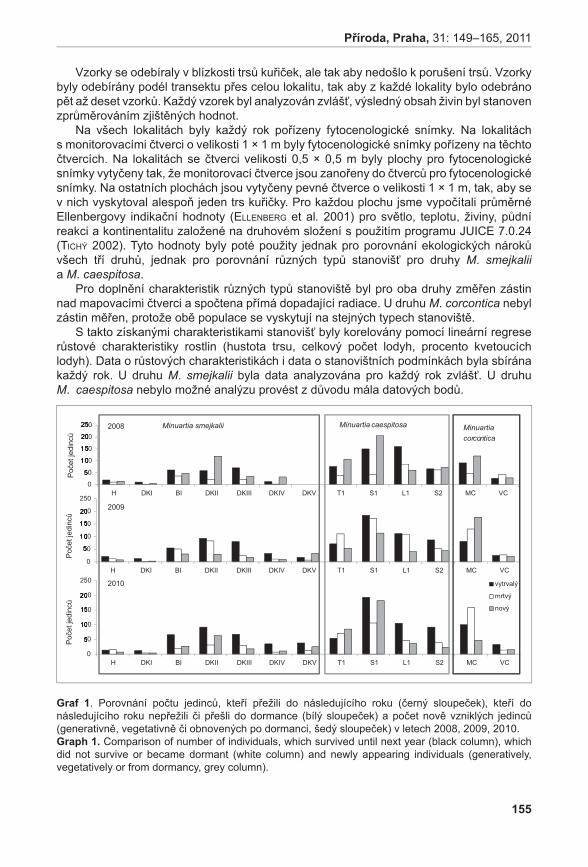
155
Příroda, Praha, 31: 149–165, 2011
Vzorky se odebíraly v blízkosti trsů kuřiček, ale tak aby nedošlo k porušení trsů. Vzorky byly odebírány podél transektu přes celou lokalitu, tak aby z každé lokality bylo odebráno pět až deset vzorků. Každý vzorek byl analyzován zvlášť, výsledný obsah živin byl stanoven zprůměrováním zjištěných hodnot.
Na všech lokalitách byly každý rok pořízeny fytocenologické snímky. Na lokalitách s monitorovacími čtverci o velikosti 1 × 1 m byly fytocenologické snímky pořízeny na těchto čtvercích. Na lokalitách se čtverci velikosti 0,5 × 0,5 m byly plochy pro fytocenologické snímky vytyčeny tak, že monitorovací čtverce jsou zanořeny do čtverců pro fytocenologické snímky. Na ostatních plochách jsou vytyčeny pevné čtverce o velikosti 1 × 1 m, tak, aby se v nich vyskytoval alespoň jeden trs kuřičky. Pro každou plochu jsme vypočítali průměrné Ellenbergovy indikační hodnoty (ellenberG et al. 2001) pro světlo, teplotu, živiny, půdní reakci a kontinentalitu založené na druhovém složení s použitím programu JUICE 7.0.24 (tichý 2002). Tyto hodnoty byly poté použity jednak pro porovnání ekologických nároků všech tří druhů, jednak pro porovnání různých typů stanovišť pro druhy M. smejkalii a M. caespitosa.
Pro doplnění charakteristik různých typů stanoviště byl pro oba druhy změřen zástin nad mapovacími čtverci a spočtena přímá dopadající radiace. U druhu M. corcontica nebyl zástin měřen, protože obě populace se vyskytují na stejných typech stanoviště.
S takto získanými charakteristikami stanovišť byly korelovány pomocí lineární regrese růstové charakteristiky rostlin (hustota trsu, celkový počet lodyh, procento kvetoucích lodyh). Data o růstových charakteristikách i data o stanovištních podmínkách byla sbírána každý rok. U druhu M. smejkalii byla data analyzována pro každý rok zvlášť. U druhu M. caespitosa nebylo možné analýzu provést z důvodu mála datových bodů.
0
50
100
150
200
250
H DKI BI DKII DKIII DKIV DKV T1 S1 L1 S2 MC VC
0
50
100
150
200
250
H DKI BI DKII DKIII DKIV DKV T1 S1 L1 S2 MC VC
0
50
100
150
200
250
H DKI BI DKII DKIII DKIV DKV T1 S1 L1 S2 MC VC
vytrvalý
mrtvý
nový
Minuartia smejkalii Minuartia caespitosa Minuartia corcontica
2008
2009
2010
50
10
15
20
25
Počet
jedi
nců
50
10
15
20
Počet
jedi
nců
50
10
15
20
Počet
jedi
nců
Graf 1. Porovnání počtu jedinců, kteří přežili do následujícího roku (černý sloupeček), kteří do následujícího roku nepřežili či přešli do dormance (bílý sloupeček) a počet nově vzniklých jedinců (generativně, vegetativně či obnovených po dormanci, šedý sloupeček) v letech 2008, 2009, 2010.Graph 1. Comparison of number of individuals, which survived until next year (black column), which did not survive or became dormant (white column) and newly appearing individuals (generatively, vegetatively or from dormancy, grey column).

156
Pánková & Münzbergová: Populační biologie a stanovištní nároky Minuartia verna agg.
výsledky a diskuse
sledování populační dynamiky
Životní cyklus
Životní cyklus je u všech tří druhů velmi podobný. Kuřičky jsou značně dynamickým druhem, kdy každým rokem u nich dochází ke vzniku mnoha nových jedinců, ale zároveň velmi mnoho jedinců uhyne či přejde do dormantního stavu (graf 1). Nicméně některé trsy kuřiček jsou dlouhověké. U druhu M. smejkalii a M. corcontica bylo po pěti letech 27 % trsů vytrvalých, u druhu M. caespitosa stále přežívalo dokonce 35 % trsů.
Kuřičky se rozmnožují především semeny. U všech druhů bylo pozorováno i vegetativní rozmnožování, zejména však u druhu M. caespitosa (graf 2). Vegetativní množení probíhalo rozpadem větších trsů na několik menších a bylo pozorováno především na zastíněných lokalitách či na lokalitách s vyšší kompeticí. U druhu M. corcontica bylo vegetativní rozmnožování pozorováno pouze na lokalitě Malá Čertova zahrádka.
Stádium semenáčku je u všech tří druhů velmi obtížně definovatelné, za semenáček je proto považován nově vzniklý jedinec, který má pouze jednu lodyhu (Pánková 2011). Vývoj nově vzniklých jedinců je velmi variabilní. Na příznivých stanovištích probíhá poměrně rychle: nově vzniklý jedinec je schopen již první vegetační sezónu dosáhnout značných rozměrů, kvést a produkovat semena. Na méně příznivých stanovištích však zůstává ve stavu semenáčku i další vegetační sezónu. U kuřiček se taktéž setkáváme i s retrogresí, kdy kvetoucí jedinec následující rok zůstane ve sterilním stavu. U všech druhů byl pozorován přechod trsů do dormance a jejich následná regenerace i po několika vegetačních sezónách. Počet jedinců obnovených po dormanci se lišil jak mezi jednotlivými druhy, tak mezi jednotlivými roky (graf 2). Zatímco v roce 2008 byl největší nárůst pozorován u druhu M. smejkalii, v následujících letech bylo nejvíce obnovených trsů pozorováno u druhu M. caespitosa. Počet trsů regenerovaných po dormanci se nelišil mezi jednotlivými typy stanoviště. Jedinci regenerovaní po dormanci byly již počátkem vegetační sezóny velicí a obvykle i kvetoucí.
Velikost a vitalita populace
Proporce nově vzniklých jedinců a mrtvých jedinců se lišila mezi jednotlivými roky, druhy i lokalitami (graf 1). U druhu M. corcontica je na lokalitě Velká Čertova zahrádka každým rokem počet nově vzniklých jedinců nižší než počet mrtvých jedinců, takže lze očekávat, že tato populace se bude při zachování stávajících podmínek neustále zmenšovat. Pokles v proporci nově vzniklých jedinců k mrtvým jedincům byl pozorován od roku 2008 u druhu M. smejkalii na lokalitách DKIII a H, což jsou lokality nejvíce ohrožené kompeticí. Naopak větší počet nově vzniklých jedinců oproti mrtvým byl pozorován u druhu M. smejkalii na lokalitě DKV, což je lokalita, která byla objevena teprve v roce 2007. Na této lokalitě se nachází velmi mnoho neobsazených hadcových skalek a lze proto předpokládat, že tato populace bude i nadále růst. U ostatních populací se poměr nově vzniklých a mrtvých jedinců lišil mezi jednotlivými roky. V roce 2008 byl počet nově vzniklých jedinců vyšší než počet mrtvých jedinců na všech lokalitách. Nejméně příznivým rokem pro růst druhu M. smejkalii a M. caespitosa byl rok 2009, který však byl vhodný pro druh M. corcontica. Naopak rok 2010 byl velmi příznivý pro růst druhů M. smejkalii a M. caespitosa, ale u druhu M. corcontica došlo ke snížení poměru nově vzniklých jedinců k mrtvým.
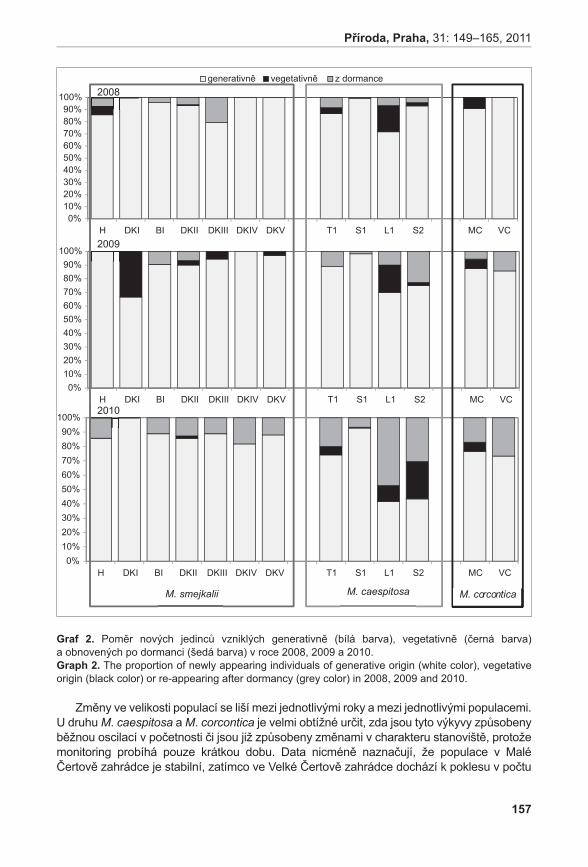
157
Příroda, Praha, 31: 149–165, 2011
0%10%20%30%40%50%60%70%80%90%
100%
H DKI BI DKII DKIII DKIV DKV T1 S1 L1 S2 MC VC
generativně vegetativně z dormance
0%10%20%30%40%50%60%70%80%90%
100%
H DKI BI DKII DKIII DKIV DKV T1 S1 L1 S2 MC VC
0%10%20%30%40%50%60%70%80%90%
100%
H DKI BI DKII DKIII DKIV DKV T1 S1 L1 S2 MC VC
M. smejkalii M. caespitosa M. corcontica
2008
2009
2010
Graf 2. Poměr nových jedinců vzniklých generativně (bílá barva), vegetativně (černá barva) a obnovených po dormanci (šedá barva) v roce 2008, 2009 a 2010.Graph 2. The proportion of newly appearing individuals of generative origin (white color), vegetative origin (black color) or re-appearing after dormancy (grey color) in 2008, 2009 and 2010.
Změny ve velikosti populací se liší mezi jednotlivými roky a mezi jednotlivými populacemi. U druhu M. caespitosa a M. corcontica je velmi obtížné určit, zda jsou tyto výkyvy způsobeny běžnou oscilací v početnosti či jsou již způsobeny změnami v charakteru stanoviště, protože monitoring probíhá pouze krátkou dobu. Data nicméně naznačují, že populace v Malé Čertově zahrádce je stabilní, zatímco ve Velké Čertově zahrádce dochází k poklesu v počtu
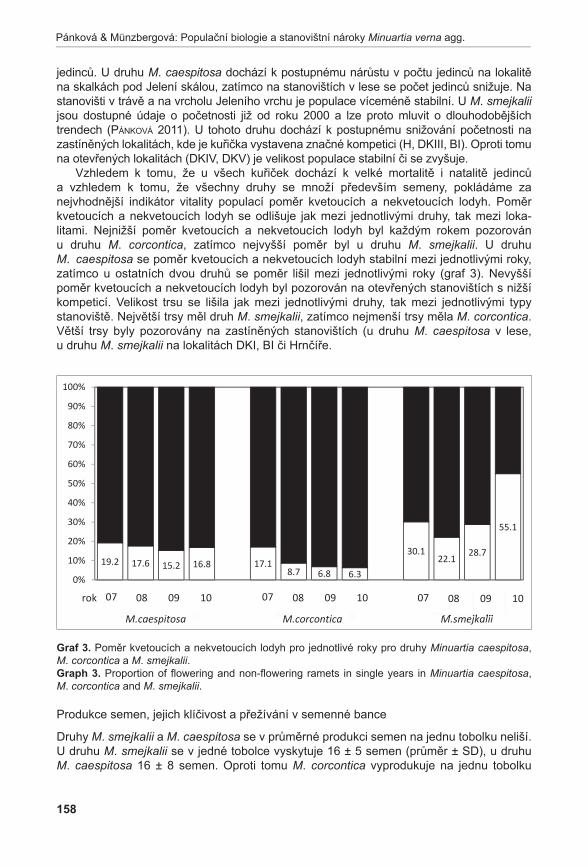
158
Pánková & Münzbergová: Populační biologie a stanovištní nároky Minuartia verna agg.
jedinců. U druhu M. caespitosa dochází k postupnému nárůstu v počtu jedinců na lokalitě na skalkách pod Jelení skálou, zatímco na stanovištích v lese se počet jedinců snižuje. Na stanovišti v trávě a na vrcholu Jeleního vrchu je populace víceméně stabilní. U M. smejkalii jsou dostupné údaje o početnosti již od roku 2000 a lze proto mluvit o dlouhodobějších trendech (Pánková 2011). U tohoto druhu dochází k postupnému snižování početnosti na zastíněných lokalitách, kde je kuřička vystavena značné kompetici (H, DKIII, BI). Oproti tomu na otevřených lokalitách (DKIV, DKV) je velikost populace stabilní či se zvyšuje.
Vzhledem k tomu, že u všech kuřiček dochází k velké mortalitě i natalitě jedinců a vzhledem k tomu, že všechny druhy se množí především semeny, pokládáme za nejvhodnější indikátor vitality populací poměr kvetoucích a nekvetoucích lodyh. Poměr kvetoucích a nekvetoucích lodyh se odlišuje jak mezi jednotlivými druhy, tak mezi lo ka-litami. Nejnižší poměr kvetoucích a nekvetoucích lodyh byl každým rokem pozorován u druhu M. corcontica, zatímco nejvyšší poměr byl u druhu M. smejkalii. U druhu M. caes pitosa se poměr kvetoucích a nekvetoucích lodyh stabilní mezi jednotlivými roky, zatímco u ostatních dvou druhů se poměr lišil mezi jednotlivými roky (graf 3). Nevyšší poměr kvetoucích a nekvetoucích lodyh byl pozorován na otevřených stanovištích s nižší kompeticí. Velikost trsu se lišila jak mezi jednotlivými druhy, tak mezi jednotlivými typy stanoviště. Největší trsy měl druh M. smejkalii, zatímco nejmenší trsy měla M. corcontica. Větší trsy byly pozorovány na zastíněných stanovištích (u druhu M. caespitosa v lese, u druhu M. smejkalii na lokalitách DKI, BI či Hrnčíře.
Graf 3. Poměr kvetoucích a nekvetoucích lodyh pro jednotlivé roky pro druhy Minuartia caespitosa, M.concortica a M.smejkalii.
19.2 17.6 15.2 16.8 17.1 8.7 6.8 6.3
30.1 22.1 28.7
55.1
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
07 08 09 10 07 08 09 10
M.caespitosa M.corcontica M.smejkalii
07
M.smejkaliiM.smejkalii
08
M.smejkaliiM.smejkalii
09 10 rok
Graf 3. Poměr kvetoucích a nekvetoucích lodyh pro jednotlivé roky pro druhy Minuartia caespitosa, M. corcontica a M. smejkalii.Graph 3. Proportion of fl owering and non-fl owering ramets in single years in Minuartia caespitosa,M. corcontica and M. smejkalii.
Produkce semen, jejich klíčivost a přežívání v semenné bance
Druhy M. smejkalii a M. caespitosa se v průměrné produkci semen na jednu tobolku neliší. U druhu M. smejkalii se v jedné tobolce vyskytuje 16 ± 5 semen (průměr ± SD), u druhu M. caespitosa 16 ± 8 semen. Oproti tomu M. corcontica vyprodukuje na jednu tobolku

159
Příroda, Praha, 31: 149–165, 2011
v průměru 12 ± 4 semena. M. smejkalii i M. caespitosa mají i průměrný stejný počet květů na jednu květonosnou lodyhu (4 květy ± 2 květy ), M. corcontica má květy pouze 3 ± 1. Všechny druhy se ale velmi liší v počtu kvetoucích lodyh a tudíž v celkové produkci semen na jeden trs. U druhu M. smejkalii mají kvetoucí trsy v průměru 24 ± 37 kvetoucích lodyh, což znamená, že jeden kvetoucí trs je schopen vyprodukovat více než 1500 semen za vegetační sezónu. Druh M. caespitosa má v průměru 8 ± 10 kvetoucích lodyh, což vede k produkci 500 ± 160 semen na jeden trs, což je 3× méně než u předchozího druhu. Nejnižší produkci semen má však druh M. corcontica, která při počtu 9 ± 10 kvetoucích lodyh na jeden trs vyprodukuje 300 ± 40 semen za vegetační sezónu.
Klíčivost semen na Petriho miskách byla nejvyšší u druhu M. corcontica a dosáhla téměř 91 %. Naopak nejnižší (53 %) byla u druhu M. caespitosa. Porovnání mezi jednotlivými typy stanoviště nebylo možné provést, protože jedinci z lesního a travinného stanoviště měli velmi nízký počet tobolek se zralými semeny. U druhu M. smejkalii dosáhla klíčivost semen na Petriho miskách 73 %, na zahradním substrátu s pískem ale pouze 46 %, mezi jednotlivými populacemi nebyl pozorován prokazatelný rozdíl. Velmi překvapivé bylo, že přestože M. smejkalii má vysokou klíčivost, v terénu nebyl na výsevových plochách nalezen ani jeden semenáček. U ostatních druhů kuřiček se z důvodu omezeného množství semen terénní výsevy neprováděly.
Jednotlivé druhy se od sebe odlišují v tvorbě semenné banky. Klíčivost semen uložených po dva roky v půdě byla pro druh M. smejkalii 80 %, což naznačuje, že tento druh vytváří vytrvalou semennou banku. Na druhou stranu klíčivost semen po jednom roce v půdě u druhu M. corcontica dosáhla pouze 1 %, takže tento druh vytrvalou semennou banku nevytváří. U druhu M. caespitosa dosáhla klíčivost semen po jednom roce v půdě 23 %.
stanovení ekologických nároků druhů
Chemické rozbory půdy ukázaly, že všechny tři druhy rostou na živinami chudých stanovištích. Minuartia smejkalii roste na hadcích, takže její půda má vysoký obsah hořčíku a niklu (tab. 1). Minuartia corcontica se vyskytuje v Obřím dole na erlánech, což jsou pyroxenové ruly vzniklé z karbonátových hornin a vysokým obsahem rudných minerálů. Proto byl v půdě nalezen vysoký obsah těžkých kovů, zejména kadmia, zinku, olova a mědi. Minuartia caespitosa osidluje vápnité pískovce, které jsou charakteristické extrémně nízkým obsahem živin a hořčíku.
tabulka 1. Chemické složení půd na lokalitách druhu Minuartia caespitosa, M. corcontica a M. smejkalii.table 1. Chemical composition of soils on localities of Minuartia caespitosa, M. corcontica andM. smejkalii.
druhpH H2O
pH KCl
N Ccelk. C karb. Corg. Ca Mg K P Zn Cu Pb Ni Co Cr CdC/N
% mg/1000g
Minuartia caespitosa
6,71 6,49 0,20 3,77 0,01 3,76 1219,29 16,84 21,52 6,09 3,14 1,50 5,67 0,13 0,10 0,10 0,09 19,303
Minuartia corcontica
6,63 6,29 0,79 11,25 0,00 11,21 3722,22 896,25 91,70 8,21 286,84 52,60 405,73 16,66 0,16 0,10 6,69 14,200
Minuartia smejkalii
6,16 5,70 0,64 10,83 0,00 10,92 892,87 3348,18 116,97 9,78 9,11 2,61 11,92 87,16 1,46 0,10 0,37 17,006
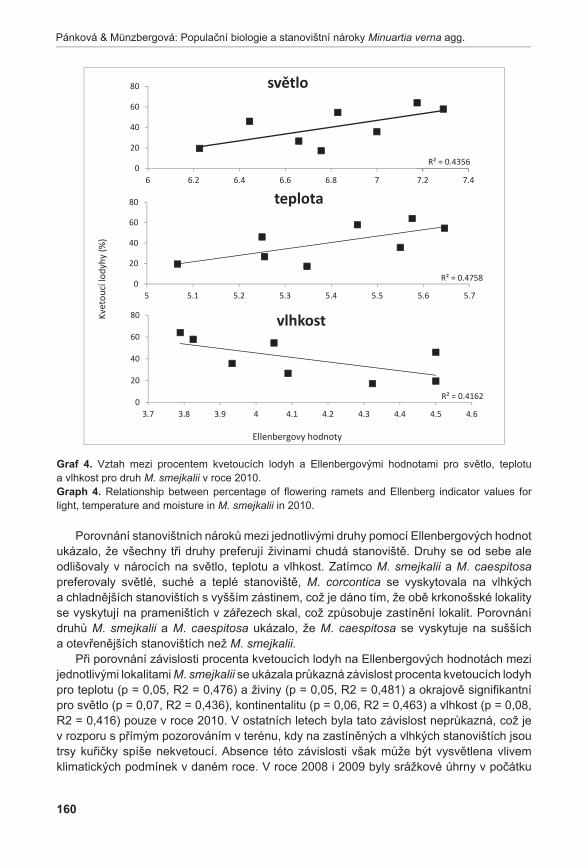
160
Pánková & Münzbergová: Populační biologie a stanovištní nároky Minuartia verna agg.
Graf 4. Vztah mezi procentem kvetoucích lodyh a Ellenbergovými hodnotami pro světlo, teplotu a vlhkost pro druh M. smejkalii v roce 2010.
0
20
40
60
80
6 6.2 6.4 6.6 6.8 7 7.2 7.4
světlo
R² = 0.4758 0
20
40
60
80
5 5.1 5.2 5.3 5.4 5.5 5.6 5.7
teplota
R² = 0.4162 0
20
40
60
80
3.7 3.8 3.9 4 4.1 4.2 4.3 4.4 4.5 4.6
vlhkost
R² = 0.4356
P *Zadejte citát z dokumentu nebo souhrn zajímavého bodu. Textové pole lze umístit kdekoli v dokumentu. Formátování textového pole citátu z vlastního textu lze změnit pomocí karty Nástroje kreslení.+
Kvet
oucí
lody
hy (%
)
Ellenbergovy hodnoty
Graf 4. Vztah mezi procentem kvetoucích lodyh a Ellenbergovými hodnotami pro světlo, teplotu a vlhkost pro druh M. smejkalii v roce 2010.Graph 4. Relationship between percentage of fl owering ramets and Ellenberg indicator values for light, temperature and moisture in M. smejkalii in 2010.
Porovnání stanovištních nároků mezi jednotlivými druhy pomocí Ellenbergových hodnot ukázalo, že všechny tři druhy preferují živinami chudá stanoviště. Druhy se od sebe ale odlišovaly v nárocích na světlo, teplotu a vlhkost. Zatímco M. smejkalii a M. caespitosa preferovaly světlé, suché a teplé stanoviště, M. corcontica se vyskytovala na vlhkých a chladnějších stanovištích s vyšším zástinem, což je dáno tím, že obě krkonošské lokality se vyskytují na prameništích v zářezech skal, což způsobuje zastínění lokalit. Porovnání druhů M. smejkalii a M. caespitosa ukázalo, že M. caespitosa se vyskytuje na sušších a otevřenějších stanovištích než M. smejkalii.
Při porovnání závislosti procenta kvetoucích lodyh na Ellenbergových hodnotách mezi jednotlivými lokalitami M. smejkalii se ukázala průkazná závislost procenta kvetoucích lodyh pro teplotu (p = 0,05, R2 = 0,476) a živiny (p = 0,05, R2 = 0,481) a okrajově signifi kantní pro světlo (p = 0,07, R2 = 0,436), kontinentalitu (p = 0,06, R2 = 0,463) a vlhkost (p = 0,08, R2 = 0,416) pouze v roce 2010. V ostatních letech byla tato závislost neprůkazná, což je v rozporu s přímým pozorováním v terénu, kdy na zastíněných a vlhkých stanovištích jsou trsy kuřičky spíše nekvetoucí. Absence této závislosti však může být vysvětlena vlivem klimatických podmínek v daném roce. V roce 2008 i 2009 byly srážkové úhrny v počátku
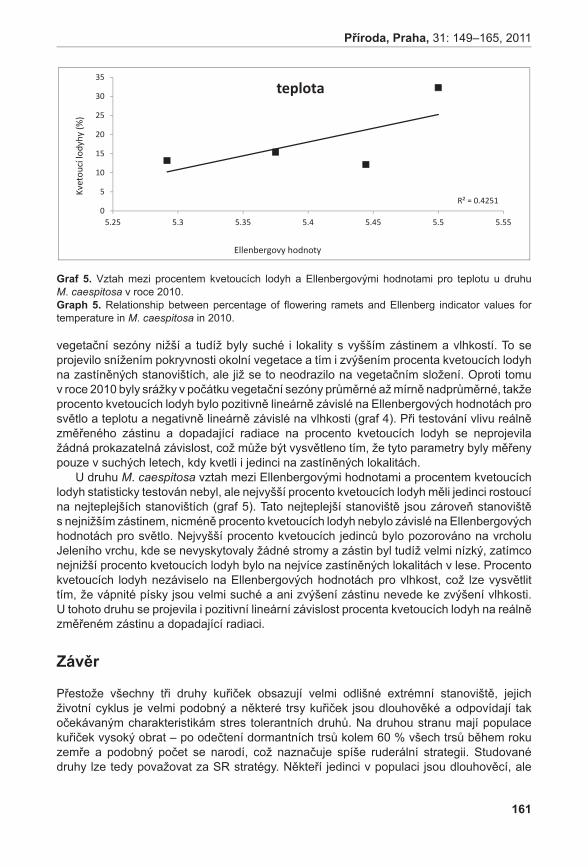
161
Příroda, Praha, 31: 149–165, 2011
Graf 5. Vztah mezi procentem kvetoucích lodyh a Ellenbergovými hodnotami pro teplotu u druhu M. caespitosa v roce 2010.
Kvet
oucí
lody
hy (%
)
Ellenbergovy hodnoty
R² = 0.4251 0
5
10
15
20
25
30
35
5.25 5.3 5.35 5.4 5.45 5.5 5.55
teplota
Graf 5. Vztah mezi procentem kvetoucích lodyh a Ellenbergovými hodnotami pro teplotu u druhu M. caespitosa v roce 2010.Graph 5. Relationship between percentage of fl owering ramets and Ellenberg indicator values for temperature in M. caespitosa in 2010.
vegetační sezóny nižší a tudíž byly suché i lokality s vyšším zástinem a vlhkostí. To se projevilo snížením pokryvnosti okolní vegetace a tím i zvýšením procenta kvetoucích lodyh na zastíněných stanovištích, ale již se to neodrazilo na vegetačním složení. Oproti tomu v roce 2010 byly srážky v počátku vegetační sezóny průměrné až mírně nadprůměrné, takže procento kvetoucích lodyh bylo pozitivně lineárně závislé na Ellenbergových hodnotách pro světlo a teplotu a negativně lineárně závislé na vlhkosti (graf 4). Při testování vlivu reálně změřeného zástinu a dopadající radiace na procento kvetoucích lodyh se neprojevila žádná prokazatelná závislost, což může být vysvětleno tím, že tyto parametry byly měřeny pouze v suchých letech, kdy kvetli i jedinci na zastíněných lokalitách.
U druhu M. caespitosa vztah mezi Ellenbergovými hodnotami a procentem kvetoucích lodyh statisticky testován nebyl, ale nejvyšší procento kvetoucích lodyh měli jedinci rostoucí na nejteplejších stanovištích (graf 5). Tato nejteplejší stanoviště jsou zároveň stanoviště s nejnižším zástinem, nicméně procento kvetoucích lodyh nebylo závislé na Ellenbergových hodnotách pro světlo. Nejvyšší procento kvetoucích jedinců bylo pozorováno na vrcholu Jeleního vrchu, kde se nevyskytovaly žádné stromy a zástin byl tudíž velmi nízký, zatímco nejnižší procento kvetoucích lodyh bylo na nejvíce zastíněných lokalitách v lese. Procento kvetoucích lodyh nezáviselo na Ellenbergových hodnotách pro vlhkost, což lze vysvětlit tím, že vápnité písky jsou velmi suché a ani zvýšení zástinu nevede ke zvýšení vlhkosti. U tohoto druhu se projevila i pozitivní lineární závislost procenta kvetoucích lodyh na reálně změřeném zástinu a dopadající radiaci.
Závěr
Přestože všechny tři druhy kuřiček obsazují velmi odlišné extrémní stanoviště, jejich životní cyklus je velmi podobný a některé trsy kuřiček jsou dlouhověké a odpovídají tak očekávaným charakteristikám stres tolerantních druhů. Na druhou stranu mají populace kuřiček vysoký obrat – po odečtení dormantních trsů kolem 60 % všech trsů během roku zemře a podobný počet se narodí, což naznačuje spíše ruderální strategii. Studované druhy lze tedy považovat za SR stratégy. Někteří jedinci v populaci jsou dlouhověcí, ale

162
Pánková & Münzbergová: Populační biologie a stanovištní nároky Minuartia verna agg.
zároveň mají rostliny vysokou produkci semen, obrat populace je velký a řada jedinců je proto velmi mladá.
Z hodnocených druhů vybočoval svými ekologickými nároky druh M. corcontica díky tomu, že roste v Krkonoších, kde jsou podmínky obecně vlhčí, chladnější. Při hodnocení tohoto druhu je však nutno brát v potaz, že druh osidluje spíše štěrbiny ve skalách než volnou půdu, kde je větší pokryvnost vegetace. Není tedy vyloučeno, že přestože Ellenbergovy hodnoty indikovaly vlhkomilnost, kuřička krkonošská bude preferovat sušší stanoviště v rámci daného území. To dokládá i pokles jak v počtu jedinců, tak v jejich vitalitě na lokalitě Velká Čertova zahrádka poté, co došlo k vytvoření pramene, který začal vyvěrat přímo nad jedním monitorovacím čtvercem.
U kuřičky krkonošské je velmi zajímavé, že přestože poměr kvetoucích lodyh k nekvetoucím je velmi nízký a druh netvoří semennou banku, kuřička krkonošská se rozmnožuje především semeny a klonální růst je zde velmi omezený. To naznačuje, že semena musí být schopna vyklíčit hned první vegetační sezónu a jedinci dorůst do dostatečné velikosti, aby byly schopni přežít zimu. U toho druhu se tedy jeví jako klíčové stádium pro udržení populací dostatečná produkce nových jedinců, aby byly schopny nahradit odumřelé jedince. Vzhledem k tomu, že trsy kuřiček jsou dlouhověké, snížení vitality populací se projeví na počtu jedinců až po delší době. Z tohoto důvodu je třeba sledovat biologické parametry rostlin, zejména však počet kvetoucích lodyh, aby nedošlo ke snížení produkce semen do takové míry, že by již nebyli stávající jedinci schopni vytvářet dostatečné množství nových jedinců. Velmi problematická se v tomto pohledu jeví populace ve Velké Čertově zahrádce, kde dochází jak k poklesu v procentu kvetoucích lodyh, tak ke snížení počtu jedinců. Vzhledem k tomu, že výzkum probíhal pouze ve spodní části lokality a pouze čtyři vegetační sezóny, je velmi obtížné odhadovat, zda tento pokles je způsoben přirozenou fluktuací jak v počtu jedinců, tak v obsazenosti lokality, či zda se jedná o dlouhodobý proces probíhající na celé lokalitě.
Kuřička hercynská je nejvíce vitální na suchých, teplých a osluněných stanovištích, její vitalita klesá s rostoucím zástinem. To dokazuje i to, že nejpříznivější poměr kvetoucích lodyh se vyskytoval na otevřené skále na vrcholu Jeleního vrchu, který není téměř zastíněn, zatímco nejméně kvetoucí jedinci se vyskytovali v podrostu borového lesa. Negativní vliv zástinu na populace kuřičky hercynské se projevil i poklesem ve velikosti populace na stanovišti v lese. I tento druh se rozmnožuje převáženě semeny, nicméně u něj bylo pozorováno vegetativní rozmnožování rozpadem velkých trsů na několik menších. Přestože druh tvoří semennou banku, klíčivost semen po prvním roce v půdě velmi poklesla. Z tohoto důvodu je i pro tento druh důležité udržovat populace ve stavu, kdy budou schopny produkovat dostatečné množství semen, aby nahradily odumřelé jedince. Aby se udržely populace tohoto druhu vitální, je potřeba udržovat jednotlivé lokality otevřené. V současné době je nejvyšší zástin v borovém lese pod oběma lokalitami, který zastiňuje i část písčiny. Na stanovišti v lese je i problematický vývoj semenáčků, které musí prorůstat přes velké množství opadu a spadaných větví.
Z měřených stanovištních nároků pro druh M. smejkalii nelze jednoznačně potvrdit původní předpoklad, že populace na zastíněných lokalitách mají každým rokem horší vitalitu než jedinci rostoucí na otevřených skalách. Vitalita jedinců u tohoto druhu je totiž značně ovlivněna počasím v daném roce. Pokud je na začátku vegetační sezóny velmi sucho, jsou schopny kvést a produkovat semena i jedinci rostoucí na zastíněných lokalitách. Naopak, pokud je rok deštivý, dochází na těchto zastíněných lokalitách k rozvoji mechu a okolní vegetace, takže kuřičky jsou velmi potlačeny. V dlouhodobém měřítku lze tedy očekávat, že populace vyskytující se na otevřených stanovištích by měly být vitálnější. V současné době jsou zástinem a kompeticí nejvíce ohroženy populace Dolnokralovické hadce III,

163
Příroda, Praha, 31: 149–165, 2011
Bernartice I a Hadce u Hrnčíř, což jsou všechno lokality vyskytující se na sekundárních stanovištích, či v kulturním borovém lese. Na těchto lokalitách již byly zahájeny částečné managementové zásahy, které by měly směřovat ke snížení zástinu a kompetice.
U druhu M. smejkalii byla zjištěna největší produkce semen, které mají taktéž velmi dobrou klíčivost. Velmi překvapivé bylo zjištění, že klíčivost semen v semenné bance je vyšší než klíčivost kontrolních semen vysetých ihned po sebrání. To naznačuje, že některá semena kuřičky hadcové jsou sice schopna klíčit okamžitě po vysemenění, ale některá potřebují ke stimulaci růstu určitý čas strávit v půdě. Bohužel není možné určit, zda je to z důvodu zimní stratifikace či narušení osemení. Dalším vysvětlením by mohlo být, že semena potřebují po určitou dobu být v temném prostředí a jakmile se vlivem rozsáhlejší disturbance dostanou na povrch, začnou okamžitě klíčit. Toto se projevilo v průběhu výzkumu na dvou lokalitách. Na podzim roku 2008 byla vytvořena velká obnažená plocha na lokalitě Bernartice I po úklidu černé skládky a v následujícím roce se na této ploše objevily čtyři nové trsy kuřičky hadcové. Obdobně byla okrajová část lokality Hadce u Hrnčíř na podzim roku 2010 značně narušena těžbou dřeva, ale na jaře 2011 se na těchto otevřených plochách objevilo velmi mnoho nových trsů kuřiček. Velmi překvapivé bylo, že neklíčila semena kuřičky hadcové vysetá přímo v terénu, i když byla vyseta do disturbovaných ploch. Vysvětlením by mohlo být, že se pro tyto výsevy použilo pouze 50 semen, zatímco průměrná produkce na trs je několikanásobně větší.
summary
There are three species from Minuartia verna agg. growing in the Czech Republic: Minuartia smejkalii, M. corcontica, and M. caespitosa. All the species are considered as critically endangered, M. corcontica and M. smejkalii are Czech endemics. All these species occur in extreme habitats. In this study we tested whether these species behave as typical S strategists or whether they have some traits characteristic for R strategists. We studied the population dynamics and the habitat preference of these species.
Although all three Minuartia species occur in diverse extreme habitats, their life-cycle is similar. Some of their tussocks are long-lived bunches, which is typical of stress tolerant species. On the other hand, populations of Minuartia species have a very high turnover. In spite of the occurrence of some long-lived tussocks, approximately 60% of all tussocks die and a similar amount of new tussocks appear each year. This is typical behaviour for R strategists. Therefore, our studied species could be considered as SR strategists. Some individuals in each population are long-lived but they also have high seed production, high turn-over of populations and therefore a lot of individuals are very young.
Minuartia corcontica has different ecological requirements than the other two plant species as it occurs in the Krkonoše Mts., where the climate is moister and colder than in other sites. This species, however, occurs more often in rock crevices than in soils with high competition. Although the Ellenbergs value indicated that this species is hygrophilous, M. corcontica prefers drier habitats in the range of the Krkonoše Mts. This is supported by a decreasing number of individuals and their vitality in the population on Velká Čertova zahrádka locality, which was a consequence a new water spring that had appeared on this site. Surprisingly, although this species has very low ratio of flowering stems to non-flowering stems and does not establish a permanent seed bank, it was proven to propagate mainly by seeds and to have a very low clonal growth. This finding suggests that the seeds have to germinate immediately in the first vegetation season and the seedlings grow up to a sufficient size that allows them to survive the winter period. Therefore,

164
Pánková & Münzbergová: Populační biologie a stanovištní nároky Minuartia verna agg.
seed production and establishment of new plants is a critical transition of the life cycle of this species. As the tussocks are long-lived, the negative impact of a decrease in the population vitality on the number of individuals is visible after a longer period. For this reason, it is necessary to monitor the biological parameters of this species, especially the number of flowering stems. A decrease in the number of flowering stems could cause that plants will not be able to compensate the amount of dead individuals. This problem could appear mainly on locality Velká Čertova zahrádka, where we observed in the monitored part of the population a decrease in the number of flowering stems as well as in the number of individuals. However, the research was only conducted in the lowest part of the population and only for four years. Therefore, it is not possible to decide whether this decrease is caused by a really lower vitality of the population or whether is it only natural fluctuation.
Minuartia caespitosa was most vital on dry, warm, and sunny habitats. Its vitality decreased with increasing shading. The highest ratio of flowering stems to non-flowering stems was observed on the open rock on the top of Jelení vrch, while the lowest ratio was observed in plants growing in a pine forest. High shading in the forest caused a decrease in the population size on this site. This species propagates mainly by seeds; nevertheless, it is able to grow clonally by disintegrating large tussocks. Although this species establishes a seed bank, germination of the seeds strongly decreased after one year in the soil. Therefore, similarly to M. corcontica, the seed production and establishment of new plants is a critical stage of the life cycle in this species. To maintain the vitality of the population it is necessary to keep all sites open. The forested site is unsuitable for the species not only due to the shading but also due to a large amount of litterfall (needles and branches).
Negative effects of shading on populations of M. smejkalii could be detected only in certain years. If the early spring was dry, not only plants on open rocks but also plants on shaded sites were able to flower and produced seeds. On the other hand, in rainy years, the shaded sites were covered with moss and other plant species and plants of M. smejkalii are strongly suppressed. In a long time perspective, we can suppose that plants growing on open sites will have higher vitality than plants on shaded sites. The populations recently most endangered with shading and competition are growing on localities Dolnokralovické hadce III, Bernartice I and Hadce u Hrnčíř. These populations occur in secondary habitats or in cultured pine forests. Management on all of these sites started to eliminate the negative aspects of shading and competition. M. smejkalii had the highest seed production of all the studied species. Seeds also had a high germination rate but germination of seeds from the seed bank was even higher. This suggests that some seeds are able to germinate immediately after seeding but other need some time in the soil to stimulate germination. However, it is not possible to distinguish whether it is due to the need of scarification or cold stratification. Alternative explanation could be that the seeds need to be exposed to full light to start to germinate. This was observed on two sites. In 2008, a large open area was created at locality Bernartice I after removing an illegal dump. In the subsequent year, four new seedlings were observed in this open area. Similarly, in 2010, a part of locality Hadce u Hrnčíř was disturbed by forestry operations and many new seedlings appeared there in the following spring. We surprisingly observed no germination in our experimental sowing plots in the field. The explanation could be that we used only 50 seeds, while the mean production of seed per plant is many times higher.

165
Příroda, Praha, 31: 149–165, 2011
Poděkování
Tato práce byla podpořena projektem MŠMT VaV 2B06178 a částečně projekty MŠMT 0021620828 a AV0Z60050516.
literatura
bauMach h. (2005): Genetische Differenzierung mitteleuropäischer Schwermetallsippen von Silene vulgaris, Minuartia verna und Armeria maritima unter Berücksichtigung biogeographischer, montanhistorischer und physiologischer Aspekte. – Ms. [Dizertační pr., depon. in: University of Jena.]
DěDina j. (1987): Selected methods of analytic atom spectrochemistry. – Československá spektroskopická společnost.
Dúbravková D., chytrý M., willner w., illyés e., janiŠová M. & kállayné szerényi j. (2010): Dry grasslands in the Western Carpathians and the northern Pannonian Basin: a numerical classification. – Preslia 82: 165–221.
Dvořáková M. (2003): Minuartia pauciflora, das karpatische Endemit aus der M. verna-Gruppe. – Preslia 75: 349–356.
Dvořáková M. (1999): Zwei zur Sektion Polymechana gehörende Minuartia-Arten (Caryophyllaceae). – Preslia 70: 335–338.
Dvořáková M. (1988): Minuartia smejkalii, eine neue Art aus der Minuartia gerardii-Gruppe. – Preslia 60: 1–9.
Dvořáková M. (1985): Minuartia glaucina, eine neue Art aus der Minuartia verna-Gruppe. – Preslia 57: 1–8.
ehrenberGer f. & Gorbach s. (1973): Methoden der organischen Elementar- und Spurenanalyse. – Verlag Chemie, Berlin.
ellenberG h., weber h. e., Düll r., wirth v. & werner w. (2001): Zeigerwerte von Pflanzen in Mitteleuropa. 3. Auflage. – Scr. Geobot. 18: 1–262.
hejný s. & slavík b. [eds] (1990): Květena České republiky 2. – Academia, Praha.linDsay w. l. & norwell w. a. (1978): Development of a DTPA soil test for zinc, iron, manganese
and copper. – Soil. Sci. Soc. Am. J. 42: 421–428.Moore P. D. & chaPMan s. b. [eds] (1986): Methods in Plant Ecology. – Blackwell Scientific
Publications, Oxford.neuMann D., zur nieDen u., schwieGer w., leoPolD i. & lichtenberGer o. (1997): Heavy metal
tolerance of Minuartia verna. – J. Plant Physiol. 151: 101–108.olsen s. r., cole c. v., watanabe f. s. & Dean l. a. (1954): Estimation of available phosphorus
in soils by extraction with sodium bicarbonate. – USDA Circular 939: 1–19.olsen s. r. & soMMers l. e. (1982): Phosphorus. – In: PaGe a. l. [ed.], Methods of soil analysis:
Part 2. Chemical and microbiological properties. Agron. Monogr. 9, 2nd ed., pp. 403-430, ASA and SSSA, Madison WI.
Pánková h. (2011): Kuřička hadcová Minuartia smejkalii. Péče o druh a jeho lokality. – Agentura ochrany přírody a krajiny České republiky, Praha.
Procházka f. [ed.] (2001): Černý a červený seznam cévnatých rostlin České republiky (stav v roce 2000). – Příroda, Praha, 18: 1–166.
sáDlo J., Petřík P., boublík k., rychtařík P. & ŠíMová i. (2011): Diverzita rostlinstva Hradčanských stěn (Dokesko) a její příčiny. – Zpr. Čes. Bot. Společ. 46: 17–38.
slavíková j. (1986): Ekologie rostlin. – Praha.tichý l. (2002): JUICE, software for vegetation classification. – J. Veg. Sci. 13: 451–453. zbíral j. (1996): Analýza půd II. Jednotné analytické postupy. – Ústřední kontrolní a zkušební
ústav zemědělský, Brno.

166
Pánková & Münzbergová: Populační biologie a stanovištní nároky Minuartia verna agg.

167
Příroda, Praha, 31: 167–183, 2011
Monitoring vstavače trojzubého (Orchis tridentata) v národní přírodní rezervaci strabišov-oulehla v období 1997–2010
Monitoring of the three-toothed orchid (Orchis tridentata) in national nature Reserve strabišov-oulehla over the period 1997–2010
sLavoMír dostaLík1, vLastiMiL rybka1, roMana ZeMánková-bartková& MiLan baLáž2
1Sagittaria – sdružení pro ochranu přírody střední Moravy, Lipové náměstí 3, CZ-783 36 Křelov; [email protected]Ústav experimentální biologie, Přírodovědecká fakulta Masarykovy univerzity, Kotlářská 2, CZ-611 37 Brno; [email protected] (corresponding author)
Abstract: Orchis tridentata (syn. Neotinea tridentata) is a tuberous terrestrial orchid with the centre of its distribution in the Mediterranean region. In the Czech Republic, where seven recent localities occur, this species is treated as critically endangered. The most important site where more than 90% of the entire Czech population of this species can be found is National Nature Reserve Strabišov-Oulehla. Since the end of 80’s of 20th century, the annual management consists of elimination of woody plants and mosaic mowing after capsules ripening, followed by biomass removal. The local population of O. tridentata has been monitored since 1997 in order to get a set of basic morphometric data, to quantify interseasonal variation of these plant characteristics and to evaluate the efficiency of the management on the stability and prosperity of evaluated O. tridentata population. We observed a marked increase in the number of plants, reaching its maximum in 2002 (695 plants). In the period 2003–2009, the lowest number of plants was 463 and the mean was 552 plants. There were high interseasonal fluctuations in the number of fertile plants (coefficient of variation calculated from annual means CV = 51.3%) and in the percentage of flowers producing capsules (CV = 57.4%), mainly due to low values observed in 2000, 2003, and 2007. These were caused by dry weather during spring months, eventually combined with spring frosts in 2007, which damaged the orchids substantially. The weather also influenced the plant height and diameter of leaf rosettes, whereas the number of flowers was not affected and its interseasonal variation was low (CV = 7.5%). Based on our data, we conclude that the condition of this O. tridentata population is very good and we recommend retaining present management practices. We suggest that the future monitoring focuses on the number of flowering plants and the proportion of flowers producing capsules in individual plants.Keywords: morphometrics, Orchidaceae, Neotinea tridentata, population biology, site management, species conservation
Abstrakt: Orchis tridentata (syn. Neotinea tridentata) je hlíznatá zemní orchidej s centrem rozšíření v mediteránních oblastech. Tento druh je v České republice chráněn v kategorii kriticky ohrožených druhů a ze sedmi recentních lokalit má zdaleka nejpočetnější populaci (více jak 90 % celé v ČR rostoucí populace) v Národní přírodní rezervaci (NPR) Strabi-šov-Ou lehla. V této rezervaci je od konce osmdesátých let 20. století každoročně aplikován ma nagement, který zpočátku spočíval zejména v redukci náletových dřevin, později pak v pravidelném mozaikovitém sečení ploch, realizovaném vždy po uzrání tobolek O. tridentata, s následným odnosem biomasy mimo NPR. Od roku 1997 je na této lokalitě

168
Dostalík et al.: Monitoring Orchis tridentata v Národní přírodní rezervaci Strabišov-Oulehla
realizován monitoring, jehož cílem bylo nejen získat základní morfometrická data o místní populaci tohoto druhu, ale rovněž blíže poznat mezisezónní fluktuace v těchto znacích a ohodnotit, nakolik aplikovaný management lokality zabezpečuje stabilitu a prosperitu místní populace O. tridentata. V období 1997 až 2002 byl pozorován výrazný nárůst počtu rostlin, který dosáhl maxima v roce 2002 (695 rostlin). V období 2003–2009 celkový počet rostlin nikdy neklesl pod 463 a v průměru činil 552 rostliny. Výraznou meziroční fluktuací se vyznačoval počet plodících rostlin (variační koeficient vypočtený z ročních průměrných hodnot CV = 51,3 %) a procentuální podíl počtu fertilních tobolek z celkového počtu květů jednotlivých rostlin (CV = 57,4 %), a to zejména kvůli nízkým hodnotám naměřeným v letech 2000, 2003 a 2007. Ty byly způsobeny suchým počasím v průběhu jarních měsíců, v r. 2007 se navíc vyskytly pozdní jarní mrazy, které rostliny výrazně poškodily. Vliv počasí bylo možné vysledovat i na hodnotách výšky kvetoucích rostlin a průměru listových růžic, zatímco průměrný počet květů počasím výrazně ovlivněn nebyl a meziročně kolísal velmi málo (CV = 7,5 %). Na základě naměřených dat vyvozujeme, že stav místní populace O. tridentata je velmi dobrý a doporučujeme zachovat stávající způsob péče o tuto lokalitu. Z hlediska monitoringu navrhujeme snížit počet sledovaných znaků na pouhé dva, počet kvetoucích rostlin a procentuální podíl počtu fertilních tobolek z celkového počtu květů jednotlivých rostlin.Klíčová slova: druhová ochrana, management lokalit, morfometrie, Orchidaceae, Neotinea tridentata, populační biologie
Úvod
Čeleď vstavačovité (Orchidaceae) je se svými přibližně 25 tisíci druhy jednou z největších rostlinných čeledí. Orchideje se vyznačují řadou biologických zvláštností, které jsou pravděpodobně evolučně propojeny. Zřejmě nejvýznačnější z nich je tvorba tzv. orchi-deoidní mykorhizy, tedy parazitické až mutualistické symbiózy, která zabezpečuje všem klíčícím orchidejím energeticky bohaté látky a dospělým rostlinám většiny druhů orchidejí pak zvýšený příjem minerálních živin (sMith & reaD 2008). Orchideoidní mykorhizní symbióza je zcela typická i pro dospělé terestrické orchideje rostoucí v České republice (GrynDler et al. 2004). Mnohé v ČR rostoucí orchideje jsou přitom velmi vzácné a pokles počtu jejich lokalit byl v minulém století velmi dramatický (jatiová & ŠMiták 1996). Tento pokles byl téměř bez výjimky způsoben činností člověka a souvisí zejména s přímým úbytkem ploch vhodných pro růst těchto rostlin, zapříčiněným zejména intenzifikací zemědělské výroby a acidifikací a eutrofizací způsobenou plynnými emisemi. Bohužel, dosud přesně neznáme mechanismy způsobující úbytek počtu lokalit a/nebo redukci populací orchidejí na lokalitách. A tak rozhodnout, zda-li se v případě negativního efektu eutrofizace jedná o přímé negativní působení, poškozující fyziologicky buď orchideje jako takové, popř. jejich mykorhizní symbiózu, nebo zda se jedná o nepřímý vliv zprostředkovaný zvýšenou kompeticí o zdroje (zejména o vodu či světlo), způsobenou akcelerací růstu zejména vysokostébelných trav, není dosud možné (beyrle et al. 1991, Dijk & olff 1994, silvertown et al. 1994, Dijk & eck 1995, wotavová et al. 2004, hejcMan et al. 2010).
Přes naši neznalost těchto mechanismů bylo v minulosti opakovaně prokázáno, že na přežívání a prosperitu populací orchidejí má vysoce pozitivní vliv pravidelné sečení, realizované ve vhodně zvoleném termínu a následované odnosem sklizené biomasy (waite & hutchinGs 1991, wotavová et al. 2004, janeČková et al. 2006). Tento způsob managementu byl od přelomu osmdesátých a devadesátých let 20. století aplikován i na nejvýznamnější lokalitě druhu Orchis tridentata v ČR, v Národní přírodní rezervaci Strabišov-Oulehla. Na

169
Příroda, Praha, 31: 167–183, 2011
téže lokalitě bylo v roce 1997 započato s intenzivní monitoringem tohoto druhu orchideje s cílem i) přinést základní morfometrická data tohoto druhu, ii) kvantifikovat mezisezónní variabilitu těchto znaků a iii) zejména vyhodnotit, zda zvolený způsob péče o lokalitu je vhodný a zabezpečuje prosperitu místní populace O. tridentata.
obr. 1. Geneta Orchis tridentata rostoucí v NPR Strabišov-Oulehla. Foto: S. Dostalík, 14. 5. 2008.Fig. 1. Genet of Orchis tridentata growing in NNR Strabišov-Oulehla. Photo: S. Dostalík, 14 May 2008.

170
Dostalík et al.: Monitoring Orchis tridentata v Národní přírodní rezervaci Strabišov-Oulehla
Materiál a metody
charakteristika zájmového druhu
Vstavač trojzubý – Orchis tridentata Scop. (obr. 1), dle nejnovějších prací označovaný jako Neotinea tridentata (Scop.) R. M. Bateman, A. M. Pridgeon & M. W. Chase (kretzschMar et al. 2007) – je orchidej s centrem rozšíření v mediteránních oblastech (Iberský poloostrov, Itálie, Sardinie, Korsika, Sicílie, Malta, státy bývalé Jugoslávie, Řecko, Kréta, Turecko a mediteránní oblast severní Afriky). Směrem na východ zasahuje areál až po Krym, Arménii, Izrael, Sýrii, Jordánsko a Libanon. Ve střední Evropě má O. tridentata značně disjunktní areál rozšíření. V České republice je jeho současný výskyt pouhým fragmentem z doložených 28 historických moravských lokalit (jatiová & ŠMiták 1996). V současnosti je známo pouze sedm lokalit tohoto druhu, z toho v Národní přírodní rezervaci (NPR) Strabišov-Oulehla roste více jak 90 % celé české populace tohoto druhu. Biotopem vstavače trojzubého jsou xerotermní až semixerotermní travinobylinná společenstva hlubších minerálně silných půd teplých oblastí s alkalickou až neutrální půdní reakcí. Jedná se o striktní heliofyt a termofyt, který nesnáší zástin a vyžaduje stanoviště jižně orientovaných svahů nebo výhřevných vápnitých substrátů. V České republice je chráněn v kategorii kriticky ohrožený, pod stejným stupněm ohrožení je uveden i v červeném seznamu cévnatých rostlin ČR (holub & Procházka 2000).
charakteristika lokality
Národní přírodní rezervace Strabišov-Oulehla se nachází v Litenčické pahorkatině ve Zlínském kraji, v katastrálním území obcí Lísky a Kunkovice u Litenčic, v nadmořské výšce 278–380 m, GPS souřadnice 49º10’16” S a 17º12’41” V. Ochrana tohoto území se datuje od roku 1953, kdy byla vyhlášena Státní přírodní rezervace (SPR) Oulehla, resp. od roku 1956, kdy byla vyhlášena SPR Strabišov. V současnosti je lokalita chráněna taktéž v rámci soustavy Natura 2000 (kód CZ0624069). O. tridentata se vyskytuje v části NPR zvané Oulehla, v širokolistých suchých trávnících svazu Bromion erecti (chytrý 2007), spolu s dalšími druhy orchidejí, střevičníkem pantoflíčkem (Cypripedium calceolus, ca 300 ks), vstavačem vojenským (Orchis militaris, stovky kusů), pětiprstkou žežulníkem (Gymnadenis conopsea, desítky rostlin), vemeníkem dvoulistým (Platanthera bifolia, desítky rostlin) a okroticí bílou (Cephalanthera damasonium, několik málo rostlin; nomenklatura dle bauMann et al. 2009). V jiných typech vegetace lze na území NPR nalézt vstavač nachový (O. purpurea, několik málo kusů), vstavač bledý (O. pallens, ca 50 ks), kruštík růžkatý (Epipactis muelleri, ca deset ks) a velmi vzácně kruštík modrofialový (E. purpurata). Kompletní inventarizační seznam druhů rostlin lze nalézt v pracích ŠeDa (1976) a rybka et al. (2005).
Management na území tehdejší SPR Oulehla byl započat v letech 1978 a 1979, kdy byly na velké části tzv. zadní Oulehly radikálně odstraněny náletové borovice, což se s odstupem času ukázalo jako opatření, díky kterému se nejenže podařilo zachovat cenná travinobylinná společenstva, ale řada druhů včetně O. tridentata se postupně rozšířila i na vykácené plochy, které úspěšně zregenerovaly. Jednotlivé borovice byly odstraňovány i v pozdějších letech, a to i v severním cípu tzv. přední Oulehly. Od přelomu osmdesátých a devadesátých let bylo započato pravidelné kosení na základě ochranářského plánu, později plánu péče. Byl zaveden střídavý režim sečení tak, aby byla každoročně pokosena přibližně polovina obou částí nelesních ploch (obr. 2). Intenzivnější kosení (2× až 3× ročně) je uplatňováno lokálně na místech s výskytem třtiny křovištní (Calamagrostis epigejos),

171
Příroda, Praha, 31: 167–183, 2011
jejíž výskyt (pokryvnost) se podařilo během posledních let výrazně snížit. Úklid posečené biomasy je prováděn pomocí obraceče sena, který zároveň do jisté míry rozrušuje vegetační kryt na lokalitě, čemuž přičítáme pozitivní vliv na možnost klíčení semen O. tridentata. Při sečení se také pravidelně odstraňují nárosty zmlazujících dřevin, zejména krušiny, svídy a borovice.
obr. 2. Ortofotomapa národní přírodní rezervace (NPR) Strabišov-Oulehla. Červeně je vyznačena hranice NPR, zeleně zájmové území, kde probíhal monitoring populace Orchis tridentata. Plochy vyznačené uvnitř zájmového území růžovou barvou ukazují mozaikovitou oblast posečenou v srpnu 2009. GPS souřadnice bodu označeného * jsou 49º10’16,139” S a 17º12’40,685” V.Fig. 2. Orthophotomap of National Nature Reserve (NNR) Strabišov-Oulehla. The border of the NNR is marked by a red line and the area of interest where the population of Orchis tridentata was monitored by a green line. The areas within it which are marked by pink color represent mosaic distribution of areas moved in August 2009. GPS coordinates of the place marked by * are 49º10’16.139” N and 17º12’40.685” E.
172
Dostalík et al.: Monitoring Orchis tridentata v Národní přírodní rezervaci Strabišov-Oulehla
tab. 1. Schéma hodnocení populační dynamiky a morfometrického hodnocení Orchis tridentata v NPR Strabišov-Oulehla.table 1. Scheme of morphometrical and population dynamics assessment of Orchis tridentata in NNR Strabišov-Oulehla.
Rok/year
datum/date
Počet rostlin/number of plants
výška kvetoucích
rostlin/Flowering plant
height (cm)
Počet listů/
number of leaves
Šířka listové růžice/
Width of leaf rosette (cm)
Počet květů/number of
flowers
Počet fertilních tobolek/
number of fertile capsules
1997 17.5.1997
12.6.1997
1998 11.5.1998
21.5.1998
10.10.1998
20.11.1998
1999 30.3.1999
11.5.1999
4.6.1999
23.10.1999
19.11.1999
2000 1.4.2000
1.5.2000
3.6.2000
6.10.2000
22.11.2000
2001 17.4.2001
17.5.2001
1.6.2001
2.10.2001
21.11.2001
2002 19.4.2002
10.5.2002
17.6.2002
2003 15.5.2003
11.6.2003
2004 10.5.2004
16.6.2004
2005 25.5.2005
21.6.2005
2006 18.5.2006
26.6.2006
2007 7.5.2007
20.6.2007
2008 13.5.2008
25.6.2008
2009 11.5.2009
19.6.2009
2010 11.5.2010
24.6.2010

173
Příroda, Praha, 31: 167–183, 2011
Hodnocené znaky a statistické zpracování dat
V návaznosti na záchranný program O. tridentata byl na lokalitě od roku 1997 zaveden pravidelný každoroční monitoring, zajišťovaný sdružením Sagittaria. V jeho rámci byly v průběhu let 1997–2010 hodnoceny počty kvetoucích, plodících a všech ostatních rostlin, jejich výška, počet listů, šířka listové růžice, počet květů a počet fertilních tobolek (tab. 1). Jednotlivé rostliny byly pro účely opakovaných měření mezi lety 1999 až 2002 postupně značeny číslovanými nerezovými štítky, dohledatelnými detektorem kovů. Počet rostlin takto označených činil v roce 1999 – 228, v r. 2000 – 382, v r. 2001 – 590 a v r. 2002 – 695. Toto značení rostlin je využíváno až do současné doby.
Meziroční rozdíly v průměrných hodnotách výšky květního stvolu, šířky listové růžice kvetoucích i nekvetoucích rostlin v době květu, rozdíly v počtu květů a v procentuálním poměru počtu fertilních tobolek k počtu květů v květenství byly hodnoceny jednocestnou analýzou rozptylu. Průměry přízemních listových růžic měřené opakovaně na jednotlivých rostlinách, 4× v průběhu každé vegetační sezóny v letech 1998 až 2002, byly hodnoceny analýzou rozptylu s opakovanými měřeními. Vstupní předpoklad homogenity rozptylů byl hodnocen pomocí grafů závislosti hodnot skupinových směrodatných odchylek na skupinových průměrech, normalita reziduí pomocí grafické shody s normálním rozložením. Procentická data podílu fertilních tobolek z počtu všech květů byla před analýzou arcsin transformována. K identifikaci rozdílů mezi jednotlivými sezónami byl použit post hoc Sheffého test na hladině významnosti 5 %. Párový t-test byl použit pro testování rozdílů výšky kvetoucích rostlin, kdy byla testována data z 17. 5. 2001 vs. 1. 6. 2001 a 10. 5. 2002 vs. 17. 6. 2002. Normalita rozdílů dat byla testována Shapiro-Wilkovým testem. Po ověření linearity vztahu mezi výškou rostliny a počtem květů byl vypočten koeficient determinance R2 (= druhá mocnina Pearsonova korelačního koeficientu). Všechny statistické výpočty byly realizovány s využitím software Statistica 9 (StatSoft Inc., Tulsa, USA).
výsledky
Celkový počet kvetoucích i nekvetoucích rostlin (obr. 3) měl v období 1997 až 2002 výrazně se zvy šující tendenci, s maximem zjištěným v roce 2002 (695 rostlin). V letech 2003–2009 pak zůstával poměrně konstantní, nejvyšší hodnota naměřená v v tomto období činila 680 rostlin (r. 2006), nejnižší pak 463 rostlin (r. 2003), tj. 66,6 % hodnoty maxima z r. 2002. Celkově byl počet všech rostlin oproti počtu rostlin kvetoucích či plodících nejméně kolísajícím znakem, s variačním koeficientem (CV) 31,2 %. Vyšší variabilita byla zjištěna pro počty kvetoucích rostlin (CV = 36,6 %) a nejvyšší pak pro počty plodících rostlin (CV = 51,3 %). Příčinou nejvyšší variability počtu plodících rostlin byly zjevně nízké hodnoty tohoto znaku zjištěné v letech 2000 a zejména 2007. Rovněž výška kvetoucích rostlin (obr. 4) byla v těchto dvou letech nejnižší, v r. 2000 činila v průměru 13,5 cm, v r. 2007 12,5 cm. Nejvyšší průměrná výška kvetoucích rostlin byla zjištěna v r. 2008 (24,5 cm), vyšší jak dvaceticentimetrová byla i v letech 1997 až 1999 a 2002. Variační koeficient vypočtený z ročních průměrných hodnot výšky kvetoucích rostlin činí 19,5 %. Postupný růst rostlin v průběhu kvetení/plození bylo potvrzeno opakovaným měřením výšky stejných rostlin na jaře 2001 a 2002. Zatímco průměrná výška kvetoucích rostlin činila 17. 5. 2001 20,1 cm, k 1. 6. 2001 byla hodnota téhož znaku 21,6 cm a tento rozdíl byl statisticky průkazný (párový t-test, n = 162, t = -12,3, P < 0,001). Analogická situace byla zjištěna o rok později, kdy rozdíl mezi výškou rostlin k 10. 5. 2002 (23,7 cm) a k 17. 6. 2002 (25,0 cm) byl opět statisticky průkazný (párový t-test, n = 371, t = -15,7, P < 0,001).
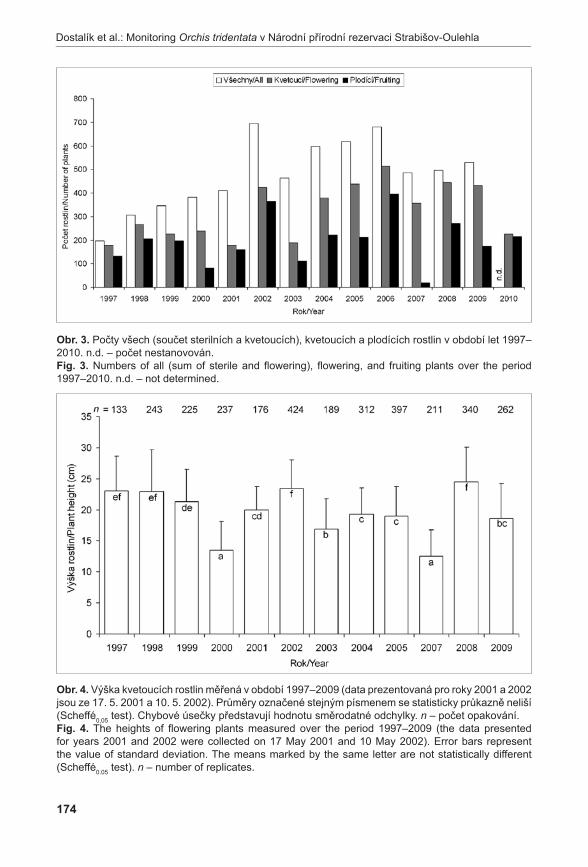
174
Dostalík et al.: Monitoring Orchis tridentata v Národní přírodní rezervaci Strabišov-Oulehla
obr. 3. Počty všech (součet sterilních a kvetoucích), kvetoucích a plodících rostlin v období let 1997–2010. n.d. – počet nestanovován.Fig. 3. Numbers of all (sum of sterile and flowering), flowering, and fruiting plants over the period 1997–2010. n.d. – not determined.
obr. 4. Výška kvetoucích rostlin měřená v období 1997–2009 (data prezentovaná pro roky 2001 a 2002 jsou ze 17. 5. 2001 a 10. 5. 2002). Průměry označené stejným písmenem se statisticky průkazně neliší (Scheffé0,05 test). Chybové úsečky představují hodnotu směrodatné odchylky. n – počet opakování. Fig. 4. The heights of flowering plants measured over the period 1997–2009 (the data presented for years 2001 and 2002 were collected on 17 May 2001 and 10 May 2002). Error bars represent the value of standard deviation. The means marked by the same letter are not statistically different (Scheffé0.05 test). n – number of replicates.
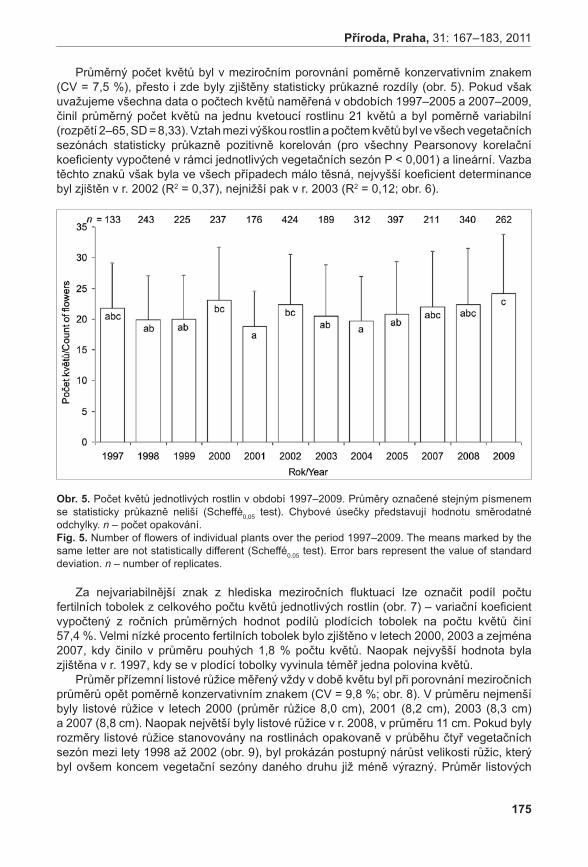
175
Příroda, Praha, 31: 167–183, 2011
Průměrný počet květů byl v meziročním porovnání poměrně konzervativním znakem (CV = 7,5 %), přesto i zde byly zjištěny statisticky průkazné rozdíly (obr. 5). Pokud však uvažujeme všechna data o počtech květů naměřená v obdobích 1997–2005 a 2007–2009, činil průměrný počet květů na jednu kvetoucí rostlinu 21 květů a byl poměrně variabilní (rozpětí 2–65, SD = 8,33). Vztah mezi výškou rostlin a počtem květů byl ve všech vegetačních sezónách statisticky průkazně pozitivně korelován (pro všechny Pearsonovy korelační koeficienty vypočtené v rámci jednotlivých vegetačních sezón P < 0,001) a lineární. Vazba těchto znaků však byla ve všech případech málo těsná, nejvyšší koeficient determinance byl zjištěn v r. 2002 (R2 = 0,37), nejnižší pak v r. 2003 (R2 = 0,12; obr. 6).
obr. 5. Počet květů jednotlivých rostlin v období 1997–2009. Průměry označené stejným písmenem se statisticky průkazně neliší (Scheffé0,05 test). Chybové úsečky představují hodnotu směrodatné odchylky. n – počet opakování.Fig. 5. Number of flowers of individual plants over the period 1997–2009. The means marked by the same letter are not statistically different (Scheffé0.05 test). Error bars represent the value of standard deviation. n – number of replicates.
Za nejvariabilnější znak z hlediska meziročních fluktuací lze označit podíl počtu fertilních tobolek z celkového počtu květů jednotlivých rostlin (obr. 7) – variační koeficient vypočtený z ročních průměrných hodnot podílů plodících tobolek na počtu květů činí 57,4 %. Velmi nízké procento fertilních tobolek bylo zjištěno v letech 2000, 2003 a zejména 2007, kdy činilo v průměru pouhých 1,8 % počtu květů. Naopak nejvyšší hodnota byla zjištěna v r. 1997, kdy se v plodící tobolky vyvinula téměř jedna polovina květů.
Průměr přízemní listové růžice měřený vždy v době květu byl při porovnání meziročních průměrů opět poměrně konzervativním znakem (CV = 9,8 %; obr. 8). V průměru nejmenší byly listové růžice v letech 2000 (průměr růžice 8,0 cm), 2001 (8,2 cm), 2003 (8,3 cm) a 2007 (8,8 cm). Naopak největší byly listové růžice v r. 2008, v průměru 11 cm. Pokud byly rozměry listové růžice stanovovány na rostlinách opakovaně v průběhu čtyř vegetačních sezón mezi lety 1998 až 2002 (obr. 9), byl prokázán postupný nárůst velikosti růžic, který byl ovšem koncem vegetační sezóny daného druhu již méně výrazný. Průměr listových
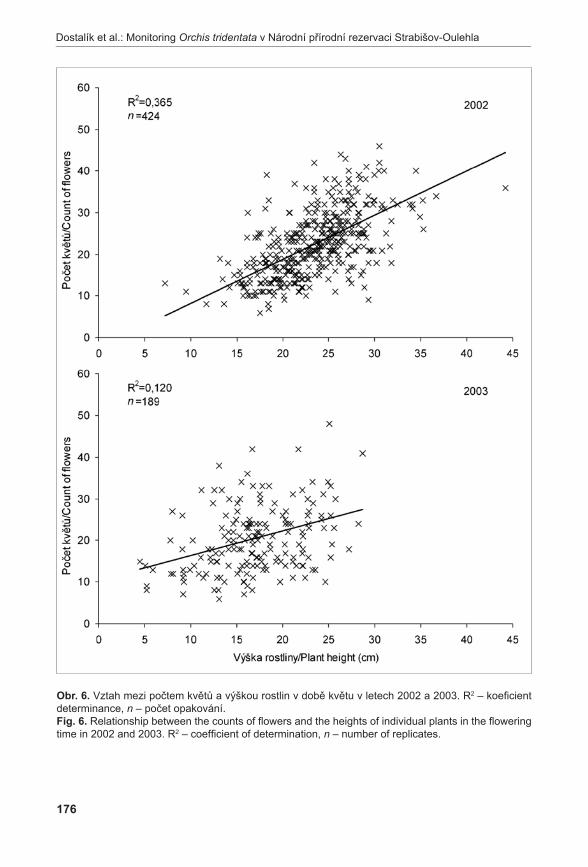
176
Dostalík et al.: Monitoring Orchis tridentata v Národní přírodní rezervaci Strabišov-Oulehla
obr. 6. Vztah mezi počtem květů a výškou rostlin v době květu v letech 2002 a 2003. R2 – koeficient determinance, n – počet opakování.Fig. 6. Relationship between the counts of flowers and the heights of individual plants in the flowering time in 2002 and 2003. R2 – coefficient of determination, n – number of replicates.

177
Příroda, Praha, 31: 167–183, 2011
růžic v měsíci říjnu dosahoval v průměru 4,3 cm (r. 1999) až 5,5 cm (r. 1998), vyjádřeno v procentech celkové maximální naměřené velikosti růžic v dané vegetační sezóně činila velikost (průměr) listové růžice 50–60 %. V průběhu vegetační sezóny docházelo nejen k nárůstu průměru listové růžice, ale rovněž ke zvyšování počtu listů jednotlivých rostlin (obr. 10). Zatímco v říjnovém termínu byly nejčastější rostliny se dvěmi až třemi listy, koncem vegetační sezóny mělo nejvíce rostlin sedm nebo osm listů. Počet listů v období květu druhu (obr. 11) ukazuje, že nejběžnější byly rostliny se šesti až sedmi listy, s výjimkou roku 2001, kdy měly rostliny výrazně méně listů (nejčastěji čtyři).
obr. 7. Procentuální podíl počtu fertilních tobolek z celkového počtu květů jednotlivých rostlin v období 1997–2009. Průměry označené stejným písmenem se statisticky průkazně neliší (Scheffé0,05 test na arcsin transformovaných datech). Chybové úsečky představují hodnotu směrodatné odchylky. n – počet opakování.Fig. 7. The percentage of fertile capsules produced from all flowers in individual plants over the period 1997–2009. Error bars represent the value of standard deviation. The means marked by the same letter are not statistically different (Scheffé0.05 test on arcsin transformed data). n – number of replicates.
diskuse
Vstavač trojzubý (Orchis tridentata) je druh s centrem rozšíření ve středomořské oblasti, vyskytující se velmi vzácně i v ČR. V této studii prezentujeme výsledky dlouhodobého monitoringu, realizovaného na nejvýznamnější české lokalitě tohoto druhu v letech 1997 až 2010. Cílem monitoringu bylo kromě zjištění základních morfometrických dat tohoto druhu zejména vyhodnocení účinnosti prováděného managementu lokality, založeného na mozaikovitém kosení, odnosu posečené biomasy a odstraňování náletových keřů a stromů. Dílčí výsledky monitoringu do roku 2000 lze nalézt v pracích bartková & rybka (1999) a rybka et al. (2001).
Data prezentovaná v tomto článku ukazují na velmi dobrý stav této lokality ve sledovaném období. Mezi lety 1997 až 2002 došlo k výraznému nárůstu populace O. tridentata (obr. 3), která se s jistými mezisezónními fluktuacemi udržuje do současnosti. Podstatný je rovněž
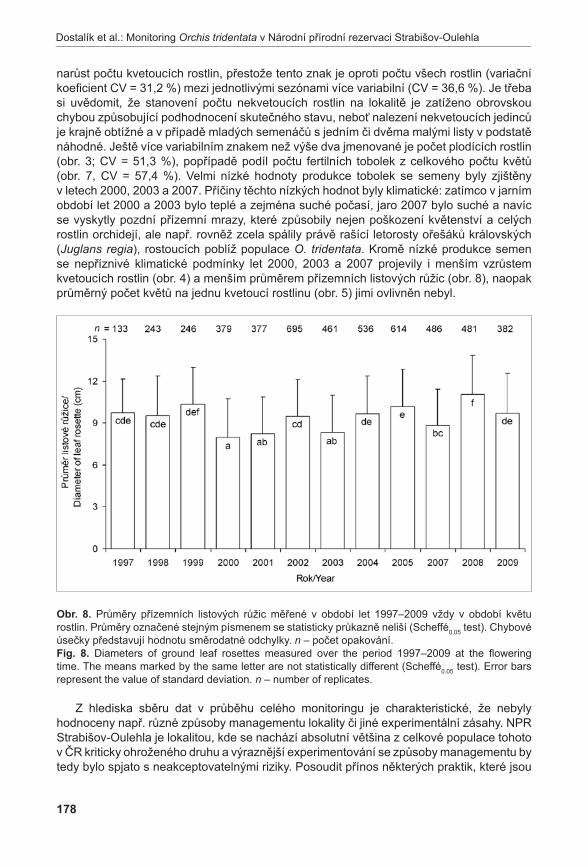
178
Dostalík et al.: Monitoring Orchis tridentata v Národní přírodní rezervaci Strabišov-Oulehla
narůst počtu kvetoucích rostlin, přestože tento znak je oproti počtu všech rostlin (variační koeficient CV = 31,2 %) mezi jednotlivými sezónami více variabilní (CV = 36,6 %). Je třeba si uvědomit, že stanovení počtu nekvetoucích rostlin na lokalitě je zatíženo obrovskou chybou způsobující podhodnocení skutečného stavu, neboť nalezení nekvetoucích jedinců je krajně obtížné a v případě mladých semenáčů s jedním či dvěma malými listy v podstatě náhodné. Ještě více variabilním znakem než výše dva jmenované je počet plodících rostlin (obr. 3; CV = 51,3 %), popřípadě podíl počtu fertilních tobolek z celkového počtu květů (obr. 7, CV = 57,4 %). Velmi nízké hodnoty produkce tobolek se semeny byly zjištěny v letech 2000, 2003 a 2007. Příčiny těchto nízkých hodnot byly klimatické: zatímco v jarním období let 2000 a 2003 bylo teplé a zejména suché počasí, jaro 2007 bylo suché a navíc se vyskytly pozdní přízemní mrazy, které způsobily nejen poškození květenství a celých rostlin orchidejí, ale např. rovněž zcela spálily právě rašící letorosty ořešáků královských (Juglans regia), rostoucích poblíž populace O. tridentata. Kromě nízké produkce semen se nepříznivé klimatické podmínky let 2000, 2003 a 2007 projevily i menším vzrůstem kvetoucích rostlin (obr. 4) a menším průměrem přízemních listových růžic (obr. 8), naopak průměrný počet květů na jednu kvetoucí rostlinu (obr. 5) jimi ovlivněn nebyl.
obr. 8. Průměry přízemních listových růžic měřené v období let 1997–2009 vždy v období květu rostlin. Průměry označené stejným písmenem se statisticky průkazně neliší (Scheffé0,05 test). Chybové úsečky představují hodnotu směrodatné odchylky. n – počet opakování.Fig. 8. Diameters of ground leaf rosettes measured over the period 1997–2009 at the flowering time. The means marked by the same letter are not statistically different (Scheffé0.05 test). Error bars represent the value of standard deviation. n – number of replicates.
Z hlediska sběru dat v průběhu celého monitoringu je charakteristické, že nebyly hodnoceny např. různé způsoby managementu lokality či jiné experimentální zásahy. NPR Strabišov-Oulehla je lokalitou, kde se nachází absolutní většina z celkové populace tohoto v ČR kriticky ohroženého druhu a výraznější experimentování se způsoby managementu by tedy bylo spjato s neakceptovatelnými riziky. Posoudit přínos některých praktik, které jsou
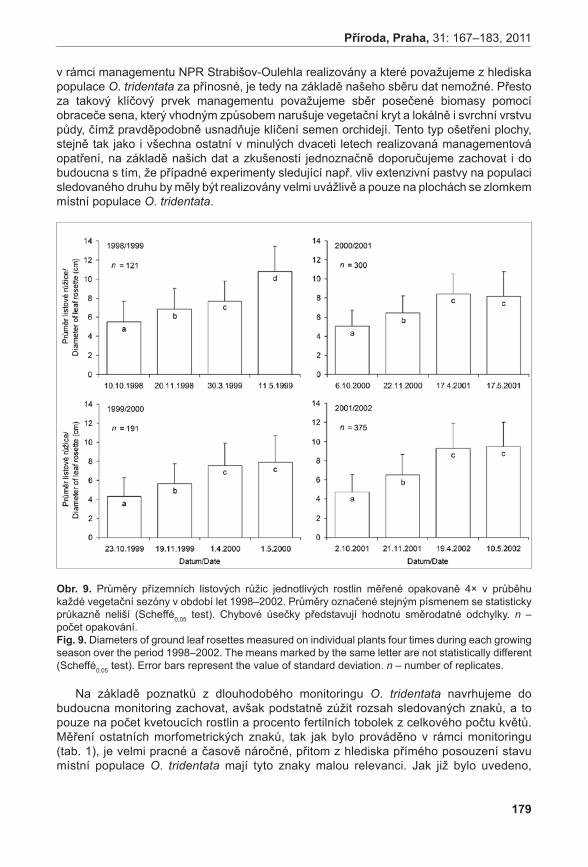
179
Příroda, Praha, 31: 167–183, 2011
v rámci managementu NPR Strabišov-Oulehla realizovány a které považujeme z hlediska populace O. tridentata za přínosné, je tedy na základě našeho sběru dat nemožné. Přesto za takový klíčový prvek managementu považujeme sběr posečené biomasy pomocí obraceče sena, který vhodným způsobem narušuje vegetační kryt a lokálně i svrchní vrstvu půdy, čímž pravděpodobně usnadňuje klíčení semen orchidejí. Tento typ ošetření plochy, stejně tak jako i všechna ostatní v minulých dvaceti letech realizovaná managementová opatření, na základě našich dat a zkušeností jednoznačně doporučujeme zachovat i do budoucna s tím, že případné experimenty sledující např. vliv extenzivní pastvy na populaci sledovaného druhu by měly být realizovány velmi uvážlivě a pouze na plochách se zlomkem místní populace O. tridentata.
obr. 9. Průměry přízemních listových růžic jednotlivých rostlin měřené opakovaně 4× v průběhu každé vegetační sezóny v období let 1998–2002. Průměry označené stejným písmenem se statisticky průkazně neliší (Scheffé0,05 test). Chybové úsečky představují hodnotu směrodatné odchylky. n – počet opakování.Fig. 9. Diameters of ground leaf rosettes measured on individual plants four times during each growing season over the period 1998–2002. The means marked by the same letter are not statistically different (Scheffé0.05 test). Error bars represent the value of standard deviation. n – number of replicates.
Na základě poznatků z dlouhodobého monitoringu O. tridentata navrhujeme do budoucna monitoring zachovat, avšak podstatně zúžit rozsah sledovaných znaků, a to pouze na počet kvetoucích rostlin a procento fertilních tobolek z celkového počtu květů. Měření ostatních morfometrických znaků, tak jak bylo prováděno v rámci monitoringu (tab. 1), je velmi pracné a časově náročné, přitom z hlediska přímého posouzení stavu místní populace O. tridentata mají tyto znaky malou relevanci. Jak již bylo uvedeno,
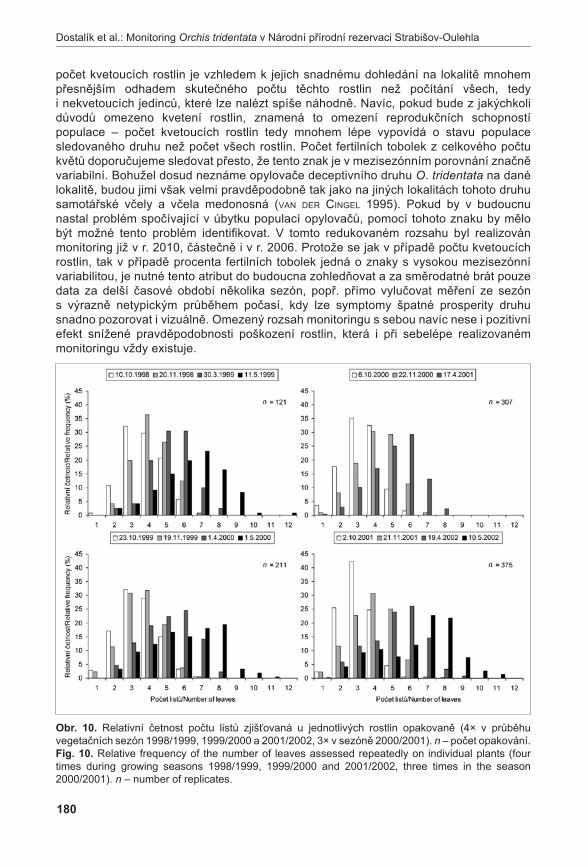
180
Dostalík et al.: Monitoring Orchis tridentata v Národní přírodní rezervaci Strabišov-Oulehla
počet kvetoucích rostlin je vzhledem k jejich snadnému dohledání na lokalitě mnohem přesnějším odhadem skutečného počtu těchto rostlin než počítání všech, tedy i nekvetoucích jedinců, které lze nalézt spíše náhodně. Navíc, pokud bude z jakýchkoli důvodů omezeno kvetení rostlin, znamená to omezení reprodukčních schopností populace – počet kvetoucích rostlin tedy mnohem lépe vypovídá o stavu populace sledovaného druhu než počet všech rostlin. Počet fertilních tobolek z celkového počtu květů doporučujeme sledovat přesto, že tento znak je v mezisezónním porovnání značně variabilní. Bohužel dosud neznáme opylovače deceptivního druhu O. tridentata na dané lokalitě, budou jimi však velmi pravděpodobně tak jako na jiných lokalitách tohoto druhu samotářské včely a včela medonosná (van Der cinGel 1995). Pokud by v budoucnu nastal problém spočívající v úbytku populací opylovačů, pomocí tohoto znaku by mělo být možné tento problém identifikovat. V tomto redukovaném rozsahu byl realizován monitoring již v r. 2010, částečně i v r. 2006. Protože se jak v případě počtu kvetoucích rostlin, tak v případě procenta fertilních tobolek jedná o znaky s vysokou mezisezónní variabilitou, je nutné tento atribut do budoucna zohledňovat a za směrodatné brát pouze data za delší časové období několika sezón, popř. přímo vylučovat měření ze sezón s výrazně netypickým průběhem počasí, kdy lze symptomy špatné prosperity druhu snadno pozorovat i vizuálně. Omezený rozsah monitoringu s sebou navíc nese i pozitivní efekt snížené pravděpodobnosti poškození rostlin, která i při sebelépe realizovaném monitoringu vždy existuje.
obr. 10. Relativní četnost počtu listů zjišťovaná u jednotlivých rostlin opakovaně (4× v průběhu vegetačních sezón 1998/1999, 1999/2000 a 2001/2002, 3× v sezóně 2000/2001). n – počet opakování.Fig. 10. Relative frequency of the number of leaves assessed repeatedly on individual plants (four times during growing seasons 1998/1999, 1999/2000 and 2001/2002, three times in the season 2000/2001). n – number of replicates.
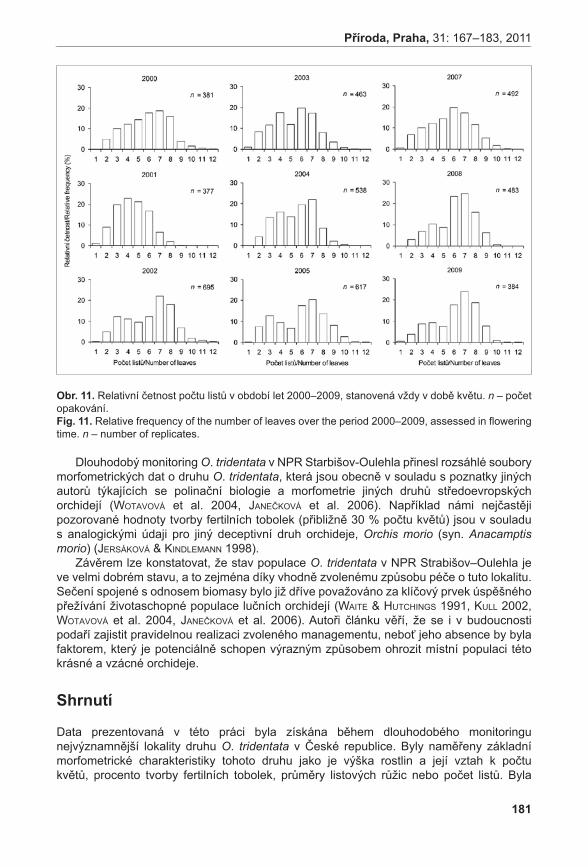
181
Příroda, Praha, 31: 167–183, 2011
obr. 11. Relativní četnost počtu listů v období let 2000–2009, stanovená vždy v době květu. n – počet opakování.Fig. 11. Relative frequency of the number of leaves over the period 2000–2009, assessed in flowering time. n – number of replicates.
Dlouhodobý monitoring O. tridentata v NPR Starbišov-Oulehla přinesl rozsáhlé soubory morfometrických dat o druhu O. tridentata, která jsou obecně v souladu s poznatky jiných autorů týkajících se polinační biologie a morfometrie jiných druhů středoevropských orchidejí (wotavová et al. 2004, janeČková et al. 2006). Například námi nejčastěji pozorované hodnoty tvorby fertilních tobolek (přibližně 30 % počtu květů) jsou v souladu s analogickými údaji pro jiný deceptivní druh orchideje, Orchis morio (syn. Anacamptis morio) (jersáková & kinDleMann 1998).
Závěrem lze konstatovat, že stav populace O. tridentata v NPR Strabišov–Oulehla je ve velmi dobrém stavu, a to zejména díky vhodně zvolenému způsobu péče o tuto lokalitu. Sečení spojené s odnosem biomasy bylo již dříve považováno za klíčový prvek úspěšného přežívání životaschopné populace lučních orchidejí (waite & hutchinGs 1991, kull 2002, wotavová et al. 2004, janeČková et al. 2006). Autoři článku věří, že se i v budoucnosti podaří zajistit pravidelnou realizaci zvoleného managementu, neboť jeho absence by byla faktorem, který je potenciálně schopen výrazným způsobem ohrozit místní populaci této krásné a vzácné orchideje.
shrnutí
Data prezentovaná v této práci byla získána během dlouhodobého monitoringu nejvýznamnější lokality druhu O. tridentata v České republice. Byly naměřeny základní morfometrické charakteristiky tohoto druhu jako je výška rostlin a její vztah k počtu květů, procento tvorby fertilních tobolek, průměry listových růžic nebo počet listů. Byla

182
Dostalík et al.: Monitoring Orchis tridentata v Národní přírodní rezervaci Strabišov-Oulehla
vyhodnocena mezisezónní variabilita těchto znaků a byl identifikován výrazný vliv nepří-znivého průběhu počasí v jarních měsících (sucho, případně mráz) zejména na tvorbu fertilních tobolek a výšku rostlin. Z našich dat vyvozujeme, že management aplikovaný na sledované lokalitě od konce osmdesátých let 20. století, založený na eliminaci náletových dřevin a mozaikovitém kosení následovaném odnosem biomasy je velmi efektivní a místní populace O. tridentata se díky němu nachází ve výborném stavu. Na základě posouzení efektivity sběru dat navrhujeme do budoucna sledování pouhých dvou znaků, a to počtu kvetoucích rostlin a procenta tvorby fertilních tobolek z celkového počtu květů. Počet kvetoucích rostlin je z hlediska posouzení stavu populace mnohem přesnějším kritériem než stanovený počet všech rostlin, neboť zjištění přesného počtu kvetoucích rostlin je zatíženo mnohem menší chybou než zjišťování počtu malých nekvetoucích rostlin, které jsou nalézány spíše náhodně. Navíc, pokud by vitalita místní populace O. tridentata byla z jakéhokoli důvodu snížena, projeví se to nutně mimo jiné právě na těchto dvou parametrech, které navrhujeme pro budoucí monitoring.
summary
The data presented in this study were obtained during long-term monitoring of the most important locality of O. tridentata in the Czech Republic. The basic morphometric characteristics of this species, such as plant height, the number of flowers, percentage of fertile capsules from the number of flowers, diameter of leaf rosette, or the number of leaves were measured. Interseasonal variability of these parameters was evaluated and the effect of unfavorable weather during spring months (drought, frosts) namely on the number of fertile capsules and plant height was identified. Based on our data we conclude that management practices applied on this site since the end of the 80’s of the 20th century, based on elimination of woody species and mosaic mowing followed by biomass removal are highly effective and that the local population of O. tridentata is in an excellent condition. Considering efficiency of data collecting, we propose to monitor only the number of flowering plants and the percentage of fertile capsules from the number of flowers in the future. The number of flowering plants is a more precise criterion when orchid population condition is assessed because the number of all plants is inaccurate as the small non-flowering plants are found rather by chance. Moreover, if the vitality of the local population of O. tridentata decreased for any reason, it would necessarily manifest on the two parameters we suggest for monitoring.
Poděkování
Autoři článku děkují všem, kteří se jakkoli podíleli na monitoringu studovaného druhu a následném zpracování dat či byli jiným způsobem vtaženi do řešené problematiky. Speciální dík patří Radce Říhovské za pomoc v terénu a za pochopení při časově náročném sběru dat. Kromě toho bychom rádi poděkovali sdružení Sagittaria za poskytnutí kvalitního zázemí a všem ostatním nadacím, organizacím a institucím za finanční podporu projektu monitoringu vstavače trojzubého – jmenovitě Ministerstvu životního prostředí ČR (projekty 496/98, 32/99, 393/00, 333/01 a 3473/432/03), Českému svazu ochrany přírody (projekty 120902, 121805 a 03011606) a Ministerstvu školství, mládeže a tělovýchovy (projekt MŠMT VaV 2B06178).

183
Příroda, Praha, 31: 167–183, 2011
literatura
bartková r. & rybka v. (1999): Sledování populací vstavače trojzubého (Orchis tridentata Scop.) na Moravě. – Příroda, Praha, 15: 135–139.
bauMann h., künkele s. & lorenz r. (2009): Orchideje Evropy a přilehlých oblastí. – Academia, Praha.beyrle h., PenninGsfelD f. & hock b. (1991): The role of nitrogen concentration in determining the
outcome of the interaction between Dactylorhiza incarnata (L) Soó and Rhizoctonia sp. – New Phytol. 117: 665–672.
Dijk e. & eck n. (1995): Ammonium toxicity and nitrate response of axenically grown Dactylorhiza incarnata seedlings. – New Phytol. 131: 361–367.
Dijk e. & olff h. (1994): Effects of nitrogen, phosphorus and potassium fertilization on field performance of Dactylorhiza majalis. – Acta Bot. Neerl. 43: 383–392.
GrynDler M., baláž M., hrŠelová h., jansa j. & vosátka M. (2004): Mykorhizní symbióza. O soužití hub s kořeny rostlin. – Academia, Praha.
hejcMan M., schellberG j. & Pavlů v. (2010): Dactylorhiza maculata, Platanthera bifolia and Listera ovata survive N application under P limitation. – Acta Oecol. 36: 684–688.
holub j. & Procházka F. (2000): Red List of the vascular plants of the Czech Republic – 2000. – Preslia 72: 187–230.
chytrý M. [ed.] (2007): Vegetace České republiky. Vol. 1. Travinná a keříčková vegetace. – Academia, Praha.
janeČková P., wotavová k., schöDelbauerová i., jersáková j. & kinDlMann P. (2006): Relative effects of management and environmental conditions on performance and survival of populations of a terrestrial orchid, Dactylorhiza majalis. – Biol. Conserv. 129: 40–49.
jatiová M. & ŠMiták j. (1996): Rozšíření a ochrana orchidejí na Moravě a ve Slezsku. – Agentura ochrany přírody a krajiny, Brno.
jersáková j. & kinDleMann P. (1998): Patterns of pollinator-generated fruit set in Orchis morio (Orchidaceae). – Folia Geobot. 33: 377–390.
kretzschMar h., eccarius w. & Dietrich h. (2007): The orchid genera Anacamptis, Orchis, Neotinea. Phylogeny, taxonomy, morphology, biology, distribution, ecology, hybridisation. – Bürgel, Germany.
kull t. (2002): Population dynamics of north temperate orchids. – In: kull t. & arDitti j. [eds], Orchid biology: Reviews and perspectives, pp. 139–165, VIII. Kluwer Academic Publishers, Dordrecht.
rybka v., krátký M. & Dostalík s. (2005): Botanický inventarizační průzkum národní přírodní rezervace Strabišov-Oulehla. – Ms. [Depon. in: Agentura ochrany přírody a krajiny ČR, středisko Brno.]
rybka v., zeMánková-bartková r. & Dostalík s. (2001): Výzkum a ochrana vstavače trojzubého (Orchis tridentata) v České republice. – Příroda, Praha, 19: 15–26.
silwertown j., wells D. a., GillMan M., DoDD M. e., robertson h. & lakhani k. h. (1994): Short-term effects and long-term after-effects or fertilizer application on the flowering population of green winged orchid Orchis morio. – Biol. Conserv. 69: 191–197.
sMith s. e. & reaD D. j. (2008): Mycorrhizal Symbiosis. 3rd edition. – Academic Press.ŠeDa z. (1976): Inventarizační průzkum vegetačního krytu státní přírodní rezervace Strabišov. – Ms.
[Depon. in: Agentura ochrany přírody a krajiny ČR, středisko Brno.]van Der cinGel N. A. (1995): An atlas of orchid pollination. European orchids. – A. A. Balkema,
Rotterdam.waite s. & hutchinGs M. j. (1991): The effects of different management regimes on the population
dynamics of Ophrys sphegodes: analysis and description using matrix models. – In: wells t. c. & willeMs j. h. [eds], Population Ecology of Terrestrial Orchids, pp. 161–175, SPB Academic Publishing, Hague.
wotavová k., balounová z. & kinDlMann P. (2004): Factors affecting persistence of terrestrial orchids in wet meadows and implications for their conservation in a changing agricultural landscape. – Biol. Conserv. 118: 271–279.

184
Dostalík et al.: Monitoring Orchis tridentata v Národní přírodní rezervaci Strabišov-Oulehla

185
Příroda, Praha, 31: 185–200, 2011
Populační dynamika, energetická náročnost kvetení a přežívání švihlíku krutiklasu (Spiranthes spiralis) na tradičně obhospodařované lokalitě „nPP Pastviště u Fínů“Population dynamics, energetic cost of flowering and survival of Spi ran thes spiralis in regularly managed national nature Monument „Pastviště u Fínů”
Jiří brabec1, 4, Zdeněk iPser2, Jana Jersáková3, taMara MaLinová3, ZuZana baLounová2, Lenka PLesková4, vácLav soMoL4, Jan brabec3, 4, Lukáš krinke4, 5, MichaL štefánek4, 6 & JarosLava nesvadbová7
1 Muzeum Cheb, příspěvková organizace Karlovarského kraje, nám. Krále Jiřího z Poděbrad 493/4, CZ-350 11 Cheb, [email protected] (korespondenční adresa)2 Zemědělská fakulta, Jihočeská univerzita v Českých Budějovicích, Branišovská 31, CZ-370 05 České Budějovice3 Přírodovědecká fakulta, Jihočeská univerzita v Českých Budějovicích, Branišovská 31, CZ-370 05 České Budějovice4 ZO ČSOP Silvatica Brejl, Krupá 28, CZ-270 08 Krupá5 Sládečkovo vlastivědné muzeum, Huťská 1375, CZ-272 01 Kladno6 Herbářové sbírky a Katedra botaniky Univerzity Karlovy, Benátská 2, CZ-128 01 Praha 27 Západočeské muzeum, Kopeckého sady 2, CZ-301 36 Plzeň
Abstract: Population biology of critically endangered species Spiranthes spiralis was studied in one of the two remaining Czech localities (National Nature Monument Pastviště u Fínů). In years 1985–2010, the total number of flowering plants varied from 15 to 181 individuals. Since 1998, we have gradually marked 540 positions with the occurrence of individual plants or dense clumps. Survival of marked individuals and their growth characteristics (plant height, number of flowers, length and width of leaves) were recorded annually from 2000 to 2010. Long-term observation enabled calculation of transition probabilities between life stages. Dormancy (survival without an appearance of the above-ground parts in one or more growing seasons) was a very rare event recorded eight times in six plants. Sterile plants were more likely to remain sterile in a subsequent year (65.5%) than becoming flowering (15.4%) or dead (19.1%). Flowering plants were prone to sterility in the following season (63.7%), rarely dying (8.6%). Despite the energy loss in favour of flowering, the probability of flowering over two consecutive years was 27.6%. The flowering plants had a significantly smaller leaf area in the following season compared to the previous one, while the leaf area of sterile plants remained similar.Keywords: average life span, critically endangered species, fluctuation of flowering plants, grazing of grassland, transition probabilities among life stages
Abstrakt: Populační biologie kriticky ohroženého druhu Spiranthes spiralis byla studována na jedné ze dvou posledních známých lokalit v České republice (NPP Pastviště u Fínů). V letech 1985–2010 byl pravidelně sledován počet kvetoucích exemplářů, který kolísal mezi 15 až 181 exempláři. Od roku 1998 bylo postupně označeno 540 pozic výskytu jednotlivých rostlin nebo těsně u sebe rostoucích skupin rostlin. Přežívání všech označených individuí

186
Brabec et al.: Populační dynamika, energetická náročnost kvetení a přežívání Spiranthes spiralis
a jejich podrobné růstové charakteristiky (výška kvetoucí lodyhy, počet květů, délka a šířka listů přízemní růžice v období maximálního rozvoje) byly pravidelně zaznamenávány v letech 2000 až 2010. Dlouhodobé sledování umožnilo stanovit pravděpodobnost přechodů mezi jednotlivými životními fázemi. Dormance hlíz (přežívání bez vytvoření nadzemní části ve vegetační sezóně) byla zaznamenána velmi výjimečně (osm případů u šesti rostlin). Sterilní rostliny zůstávají v následujícím roce nejčastěji sterilní (65,5 %), méně často kvetou (15,4 %) nebo zcela odumírají (19,1 %). Kvetoucí rostliny zůstávají v následující sezóně nejčastěji sterilní (63,7 %), hynou pouze zřídka (8,6 %). I přestože je kvetení poměrně energeticky náročné, dosahuje pravděpodobnost květu stejné rostliny dva roky po sobě v průměru 27,6 %. Kvetoucí rostliny mají v následujícím roce průkazně menší listovou plochu než v roce předešlém, zatímco listová plocha sterilních rostlin se téměř nezměnila.Klíčová slova: fluktuace kvetení, kriticky ohrožený druh, pastva travinných porostů, pravděpodobnost přechodu mezi životními fázemi, průměrná doba života nomenklatura: kubát et al. 2002 (cévnaté rostliny), Moravec et al. 1995 (vegetace)
Úvod
Populace nelesních orchidejí vesměs vyžadují pravidelné tradiční selské obhospodařování (např. seč, pastva, vláčení, vyřezávání náletu) svých stanovišť (jersáková & kinDlMann 2004). V mnoha oblastech Evropy však tradiční obhospodařování zcela mizí a je nahrazováno různě intenzivními ochranářskými zásahy (PetříČek et al. 1999). Jejich potřebnost a kvalita je vyhodnocována monitoringem populací cílových druhů. Nejčastěji se zjišťuje vliv managementu na počet kvetoucích jedinců, avšak terestrické orchideje patří k druhům, u kterých jsou velmi často zaznamenávány velké meziroční fluktuace v počtech kvetoucích exemplářů (např. hutchinGs 1987, 2010, welss et al. 1998, kinDlMann & balounová 2001, shefferson et al. 2003). U mnohých je také častá dormance hlíz (shefferson et al. 2003, shefferson 2009) nebo je velmi obtížné nové semenáčky a sterilní dospělce v porostu nalézt (brabec et al. 2004). Objektivní vyhodnocení kvality obhospodařování orchidejových stanovišť je tak bez podrobnější znalosti biologie cílových druhů a pravidelného záznamu vypovídajících parametrů jejich populací téměř nemožné (jersáková & kinDlMann 2004). Zároveň nelze vzhledem k finanční a časové náročnosti sledovat populační dynamiku orchidejí na všech potenciálně ohrožených lokalitách. Nejefektivnější je pomocí několika podrobnějších studií vytipovat a ověřit pro vybrané druhy parametry, které mají vysokou vypovídající hodnotu o vitalitě populace a zároveň jsou co nejjednodušeji sledovatelné.
Švihlík krutiklas (Spiranthes spiralis) je ve střední Evropě typickým příkladem extrémně vzácného druhu, který ztratil většinu svých historických lokalit (souhrnně brabec et al. 2004, jacQueMyn & hutchinGs 2010). Jde o druh poměrně dlouhověký (wells 1981, willeMs & DorlanD 2000, kull 2002), s velkou fluktuací v počtu kvetoucích i sterilních jedinců (wells 1967, 1981, willeMs 1989, willeMs & DorlanD 2000, jacQueMyn et al. 2007, nesvaDbová et al. 2003), s nejasným podílem vegetativního a generativního rozmnožování (Machon et al. 2003) a vzhledem ke své nenápadnosti i o druh obtížněji monitorovatelný (brabec et al. 2004). V této studii jsou zpracovány výsledky čtvrtstoletí sledování populace kriticky ohrožené orchideje švihlíku krutiklasu na jediné známé recentní lokalitě v Čechách a současně jsou diskutovány jejich možné aplikace v ochranářské praxi.
Cílem práce bylo (i) popsat populační dynamiku druhu na jedné lokalitě a porovnat ji se zjištěnými parametry obdobně sledovaných populací na jiných lokalitách, (ii) vyhodnotit vliv velikosti a tvaru listové plochy na kvetení (a přežívání) jedince, (iii) odhadnout energetickou náročnost kvetení a (iv) najít parametry populace, které jsou nejvhodnější k posouzení kvality péče o lokalitu a lze je doporučit k dlouhodobému sledování populace.

187
Příroda, Praha, 31: 185–200, 2011
Metodika
Pastviště u Fínů
Národní přírodní památka (dále NPP) Pastviště u Fínů se nachází 4,5 km jihovýchodně od Sušice, 1 km severovýchodně od obce Albrechtice pod samotou „U Fínů“. Populaci Spiranthes spiralis zde nalezl v roce 1980 ornitolog L. Kučera (kuČera 1980, 1982, anonyMus 1982, Procházka & velísek 1983). Jde o komplex několika typů travinných společenstev, kamenných snosů, mezí a remízků. Následný botanický průzkum ukázal, že jde o mimořádně cennou lokalitu s bohatým výskytem ohrožených typů vegetace a vzácných druhů rostlin, které se zde udržely především díky tradičnímu obhospodařování obyvateli okolních usedlostí (nesvaDbová et al. 1987, 1989 a Matějková & nesvaDbová 1999). Na území NPP Pastviště u Fínů (4,2 ha) a jeho ochranného pásma (3 ha) byl zaznamenán výskyt více než 260 druhů cévnatých rostlin, z nich jsou dle vyhlášky MŽP ČR č. 395/1992 Sb. tři druhy kriticky ohrožené (Botrychium matricariifolium, Gentianella praecox subsp. bohemica a Spiranthes spiralis), sedm druhů silně ohrožených (Coeloglossum viride, Drosera rotundifolia, Epipactis palustris, Orchis morio, Orchis ustulata, Pedicularis syl vatica a Pinguicula vulgaris) a 11 druhů ohrožených (Carex davalliana, Carex pulicaris, Dactylorhiza fuchsii, Dactylorhiza majalis, Gymnadenia conopsea subsp. conopsea, Par na ssia palustris, Platanthera bifolia, Salix repens, Taraxacum madidum, Taraxacum trilobifolium a Willemetia stipitata). Některé z uvedených druhů nebyly však již dlouho dobu ověřeny (např. Botrychium matricariifolium, Taraxacum madidum a Taraxacum trilo bifolium). Z dalších zajímavých druhů se zde vyskytují dva silně ohrožené druhy podle červeného seznamu (Procházka 2001) – Triglochin palustre a Eriophorum latifolium (podle nesvaDbová et al. 1989 a dalších, nepublikovaných záznamů).
Populace S. spiralis se nachází v severní části území na ploše ca 1 ha, na jiho-východně orientovaných svazích se sklonem 0–20° (nesvaDbová et al. 1987, nesvaDbová & Pivoňková 1996). Druh roste v mozaice krátkostébelných porostů svazů Violion caninae (as. Polygalo-Nar detum a as. Thymo-Festucetum ovinae), Arrhenatherion (as. Trifolio-Festucetum ru brae subas. nardetosum) a v místech s vystupujícím skalním podložím ve společenstvech svazu Hyperico perforati-Scleranthion perennis (as. Jasiono montanae-Festucetum ovinae) (nesvaDbová et al. 1989, 2003, brabec et al. 2004). Podle historických materiálů (Parcellen – Protokoll der Gemeinde Albrechtsried) z roku 1895 byla lokalita v minulosti využívána převážně jako louka, méně často jako orná půda a pastvina (nesvaDbová et al. 1987). Zajímavé je, že právě parcela se současným výskytem S. spiralis je zde uváděna jako orná půda a dle ústního sdělení starousedlíků byla takto využívána ještě po roce 1945 (nesvaDbová et al. 1987). V době nálezu Spiranthes spiralis byly pozemky v NPP Pastviště u Fínů užívány Státním statkem Sušice, později Jednotným zemědělským družstvem Žihobce. Ani jedna z těchto organizací neměla o svažité pozemky skutečný zájem a systematicky zde nehospodařila, jen příležitostně byly travnaté stráně přepásány menšími stády hovězího dobytka. V roce 1985 byla vyhlášena ochrana územní (formou chráněného přírodního výtvoru), zároveň však byl zjištěn rapidní úbytek počtu kvetoucích švihlíků a zaznamenány změny ve složení a struktuře travního porostu, včetně nárůstu náletových dřevin (nesvaDbová & Pivoňková 1996). Díky práci dobrovolníků a pochopení úředníků ochrany přírody se podařilo postupně vyřezat náletové dřeviny a od roku 1988 zajistit sezónní pastvu ovcí, která probíhá dodnes (nesvaDbová & Pivoňková 1996, nesvaDbová et al. 2003, brabec et al. 2004).

188
Brabec et al.: Populační dynamika, energetická náročnost kvetení a přežívání Spiranthes spiralis
V současné době je plocha s výskytem S. spiralis oplocena a je na ní aplikována tzv. řízená pastva (podle plánu péče na léta 2005 až 2014, slaDký & liŠková 2004). Pastva ovcí je prováděna každoročně ca od 1. 6. do 15. 8., tedy v období kdy S. spiralis nemá nadzemní vegetativní orgány, a následně pak od 15. 10. do zámrazu. V případě nárůstu většího množství biomasy je lokalita převláčena (naposledy bylo vláčení provedeno na podzim 2009).
Spiranthes spiralis
Spiranthes spiralis (L.) Chevall. je považován za oceánický a atlantsko-mediteránní druh. Jeho areál se táhne od západního Středozemí (včetně severní Afriky) přes celou Evropu až do Malé Asie (Turecko, Libanon) a na Kavkaz. Severní hranice výskytu prochází Irskem, severní Anglií, Belgií, Nizozemím, Dánskem, švédským ostrovem Öland, pobaltskými republikami a evropskou částí Ruska (brabec et al. 2004, jacQueMyn & hutchinGs 2010). S výjimkou několika středomořských států a Velké Británie jde o velmi vzácný a ohrožený druh (podrobněji viz brabec et al. 2004). Hlavním důvodem mizení tohoto druhu ve velké části jeho areálu je změna způsobů obhospodařování lokalit a likvidace vhodných stanovišť (jacQueMyn & hutchinGs 2010). Přitom kromě malé konkurence okolní vegetace nevyžaduje tento druh žádné specifické podmínky a dovede osídlit celou škálu travinných porostů na kyselých i bazických substrátech. Roste od mořského pobřeží (jacQueMyn & hutchinGs 2010) až do vysokých hor (max. 2260 m n. m., V. Vlačiha a J. Brabec, vlastní pozorování z roku 2005) na vlhčích rašelinných a slatinných loukách, v xerotermních a semixerotermních trávnících, na pastvinách, na vřesovištích a na písčinách (brabec et al. 2004, jacQueMyn & hutchinGs 2010).
Fenologie S. spiralis odpovídá sezónnímu rytmu klimatu v mediteránní oblasti (viz obr. 1). Švihlík krutiklas tak má přezimující růžici zelených listů, což je mezi středoevropskými orchidejemi vzácný jev (z našich druhů takto přezimují např. Orchis morio, Anacamptis pyramidalis či Himantoglossum adriaticum; jersáková & kinDlMann 2004). Ve vývoji a diferenciaci podzemních částí rostliny panují evidentně stále nejasnosti (wells 1967, brabec et al. 2004, jacQueMyn & hutchinGs 2010). V lednu se u dospělé rostliny nacházejí pod zemí jedna až tři (nejčastěji dvě) světle hnědé, hladké, ztlustlé kořenové hlízy vytvořené v předchozím roce, a dále scvrklé zbytky hlíz, které patřily loňské kvetoucí lodyze, tj. v květnu zatahující listové růžici. Kořenové hlízy jsou navzájem spojeny bází růžice. Navíc jsou již založeny a dobře patrné nové hlízy (malé oválné až hruškovité útvary) (wells 1967, brabec et al. 2004). Nad zemí se v lednu nachází přezimující růžice zelených listů. Až do přelomu května a června růžice stále narůstá, její listy vždy v klimaticky vhodném období asimilují a pod zemí se z malých nových hlíz postupně vytváří dospělé kořenové hlízy (viz obr. 1, brabec et al. 2004, jacQueMyn & hutchinGs 2010). Listová růžice dosahuje své maximální velikosti v květnu a poté postupně odumírá. Během června a července jsou listy zcela odumřelé a rostlina je bez viditelné nadzemní části. V průběhu srpna vyrůstá ze staré hlízy květonosná lodyha (willeMs & lahtinen 1997, jacQueMyn & hutchinGs 2010). Rostliny kvetou od srpna do poloviny září. Od konce srpna či v září se nad zemí objevuje listová růžice, která v případě fertilní rostliny vyrůstá vedle květonosného stonku (wells 1981). Kvetení a následné tvorby plodů se zřejmě zčásti účastní i nové hlízy, které koncem srpna nebo během září vytváří novou listovou růžici (willeMs & lahtinen 1997).
Květonosná lodyha je (4,5–)7–20(–35) cm dlouhá. Na lodyze se nachází 3–7 blanitých přitisklých lístků a (1–)10–25(–30) květů v charakteristicky spirálovitě stočeném květenství (brabec et al. 2004, jacQueMyn & hutchinGs 2010). Přízemní listová růžice je podle literatury složená ze 3–7 (zřídka přes 10) listů o délce 2,0–4,0(–5,5) a šířce (0,6–)0,8–1,8 cm

189
Příroda, Praha, 31: 185–200, 2011
obr. 1. Schematický nákres životních fází švihlíku krutiklasu během jednoho roku. Upraveno podle brabec et al. (2004).Fig. 1. A schematic view of the Spiranthes spiralis life stages during a year (modified from brabec et al. 2004).
(jacQueMyn & hutchinGs 2010). Listové růžice zaznamenané v letech 2000 až 2010 na Pastvišti u Fínů byly v době svého největšího rozvoje (tj. v květnu) tvořené průměrně 6–7 (min. 3, max. 19) listy o délce až 6,3 cm a šířce až 2,4 cm (brabec et al. 2004, Brabec a kol., nepublikované údaje). Údaje o menších rozměrech listů na jiných lokalitách ve srovnání s populací na Pastvišti u Fínů jsou způsobeny jejich popisem v době květu, tj. v době první fáze růstu. Drobné květy jsou bílé nebo nazelenale bílé s nažloutle zeleným středem. Korunní lístky jsou 5 až 7 mm dlouhé, celý plně vyvinutý květ je jen zřídka větší než 10 mm (brabec et al. 2004). Květy v květenství se otevírají odspodu a to často synchronně ve skupinách po třech až pěti květech. Opylovány jsou hmyzem, zejména čmeláky. Plodem je tobolka s velkým množstvím větrem šířených semen. Počet semen v jedné tobolce je silně variabilní, willeMs & lahtinen (1997) udávají 0 až méně než 2000 semen na tobolku. Semena švihlíku patří k nejmenším u středoevropských orchidejí, jejich rozměry jsou jen 0,32–0,38 mm × 0,1–0,15 mm (Procházka & velísek 1983).
Metody sledování
Sledování počtu kvetoucích exemplářů probíhá na lokalitě nepřetržitě od roku 1985, přičemž do roku 1995 nebyla prohledávána celá pastvina, proto mohou být údaje z let 1986 až 1994 mírně podhodnocené. V letech 1998 až 2010 byly označovány všechny postupně nalézané exempláře Spiranthes spiralis, respektive všechny pozice výskytu, tj. skupiny rostlin rostoucí vedle sebe. K označení jednotlivých rostlin (popř. pozic výsky-

190
Brabec et al.: Populační dynamika, energetická náročnost kvetení a přežívání Spiranthes spiralis
tu) bylo využito dvou způsobů: 1) přímé označení očíslovaným hřebíkem v přesně de-fino vané vzdálenosti a směru od jedince; 2) zaměření jednotlivých rostlin (popř. pozic výskytu) s přesností na 0,5 cm k pevně fixovaným liniím či plochám. Druhý způsob byl použit v místech nejhustšího výskytu. Přítomnost rostlin a jejich stav byly zjišťovány dvakrát ročně: počátkem května (2000–2010), kdy končí vegetační sezóna zelených nadzemních růžic, a na přelomu srpna a září (1998–2010), tj. v době květu a nárůstu nových následně přezimujících růžic.
Zaznamenány byly následující parametry – počátek května: • prezence/absence nadzemních částí rostliny na pozici výskytu,• počet listových růžic na pozici výskytu,• délka a šířka všech listů každé listové růžice a jejich stav,• plocha půdorysu každé listové růžice (fotodokumentace s měřítkem),srpen/září: • prezence/absence květonosných stonků a počet kvetoucích jedinců na pozici výskytu,• výška každého stonku a počet květů,• počet nově vyrůstajících růžic na pozici výskytu.
Z délky a šířky listů byla extrapolována velikost listové plochy růžice jako 2/3 × součet násobků maximální délky a maximální šířky každého listu růžice. Korekční koeficient listové plochy byl spočten jako podíl reálné listové plochy (zjištěné překreslením na milimetrový papír) u deseti náhodně vybraných listů a plochy spočtené jako obdélník (výška × délka listu). Průměr tohoto podílu byl 0,69, tj. přibližně 2/3.
Fotodokumentace pozice výskytu byla prováděna při jarním monitoringu (duben/ květen) v letech 2000–2010. Fotografie byly pořizovány kolmo na listovou růžici, popřípadě na skupinu růžic. Před fotografováním byl v blízkosti růžice umístěn kalibrační čtvereček (o velikosti 1 cm2) a označení „pozice výskytu“. Fotografie byly zpracovány analýzou obrazu v softwaru NIS Elements, varianta Advanced Research, verze 3.0 (výrobce Laboratory Imaging s.r.o., Praha).
Velikost listů byla charakterizována třemi způsoby – poměrem průměrné délky listu růžice k průměrné šířce listu růžice, průměrnou délkou listu v růžici a průměrnou šířkou listu v růžici.
vyhodnocení dat
Pro analýzu populační dynamiky kvetoucích a sterilních rostlin Spiranthes spiralis byly použity kompletní údaje ze sledování populace v letech 1986 až 2010, tj. 663 rostlin z 540 pozic výskytu.
Odhad průměrné délky života jednotlivých rostlin (tj. počet let, kdy byla zaznamenána nadzemní část rostliny) byl vypočten pro jednotlivé kohorty rostlin z let 1998 až 2010 (tj. pro rostliny poprvé zaznamenané v těchto jednotlivých letech). Do odhadu byly zahrnuty pouze rostliny (celkem 438), u nichž existoval reálný předpoklad, že pocházejí z jedné genety, tj. byly hodnoceny pouze pozice výskytu s jednou nadzemní částí rostliny (listovou růžicí) a pozice výskytu, na nichž došlo ke zřejmému vegetativnímu rozdělení rostliny během monitoringu, a zároveň vegetativně vzniklá růžice během dalšího monitoringu nezanikla. V případě, že jedna růžice zanikla, není jasné, jak počítat věk zbývající rostliny. Stejná kritéria výběru sledovaných rostlin byla použita při odhadu průměrné doby přežívání na lokalitě. Průměr byl stanoven z 252 rostlin sledovaných déle než pět let.
Pravděpodobnosti přechodů mezi jednotlivými fenofázemi rostliny byly spočteny na základě údajů o zaznamenaných přechodech z roku t na rok t+1 z let 1998 až 2010 celkem u 1522 rostlin.

191
Příroda, Praha, 31: 185–200, 2011
Statistické analýzy byly provedeny v programech Statistica v. 10 (statsoft 2011) a R package v. 2.12 (r DeveloPMent core teaM 2006). Pro analýzu vlivu velikosti a tvaru listové plochy na životaschopnost rostlin byly použity údaje z celkem 1070 samostatně rostoucích rostlin měřených a zaznamenaných v letech 2000 až 2009. Vyřazeny byly rostliny, jejichž listové růžice byly v době měření významně poničené, a rostliny, u nichž byla zaznamenána dormance nadzemních částí. Regresní koeficient vyjadřující míru těsnosti vztahu mezi měřenou a vyfotografovanou listovou plochou byl vypočten pomocí lineární regrese. Analýza stavu rostlin v následujícím období (nezávislá kategoriální proměnná) na základě měřené velikosti listové plochy, fotografované velikosti listové plochy, počtu listů růžice na jaře a charakteristik tvarů listu (vždy jako spojitá závislá proměnná) byla provedena jednocestnou analýzou variance. Zjištění rozdílů mezi jednotlivými proměnnými bylo provedeno následným mnohonásobným porovnáním Tukey HSD testem. Z charakteristik tvarů listů byly testovány poměr průměrné délky listu růžice k průměrné šířce listu růžice, průměrná délka listu v růžici a průměrná šířka listu v růžici. Proměnné listová plocha a tvar listů byly normalizovány pomocí odmocninové transformace. Pro analýzu nákladů na kvetení promítnutých do velikosti listové plochy jsme použili údaje z celkem 557 rostlin, u nichž byla známá velikost měřené listové plochy v roce t a t+1 a zároveň jejich stav na podzim roku t (140 kvetoucích a 417 sterilních). Meziroční rozdíly ve velikosti listové plochy kvetoucích a sterilních rostlin byly analyzovány dvoucestnou analýzou variance s interakcemi a následným mnohonásobným porovnáváním Tukey HSD testem.
výsledkyPopulační dynamika
V letech 1986 až 1997 byly sledovány počty kvetoucích jedinců, následně v letech 1998 až 2010 bylo celkem na 540 pozicích výskytu sledováno 663 rostlin Spiranthes spiralis. Fluktuace počtu kvetoucích a sterilních exemplářů ukazuje obr. 2. V letech 1986 až 2010 byly zaznamenány tři výraznější nárůsty počtu kvetoucích rostlin: 1995 (181 kvetoucích), 2005 (140) a 2010 (113). Naopak méně než 25 kvetoucích exemplářů bylo zjištěno v letech 1988 až 1992, 2003, 2004 (absolutní minimum 15 kvetoucích) a 2007. S propady počtu kvetoucích exemplářů koreluje v letech 2000 až 2010 i snížení počtu živých rostlin (obr. 2).
Rostliny S. spiralis jsou relativně dlouhověké. Průměrná doba přežívání jednotlivých rostlin zaznamenaná na lokalitě Pastviště u Fínů je 4,7 roku (průměr z 252 rostlin sledovaných déle než pět let). Průměrnou dobu přežívání kohort z jednotlivých roků ukazuje obr. 3. Jednotlivých rostlin přežívajících více než deset let bylo zaznamenáno 20 (z celkem 95 takto dlouho sledovaných). Rostlin přežívajících celou dobu sledování, tj. 13 let, bylo zaznamenáno celkem devět.
V jednotlivých letech života prochází rostlina vždy některou ze tří fenofází: kvetení, přežívání ve formě nadzemní růžice nebo přežívání podzemní části (tzv. dormance). Pravděpodobnosti přechodů mezi jednotlivými fenofázemi z roku na rok ukazuje obr. 4. Dormance byla u sledované populace zaznamenána velmi vzácně, v celém souboru dat existuje pouze osm případů u šesti rostlin. Jednalo se o roky 2003, 2004 a 2007, ve dvou případech byl tentýž exemplář dormantní dvakrát za dobu sledování (vždy 2003 a 2007). Vzhledem k výjimečnosti není dormance v obr. 4 zohledněna.
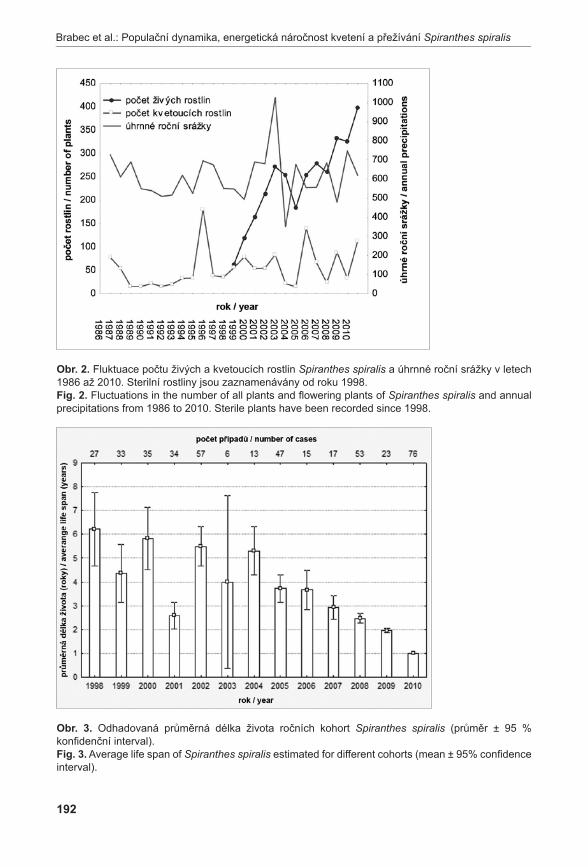
192
Brabec et al.: Populační dynamika, energetická náročnost kvetení a přežívání Spiranthes spiralis
obr. 2. Fluktuace počtu živých a kvetoucích rostlin Spiranthes spiralis a úhrnné roční srážky v letech 1986 až 2010. Sterilní rostliny jsou zaznamenávány od roku 1998.Fig. 2. Fluctuations in the number of all plants and flowering plants of Spiranthes spiralis and annual precipitations from 1986 to 2010. Sterile plants have been recorded since 1998.
obr. 3. Odhadovaná průměrná délka života ročních kohort Spiranthes spiralis (průměr ± 95 % konfidenční interval).Fig. 3. Average life span of Spiranthes spiralis estimated for different cohorts (mean ± 95% confidence interval).

193
Příroda, Praha, 31: 185–200, 2011
obr. 4. Pravděpodobnosti přechodů (počet případů) mezi jednotlivými fenofázemi z roku t na rok t+1 (na základě údajů sloučených z let 1998–2010).Fig. 4. Transition probabilities (number of cases) of individual plants among stages from year t to year t+1 (data pooled from period 1998–2010).
Pravděpodobnosti přechodů na obr. 4 zahrnují též údaje o 94 rostlinách, které se z roku na rok vegetativně rozdělily. Ve sledovaném souboru bylo zaznamenáno rozdělení 52 sterilních rostlin (4,9 %) a 42 kvetoucích rostlin (9,1 %). Z rozdělených 52 sterilních rostlin v následujícím roce kvetla jedna nová rostlina ve 14 případech, z rozdělených 42 kvetoucích rostlin kvetla téměř polovina (20 případů).
vliv velikosti a tvaru listové plochy na životaschopnost rostlinyVelikost asimilační listové plochy na jaře silně ovlivňuje životaschopnost (stav) rostliny v dalším období, zejména na podzim téhož roku. Oba způsoby stanovení velikosti listové plochy (výpočet z měření a analýza fotografie) vykazují shodný efekt, mezi měřenou a focenou listovou plochou je totiž vysoká míra korelace (regresní koeficient R2 = 0,89). Měřená plocha vysvětluje 25 % variability v chování rostlin (F(4, 1065) = 90,5; p < 0,0001), focená jen o 3 % méně (F(4, 1065) = 77,8; p < 0,0001). Jak ukazuje obr. 5, na podzim kvetoucí rostliny měly na jaře průkazně větší listovou plochu než rostliny na podzim sterilní či následně uhynulé. Listová plocha kvetoucích rostlin, které se následující rok spontánně rozdělily na dvě a více růžic, byla na jaře průkazně větší než u těch, které se nerozdělily. V následujícím období uhynulé rostliny měly na jaře dle očekávání listovou plochu nejmenší.
Zcela shodný vliv na životaschopnost (stav) rostliny v dalším období vykazuje též počet listů jarní růžice (viz obr. 6), která vysvětluje 22 % variability v datech(F(4,1065) = 76,9; p < 0,0001), tedy shodně jako focená listová plocha. Na podzim kvetoucí rostliny měly na jaře průkazně více listů než rostliny, které zůstaly sterilní (vytvořily pouze novou růžici) nebo které v následujícím období nebyly nalezené (uhynuly). Počet listů kvetoucích rostlin, které se rozdělily na dvě a více růžic, byl na jaře průkazně větší než u těch, které se nerozdělily. Rostliny, které na podzim a ani v následujících letech nebyly nalezeny, měly na jaře sledovaného roku nejmenší počet listů.
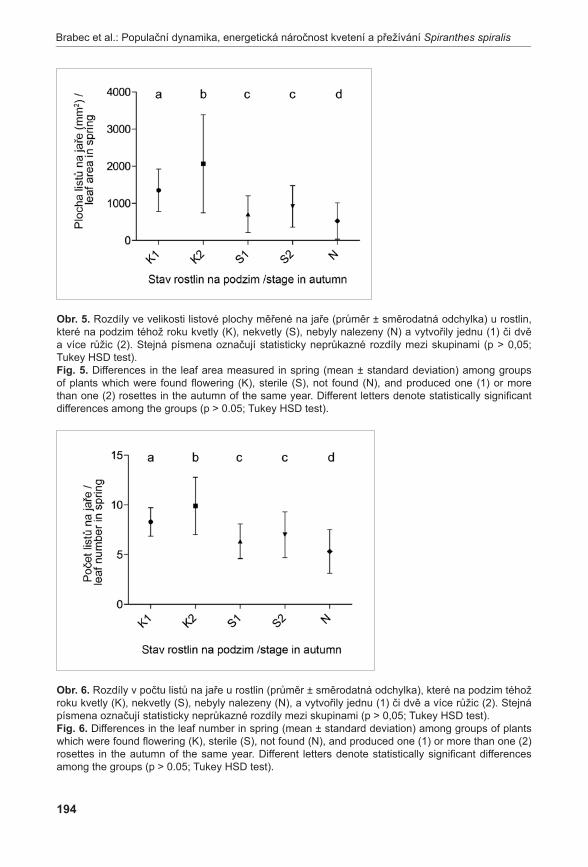
194
Brabec et al.: Populační dynamika, energetická náročnost kvetení a přežívání Spiranthes spiralis
obr. 5. Rozdíly ve velikosti listové plochy měřené na jaře (průměr ± směrodatná odchylka) u rostlin, které na podzim téhož roku kvetly (K), nekvetly (S), nebyly nalezeny (N) a vytvořily jednu (1) či dvě a více růžic (2). Stejná písmena označují statisticky neprůkazné rozdíly mezi skupinami (p > 0,05; Tukey HSD test).Fig. 5. Differences in the leaf area measured in spring (mean ± standard deviation) among groups of plants which were found flowering (K), sterile (S), not found (N), and produced one (1) or more than one (2) rosettes in the autumn of the same year. Different letters denote statistically significant differences among the groups (p > 0.05; Tukey HSD test).
obr. 6. Rozdíly v počtu listů na jaře u rostlin (průměr ± směrodatná odchylka), které na podzim téhož roku kvetly (K), nekvetly (S), nebyly nalezeny (N), a vytvořily jednu (1) či dvě a více růžic (2). Stejná písmena označují statisticky neprůkazné rozdíly mezi skupinami (p > 0,05; Tukey HSD test).Fig. 6. Differences in the leaf number in spring (mean ± standard deviation) among groups of plants which were found flowering (K), sterile (S), not found (N), and produced one (1) or more than one (2) rosettes in the autumn of the same year. Different letters denote statistically significant differences among the groups (p > 0.05; Tukey HSD test).
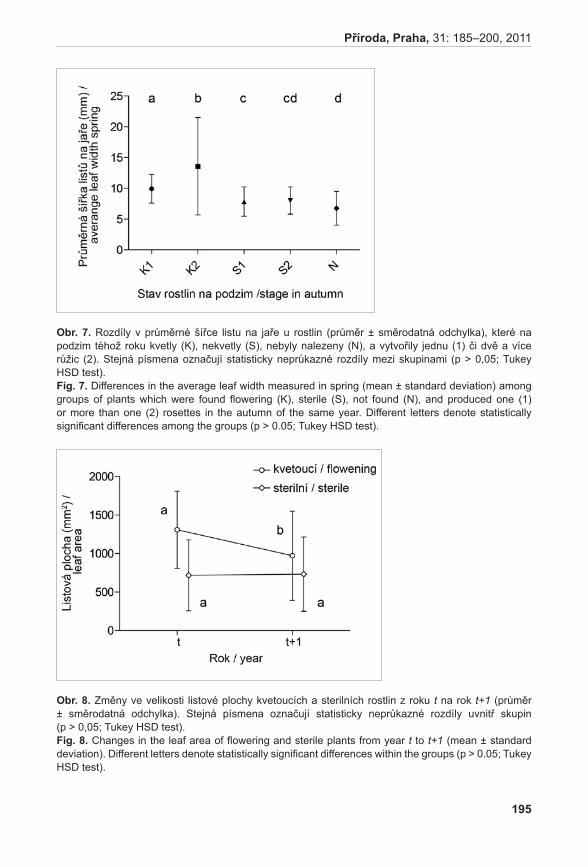
195
Příroda, Praha, 31: 185–200, 2011
obr. 7. Rozdíly v průměrné šířce listu na jaře u rostlin (průměr ± směrodatná odchylka), které na podzim téhož roku kvetly (K), nekvetly (S), nebyly nalezeny (N), a vytvořily jednu (1) či dvě a více růžic (2). Stejná písmena označují statisticky neprůkazné rozdíly mezi skupinami (p > 0,05; Tukey HSD test).Fig. 7. Differences in the average leaf width measured in spring (mean ± standard deviation) among groups of plants which were found flowering (K), sterile (S), not found (N), and produced one (1) or more than one (2) rosettes in the autumn of the same year. Different letters denote statistically significant differences among the groups (p > 0.05; Tukey HSD test).
obr. 8. Změny ve velikosti listové plochy kvetoucích a sterilních rostlin z roku t na rok t+1 (průměr ± směrodatná odchylka). Stejná písmena označují statisticky neprůkazné rozdíly uvnitř skupin (p > 0,05; Tukey HSD test).Fig. 8. Changes in the leaf area of flowering and sterile plants from year t to t+1 (mean ± standard deviation). Different letters denote statistically significant differences within the groups (p > 0.05; Tukey HSD test).

196
Brabec et al.: Populační dynamika, energetická náročnost kvetení a přežívání Spiranthes spiralis
Z testovaných charakteristik tvaru listů (poměr průměrné délky listu růžice k průměrné šířce listu růžice, průměrná délka listu v růžici a průměrná šířka listu v růžici) vysvětluje nejvíce variability (16 %) průměrná šířka listu (F(4,1065) = 53,3; p < 0,0001), přičemž rostliny sterilní a nenalezené mají užší listy než rostliny kvetoucí (viz obr. 7).
Energetická náročnost kvetení
Kvetení je pro rostliny energeticky poměrně náročné. Obr. 8 srovnává změnu velikosti listové plochy u kvetoucích a sterilních rostlin z roku t na rok t+1. Průkazná interakce(F(1,1110) = 25,74; p < 0,0001) mezi stavem rostliny (kvetoucí a sterilní) a rokem (t a t + 1) ukazuje, že kvetoucí rostliny měly v následujícím roce průkazně menší listovou plochu než v roce předešlém, zatímco listová plocha sterilních rostlin se téměř nezměnila.
diskusePopulační dynamika
Celkem 25 let pravidelného sledování počtu kvetoucích exemplářů Spiranthes spiralis na lokalitě Pastviště u Fínů ukázalo výrazné meziroční rozdíly. Taktéž celkový počet živých exemplářů vykazoval podobné, byť ne tak výrazné fluktuace. Tento jev je znám u řady terestrických orchidejí (např. shefferson et al. 2003, hutchinGs 2010), přímo u druhu S. spiralis byl popsán např. v dlouhodobě sledované populaci v Nizozemí (jacQueMyn et al. 2007). Fluktuace jsou většinou přičítány „ceně za kvetení“, klimatickým faktorům nebo kombinaci obojího (jacQueMyn et al. 2007). V populaci na Pastvišti u Fínů však nebyly zaznamenány pravidelné propady počtu kvetoucích exemplářů následující rok po kvetení většího počtu rostlin, jako tomu je podle Willemse & Dorlanda (Willems & Dorland 2000)v Nizozemí. Sledované propady jsou těmito autory přičítány energetické náročnosti kvetení, která je dokládána jako pokles počtu listů růžice v době kvetení (v témže roce) a jako významně velký podíl (více než 80 %) rostlin, které zůstanou následující rok sterilní (willeMs & DorlanD 2000). V námi sledované populaci bylo však v roce, následujícím po kvetení sterilních pouze 63,7 % rostlin, ca 8,6 % nebylo nalezeno (vesměs uhynulo) a více než 25 % (128 případů) opět kvetlo (viz obr. 4). Pro zajímavost lze uvést, že za 13 let sledování kvetlo třikrát po sobě deset rostlin, čtyřikrát čtyři rostliny, pětkrát dvě rostliny a šestkrát jedna rostlina (ze 272 údajů, kdy bylo sledování pozice s jedinou růžicí delší než dva roky). Zdá se tedy, že násobné kvetení je v šumavské populaci mnohem častější než v Nizozemí. Velmi nízký počet zaznamenané dormance (pouze osm případů) odpovídá údajům z nizozemské populace (jacQueMyn et al. 2007). Roky, v nichž se vyskytly případy dormance na Pastvišti u Fínů, byly buď nadprůměrně suché (2003 a 2007), nebo šlo o rok 2004, který bezprostředně navazoval a nesl v sobě důsledky extrémně suchého roku 2003.
Jak ukazuje průběh úhrnného množství srážek na obr. 2, lze určitou korelaci mezi klimatickými faktory a fluktuací počtu živých a kvetoucích exemplářů očekávat. Identifikace těchto faktorů však bude vyžadovat analýzu podrobnějších klimatických dat, zejména srážek v období výskytu nadzemních částí S. spiralis, a to včetně množství a délky sněhové pokrývky ve sledovaných letech. Ve srovnání s ostatními dlouhodobě sledovanými lokalitami druhu (Anglie, Nizozemí) se šumavská populace jako jediná nachází v oblasti s dlouhodobou sněhovou pokrývkou (sezónní průměr 60 až 80 dní; tolasz et al. 2007).
Průměrná délka života jednotlivých kohort S. spiralis (měřená jako počet let od prvního záznamu nadzemní části rostliny) se pohybovala v rozmezí 2,6 až 6,2 roku (viz obr. 3,

197
Příroda, Praha, 31: 185–200, 2011
u kohort sledovaných déle než tři roky, tj. 1998 až 2007). To poměrně dobře koresponduje se shodně získanými údaji z Nizozemí (1,33 až 9 let, viz jacQueMyn et al. 2007), či z Anglie, kde wells (1981) dokládá poločasy kohort S. spiralis mezi 4,6 až 9,2 lety. Mladší kohorty (tj. 2008 až 2010) nemá smysl vyhodnocovat, neboť jsou sledovány příliš krátkou dobu. Doba sledování ovlivňuje průměrnou délku života jednotlivých kohort až do doby, kdy uhynou všechny rostliny dané kohorty. Z nejstarší námi sledované kohorty přežívá stále devět rostlin.
vliv velikosti a tvaru listové plochy na životaschopnost rostliny a energetická náročnost kvetení
Tato studie ukázala, že velikost listové plochy u druhu S. spiralis ovlivňuje pravděpodobnost kvetení a zároveň kvetení má vliv na velikost listové plochy v následujícím roce. Tvorba květů a plodů je u vstavačovitých rostlin obecně náročná na energetické zdroje. Kvetení a vysoká produkce plodů v jedné sezóně může u orchidejí negativně ovlivnit budoucí růst a velikost rostlin, jejich reprodukci a přežívání – např. Tipularia discolor (snow & whiGhaM 1989), Epidendrum ciliare (ackerMan & Montalvo 1990), Dendrobium monophyllum (bartareau 1995), Orchis purpurea (jacQueMyn et al. 2010). Vliv energetických nákladů na kvetení na budoucí fitness rostlin druhu Spiranthes spiralis byl dokumentován již v předešlých studiích (willeMs & DorlanD 2000, willeMs et al. 2001). Nicméně existují druhy, jako Cypripedium acaule (PriMack & stacy 1998) či C. calceolus subsp. parviflorum (shefferson et al. 2003), které musí kvést a tvořit plody po několik let za sebou, než se u nich projeví limitace zdroji. Podobně studie provedená na Cypripedium calceolus a Cephalanthera longifolia neprokázala negativní vliv kvetení na fitness rostlin (shefferson et al. 2005). Schopnost druhů vyrovnat se s náklady na kvetení a tvorbu plodů by mohla záviset na typu kořenového systému jednotlivých druhů. Zatímco S. spiralis a Orchis purpurea jsou hlíznaté druhy s několika adventivními kořeny, Cypripedium spp. a Cephalanthera longifolia jsou klonální oddenkaté druhy s rozsáhlým kořenovým systémem. Protože hlíznaté druhy musí každoročně obměnit starou hlízu za novou, je pravděpodobné, že jejich vitalita je mnohem více závislá na množství naakumulovaných fotosyntetických asimilátů, než je tomu u druhů oddenkatých, které mohou skladovat zdroje ve vytrvalém podzemním systému.
Listová plocha zjišťovaná pomocí fotografie vysvětluje jen o 3 % méně variability než měřená plocha. Navíc stejné procento variability v datech vysvětluje pouhý počet listů v růžici. Z toho lze vyvodit, že pro odhad životaschopnosti rostliny a predikci jejího budoucího vývoje tak zcela dostačuje zjistit počet listů v růžici v době maximálního rozvoje, tj. v květnu. Vzhledem ke značným rozdílům v pracnosti a časové náročnosti mezi zmiňovanými metodami sběru dat lze považovat toto zjištění za významné, zejména pro ochranářskou praxi.
Původním impulsem ke stanovení asimilační plochy listové růžice dvěma způsoby (měřením listů a fotografováním růžice kolmo k zemi) byla snaha kvantifikovat vliv zastínění na životaschopnost rostliny. Experiment vycházel z pozorovaného a následně experimentálně ověřeného předpokladu (willeMs et al. 2001), že nezastíněná listová růžice Spiranthes spiralis má listy široké, dobře vyvinuté, přitisknuté k zemi. Oproti tomu zastíněná listová růžice má listy užší, vztyčené od země. Míra zástinu byla v našem případě kvantifikována tzv. „indexem zastínění“, který byl vypočten jako poměr měřené velikosti listové plochy a listové plochy, zjištěné pomocí obrazové analýzy. Rozdíl mezi oběma hodnotami měl být úměrný míře zkreslení u zastíněných listů (tedy těch, které nebyly přitisklé, ale svíraly s povrchem půdy úhel, úměrný míře zastínění). Index zastínění byl použit pro testování hypotézy, že životaschopnost rostliny (tj. schopnost přežít, vykvést,

198
Brabec et al.: Populační dynamika, energetická náročnost kvetení a přežívání Spiranthes spiralis
vegetativně se rozmnožit apod.) je ovlivňována mírou zastínění jejího asimilačního aparátu okolní vegetací. Jak se však ukázalo, oba typy stanovení velikosti asimilační plochy byly silně korelovány a vypočtený index zastínění měl na životaschopnost (stav) rostliny v následujícím období minimální vliv (podrobně viz iPser 2010).
Jak však ukazuje obr. 7, životaschopnost rostliny výrazně ovlivňuje již průměrná šířka listu, která pravděpodobně koreluje s mírou zástinu (viz willeMs et al. 2001). Na základě tohoto výsledku lze S. spiralis považovat za druh tolerantní ke střednímu zastínění. Je tedy pravděpodobné, že na lokalitě Pastviště u Fínů, obhospodařované více než 20 let pravidelnou a pravděpodobně dostatečně intenzivní pastvou ovcí, většina rostlin S. spiralis stresem ze zastínění netrpí.
Závěr
Čtvrtstoletí sledovaná populace Spiranthes spiralis v NPP Pastviště u Fínů vykazuje stejnou populační dynamiku a podobné charakteristiky jako dlouhodobě sledované populace v Nizozemí (willeMs 1989, willeMs & DorlanD 2000, jacQueMyn et al. 2007) a v Anglii (wells 1967, 1981). Přestože se jako jediná z těchto populací nachází v oblasti s dlouhodobou sněhovou pokrývkou (sezónní průměr 60 až 80 dní; tolasz et al. 2007) a zároveň se nachází mimo historické těžiště výskytu (brabec et al. 2004, jacQueMyn & hutchinGs 2010), nevykazuje nikterak sníženou životaschopnost. Více než 20 let trvající pravidelné obhospodařování lokality pastvou ovcí a občasným převláčením (nesvaDbová & Pivoňková 1996, nesvaDbová et al. 2003, brabec et al. 2004) evidentně dostatečně odstraňuje zástin vegetace nad rostlinami Spiranthes spiralis. Pokud se podaří toto tradiční obhospodařování zachovat, lze podrobný monitoring populace tohoto extrémně vzácného druhu květeny České republiky zúžit na pravidelné zaznamenání přežívajících a kvetoucích exemplářů (dvakrát ročně) a na měření jednodušších charakteristik pro záznam životaschopnosti populace, např. počtu listů růžice a délky a šířky největšího listu (jedenkrát ročně).
summary
A twenty-five-year study of Spiranthes spiralis population in National Nature Monument Pastviště U Fínů shows same pattern in population dynamics and similar characteristics as long-term studies of Dutch and British populations. Though the Czech population occurs in an area with long-lasting snow cover (seasonal average is 60 to 80 days) and outside the species’ historical range, it does not show decreased vitality. Regular management (sheep grazing and occasional harrowing) for more than 20 years sufficiently eliminates effect of vegetation shading on Spiranthes spiralis plants. If such traditional management is also kept in future, the detailed monitoring of the population of an extremely rare species of the Czech flora can be reduced to regular census of surviving and flowering plants (twice a year) and measurement of simpler characteristics recording population vitality, e.g. leaf number in a rosette and length and width of the largest leaf (once a year).
Poděkování
Autoři velmi děkují recenzentovi za podnětné připomínky k textu. Čtvrtstoletí bádání na lokalitě Pastviště u Fínů a dlouhodobé sledování populace švihlíku krutiklasu se neobešlo bez pomocí více než šesti desítek dobrovolníků zejména z řad členů a přátel základní organizace ČSOP Silvatica

199
Příroda, Praha, 31: 185–200, 2011
Brejl a botanického oddělení Západočeského muzea v Plzni. Všem jim patří velký dík. Prezentovaná studie byla v letech 2006 až 2011 součástí projektu MŠMT VaV 2B06178. V letech 2000 až 2003 byly výzkumy podpořeny v rámci projektu VaV 610/10/00 Vliv hospodářských zásahů na změnu biologické diverzity ve zvláště chráněných územích.
literatura
ackerMan j. D. & Montalvo a. M. (1990): Short- and long-term limitations to fruit production in a tropical orchid. – Ecology 71: 263–272.
anonyMus (= kuČera l.) (1982): Nová lokalita švihlíku krutiklasu (Spiranthes spiralis) na Sušicku. – Památ. a Přír. 7: 127.
bartareau t. (1995): Pollination limitation, costs of capsule production and the capsule-to-flower ratio in Dendrobium monophylum F. Muell. (Orchidaceae). – Austr. J. Ecol. 20: 257–265.
brabec j., křenová z. & nesvaDbová j. (2004): Švihlík krutiklas – pozoruhodný druh květeny ČR. – Živa 2004/5: 209–211.
hutchinGs M. j. (1987): The population biology of the early spider orchid, Ophrys sphegodes Mill. II. Temporal patterns in behaviour. – J. Ecol. 75: 729–742.
hutchinGs M. j. (2010) The population biology of the early spider orchid, Ophrys sphegodes Mill. III. Demography across three decades. – J. Ecol. 98: 867–878.
iPser z. (2010): Využití obrazové analýzy při monitoringu kriticky ohroženého druhu Spiranthes spiralis. – Ms. [Bakalářská pr., depon in: Katedra biologických disciplín, Zemědělská fakulta Jihočeské univerzity, České Budějovice.]
jacQueMyn h. & hutchinGs M. j. (2010): Biological flora of the British Isles: Spiranthes spiralis (L.) Chevall. – J. Ecol. 98: 1253–1267.
jacQueMyn h., brys r. & jonGejans e. (2010): Size-dependent flowering and costs of reproduction affect population dynamics in a tuberous perennial woodland orchid. – J. Ecol. 98: 1204–1215.
jacQueMyn h., brys r., herMy M. & willeMs j. h. (2007): Long-term dynamics and population viability in one of the last populations of the endangered Spiranthes spiralis (Orchidaceae) in the Netherlands. – Biol. Conserv. 134: 14–21.
jersáková j. & kinDlMann P. (2004): Zásady péče o orchidejová stanoviště. – Kopp, České Budějovice.
kinDlMann P. & balounová z. (2001): Irregular flowering patterns in terrestrial orchids: theories vs empirical data. – Web Ecol. 2: 75–82.
kubát k., hrouDa l., chrtek j. jun., kaPlan z., kirschner j. & ŠtěPánek j. [eds] (2002): Klíč ke květeně České republiky. – Academia, Praha.
kuČera l. (1980): Nová lokalita švihlíku krutiklasu (Spiranthes spiralis /L./ Chevall.) v západních Čechách. – Zprav. Západočes. Poboč. Čs. Bot. Společ. 1980/2: 6–7.
kuČera l. (1982): Příroda Sušicka a její ochrana. – Městské kulturní středisko v Sušici, Sušice.kull t. (2002): Population dynamics of north temperate orchids. – In: kull t. & arDitti j.
[eds], Orchid Biology: Review and Perspectives, Vol. VIII, pp. 139–165, Kluwer Academic Publishers, Dordrecht.
Machon n., barDin P., Mazer s. j., Moret j., GoDelle b. & austerlitz f. (2003): Relationship between genetic structure and seed and pollen dispersal in the endangered orchid Spiranthes spiralis. – New Phytol. 157: 677–687.
Matějková i. & nesvaDbová j. (1999): Změny ve vegetačních poměrech NPP Pastviště u Fínů po desetiletém řízeném obhospodařování. – Erica 8: 93–116.
Moravec j. et al. (1995): Rostlinná společenstva ČR a jejich ohrožení. Ed. 2. – Severočeskou přírodou, Příloha 1995, Okresní Muzeum Litoměřice.
nesvaDbová j. & Pivoňková l. (1996): Monitoring populace Spiranthes spiralis (L.) Chevall. na trvalých plochách v národní přírodní památce Pastviště u Fínů. – Příroda, Praha, 6: 95–104.
nesvaDbová j., brabec j., Matějková i. & křenová z. (2003) [2006]: Populační biologie Spiranthes

200
Brabec et al.: Populační dynamika, energetická náročnost kvetení a přežívání Spiranthes spiralis
spiralis (L.) Chevall. na tradičně obhospodařované lokalitě „NPP Pastviště u Fínů“. – In.: PivniČková M. [ed.], Sborník dílčích zpráv z grantového projektu VaV 610/10/00 „Vliv hospodářských zásahů na změnu v biologické rozmanitosti ve zvláště chráněných územích“, Příroda, supplementum, pp. 189–195, 386.
nesvaDbová j., ČeČil f., Gryc j., hostiČka M., kraft j., neDorostová b., sofron j., vaněČek j., vonDráČek M. & žán M. (1987): Chráněný přírodní výtvor Pastviště U Fínů. – Ms. [Inventarizační průzkum; depon. in: Agentura ochrany přírody a krajiny ČR, středisko Plzeň.]
nesvaDbová j., sofron j. & vonDráČek M. (1989): Die Vegetation des geschütztes Naturgebildes Weide „U Fínů“ (Südwest Böhmen, Gebiet von Sušice). – Folia Mus. Rer. Natur. Bohem. Occid. 29: 1–42.
PetříČek v. [ed.] (1999): Péče o chráněná území I. Nelesní společenstva. – Agentura ochrany přírody a krajiny ČR, Praha.
PriMack r. b. & stacy e. (1998): Cost of reproduction in the pink lady’s slipper orchid (Cypripedium acaule, Orchidaceae): an eleven-year experimental study of three populations. – Am. J. Bot. 85: 1672–1679.
Procházka f. & velísek v. (1983): Orchideje naší přírody. – Praha.Procházka f. [ed.] (2001): Černý a červený seznam cévnatých rostlin České republiky (stav
v roce 2000). – Příroda, Praha, 18: 1–166.r DeveloPMent core teaM (2009): R: A language and environment for statistical computing. –
Vienna.shefferson r. P. (2009): The evolutionary ecology of vegetative dormancy in mature herbaceous
perennial plants. – J. Ecol. 97: 1000–1009.shefferson r. P., kull t. & tali k. (2005): Adult whole-plant dormancy induced by stress in long-
lived orchids. – Ecology 86: 3099–3104.shefferson r. P., ProPer j., beissinGer s. r. & siMMs e. l. (2003): Lifehistory trade-offs in a rare
orchid: the costs of flowering, dormancy and sprouting. – Ecology 84: 1199–1206.slaDký j. & liŠková D. (2004): Plán péče pro Národní přírodní památku Pastviště u Fínů a její
ochranné pásmo na období 2005–2014. – Ms. [Depon. in: Agentura ochrany přírody a krajiny České republiky, středisko Plzeň.]
snow a. a. & whiGhaM D. f. (1989): Costs of flower and fruit production in Tipularia discolor (Orchidaceae). – Ecology 70: 1286–1293.
statsoft (2011): Electronic Statistics Textbook. – Tulsa, http://www.statsoft.com/textbook/.tolasz r., Míková t., valeriánová a. & voženílek v. [eds] (2007): Atlas podnebí Česka. – Český
hydrometrorologický ústav a Univerzita Palackého v Olomouci.wells t. c. e. (1967): Changes in a population of Spiranthes spiralis (L.) Chevall. at Knocking
Hoe National Nature Reserve, Bedfordshie, 1962–65. – J. Ecol. 55: 83–99.wells t. c. e. (1981): Population ecology of terrestrial orchids. – In: h. synGe [ed.], The
biological aspects of rare plant conservation, pp. 281–295, Wiley & Sons, Chichester, New York, Brisbane, Toronto.
wells t. c. e., rothery P., coX r. & baMforD s. (1998): Flowering dynamics of Orchis morio L. and Herminium monorchis (L.) R. Br. at two sites in eastern England. – Bot. J. Linn. Soc. 126: 39–48.
willeMs j. h. & DorlanD e. (2000): Flowering frequency and plant performance and their relation to age in the perennial orchid Spiranthes spiralis (L.) Chevall. – Plant Biol. 2: 344–349.
willeMs j. h. & lahtinen M. l. (1997): Impact of pollination and resource limitation on seed production in a border population of Spiranthes spiralis (Orchidaceae). – Acta Bot. Neerl. 46: 365–375.
willeMs j. h. (1989): Population dynamics of Spiranthes spiralis in South-Limburg, The Nederlands. – Mém. Soc. Roy. Bot. Belg. 11: 115–121.
willeMs j. h., balounová z. & kinDlMann P. (2001): The effect of experimental shading on seed production and plant survival in the threatened species Spiranthes spiralis (Orchidaceae). – Lindleyana 16: 31–37.

Příroda, Praha, 31: 201–225, 2011
201
stanovištní nároky a výběr hostitelů u poloparazitů rodu lněnka (Thesium)Habitat requirements and host selectivity in hemiparasites from the genus Thesium
toMáš dostáLek1, 2 & ZuZana MünZbergová1, 2
1Botanický ústav Akademie věd ČR, Zámek 1, CZ-252 43 Průhonice; [email protected] botaniky, Přírodovědecká fakulta Univerzity Karlovy, Benátská 2, CZ-128 01 Praha 2
Abstract: The aim of this study was to identify the habitat requirements and host specificity of critically endangered species Thesium ebracteatum that is a root hemiparasite. We conducted a vegetation survey and tested whether some plant species occur significantly more often in plots with Thesium than expected, thereby representing potential hosts of hemiparasites. To test whether the potential hosts are the species to which Thesium is physically attached we conducted excavating and sowing experiment. Such kind of experiment, however, could endanger performance of populations of rare species and that is why we used related common T. linophyllon for this experiment. Thesium bavarum was added to our study as another rare species for comparison of results. Moreover we studied population performance in two populations of T. ebracteatum – Velenka in the Czech Republic and Schönwalde in Germany.
We found positive association with surrounding species for T. linophyllon but not for the other two species. The number of haustoria of T. linophyllon attached to roots of host species was significantly affected by the number of roots of those species, but also by their identity (species, family and plant group). Thesium linophyllon was, however, attached to 94% of all species occurring in the plots. We suggest that Thesium species do not specifically select hosts but rather occur in microhabitats with specific conditions. This conclusion is also supported by the fact that there were differences in microhabitats with and without studied Thesium species. We found microhabitats with T. ebracteatum to have higher values for soil reaction in comparison with microhabitats without T. ebracteatum, and microhabitats with T. linophyllon to be lighter, drier, and nutrient poorer in comparison with microhabitats without T. linophyllon. We also found high spatial and temporal variability in the population performance of T. ebracteatum, which was probably caused by climate fluctuation or local disturbances. In Czech population Velenka we observed systematic decline in the number of shoots. In German population Schönwalde we observed a negative effect of an extremely dry spring in 2007 on the proportion of fertile shoots.Keywords: hemiparasite, host range, host selectivity, host specificity, Santalales, Thesium ebracteatum
Abstrakt: Cílem této studie bylo identifikovat stanovištní nároky a hostitelské druhy kriticky ohroženého kořenového poloparazita Thesium ebracteatum. Snažili jsme se určit, zda existují druhy, které se průkazně častěji vyskytují v plochách s Thesium, než bychom očekávali náhodou a které mohou sloužit jako potenciální hostitelé tohoto poloparazita. Pro testování toho, zda potenciální hostitelé jsou opravdu druhy, na které je Thesium fyzicky napojeno, jsme použili vykopávací a výsevový experiment. Ten by však mohl ohrozit vitalitu populací

vzácného druhu, a tak jsme pro tento pokus použili příbuzný hojný druh T. linophyllon. Dále jsme ještě do naší studie pro srovnání zahrnuli vzácný druh T. bavarum. Navíc jsme ještě sledovali variabilitu v populační dynamice T. ebracteatum na dvou lokalitách – Velenka v ČR a Schönwalde v SRN.
V naší práci jsme nalezli pozitivní asociaci s okolními druhy pro T. linophyllon, ne však pro ostatní dva druhy z rodu Thesium. Počet haustorií T. linophyllon přichycených na kořeny hostitelských druhů byl průkazně ovlivněn množstvím kořenů těchto druhů, ale také jejich identitou (druh, čeleď a skupina rostlin). Thesium linophyllon však bylo přichyceno k 94 % všech druhů vyskytujících se ve studovaných plochách. Předpokládáme proto, že druhy rodu Thesium si nijak specificky své hostitele nevybírají, ale spíše se vyskytují na mikrostanovištích se specifickými podmínkami. Tento závěr je podpořen i skutečností, že jsme nalezli rozdíly mezi mikrostanovišti, kde rostly a nerostly jednotlivé studované druhy Thesium. Stanoviště s T. ebracteatum měla průkazně vyšší hodnoty pro půdní reakci než stanoviště bez T. ebracteatum a stanoviště s T. linophyllon byla průkazně světlejší, sušší a živinami chudší než stanoviště bez T. linophyllon. Nalezli jsme také velkou prostorovou i časovou variabilitu v populační dynamice T. ebracteatum, která je pravděpodobně způsobena změnami počasí a lokálními disturbancemi. V české populaci Velenka jsme pozorovali soustavný pokles v počtu prýtů ve sledovaných plochách. V německé populaci Schönwalde jsme zaznamenali negativní vliv suchého jara 2007 na proporci fertilních prýtů.Klíčová slova: poloparazité, Santalales, specificita, Thesium ebracteatum, výběr hostitelů
Úvod
Parazitické rostliny představují specifickou skupinu rostlinných druhů a už dlouhou dobu jsou předmětem zájmu mnoha studií (např. Gibson & watkinson 1989, Press & Graves 1995, aDler 2003, PhoeniX & Press 2005a,b, Mueller & GehrinG 2006). I přes relativně veliké množství publikované literatury však víme překvapivě málo o ekologii kořenových (polo)parazitických druhů. Tento nedostatek znalostí je zjevně spojen s obtížemi při studiu podzemních spojení mezi parazity a jejich hostiteli.
V literatuře o kořenových poloparazitech můžeme najít poměrně hodně informací o druzích z čeledi Orobanchaceae (např. Gibson & watkinson 1989, 1991, 1992, Matthies 1996, joshi et al. 2000, PhoeniX & Press 2005b, ren et al. 2010), ale znalosti o dalších skupinách téměř chybí. Kořenoví poloparazité z řádu Santalales jsou velice málo prozkoumanou skupinou, která obsahuje jak významné plevele, tak i mnoho ohrožených druhů (MusselMan & haynes 1996, Marvier & sMith 1997, libby & blooM 1998). Znalost jejich biologie je tedy klíčová jak pro jejich likvidaci, tak pro efektivní ochranu. V řádu Santalales můžeme najít druhy, které parazitují pouze na úzkém okruhu hostitelských rostlin, jako jsou třeba někteří zástupci z rodu Viscum (zuber & wiDMer 2000), ale i druhy, které parazitují na velkém množství rostlin, jako je například druh Osyris alba (QaseM 2006). Avšak u mnohých dalších druhů z řádu Santalales máme pouze velice málo informací o jejich hostitelích nebo je tato informace dokonce úplně neznámá.
Rod Thesium zahrnuje bylinné i dřevinné zástupce kořenových poloparazitů z čeledi Santalaceae a je široce rozšířen v oblasti Starého světa (PilGer 1935), přičemž na území Evropy se vyskytuje z tohoto rodu 18 druhů (henDrych 1980). Někteří zástupci rodu Thesium patří v Evropě mezi zemědělské plevele (T. humile − fer et al. 1993), zatímco jiní patří mezi kriticky ohrožené druhy (T. ebracteatum – euroPean coMMission 1992). Zástupci rodu Thesium jsou schopni vytvořit haustoria na široké škále krytosemenných rostlin (Themeda a Poa – scarlett et al. 2003, Galium − renauDin et al. 1981, ječmen
202
Dostálek & Münzbergová: Stanovištní nároky a výběr hostitelů u rodu Thesium

a cibule – abu-irMaileh 1980, vinná réva – DasGuPta 1988), avšak je velice málo známé, jak si Thesium skutečně vybírá hostitele ve svých přirozených populacích. Existuje jediná studie, která se zabývá výběrem hostitelů druhu Thesium chinense v jeho přirozených populacích (suetsuGu et al. 2008). suetsuGu et al. (2008) poprvé využili nepřímou metodu identifikace hostitelů u druhu z čeledi Santalaceae a sledovali, zda jsou některé druhy rostlin, které se vyskytují častěji s druhem T. chinense a mohly by tak sloužit jako potenciální hostitelé tohoto druhu. Potom použili i přímou metodu, kdy vykopávali kořeny T. chinense a zkoumali přímé napojení na hostitelské druhy pomocí haustorií. Zjistili, že pouze dva z 38 celkově zaznamenaných druhů, Lespedeza juncea a Eragrostis curvula, se vyskytovaly v plochách s Thesium častěji než bychom mohli očekávat jen náhodou. Podle jejich studie T. chinense parazituje na 22 druzích z 11 čeledí, což odpovídá téměř 58 % všech druhů rostoucích na lokalitě, kde studie probíhala. Haustoria však byla nenáhodně rozmístěna mezi hostitelskými druhy, což ukazuje na určitou míru selektivity ve výběru hostitelů. Thesium chinense obecně preferovalo druhy z čeledi Poaceae, přestože haustoria vytvořená na kořenech druhů z čeledi Fabaceae byla v porovnání s haustorii vytvořenými na ostatních druzích větší. Tato studie (suetsuGu et al. 2008) byla úplně první, která se zabývala specifičností vazby na hostitele u druhu z rodu Thesium. Byla však provedena pouze na jedné lokalitě a pouze na jedné prostorové škále (30 × 30 cm). Obecné závěry proto mohou být nejisté. Navíc zůstává neznámá prostorová škála, na které se výběr hostitelů odehrává.
Naším prvním cílem bylo identifikovat možné hostitelské druhy pro v ČR kriticky ohrožený druh Thesium ebracteatum a zjistit, jak se výsledky liší mezi jednotlivými lokalitami. Na základě předchozích studií jsme vytvořili následující hypotézy: 1) Thesium bude průkazně více asociováno s některými hostitelskými druhy v nadzemní vegetaci a tyto druhy budou stejné jako druhy, se kterými je Thesium propojeno i pod zemí, 2) rozdíly mezi hostitelskými druhy na jednotlivých lokalitách budou způsobeny velkým rozsahem oblasti studie, tyto rozdíly však zmizí, když v analýze použijeme místo druhů vyšší taxonomické jednotky (např. čeledi).
Nepřímá metoda identifikace hostitelů je nedestruktivní a může být jednoduše použita i pro vzácné druhy. Použití přímé metody je však omezeno pouze na hojné druhy, protože by při jejím použití mohlo dojít k ohrožení vitality zejména menších populací. Při práci se vzácnými druhy však můžeme testovat užitečnost nepřímé metody na příbuzném hojném druhu a tuto znalost pak aplikovat také u druhu vzácného. Abychom otestovali naše předpoklady, použili jsme kromě kriticky ohroženého Thesium ebracteatum také vzácný druh T. bavarum, a v teplejších oblastech relativně běžný druh T. linophyllon. Pro určení potenciálních hostitelů jsme použili nepřímou metodu (zda jsou některé druhy ve vegetaci více asociovány s parazitem) pro všechny tři druhy z rodu Thesium. Abychom určili, zda jsou potenciální hostitelé skutečně také druhy, na které se Thesium haustorii přichycuje, použili jsme též přímou metodu (hledání fyzického napojení parazita haustorii na hostitele) u T. linophyllon. Avšak ani nalezení přímého fyzického napojení na hostitelskou rostlinu nemusí znamenat, že je poloparazit závislý na tomto druhu. Toto musí být prokázáno s použitím manipulativního experimentu, při kterém se prokáže, zda rostliny poloparazita rostou lépe při napojení na svého hostitele, než když rostou samostatně. Provedli jsme proto takovýto experiment s hojným T. linophyllon v experimentální zahradě.
Druhým cílem bylo použít data z nepřímé metody identifikace hostitelských druhů k odvození ekologických nároků jednotlivých druhů rodu Thesium v rámci jejich lokalit a porovnat ekologické nároky jednotlivých studovaných druhů Thesium. Tyto dva cíle projektu byly předmětem předchozí práce publikované v anglickém jazyce (Dostálek & MünzberGová 2010), z níž tento článek vychází.
Příroda, Praha, 31: 201–225, 2011
203
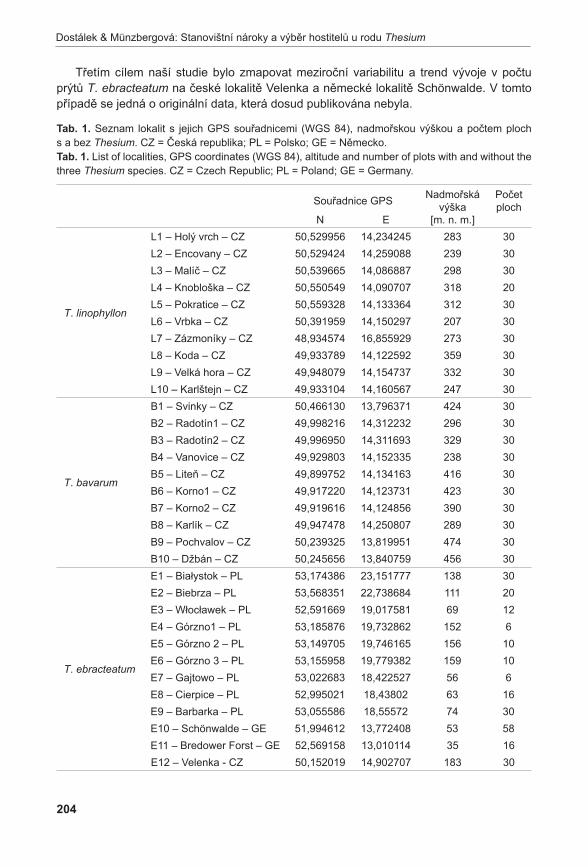
Třetím cílem naší studie bylo zmapovat meziroční variabilitu a trend vývoje v počtu prýtů T. ebracteatum na české lokalitě Velenka a německé lokalitě Schönwalde. V tomto případě se jedná o originální data, která dosud publikována nebyla.
tab. 1. Seznam lokalit s jejich GPS souřadnicemi (WGS 84), nadmořskou výškou a počtem ploch s a bez Thesium. CZ = Česká republika; PL = Polsko; GE = Německo.tab. 1. List of localities, GPS coordinates (WGS 84), altitude and number of plots with and without the three Thesium species. CZ = Czech Republic; PL = Poland; GE = Germany.
Souřadnice GPS Nadmořská výška
Počet ploch
N E [m. n. m.]
T. linophyllon
L1 – Holý vrch – CZ 50,529956 14,234245 283 30L2 – Encovany – CZ 50,529424 14,259088 239 30L3 – Malíč – CZ 50,539665 14,086887 298 30L4 – Knobloška – CZ 50,550549 14,090707 318 20L5 – Pokratice – CZ 50,559328 14,133364 312 30L6 – Vrbka – CZ 50,391959 14,150297 207 30L7 – Zázmoníky – CZ 48,934574 16,855929 273 30L8 – Koda – CZ 49,933789 14,122592 359 30L9 – Velká hora – CZ 49,948079 14,154737 332 30L10 – Karlštejn – CZ 49,933104 14,160567 247 30
T. bavarum
B1 – Svinky – CZ 50,466130 13,796371 424 30B2 – Radotín1 – CZ 49,998216 14,312232 296 30B3 – Radotín2 – CZ 49,996950 14,311693 329 30B4 – Vanovice – CZ 49,929803 14,152335 238 30B5 – Liteň – CZ 49,899752 14,134163 416 30B6 – Korno1 – CZ 49,917220 14,123731 423 30B7 – Korno2 – CZ 49,919616 14,124856 390 30B8 – Karlík – CZ 49,947478 14,250807 289 30B9 – Pochvalov – CZ 50,239325 13,819951 474 30B10 – Džbán – CZ 50,245656 13,840759 456 30
T. ebracteatum
E1 – Białystok – PL 53,174386 23,151777 138 30E2 – Biebrza – PL 53,568351 22,738684 111 20E3 – Włocławek – PL 52,591669 19,017581 69 12E4 – Górzno1 – PL 53,185876 19,732862 152 6E5 – Górzno 2 – PL 53,149705 19,746165 156 10E6 – Górzno 3 – PL 53,155958 19,779382 159 10E7 – Gajtowo – PL 53,022683 18,422527 56 6E8 – Cierpice – PL 52,995021 18,43802 63 16E9 – Barbarka – PL 53,055586 18,55572 74 30E10 – Schönwalde – GE 51,994612 13,772408 53 58E11 – Bredower Forst – GE 52,569158 13,010114 35 16E12 – Velenka - CZ 50,152019 14,902707 183 30
204
Dostálek & Münzbergová: Stanovištní nároky a výběr hostitelů u rodu Thesium

Metodika
studované druhy
V Evropě má rod Thesium 18 druhů, které se vyskytují v širokém spektru stanovišť od nížin až po hory (henDrych 1980). Jako modelové druhy jsme v naší studii zvolili tři druhy: T. ebracteatum je v České republice kriticky ohrožený druh (holub & Procházka 2000) a je to také druh s vysokou prioritou ochrany v rámci celé Evropy (natura 2000; euroPean coMMission 1992). Dalším studovaným druhem bylo T. linophyllon, které je hojné na mnoha lokalitách v České republice a jeho populace čítají často tisíce jedinců. Kvůli jeho četnosti jsme u tohoto druhu mohli použít jak přímou tak nepřímou metodu identifikace hostitelů. Posledním druhem je T. bavarum, které je v České republice silně ohroženým druhem (holub & Procházka 2000).
Thesium ebracteatum Hayne je vytrvalý druh s centrem rozšíření ve východní Evropě (Pobaltí, Bělorusko, Rusko až po Ural, severní Ukrajina, Moldavsko a střední Rumunsko) a jednotlivými výskyty zasahujícími až do střední Evropy (Polsko, severní Německo, Čechy, Rakousko, kdysi na Slovenku). V ČR byla především malá arela ve středním Polabí, z níž se dochovala jediná lokalita u Velenky (henDrych 1969b). V roce 2008 byla nalezena další malá lokalita u obce Běštín na okraji Brd, která však není zahrnuta v naší studii (Prach & zajíČková 2009). Tento druh patří mezi rostliny vlhkých, rašelinných nebo slatinných luk a pastvin, lesních světlin a světlých hájů (Grulich in slavík 1997, Grulich in kubát et al. 2002). V Polsku a Německu roste i ve světlých lesích na písku u cest a železničních tratí (T. Dostálek, osobní pozorování). Druh je hemikryptofyt, jehož tenký oddenek tvoří plazivé výběžky. Kvete od května do června (Grulich in kubát et al. 2002).
Thesium linophyllon L. je vytrvalá rostlina, rostoucí často na suchých travnatých nebo kamenitých stráních nebo ve světlých listnatých lesích (Grulich in slavík 1997, Grulich in kubát et al. 2002). Je naším nejrozšířenějším zástupcem z rodu Thesium. Je to druh s centrem areálu ve střední Evropě, který je znám z Itálie, Německa, Rakouska, ČR, SR, Maďarska, Polska, Rumunska, Balkánského poloostrova a Ukrajiny (henDrych 1969a). Je to hemikryprofyt, který tvoří výběžkatý oddenek a kvete v květnu až srpnu (Grulich in kubát et al. 2002).
Thesium bavarum Schrank je vytrvalý druh vyskytující se vzácně v pahorkatinách středních a severních Čech na stepních svazích a ve světlých lesích (Grulich in slavík 1997, Grulich in kubát et al. 2002). Jeho rozšíření se táhne od Francie, přes Itálii, Německo, ČR, Rakousko, Balkánský poloostrov až do Rumunska a Bulharska (henDrych 1969a). Rostliny mají nevýběžkatý mnohohlavý oddenek. Je to rovněž hemikryptofyt a kvete také v květnu až srpnu (Grulich in kubát et al. 2002).
studované lokality
Thesium ebracteatum bylo studováno na poslední větší recentní lokalitě v ČR u obce Velenka ve středních Čechách (Čeřovský 1999). Dále bylo studováno v Braniborsku v SRN na lokalitách Schönwalde a Bredower Forst; v Polsku na lokalitách Biebrza (Biebrzański Park Narodowy), Białystok (SV Polsko), na třech lokalitách v Górznieńsko-Lidzbarski Park Krajobrazowy a čtyřech lokalitách poblíž Toruně ve středním Polsku. Thesium linophyllon bylo studováno pouze v České republice (šest populací v Českém středohoří, tři populace v Českém krasu a jedna populace v PP Zázmoníky na jižní Moravě). V těchto územích se T. linophyllon vyskytuje hojně. Thesium bavarum bylo studováno na sedmi lokalitách v Českém krasu, dvou lokalitách v PP Džbán a jedné lokalitě v Českém středohoří (tab. 1).
Příroda, Praha, 31: 201–225, 2011
205

Velikosti populací sahaly od stovek po desítky tisíc ramet pro T. ebracteatum, od tisíců po desítky tisíc ramet pro T. linophyllon a od desítek až po stovky trsů pro T. bavarum. Vzdálenost mezi jednotlivými populacemi byla několik km až 700 km pro T. ebracteatum (zahrnuty byly populace v ČR, SRN a Polsku) a několik km až 250 km pro T. linophyllon a T. bavarum.
stanovištní nároky
Abychom určili stanovištní nároky modelových druhů z rodu Thesium, zaznamenávali jsme vegetační složení (pokryvnost každého druhu v %) ve čtvercích 20 × 20, 50 × 50 a 100 × 100 cm. Čtverce byly umístěny na transektech přes každou ze sledovaných lokalit a menší velikosti čtverce byly umístěny uvnitř větších. Na transektech jsme střídali plochy ve vegetaci s a bez přítomnosti Thesium. Minimální vzdálenost mezi dvěmi plochami byla jeden metr. Na každé lokalitě jsme zaznamenali, pokud to bylo možné, pokryvnosti v 15 plochách s Thesium a v 15 plochách bez Thesium (tab. 1). Použili jsme tři velikosti ploch, protože délka rhizomů těchto druhů není známá a chtěli jsme se dozvědět, ve které velikosti ploch se nejlépe projeví stanovištní nároky a potenciální hostitelé.
S použitím programu JUICE 7.0.24 (tichý 2002) jsme pro každou plochu vypočítali průměrné Ellenbergovy indikační hodnoty (ellenberG et al. 2001) pro světlo, teplotu, kontinentalitu, vlhkost, půdní reakci a živiny založené na druhovém složení. Spočítali jsme také počet druhů v každé ploše. Pak jsme testovali rozdíly mezi Ellenbergovými indikačními hodnotami a počtem druhů mezi plochami s a bez Thesium po odečtení vlivu lokality. Dále jsme testovali i interakci mezi přítomností Thesium a lokalitou, abychom zjistili, zda se výše zjištěné rozdíly liší mezi lokalitami. Sledovali jsme také rozdíly mezi počty druhů v plochách s a bez Thesium po odečtení vlivu podmínek prostředí reprezentovaných Ellenbergovými indikačními hodnotami. Pro testování byla použita lineární regrese v programu s-Plus (2000). Thesium bylo vyloučeno z dat při výpočtu průměrných Ellenbergových indikačních hodnot a počtu druhů na plochu.
Abychom mohli porovnat stanovištní nároky tří druhů z rodu Thesium, testovali jsme také rozdíly v Ellenbergových indikačních hodnotách a počtech druhů mezi plochami s různými druhy Thesium. Protože rozdíly mezi plochami s různými druhy Thesium by mohly být dány pouze rozdíly mezi jednotlivými lokalitami, mezi různými vlivy jednotlivých druhů Thesium na okolní vegetaci nebo rozdílným výběrem mikrostanovišť jednotlivými druhy Thesium, testovali jsme také rozdíly v Ellenbergových indikačních hodnotách a počtech druhů mezi plochami bez přítomnosti Thesium mezi lokalitami jednotlivých druhů Thesium. Test byl proveden pomocí ANOVA v programu S-PLUS (2000) s lokalitami vřazenými do druhů. Ellenbergovy indikační hodnoty pro světlo, teplotu, kontinentalitu, vlhkost, půdní reakci a živiny jsou 7, 6, 6, 4, 2, 2 pro T. ebracteatum, 8, 7, 5, 2, 8, 1 for T. linophyllon a 7, 6, 4, 3, 8, 2 pro T. bavarum (ellenberG et al. 2001).
Potenciální asociace s hostiteli
Data o vegetačním složení (viz výše) byla použita rovněž k identifikaci druhů, které mají tendenci častěji se vyskytovat s Thesium. Použili jsme kanonickou korespondenční analýzu (CCA) a testovali jsme vliv přítomnosti Thesium na druhové složení. Naše hypotéza byla, že jsou druhy, které jsou pozitivně a negativně asociovány s Thesium a že se neliší na vybraných prostorových škálách (20 × 20 cm, 50 × 50 cm a 100 × 100 cm). Počítali jsme podíl variability v druhovém složení vysvětlený přítomností Thesium k celkové variabilitě v druhovém složení. Lokality byly použity jako kovariáty a provedli jsme randomizační test v blocích, které byly určeny kovariátami (lePŠ & ŠMilauer 2003). Provedli jsme také test interakce mezi přítomností Thesium a lokalitou, přičemž jsme použili lokalitu a přítomnost
206
Dostálek & Münzbergová: Stanovištní nároky a výběr hostitelů u rodu Thesium

Thesium jako kovariáty a jejich interakci jako nezávislou proměnnou. Průkazná interakce by znamenala, že na každé lokalitě jsou jiné druhy pozitivně nebo negativně asociované s Thesium. Data o pokryvnosti jednotlivých druhů byla odmocninově transformována a byla snížena hodnota vzácných druhů (lePŠ & ŠMilauer 2003).
Všechny výše popsané testy byly provedeny pro každý ze studovaných druhů Thesium a pro každou velikost plochy zvlášť. Testy byly také zopakovány a místo druhů byly použity rostlinné čeledi, abychom viděli, jak se výsledky změní s nižším taxonomickým rozlišením. Rozřazení druhů do čeledí bylo provedeno podle International Plant Names Index (IPNI; http://www.ipni.org). Pro každý druh (čeleď) byla také spočítána hodnota N2, což je převrácená hodnota Simpsonova indexu (siMPson 1949), která popisuje, jak často se druh (čeleď) v datovém souboru vyskytuje. Druhy s N2 nižší než tři a čeledi s N2 nižší než pěti byly vyloučeny z konečných analýz (tj. druhy s nízkým počtem výskytů nebo s nízkou pokryvností) a nejsou tedy uváděny ve výsledcích. Použili jsme tedy výsledky pouze ze 146 nejběžnějších druhů a 35 nejběžnějších čeledí pro T. ebracteatum, 155 druhů a 35 čeledí pro T. linophyllon a 152 druhů a 37 čeledí pro T. bavarum (pro velikost ploch 100 cm × 100 cm). Mnohorozměrné analýzy a výpočet hodnot N2 byly provedeny v programu Canoco for Windows 4.5 (ter braak & ŠMilauer 2002). Stejné analýzy byly také provedeny samostatně pro lokality T. linophyllon L1, L2 a L3 (tab. 1). V tomto případě byly vyloučeny druhy a čeledi s N2 nižší než tři.
Testovali jsme také vztah mezi přítomností každého z druhů Thesium ve třech velikostech ploch a pokryvností graminoidů (druhy z čeledí Cyperaceae a Poaceae), druhů z čeledi Fabaceae a druhů z ostatních čeledí. Data o pokryvnosti druhů byla logaritmicky transformována. K testu jsme použili logistickou regresi s přítomností/nepřítomností Thesium jako závislou proměnnou. Test byl proveden v programu S-PLUS (2000).
Fyzická asociace s hostiteli
Fyzické spojení mezi Thesium a jeho hostiteli jsme studovali pouze u druhu T. linophyllon, protože tento typ studie je destruktivní a nemůže být prováděn v populacích vzácných druhů bez rizika ohrožení vitality jejich populací. Abychom prozkoumali napojení haustorii na hostitelské druhy, vykopali jsme na třech lokalitách v Českém středohoří (L1, L2, L3) deset drnů o velikosti 20 × 20 × 15 (hloubka) cm, které obsahovaly přibližně pět ramet T. linophyllon. Každý drn byl rozdělen na čtyři části (10 cm × 10 cm × 15 cm). Potom byly drny pečlivě vymyty vodou a hostitelské druhy byly určeny pomocí sledování kořenů k nadzemní části. Po spočtení počtu haustorií na každém rostlinném druhu v každé části drnu, jsme odhadli množství kořenů každého druhu v každé ze čtyř částí drnu na stupnici 0–5 (0 – žádné kořeny, 5 – kořeny přítomné všude v drnu). Počet haustorií a množství kořenů v každém drnu bylo sečteno přes čtyři části drnu. Konečné množství kořenů mohlo tedy v drnu 20 cm × 20 cm × 15 cm sahat na stupnici od 0 do 20.
Testovali jsme vztah mezi množstvím kořenů každého druhu v ploše a počtem haustorií (logaritmicky transformováno) přichycených na tomto druhu po odečtení vlivu lokality. Testovali jsme také vliv čeledi, druhu včleněného do čeledi (druhy s méně než šesti záznamy byly z analýzy vyloučeny), růstové formy (graminoidy, tj. Poaceae a Cyperaceae, vs. druhy z ostatních čeledí a graminoidy vs. Fabaceae vs. druhy z ostatních čeledí) na počet haustorií. Množství kořenů každého druhu a lokalita byly použity jako kovariáty. Všechny testy byly provedeny s použitím GLM s Poissonovým rozdělením v programu S-PLUS (2000).
V případě průkazné asociace Thesium s potenciálními hostiteli v datech o vegetačním složení (popsáno výše) jsme testovali, zda je průkazná shoda mezi druhy (čeleděmi)
Příroda, Praha, 31: 201–225, 2011
207

určenými jako potenciální hostitelé a druhy (čeleděmi) s fyzickým propojením s Thesium. Technicky jsme testovali korelaci mezi skóry na první. ose v analýze CCA (ukazující, jak jsou druhy asociovány s Thesium) a průměrným počtem haustorií na kořenech testovaného druhu. Tato analýza byla provedena pouze pro tři lokality T. linophyllon, kde jsme sbírali data jak o potenciálních hostitelích, tak o fyzickém propojení s Thesium. Korelace na každé ze tří lokalit byla testována zvlášť. Analýzy byly provedeny v programu S-PLUS (2000).
výsevový experiment
V červenci 2007 jsme odebrali semena T. linophyllon ze tří populací v Českém středohoří (L1, L2, L3) a vyseli je do květináčů s potenciálními hostiteli. Jako potenciální hostitelé byly použity dominantní druhy na lokalitách: Bromus erectus, Brachypodium pinnatum, Carex flacca, Carex tomentosa a Inula salicina. Semena potenciálních hostitelů byla vyseta samostatně do květináčů v květnu 2007, tj. dva měsíce před přidáním semen T. linophyllon. Množství vysetých semen bylo zvoleno u každého druhu tak, aby hostitelská rostlina nezabrala více než 50 % plochy květináče. Pokud to bylo potřeba, tak byly některé rostliny hostitelských rostlin z květináče odstraněny. Do každého květináče jsme vyseli 100 semen T. linophyllon. Stejný počet semen jsme vyseli i do květináče bez vysetých hostitelských rostlin. Každá z variant měla deset opakování, což je dohromady 60 květináčů s 6 000 semeny T. linophyllon. Ve všech květináčích jsme zaznamenávali rostliny Thesium každých šest měsíců do června 2009. Pomocí logistické regrese jsme chtěli testovat, zda je více semenáčků v květináčích s hostitelskými rostlinami nebo bez nich. Dále jsme chtěli testovat, který z hostitelských druhů je nejvhodnější pro vzcházení a růst semenáčů. Vzhledem k celkově malé klíčivosti semen T. linophyllon, jsme data nakonec netestovali (viz Výsledky).
Meziroční variabilita v počtu prýtů
Ve dvou populacích T. ebracteatum (Velenka v ČR a Schönwalde v SRN) jsme v každé z pěti náhodně vybraných ploch o velikosti 50 × 50 cm v červnu 2006–2009 (v populaci Velenka byl odečet proveden i v roce 2010) zaznamenali počet sterilních a fertilních prýtů. Testovali jsme pak, zda se mezi jednotlivými roky liší počet všech prýtů a proporce fertilních prýtů. Pomocí GLM s Poissonovým rozdělením jsme testovali rozdíly mezi lokalitami, roky, plochami vřazenými do lokality a interakce mezi rokem a lokalitou v S-PLUS (2000); pro test proporce fertilních prýtů byla použita logistická regrese. V tomto testu jsme použili rok jako kategoriální proměnnou. V dalším testu jsme použili rok jako spojitou proměnnou a testovali jsme pro každou lokalitu zvlášť, zda se mění počet prýtů během studovaných let a zda je trend konzistentní mezi jednotlivými plochami.
výsledky
stanovištní nároky
Plochy s T. ebracteatum a T. linophyllon obsahovaly průkazně více druhů než plochy bez přítomnosti Thesium (obr. 1, tab. 2). Rozdíl mezi počtem druhů mezi plochami s a bez Thesium stoupal s velikostí plochy. Druhy přítomné v plochách s T. ebracteatum měly průkazně vyšší indikační hodnoty pro půdní reakci. Druhy přítomné v plochách s T. linophyllon měly průkazně vyšší indikační hodnoty pro světlo a nižší indikační hodnoty pro vlhkost a živiny (obr. 2, tab. 2). U druhu T. bavarum jsme však nenalezli žádný průkazný rozdíl mezi plochami s a bez přítomnosti Thesium kromě vyšších hodnot pro kontinentalitu v plochách 100 × 100 cm s T. bavarum (tab. 2, příloha 1). Nalezli
208
Dostálek & Münzbergová: Stanovištní nároky a výběr hostitelů u rodu Thesium
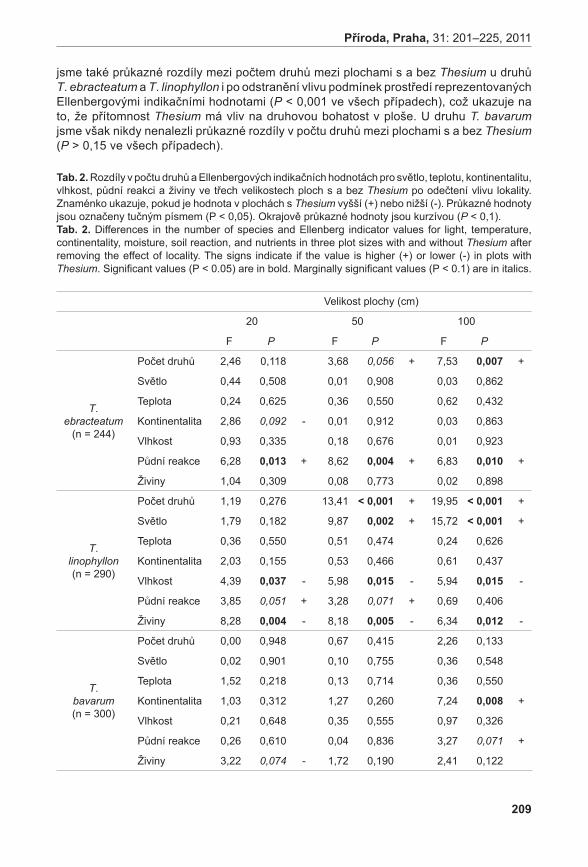
jsme také průkazné rozdíly mezi počtem druhů mezi plochami s a bez Thesium u druhů T. ebracteatum a T. linophyllon i po odstranění vlivu podmínek prostředí reprezentovaných Ellenbergovými indikačními hodnotami (P < 0,001 ve všech případech), což ukazuje na to, že přítomnost Thesium má vliv na druhovou bohatost v ploše. U druhu T. bavarum jsme však nikdy nenalezli průkazné rozdíly v počtu druhů mezi plochami s a bez Thesium (P > 0,15 ve všech případech).
tab. 2. Rozdíly v počtu druhů a Ellenbergových indikačních hodnotách pro světlo, teplotu, kontinentalitu, vlhkost, půdní reakci a živiny ve třech velikostech ploch s a bez Thesium po odečtení vlivu lokality. Znaménko ukazuje, pokud je hodnota v plochách s Thesium vyšší (+) nebo nižší (-). Průkazné hodnoty jsou označeny tučným písmem (P < 0,05). Okrajově průkazné hodnoty jsou kurzívou (P < 0,1).tab. 2. Differences in the number of species and Ellenberg indicator values for light, temperature, continentality, moisture, soil reaction, and nutrients in three plot sizes with and without Thesium after removing the effect of locality. The signs indicate if the value is higher (+) or lower (-) in plots with Thesium. Significant values (P < 0.05) are in bold. Marginally significant values (P < 0.1) are in italics.
Velikost plochy (cm)
20 50 100
F P F P F P
T. ebracteatum
(n = 244)
Počet druhů 2,46 0,118 3,68 0,056 + 7,53 0,007 +
Světlo 0,44 0,508 0,01 0,908 0,03 0,862
Teplota 0,24 0,625 0,36 0,550 0,62 0,432
Kontinentalita 2,86 0,092 - 0,01 0,912 0,03 0,863
Vlhkost 0,93 0,335 0,18 0,676 0,01 0,923
Půdní reakce 6,28 0,013 + 8,62 0,004 + 6,83 0,010 +
Živiny 1,04 0,309 0,08 0,773 0,02 0,898
T. linophyllon(n = 290)
Počet druhů 1,19 0,276 13,41 < 0,001 + 19,95 < 0,001 +
Světlo 1,79 0,182 9,87 0,002 + 15,72 < 0,001 +
Teplota 0,36 0,550 0,51 0,474 0,24 0,626
Kontinentalita 2,03 0,155 0,53 0,466 0,61 0,437
Vlhkost 4,39 0,037 - 5,98 0,015 - 5,94 0,015 -
Půdní reakce 3,85 0,051 + 3,28 0,071 + 0,69 0,406
Živiny 8,28 0,004 - 8,18 0,005 - 6,34 0,012 -
T.bavarum(n = 300)
Počet druhů 0,00 0,948 0,67 0,415 2,26 0,133
Světlo 0,02 0,901 0,10 0,755 0,36 0,548
Teplota 1,52 0,218 0,13 0,714 0,36 0,550
Kontinentalita 1,03 0,312 1,27 0,260 7,24 0,008 +
Vlhkost 0,21 0,648 0,35 0,555 0,97 0,326
Půdní reakce 0,26 0,610 0,04 0,836 3,27 0,071 +
Živiny 3,22 0,074 - 1,72 0,190 2,41 0,122
Příroda, Praha, 31: 201–225, 2011
209
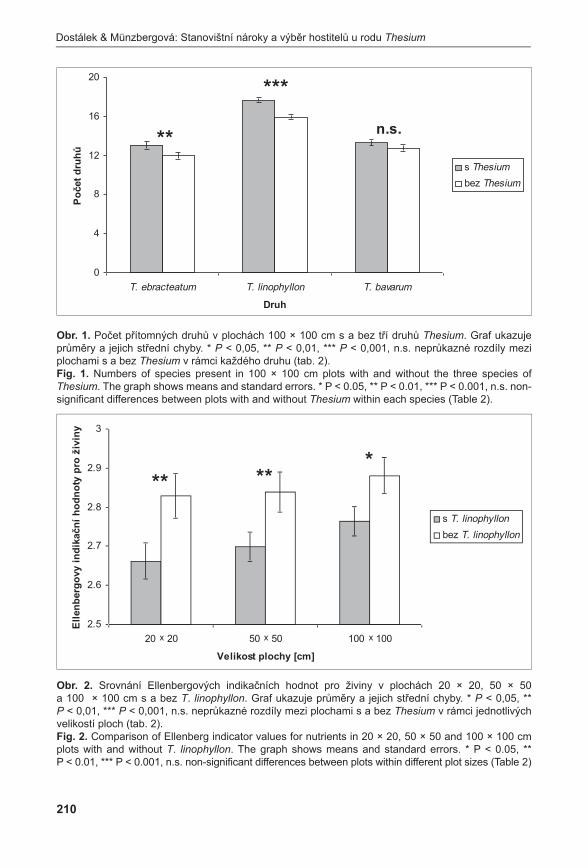
obr. 1. Počet přítomných druhů v plochách 100 × 100 cm s a bez tří druhů Thesium. Graf ukazuje průměry a jejich střední chyby. * P < 0,05, ** P < 0,01, *** P < 0,001, n.s. neprůkazné rozdíly mezi plochami s a bez Thesium v rámci každého druhu (tab. 2).Fig. 1. Numbers of species present in 100 × 100 cm plots with and without the three species of Thesium. The graph shows means and standard errors. * P < 0.05, ** P < 0.01, *** P < 0.001, n.s. non-signifi cant differences between plots with and without Thesium within each species (Table 2).
obr. 2. Srovnání Ellenbergových indikačních hodnot pro živiny v plochách 20 × 20, 50 × 50 a 100 × 100 cm s a bez T. linophyllon. Graf ukazuje průměry a jejich střední chyby. * P < 0,05, ** P < 0,01, *** P < 0,001, n.s. neprůkazné rozdíly mezi plochami s a bez Thesium v rámci jednotlivých velikostí ploch (tab. 2).Fig. 2. Comparison of Ellenberg indicator values for nutrients in 20 × 20, 50 × 50 and 100 × 100 cm plots with and without T. linophyllon. The graph shows means and standard errors. * P < 0.05, ** P < 0.01, *** P < 0.001, n.s. non-signifi cant differences between plots within different plot sizes (Table 2)
210
Dostálek & Münzbergová: Stanovištní nároky a výběr hostitelů u rodu Thesium
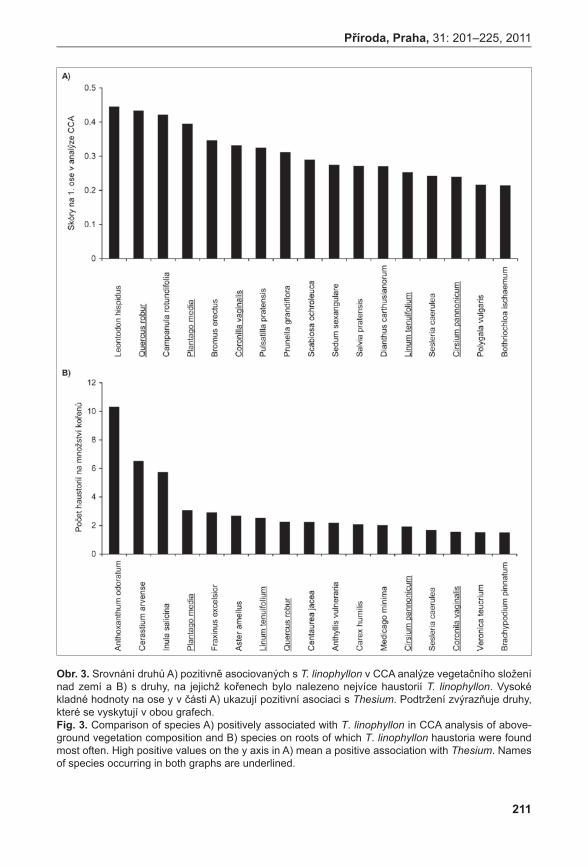
obr. 3. Srovnání druhů A) pozitivně asociovaných s T. linophyllon v CCA analýze vegetačního složení nad zemí a B) s druhy, na jejichž kořenech bylo nalezeno nejvíce haustorií T. linophyllon. Vysoké kladné hodnoty na ose y v části A) ukazují pozitivní asociaci s Thesium. Podtržení zvýrazňuje druhy, které se vyskytují v obou grafech.Fig. 3. Comparison of species A) positively associated with T. linophyllon in CCA analysis of above-ground vegetation composition and B) species on roots of which T. linophyllon haustoria were found most often. High positive values on the y axis in A) mean a positive association with Thesium. Names of species occurring in both graphs are underlined.
Příroda, Praha, 31: 201–225, 2011
211

Nalezli jsme průkazné rozdíly mezi indikačními hodnotami vypočtenými z druhů rostoucích s jednotlivými druhy Thesium. Dále jsme nalezli průkazné rozdíly i v počtu druhů mezi plochami s jednotlivými studovanými druhy z rodu Thesium, což znamená, že se jednotlivé druhy Thesium vyskytují v odlišných podmínkách prostředí. Plochy s T. ebracteatum měly nejvyšší indikační hodnoty pro vlhkost (F2,29 = 231,0; P < 0,001). Plochy s T. linophyllon měly nejvyšší indikační hodnoty pro světlo (F2,29 = 476,1; P < 0,001), teplotu (F2,29 = 137,6; P < 0,001), kontinentalitu (F2,29 = 69,0; P < 0,001) a půdní reakci (F2,29 = 509,1; P < 0,001) a nejnižší hodnoty pro živiny (F2,29 = 141,7; P < 0,001). Plochy s T. bavarum měly nejnižší indikační hodnoty pro světlo. V plochách s T. linophyllon jsme nalezli více druhů než v plochách s T. ebracteatum a T. bavarum (F2,29 = 111,4; P < 0,001) (příloha 1). Ve všech testech jsme nalezli také průkazný vliv lokality vřazené do druhu Thesium, což ukazuje na rozdíly mezi lokalitami v rámci jednoho druhu Thesium. Když jsme testovali rozdíly v Ellenbergových indikačních hodnotách a počtu druhů mezi lokalitami tří druhů Thesium pouze s použitím ploch, ve kterých se Thesium nevyskytovalo, byly výsledky velice podobné. Výsledky byly také velice podobné mezi třemi velikostmi ploch. Rozdíly mezi třemi druhy Thesium byly většinou v souladu s rozdíly v Ellenbergových indikačních hodnotách pro tři studované druhy rodu Thesium (ellenberG et al. 2001).
Potenciální asociace s hostitelskými rostlinami
U druhů T. ebracteatum a T. bavarum jsme nenalezli žádný průkazný vztah mezi druhovým složením a přítomností Thesium v žádné velikosti ploch. Ani interakce mezi přítomností Thesium a lokalitou nebyla průkazná, což ukazuje na to, že ani v rámci jednotlivých lokalit neexistuje asociace Thesium s některými druhy.
Vztah mezi přítomností Thesium a druhovým složením v plochách byl však průkazný ve všech třech studovaných velikostech ploch pro T. linophyllon. Přítomnost Thesium vysvětlila 12,2 % z celkové variability v druhovém složení, která může být vysvětlena první ordinační osou v analýze CCA, pro plochy o velikosti 20 × 20 cm, P = 0,006; 18,9 % pro plochy o velikosti 50 × 50 cm, P = 0,002; a 13,6 % pro plochy o velikosti 100 × 100 cm, P = 0,002. Druhy, které byly nejvíce asociované s T. linophyllon jsou uvedeny na obr. 3A. Mezi druhy nejvíce negativně asociovány patřily zejména Trifolium medium, Cornus sanguinea, Arrhenatherum elatius, Prunus spinosa, Salvia nemorosa a Elytrigia intermedia. Asociované druhy byly podobné ve všech velikostech ploch. Interakce mezi přítomností Thesium a lokalitou byla ve všech velikostech ploch neprůkazná (P > 0,15 ve všech případech), což ukazuje na podobnou skupinu asociovaných druhů na všech studovaných lokalitách. Když jsme testovali vztah mezi přítomností T. linophyllon a druhovým složením zvlášť pro každou ze tří lokalit, kde byly identifikovány také druhy s fyzickým napojením, tak byly výsledky průkazné pouze pro lokalitu L3.
Když jsme testovali vztah mezi přítomností Thesium a přítomnými čeleděmi (namísto dru hového složení), nalezli jsme průkazný vztah nejen pro T. linophyllon (6,9 % pro plochy 20 × 20 cm, P = 0,044; 8,8 % pro plochy 50 × 50 cm, P = 0,004; 9,5 % pro plochy 100 × 100 cm, P = 0,002) ale i pro T. ebracteatum (10,0% pro plochy 20 × 20 cm, P = 0,008; 8,2 % pro plochy 50 × 50 cm, P = 0,060; 8,1 % pro plochy 100 × 100 cm, P = 0,032). Čeledi pozitivně nebo negativně asociované s Thesium byly podobné ve všech studovaných velikostech ploch. Avšak průkazná byla i interakce mezi přítomností Thesium a lokalitou, což naznačuje rozdíly v asociovaných čeledích mezi jednotlivými lokalitami. Čeledi pozitivně a negativně asociované s T. ebracteatum a T. linophyllon jsou ukázány na obr. 4.
212
Dostálek & Münzbergová: Stanovištní nároky a výběr hostitelů u rodu Thesium
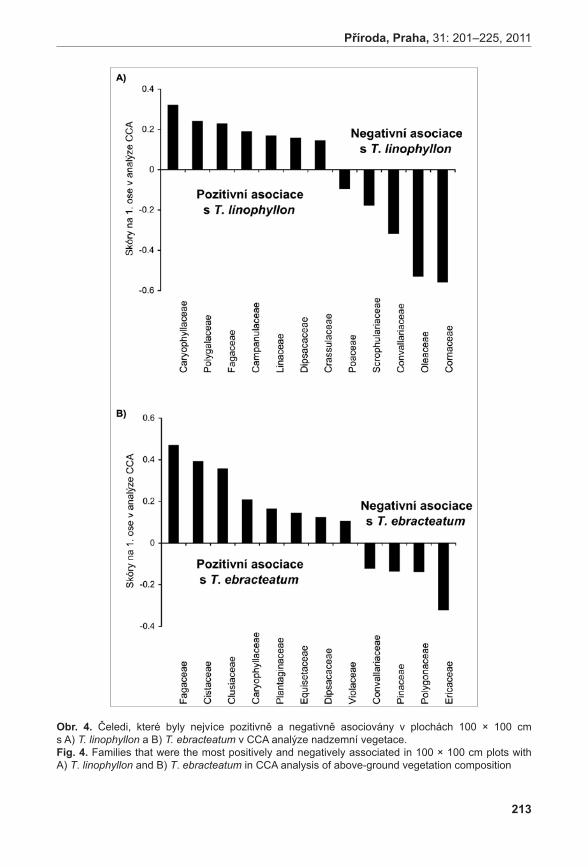
obr. 4. Čeledi, které byly nejvíce pozitivně a negativně asociovány v plochách 100 × 100 cm s A) T. linophyllon a B) T. ebracteatum v CCA analýze nadzemní vegetace.Fig. 4. Families that were the most positively and negatively associated in 100 × 100 cm plots with A) T. linophyllon and B) T. ebracteatum in CCA analysis of above-ground vegetation composition
Příroda, Praha, 31: 201–225, 2011
213
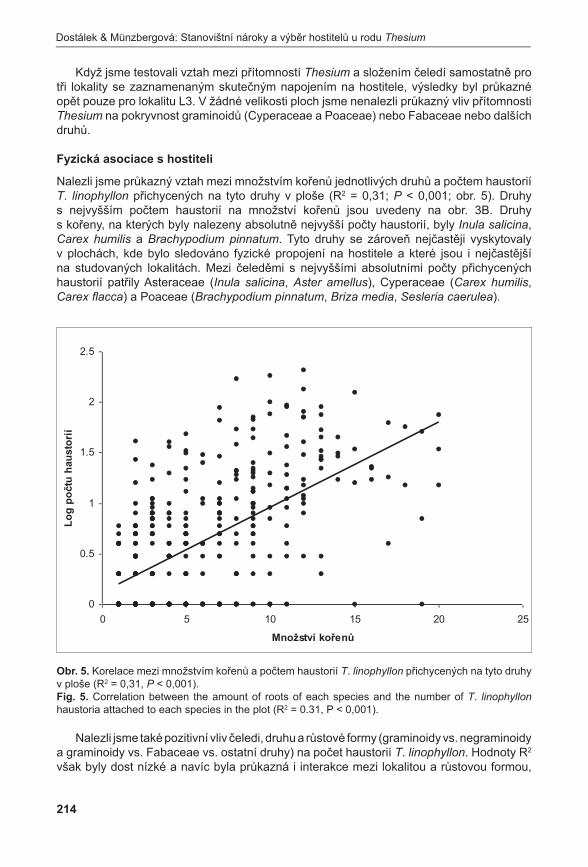
Když jsme testovali vztah mezi přítomností Thesium a složením čeledí samostatně pro tři lokality se zaznamenaným skutečným napojením na hostitele, výsledky byl průkazné opět pouze pro lokalitu L3. V žádné velikosti ploch jsme nenalezli průkazný vliv přítomnosti Thesium na pokryvnost graminoidů (Cyperaceae a Poaceae) nebo Fabaceae nebo dalších druhů.
Fyzická asociace s hostiteli
Nalezli jsme průkazný vztah mezi množstvím kořenů jednotlivých druhů a počtem haustorií T. linophyllon přichycených na tyto druhy v ploše (R2 = 0,31; P < 0,001; obr. 5). Druhy s nejvyšším počtem haustorií na množství kořenů jsou uvedeny na obr. 3B. Druhy s kořeny, na kterých byly nalezeny absolutně nejvyšší počty haustorií, byly Inula salicina, Carex humilis a Brachypodium pinnatum. Tyto druhy se zároveň nejčastěji vyskytovaly v plochách, kde bylo sledováno fyzické propojení na hostitele a které jsou i nejčastější na studovaných lokalitách. Mezi čeleděmi s nejvyššími absolutními počty přichycených haustorií patřily Asteraceae (Inula salicina, Aster amellus), Cyperaceae (Carex humilis, Carex fl acca) a Poaceae (Brachypodium pinnatum, Briza media, Sesleria caerulea).
obr. 5. Korelace mezi množstvím kořenů a počtem haustorií T. linophyllon přichycených na tyto druhy v ploše (R2 = 0,31, P < 0,001).Fig. 5. Correlation between the amount of roots of each species and the number of T. linophyllon haustoria attached to each species in the plot (R2 = 0.31, P < 0,001).
Nalezli jsme také pozitivní vliv čeledi, druhu a růstové formy (graminoidy vs. negraminoidy a graminoidy vs. Fabaceae vs. ostatní druhy) na počet haustorií T. linophyllon. Hodnoty R2 však byly dost nízké a navíc byla průkazná i interakce mezi lokalitou a růstovou formou,
214
Dostálek & Münzbergová: Stanovištní nároky a výběr hostitelů u rodu Thesium
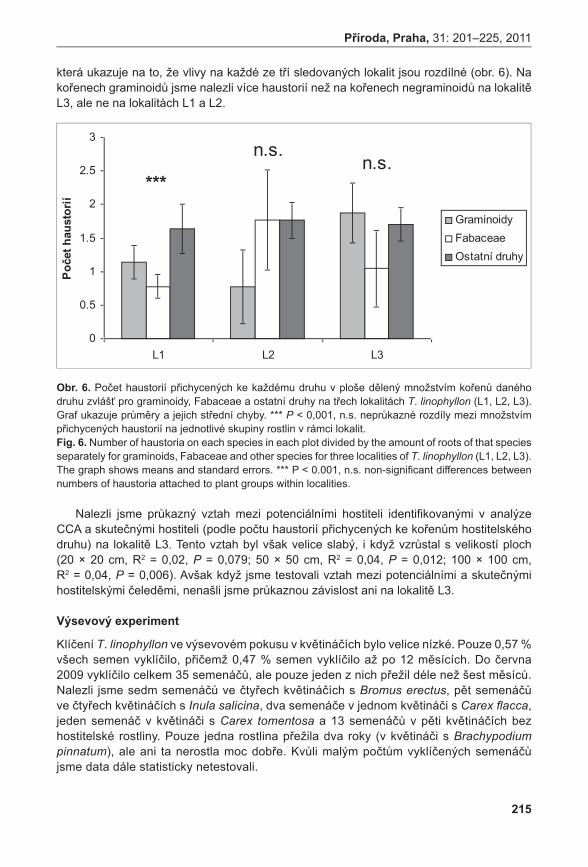
která ukazuje na to, že vlivy na každé ze tří sledovaných lokalit jsou rozdílné (obr. 6). Na kořenech graminoidů jsme nalezli více haustorií než na kořenech negraminoidů na lokalitě L3, ale ne na lokalitách L1 a L2.
obr. 6. Počet haustorií přichycených ke každému druhu v ploše dělený množstvím kořenů daného druhu zvlášť pro graminoidy, Fabaceae a ostatní druhy na třech lokalitách T. linophyllon (L1, L2, L3). Graf ukazuje průměry a jejich střední chyby. *** P < 0,001, n.s. neprůkazné rozdíly mezi množstvím přichycených haustorií na jednotlivé skupiny rostlin v rámci lokalit.Fig. 6. Number of haustoria on each species in each plot divided by the amount of roots of that species separately for graminoids, Fabaceae and other species for three localities of T. linophyllon (L1, L2, L3). The graph shows means and standard errors. *** P < 0.001, n.s. non-signifi cant differences between numbers of haustoria attached to plant groups within localities.
Nalezli jsme průkazný vztah mezi potenciálními hostiteli identifi kovanými v analýze CCA a skutečnými hostiteli (podle počtu haustorií přichycených ke kořenům hostitelského druhu) na lokalitě L3. Tento vztah byl však velice slabý, i když vzrůstal s velikostí ploch (20 × 20 cm, R2 = 0,02, P = 0,079; 50 × 50 cm, R2 = 0,04, P = 0,012; 100 × 100 cm, R2 = 0,04, P = 0,006). Avšak když jsme testovali vztah mezi potenciálními a skutečnými hostitelskými čeleděmi, nenašli jsme průkaznou závislost ani na lokalitě L3.
výsevový experiment
Klíčení T. linophyllon ve výsevovém pokusu v květináčích bylo velice nízké. Pouze 0,57 % všech semen vyklíčilo, přičemž 0,47 % semen vyklíčilo až po 12 měsících. Do června 2009 vyklíčilo celkem 35 semenáčů, ale pouze jeden z nich přežil déle než šest měsíců. Nalezli jsme sedm semenáčů ve čtyřech květináčích s Bromus erectus, pět semenáčů ve čtyřech květináčích s Inula salicina, dva semenáče v jednom květináči s Carex fl acca, jeden semenáč v květináči s Carex tomentosa a 13 semenáčů v pěti květináčích bez hostitelské rostliny. Pouze jedna rostlina přežila dva roky (v květináči s Brachypodium pinnatum), ale ani ta nerostla moc dobře. Kvůli malým počtům vyklíčených semenáčů jsme data dále statisticky netestovali.
Příroda, Praha, 31: 201–225, 2011
215
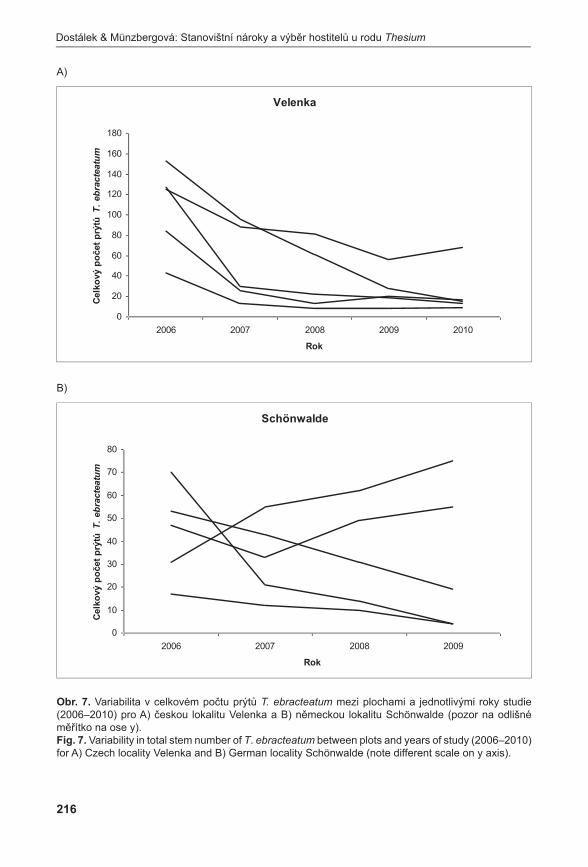
A)
B)
obr. 7. Variabilita v celkovém počtu prýtů T. ebracteatum mezi plochami a jednotlivými roky studie (2006–2010) pro A) českou lokalitu Velenka a B) německou lokalitu Schönwalde (pozor na odlišné měřítko na ose y).Fig. 7. Variability in total stem number of T. ebracteatum between plots and years of study (2006–2010) for A) Czech locality Velenka and B) German locality Schönwalde (note different scale on y axis).
216
Dostálek & Münzbergová: Stanovištní nároky a výběr hostitelů u rodu Thesium
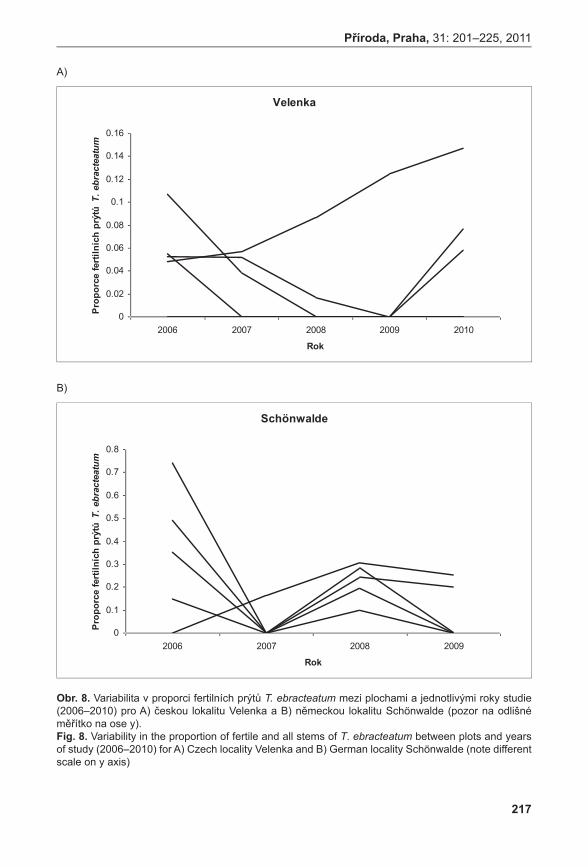
A)
B)
obr. 8. Variabilita v proporci fertilních prýtů T. ebracteatum mezi plochami a jednotlivými roky studie (2006–2010) pro A) českou lokalitu Velenka a B) německou lokalitu Schönwalde (pozor na odlišné měřítko na ose y).Fig. 8. Variability in the proportion of fertile and all stems of T. ebracteatum between plots and years of study (2006–2010) for A) Czech locality Velenka and B) German locality Schönwalde (note different scale on y axis)
Příroda, Praha, 31: 201–225, 2011
217
Meziroční variabilita v počtu prýtů
Počty prýtů T. ebracteatum kolísaly v jednotlivých plochách a letech od 8 do 153 na lokalitě Velenka a od 4 do 75 na lokalitě Schönwalde (obr. 7). V celkovém počtu prýtů jsme nalezli průkazné rozdíly mezi lokalitami, plochami i roky, přičemž největším zdrojem variability byly rozdíly mezi plochami v rámci lokalit. Proporce fertilních prýtů byla průkazně vyšší na německé lokalitě Schönwalde než na české Velence. Tato proporce se však také průkazně lišila mezi roky (obr. 8).
Velkou variabilitu jsme nalezli i ve směru vývoje počtu prýtů na obou sledovaných lokalitách (tab. 3). Na lokalitě Velenka se celkový počet prýtů ve všech plochách zmenšoval, avšak na lokalitě Schönwalde docházelo v některých plochách ke zvětšování počtu všech prýtů a v jiných ke zmenšování (obr. 7). Proporce fertilních prýtů kolísala u obou sledovaných lokalit (obr. 8).
tab. 3. Výsledky testů variability v počtu prýtů T. ebracteatum mezi plochami a jednotlivými roky studie (Velenka 2006–2010, Schönwalde 2006–2009). * P < 0,05, ** P < 0,01, *** P < 0,001, n.s. neprůkazné.tab. 3. Results of tests of variability in stem number of T. ebracteatum between plots and years of study (Velenka 2006–2010, Schönwalde 2006–2009). * P < 0.05, ** P < 0.01, *** P < 0.001, n.s. non-significant.
Velenka Schönwalde df R2 P R2 P
Počet všech prýtůplocha 4 0,39 *** 0,52 ***rok 1 0,45 *** 0,03 *plocha*rok 4 0,09 * 0,41 ***
Proporce fertilních prýtůplocha 4 0,41 ** - n.s.rok 1 - n.s. - n.s.plocha*rok 4 0,32 * - n.s.
diskuse
Cílem této studie bylo identifikovat stanovištní nároky a hostitele tří druhů kořenových poloparazitů z rodu Thesium s použitím přímé a nepřímé metody a manipulativního experimentu. Oproti našim předpokladům jsme zjistili, že studovaní zástupci z rodu Thesium si své hostitele specificky nevybírají a jsou schopni vyklíčit i bez přítomnosti hostitelské rostliny. Výsledky také ukazují, že se studované druhy Thesium vyskytují v rámci svých stanovišť ve specifických mikrostanovištích. Tento výskyt může být teoreticky také způsoben vlivem poloparazitů na okolní vegetaci. Dále jsme se snažili zjistit, jaká je variabilita v populační dynamice T. ebracteatum. Zjistili jsme, že variabilita na všech sledovaných úrovních byla poměrně značná, což ukazuje na velkou dynamiku v populacích tohoto druhu. Změny v populacích mohou být způsobeny výkyvy počasí nebo i lokálními disturbancemi.
Náš výsledek, že místa, kde roste Thesium, jsou druhově bohatší, je v souladu i s dalšími studiemi zabývajícími se kořenovými poloparazity. Je známo, že kořenoví poloparazité z rodu Rhinanthus (Orobanchaceae) podporují v rámci společenstva biodiverzitu a uchycování nových druhů (joshi et al. 2000, Pywell et al. 2004). Gibson & watkinson (1991) také objevili, že poloparaziti mají vliv na druhovou bohatost, což může být způsobeno selektivním potlačováním části společenstva a změnou kompetičních
218
Dostálek & Münzbergová: Stanovištní nároky a výběr hostitelů u rodu Thesium

vzta hů mezi jednotlivými druhy. Je proto možné, že i Thesium by šlo využít ke zvýšení diverzity ve společenstvech. Naše studie však také ukázala, že je pěstování Thesium dost obtížné. Existuje však i jiné vysvětlení pro větší bohatost v porostech s Thesium. A to je, že tato druhová bohatost je způsobena specifickými podmínkami prostředí, které umožňují uchycení více druhů včetně Thesium.
U druhů rostoucích v plochách s T. linophyllon jsme nalezli nižší indikační hodnoty pro živiny a vyšší indikační hodnoty pro světlo. Matthies (1995) dospěl k podobnému závěru, když zjistil, že rovnováha mezi vlivem kořenových (polo)parazitů na hostitele (např. kompeticí o živiny) a vlivem kompetice (polo)parazita s hostitelem o světlo vytěsňuje (polo)parazity na živinami relativně chudá místa. Tento jev jsme však nepozorovali pro další dva studované druhy T. bavarum a T. ebracteatum. Důvodem může být to, že T. ebracteatum kvete a plodí většinou v květnu, kdy ještě není okolní vegetace příliš vysoká a tudíž T. ebracteatum netrpí tolik kompeticí s okolními druhy jako T. linophyllon, které kvete a plodí až o měsíc nebo dva později. Na rozdíl od T. linophyllon a T. ebracteatum, T. bavarum tvoří trsy a je mnohem větší než další dva druhy. Z tohoto důvodu je pravděpodobně mnohem lepší kompetitor a nemusí tak ustupovat na rozvolněnější a živinami chudší místa.
Nepodařilo se nám najít druhy, které by se častěji vyskytovaly s T. ebracteatum nebo T. bavarum a mohly by tak sloužit jako jejich potenciální hostitelé. Asociace s ostatními druhy byly průkazné pouze u druhu T. linophyllon. Když jsme však analyzovali zvlášť tři lokality, kde jsme sledovali i fyzické napojení T. linophyllon na hostitele, tak asociace zůstala průkazná pouze na jedné lokalitě. To může být způsobeno velkou variabilitou v druhovém složení na plochách anebo relativně malým počtem studovaných ploch na lokalitu. Velkou variabilitu mezi plochami ve své studii zaznamenali i Gibson & watkinson (1989), kterým se také nepodařilo najít jakoukoli zobecnitelnou asociaci mezi druhy na lokalitě a poloparazickým Rhinanthus minor. Absenci průkazného vztahu u T. ebracteatum bychom mohli vysvětlit také tím, že studované lokality jsou kvůli vzácnosti druhu ve střední Evropě roztroušené na relativně velkém území a druhové složení na jednotlivých lokalitách se tak značně liší. Závěr, že jsme nenalezli žádný vztah mezi druhovým složením a přítomností T. ebracteatum kvůli velké variabilitě v druhovém složení mezi lokalitami, je také podpořen tím, že jsme nalezli průkazný vztah poté, co jsme místo druhů použili čeledi. Další možné vysvětlení je, že žádná asociace T. ebracteatum s hostiteli opravdu neexistuje.
Druhy, na které se T. linophyllon nejčastěji přichytávalo svými haustorii, byly Inula salicina, Carex humilis a Brachypodium pinnatum patřící do čeledí Asteraceae, Cyperaceae a Poaceae (i přes negativní asociaci s Poaceae v nadzemní vegetaci). Nezjistili jsme však, že by si T. linophyllon nějak zvlášť vybíralo své hostitele, protože tyto druhy byly také nejhojnější na studovaných lokalitách. Za zmínku určitě také stojí průkazný vztah mezi množstvím kořenů a počtem haustorií T. linophyllon pro jednotlivé druhy v plochách. Tento vztah ukazuje, že T. linophyllon tvoří haustoria téměř na všech kořenech v půdě a tedy čím více má druh kořenů, tím více je haustorii napaden. K podobnému závěru došli ve své studii i suetsuGu et al. (2008), kteří také nenalezli zřetelnou specificitu ve výběru hostitelských druhů u T. chinense.
Shoda mezi potenciálními a reálnými hostiteli byla průkazná pouze v jedné populaci T. linophyllon, ale tento vztah byl velice slabý. Předpokládáme tedy, že některé druhy jsou lepšími nebo horšími hostiteli pro T. linophyllon, ale že T. linophyllon nepotřebuje ke svému růstu konkrétní druhy hostitelských rostlin. To dokazuje i fakt, že v naší studii jsme nalezli haustoria T. linophyllon dokonce na 94 % všech druhů rostoucích v plochách s T. linophyllon. Podobně i suetsuGu et al. (2008) zjistili, že T. chinense bylo haustorii napojeno na 58 % všech druhů rostoucích na lokalitě. Bylo by však ještě potřeba v dalších pokusech ověřit, zda jsou haustoria přichycená na různé hostitelské rostliny stejně funkční
Příroda, Praha, 31: 201–225, 2011
219

a efektivní v přenosu látek od hostitele k parazitovi. Existuje teoretická možnost, že přestože Thesium vytváří haustoria prakticky na kořenech všech okolních rostlin, jenom některé z nich ve skutečnosti využívá jako své hostitele.
Původní motivací naší studie bylo použít informaci o vztahu mezi potenciálními a skutečnými hostiteli k identifikaci hostitelských druhů vzácných druhů, aniž bychom v jejich populacích museli vykopávat rostliny a ohrozit tak vitalitu často malých populací. Ani u jednoho ze dvou studovaných vzácných druhů (T. ebracteatum a T. bavarum) jsme však nenalezli průkazný vztah mezi přítomností Thesium a ostatních druhů rostlin. Na druhou stranu se naše data ukázala být užitečná pro identifikaci stanovištních nároků Thesium, které se zdají být mnohem významnější pro uchycení a přežívání druhu než specifický druh hostitele.
Při našich pokusech o pěstování T. linophyllon jsme byli poměrně neúspěšní, přestože v mnoha studiích je popisováno úspěšné pěstování poloparazitů s jejich hostiteli a dokonce i bez nich (např. Castilleja levisecta – lawrence & kaye 2008, Rhinanthus minor – Gibson & watkinson 1991, Rhinanthus serotinus – Matthies 1995, Odontites rubra – Matthies 1995, Melampyrum arvense – Matthies 1996). Všechny výše zmíněné studie se však zabývaly druhy z čeledi Orobanchaceae, které mají semena často malá bez většího množství zásobních látek. Navíc pro vytvoření haustorií u kořenových parazitů z čeledi Orobanchaceae jsou také zapotřebí chemické stimulanty (MusselMan 1980, yoDer 2001, bouwMeester et al. 2003). Rostliny z řádu Santalales mají naopak často semena relativně velká s velkým množstvím zásobních látek. Z tohoto důvodu by také mohly mít menší nároky na přítomnost chemických stimulantů pro samotné klíčení semen. To ukázali i fer et al. (1993, 1994), kteří zjistili, že T. humile klíčí i bez přítomnosti chemických stimulantů. Při našich pokusech jsme však byli schopni vypěstovat pouze několik semenáčů, přičemž pouze jedna rostlina přežila déle než šest měsíců. Jedním z důvodů malého počtu semenáčů může být malé procento vyvinutých semen T. linophyllon (8–20 %, T. Dostálek, osobní pozorování). Pozorovali jsme také velmi málo semenáčů v přirozených populacích, což ukazuje na převládající vegetativní rozmnožování v přirozených populacích. Z tohoto důvodu jsme také nemohli provést původně naplánovaný experiment, kdy jsme chtěli pěstovat T. linophyllon s rostlinami identifikovanými jako potenciální hostitelé.
V poslední části studie jsme nalezli značnou variabilitu v populační dynamice T. ebracteatum. Zaznamenali jsme velké rozdíly v počtu prýtů jak mezi jednotlivými plochami v populacích, tak i mezi jednotlivými lety studie. Velká variabilita v počtu prýtů mezi plochami ukazuje opět na velký vliv mikrostanovištních podmínek na populační dynamiku T. ebracteatum. Na české lokalitě Velenka může být variabilita v počtu prýtů také způsobena lokálními disturbancemi způsobenými při sečení lokality. Přestože je tento druh managementu celkově pro druh prospěšný, protože eliminuje kompetičně silnější druhy na lokalitě, v menších velikostech ploch se může dočasně projevit negativně. To bude také pravděpodobně důvod, proč je na lokalitě Velenka soustavný úbytek počtu prýtů ve sledovaných plochách, přestože celkově se populace zdá být stabilní. Proporce kvetoucích prýtů je pravděpodobně také velice závislá na výkyvech počasí. Zaznamenali jsme vliv extrémně suchého dubna 2007 (5 mm/měsíc) na pokles proporce fertilních prýtů, který byl patrný zejména na německé lokalitě Schönwalde.
Naše studie je jedna z prvních prací, která se zabývá stanovištními nároky a asociací s hostitelskými druhy poloparazitických druhů z rodu Thesium a která se zabývá najednou více druhy ve více populacích (viz také Dostálek & MünzberGová 2010). Jsou však zapotřebí ještě další studie, které by se zabývaly hostitelskými nároky Thesium, zvláště pokusy s klíčením Thesium v různých podmínkách a pokusy zkoumající vliv přítomnosti Thesium na okolní vegetaci v přirozených populacích.
220
Dostálek & Münzbergová: Stanovištní nároky a výběr hostitelů u rodu Thesium

shrnutí a závěry pro ochranu studovaných druhů
V naší studii jsme zjistili, že místa, kde roste T. ebracteatum a T. linophyllon, byla ve srovnání s okolím druhově bohatší. Také se ukázalo, že T. linophyllon roste na místech, která jsou světlejší, živinami chudší a sušší než zbytek lokality. Našli jsme také velké rozdíly ve stanovištních nárocích jednotlivých studovaných druhů Thesium, což ukazuje na jejich specifické mikrostanovištní nároky a/nebo na jejich vliv na společenstvo hostitelských rostlin. Nalezli jsme také pozitivní asociaci některých druhů s T. linophyllon, ne však se zbývajícími druhy Thesium. Počet haustorií fyzicky napojených na kořeny hostitelských druhů byl průkazně ovlivněn množstvím kořenů druhů v ploše, ale také tím, ke kterému druhu, čeledi nebo rostlinné skupině kořeny hostitelů patřily. Thesium linophyllon bylo celkem napojeno dokonce na 94 % všech druhů, se kterými rostlo. Z výsledků naší studie tedy vyplývá, že Thesium si své hostitele specificky nevybírá, ale vyskytuje se na mikrostanovištích se specifickými podmínkami. Tento závěr potvrzuje i fakt, že bylo schopné vyklíčit i v květináčích bez přidaného hostitele. Je proto velice pravděpodobné, že není možné podpořit současné populace těchto kořenových poloparazitů přidáním nebo podporou několika málo specifických hostitelů na lokalitách. To ukazují i výsledky sledování populační dynamiky T. ebracteatum, kdy jsme na dvou sledovaných lokalitách nalezli značnou variabilitu mezi počty prýtů v jednotlivých plochách a letech bez ohledu na změnu v přítomnosti potenciálních hostitelských rostlin. V naší studii jsme nezkoumali různé parametry prostředí, které by mohly být za tyto změny zodpovědné, soudíme však, že velký vliv mohou mít změny klimatu (viz negativní efekt extrémně suchého dubna 2007 na proporci fertilních prýtů). Dalším důležitým faktorem může být management sečení, který je prováděn na lokalitách T. ebracteatum v ČR. Přestože management lokalit s T. ebracteatum může mít krátkodobě negativní vliv (poničení malé části populace), dlou ho době potlačuje kompetičně silné druhy, které by jinak pravděpodobně T. ebracteatum vytlačily. Populace T. ebracteatum v Německu nebo Polsku většinou sečené nejsou, jsou ale často na místech, která jsou disturbovaná (okraje cest nebo železničních tratí na písku), což zvýhodňuje tento kompetičně slabší druh. Na lokalitách s T. linophyllon a T. bavarum není potřeba provádět management sečení, protože jsou často na místech, kde se silní luční kompetitoři nemohou prosadit (živinami chudá stanoviště, suťové a skalnaté svahy). Lokality s těmito druhy však mohou zarůstat dřevinami a křovinami, které je potřeba jednou za čas vyřezávat.
summary
We found in our study that sites with T. linophyllon and T. ebracteatum had higher species diversity and sites with T. linophyllon had more light, less nutrients and less moisture than sites without Thesium. There were also strong differences in habitat requirements among the three Thesium spp. This suggests that Thesium spp. have specific microhabitat requirements and/or they affect their host community. We also found positive associations with surrounding species for T. linophyllon, but not for the other Thesium spp. The number of haustoria physically attached to roots of other species was significantly affected by the number of roots of the species, but also by its identity (species, family and plant group); in fact, T. linophyllon was attached to 94% of all species occurring in the plots. All this suggests that Thesium spp. do not specifically select hosts, but rather occur in microhabitats with specific conditions. This conclusion is also supported by the fact that germination of
Příroda, Praha, 31: 201–225, 2011
221

Thesium in the sowing experiment was higher in pots without any host. Our study suggests it is not very useful to support current population of studied Thesium spp. by favoring a few specific host plants in the localities. This is also supported by population dynamics study when we found high variability in number of Thesium stems in individual plots and years no matter if the proportion of potential host plants was high or low. In our study we did not focus on various environmental factors which could be responsible for variability in number of Thesium stems in the plots. However, we suggest climate fluctuations can influence population dynamics a lot (see negative effect of extremely dry April 2007 on proportion of fertile stems of T. ebracteatum). Another important factor could be management of mowing performed on T. ebracteatum localities in the Czech Republic. Although mowing can negatively influence the population in short term view (destroying small part of population by mowing machine), in long term view it reduces competively strong species which would probably force T. ebracteatum out. Populations of T. ebracteatum in Germany or in Poland are usually in disturbed places (surroundings of roads or railways on sand), which favour this competively weak species. In the localities with T. linophyllon and T. bavarum mowing management is not necessary because these places are not usually much suitable for meadow competitors (nutrient poor sites, scree and rock slopes). However, localities with T. linophyllon and T. bavarum can be overgrown with trees and shrubs which need to be reduced.
Poděkování
Autoři by rádi poděkovali těm, kteří nám poskytli užitečné informace o populacích druhu, jsou to zej-ména A. Wrobłewska, L. Rutkowski a T. Załuski z Polska a H. Illig a A. Herrmann z Německa. Řadu užitečných komentářů k práci a k rukopisu článku nám poskytli také lidé ze semináře POPEKOL. Tato práce byla podpořena MŠMT VaV 2B06178, GAUK 148/2006, FRVŠ 1897/2006, GAČR 206/08/H049, MŠMT 0021620828 a AV0Z60050516.
literatura
aDler l. s. (2003): Host species affects herbivory, pollination, and reproduction in experiments with parasitic Castilleja. − Ecology 84: 2083–2091.
abu-irMaileh b. e. (1980): Thesium humile on onion. − Haustorium 6: 2.bouwMeester h. j., Matusova r., zhonGkui s. & beale M. h. (2003): Secondary metabolite
signalling in host-parasitic plant interactions. − Curr. Opin. Plant Biol. 6: 358–364.DasGuPta M. K. (1988): Principles of plant pathology. − Allied Publisher, New Delhi.Dostálek t. & MünzberGová z. (2010): Habitat requirements and host selectivity of hemiparasite
Thesium species. − Bot. J. Linn. Soc. 164: 394–408.Čeřovský J. (1999): Thesium ebracteatum L. − In: Čeřovský j., feráková v., holub j., MaGlocký Š.
& Procházka f. [eds], Červená kniha ohrožených a vzácných druhů rostlin a živočichů ČR a SR, vol. 5, Vyšší rostliny, pp. 373, Príroda, Bratislava.
ellenberG h., weber h. e., Düll r., wirth v., werner w. & Paulissen D. (2001): Indicator values of vlants in Central Europe. − Goltze, Göttingen.
euroPean coMMission (1992): Directive 92/43/EEC of the European Parliament and of the Council of 21st May 1992: conservation of natural habitats and of wild fauna and flora. − Official Journal of the European Communities 206: 7–50.
fer a., siMier P., arnauD M., rey l. & renauDin s. (1993): Carbon acquisition and metabolism in a root hemiparasitic angiosperm, Thesium humile (Santalaceae) growing on wheat (Triticum vulgare). − Funct. Plant Biol. 20: 15–24.
222
Dostálek & Münzbergová: Stanovištní nároky a výběr hostitelů u rodu Thesium

fer a., russo n., siMier P., arnauD M. c. & thalouarn P. (1994): Physiological changes in a root hemiparasitic angiosperm, Thesium humile (Santalaceae), before and after attachment to the host plant (Triticum vulgare). − J. Plant Physiol. 143: 704–710.
Gibson c. c. & watkinson a. r. (1989): The host range and selectivity of a parasitic plant: Rhinanthus minor L. − Oecologia 78: 401–406.
Gibson c. c. & watkinson a. r. (1991): Host Selectivity and the Mediation of Competition by the Root Hemiparasite Rhinanthus minor. − Oecologia 86: 81–87.
Gibson c. c. & watkinson a. r. (1992): The role of the hemiparasitic annual Rhinanthus minor in determining grassland community structure. − Oecologia 89: 62–68.
henDrych r. (1969a): The outline of the taxonomy and chorology of Thesium linophyllon. − Acta Univ. Carol. - Biol. 1969: 119–166.
henDrych r. (1969b): Systematic outline of Thesium ebracteatum. − Preslia 41: 229–240.henDrych r. (1980): Thesium L. − In: tutin t. G., heywooD v. h., burGes n. a., valentin D.,
walter s. M. & webb D. a. [eds], Flora Europaea, vol. 5, pp. 70−72, Cambridge University Press.
holub j. & Procházka f. (2000): Red list of vascular plants of the Czech Republic. − Preslia 72: 187–230.
joshi j., Matthies D. & schMiD b. (2000): Root hemiparasites and plant diversity in experimental grassland communities. − J. Ecol. 88: 634–644.
kubát k., hrouDa l., chrtek j. jun., kaPlan z., kirschner j. & ŠtěPánek j. [eds] (2002): Klíč ke květeně České republiky. – Academia, Praha.
lawrence b. a. & kaye t. n. (2008): Direct and indirect effects of host plants: implications for reintroduction of an endangered hemiparasitic plant (Castilleja levisecta). − Madroño 55: 151–158.
lePŠ j. & ŠMilauer P. (2003): Multivariate analysis of ecological data using CANOCO. − Cambridge University Press.
libby G. w. & blooM c. t. (1998): Nestronia umbellula Rafinesque (Santalaceae) from the Highland Rim of Kentucky. − Castanea 63: 161–164.
Marvier M. a. & sMith D. l. (1997): Conservation implications of host use for rare parasitic plants. − Conserv. Biol. 11: 839–848.
Matthies D. (1995): Parasitic and competitive interactions between the hemiparasites Rhinanthus serotinus and Odontites rubra and their host Medicago sativa. − J. Ecol. 83: 245–251.
Matthies D. (1996): Interactions between the root hemiparasite Melampyrum arvense and mixtures of host plants: heterotrophic benefit and parasite-mediated competition. − Oikos 75: 118–124.
Mueller r. c. & GehrinG c. a. (2006): Interactions between an above-ground plant parasite and below-ground ectomycorrhizal fungal communities on pinyon pine. − Ecology 94: 276–284.
MusselMan L. J. (1980): The biology of Striga, Orobanche, and other root-parasitic weeds. − Ann. Rev. Phytopathol. 18: 463–489.
MusselMan l. j. & haynes s. c. (1996): Santalaceae with weed potential in the United States. − Congresos y Jornadas-Junta de Andalucia (Espana).
PhoeniX G. k. & Press M. c. (2005a): Linking physiological traits to impacts on community structure and function: the role of root hemiparasitic Orobanchaceae (ex-Scrophulariaceae). − J. Ecol. 93: 67–78.
PhoeniX G. & Press M. c. (2005b): Effects of climate change on parasitic plants: the root hemiparasitic Orobanchaceae. − Folia Geobot. 40: 205–216.
PilGer r. (1935): Santalaceae. − In: enGler a. & harMs h. [eds], Die Naturlichen Pflanzen Familien, vol. 16, Duncker & Humblot, Leipzig, pp. 52−91
Prach j. & zajíČková l. (2009): Thesium ebracteatum Hayne. − In: haDinec j. & lustyk P. [eds], Additamenta ad floram Reipublicae Bohemicae. VIII., Zpr. Čes. Bot. Společ. 44: 185−319.
Press M. c. & Graves J. D. (1995): Parasitic plants. − Chapman and Hall, London.
Příroda, Praha, 31: 201–225, 2011
223

Pywell r. f., bullock j. M., walker k. j., coulson s. j., GreGory s. j. & stevenson M. j. (2004): Facilitating grassland diversification using the hemiparasitic plant Rhinanthus minor. − J. Appl. Ecol. 41: 880–887.
QaseM j. R. (2006): Host range of the parasitic weed Osyris alba L. in Jordan. − Weed Biol. Manag. 6: 74–78.
ren y-Q., Guan k-y., li a-r., hu X-j. & zhanG l. (2010): Host dependence and preference of the root hemiparasite, Pedicularis cephalantha Franch. (Orobanchaceae). − Folia Geobot. 45: 443−455.
renauDin s., cheGuillauMe n. & Gallant D. J. (1981): Distribution and role of mineral compounds in the haustorium of a parasite of Galium arenarium, Thesium humifusum, before flowering. − Can. J. Bot. 59: 1998–2002.
scarlett n., braMwell M. & earl G. (2003): Austral toad flax. − Flora and Fauna Guarantee Action Statement 56: 1–5.
siMPson e. h. (1949): Measurement of diversity. − Nature 163: 688. slavík b. [ed.] (1997): Květena České republiky. Vol. 5. − Academia, Praha.S-PLUS (2000): Professional edition for Windows, release 2. − MathSoft, Inc., Massachusetts.suetsuGu k., kawakita a. & kato M. (2008): Host range and selectivity of the hemiparasitic plant
Thesium chinense (Santalaceae). − Ann. Bot. 102: 49–55. ter braak c. j. & ŠMilauer P. (2002): CANOCO reference manual and CanoDraw for Windows
user’s guide: software for canonical community ordination (version 4.5). − Microcomputer Power Ithaca, NY.
tichý L. (2002): JUICE, Software for vegetation classification. − J. Veg. Sci. 13: 451–453.yoDer j. i. (2001): Host plant recognition by parasitic Scrophulariaceae. − Curr. Opin. Plant Biol.
4: 359–365.zuber D. & wiDMer a. (2000): Genetic evidence for host specificity in the hemi-parasitic Viscum
album L. (Viscaceae). − Mol. Ecol. 9: 1069−1073.
224
Dostálek & Münzbergová: Stanovištní nároky a výběr hostitelů u rodu Thesium
Přílo
ha 1
. Prů
měr
y (M
) a
směr
odat
né o
dchy
lky
prům
ěru
(SD
) E
llenb
ergo
vých
indi
kačn
ích
hodn
ot p
ro s
větlo
, tep
lotu
, kon
tinen
talit
u, v
lhko
st, p
ůdní
re
akci
, živ
iny
a po
čet d
ruhů
ve
třech
vel
ikos
tech
plo
ch s
(+) a
bez
(-) T
hesi
um. N
= c
elko
vý p
očet
plo
ch.
App
endi
x 1.
Mea
ns (
M)
and
stan
dard
dev
iatio
n (S
Ds)
of
Elle
nber
g in
dica
tor
valu
es f
or li
ght,
tem
pera
ture
, co
ntin
enta
lity,
moi
stur
e, s
oil r
eact
ion,
nu
trien
ts, a
nd th
e nu
mbe
r of s
peci
es in
thre
e pl
ot s
izes
with
(+) a
nd w
ithou
t (-)
The
sium
. N =
sam
ple
size
.
Velik
ost p
loch
y (c
m)
2050
100
Přít
omno
st T
hesi
um+
-+
-+
-M
SD
MS
DM
SD
MS
DM
SD
MS
D
T. li
noph
yllo
n(N
= 1
45)
Poč
et d
ruhů
6,4
2,2
6,1
212
,32,
711
,13,
217
,73,
3715
,93,
68S
větlo
7,3
0,4
7,3
0,5
7,4
0,3
7,3
0,4
7,35
0,27
7,26
0,34
Tepl
ota
5,9
0,4
5,9
0,4
6,0
0,3
6,0
0,3
5,99
0,25
5,98
0,25
Kon
tinen
talit
a4,
60,
54,
70,
64,
70,
44,
60,
44,
650,
34,
630,
34V
lhko
st3,
30,
63,
40,
83,
30,
53,
40,
63,
330,
443,
410,
49P
ůdní
reak
ce7,
70,
77,
60,
77,
60,
67,
60,
77,
610,
517,
580,
58Ži
viny
2,7
0,6
2,8
0,7
2,7
0,5
2,8
0,6
2,76
0,45
2,88
0,56
T. b
avar
um(N
= 1
50)
Poč
et d
ruhů
4,1
1,9
4,1
2,0
8,7
3,1
8,4
3,1
13,2
94,
0412
,79
4,24
Svě
tlo6,
30,
86,
30,
96,
20,
76,
20,
86,
260,
606,
220,
68Te
plot
a5,
20,
95,
30,
85,
50,
55,
50,
55,
610,
325,
590,
40K
ontin
enta
lita
3,9
0,8
4,0
0,8
4,1
0,4
4,1
0,5
4,23
0,33
4,15
0,36
Vlh
kost
3,8
0,6
3,8
0,6
3,8
0,5
3,8
0,5
3,77
0,45
3,80
0,49
Půd
ní re
akce
7,3
1,3
7,3
1,0
7,4
0,8
7,3
0,7
7,41
0,54
7,31
0,59
Živi
ny3,
31,
03,
50,
93,
50,
83,
60,
93,
500,
683,
570,
75
T. e
brac
teat
um(N
= 1
22)
Poč
et d
ruhů
5,2
2,2
4,9
2,1
9,3
3,9
8,6
3,7
13,0
54,
9811
,96
4,59
Svě
tlo6,
80,
46,
90,
56,
90,
36,
90,
46,
900,
276,
900,
35Te
plot
a5,
70,
35,
70,
45,
70,
35,
70,
35,
760,
245,
740,
26K
ontin
enta
lita
4,1
0,8
4,3
0,9
4,2
0,7
4,2
0,6
4,22
0,59
4,21
0,53
Vlh
kost
4,2
0,8
4,3
14,
20,
74,
30,
94,
230,
664,
240,
77P
ůdní
reak
ce5,
91,
55,
51,
65,
91,
25,
51,
25,
930,
965,
641,
19Ži
viny
3,4
13,
61,
13,
50,
93,
60,
93,
580,
763,
570,
75
Příroda, Praha, 31: 201–225, 2011
225

226
Dostálek & Münzbergová: Stanovištní nároky a výběr hostitelů u rodu Thesium

227
Příroda, Praha, 31: 227–246, 2011
současné poznatky o mateřídoušce vejčité kraňské (Thymus pulegioides subsp. carniolicus) v České republice
current knowledge on Thymus pulegioides subsp. carniolicus in the czech Republic
JarosLav ČáP1 & hana kLeinová2
1Kšírova 182, CZ-619 00 Brno 192Agentura ochrany přírody a krajiny ČR, Správa CHKO Litovelské Pomoraví a krajské středisko Olomouc, Lafayettova 13, CZ-779 00 Olomouc, [email protected]
Abstract: This research explores taxon Thymus pulegioides subsp. carniolicus. The introduction gives an outline on the subject of a Thymus pulegioides L. complex. Hybridisation at isolated locations might lead to extinction of the taxon gene pool on this sites. The area of the taxon is mentioned as well as its probable migration to localities in the Czech Republic and Slovakia. A global survey of the population was carried out at Hněvotín location (Olomouc, Central Moravia). There are 600 specimens. The phenological observations confirmed concurrent flowering periods in Thymus pulegioides subsp. carniolicus and subsp. chamaedrys. This excludes mowing as a possible measure to eliminate subsp. chamaedrys in the period of full flowering and pollen production. Karyological investigation of the taxon and comparison conducted by flow cytometry showed that there is no possibility to distinguish the two subspecies and their hybrids; this is currently possible at localities common to both subspecies only based on their morphological signs. In two plants from Hněvotín the number of chromosomes was 2n = 28. Other plants from Hněvotín and from Janov and Vršatské Podhradie genetically correspond diploid plants. Attached you can find phytosociological relevés of the vegetation, the results of phytosociological observations and a list of discovered taxons of the genus Thymus at Hněvotín location. Keywords: Thymus pulegioides subsp. carniolicus, distribution, genetics, phenology
Abstrakt: Práce se zabývá taxonem Thymus pulegioides subsp. carniolicus. Úvod podává nástin problematiky komplexu Thymus pulegioides L. Hybridizace mezi taxony na izolovaných lokalitách může vést k zániku genofondu taxonu, který je zde na exklávním stanovišti. Je uveden areál taxonu a jeho pravděpodobná migrační cesta na lokality v České republice a na Slovensku. Bylo provedeno plošné mapování populace na lokalitě u Hněvotína (Olomouc, střední Morava); čítá ca 600 exemplářů. Fenologická pozorování potvrdila současnou dobu kvetení Thymus pulegioides subsp. carniolicus a subsp. chamaedrys. To vylučuje sečení jako možné opatření, které by eliminovalo subsp. chamaedrys v době plného kvetení a tím i produkci jeho pylu. Karyologické vyšetření taxonu a srovnání průtokovou cytometrií ukázalo, že touto metodou není možné odlišit obě subspecie a jejich křížence, to je možné na společných lokalitách zatím jen podle morfologických znaků. U dvou rostlin od Hněvotína byl zjištěn počet chromozomů 2n = 28. Další rostliny z Hněvotína a od Janova a Vršateckého Podhradie geneticky odpovídají diploidním rostlinám. Přiloženy jsou fytocenologické snímky vegetace, výsledky fenologických pozorování a seznam zjištěných taxonů rodu Thymus na lokalitě u Hněvotína.

228
Čáp & Kleinová: Současné poznatky Thymus pulegioides subsp. carniolicus v České republice
Klíčová slova: fenologie, genetika, rozšíření, Thymus pulegioides subsp. carniolicusnomenklatura: hejný & slavík 1988, hejný & slavík 1990, hejný & slavík 1992, slavík 1995, slavík 1997, slavík 2000, slavík & ŠtěPánková 2004; rod Taraxacum a jednoděložné - neuhäuslová & kolbek (1982); rod Thymus samostatně.
Úvod
Mateřídouška vejčitá, Thymus pulegioides L., tvoří složitý soubor vnitrodruhových taxonů, jejichž taxonomie a nomenklatura není dosud dostatečně vyřešena. V Evropě se vyskytuje ve čtyřech poddruzích. Thymus pulegioides subsp. pulegioides je taxon západoevropský, rozšířený od Španělska přes Francii po Anglii. Přesné rozšíření není dosud spolehlivě známo. Thymus pulegioides subsp. chamaedrys (Fries) Gusuleac je nejrozšířenějším poddruhem; roste v severní, střední, západní i východní Evropě. Thymus pulegioides subsp. carniolicus (Borbás) P. A. Schmidt má centrum výskytu na jižních svazích Alp a v Pyrenejích. Thymus pulegioides subsp. montanus (Bentham) Ronnig roste hlavně na Balkánském poloostrově, odkud vybíhá na jednotlivých lokalitách k severu do Maďarska a vzácně až na jižní Slovensko; kdysi byl sbírán i na jižní Moravě (často v netypických a přechodných formách) a dokonce existuje i takto určený doklad z Českého středohoří. V současné době se zřejmě v České republice nevyskytuje. Každá z uvedených subspecií má ještě další vnitrodruhovou (alespoň u subsp. chamaedrys geneticky potvrzenou) variabilitu.
Cílem této práce je shrnout dosavadní poznatky o Thymus pulegioides subsp. carniolicus. Mateřídoušky se mezi sebou mohou křížit, zvláště taxony stejné ploidie. U studovaných taxonů okruhu Thymus pulegioides agg. vznikají fertilní hybridní populace; z toho důvodu hrozí na lokalitě u Hněvotína, kde je největší středoevropská populace Thymus pulegioides subsp. carniolicus, vlivem hybridizace tzv. koroze genofondu této vzácné mateřídoušky.
Materiál a metody
Byla prostudována obsáhlá literární pojednání o celém komplexu Thymus pulegioides agg. a také herbářové položky v herbářích v ČR a na Slovensku (PR, PRC, BRNU, BRNM, CHOM, CHEB, MMI, VYM, HOMP, HR, GM, MZ, LIT, OLM, OLP, OP, PL, ROZ, BRNL, Rokycany; BRA, NI, SAV, SLO – zkratky podle vozárová & sutorý 2001, soukromé herbáře Čáp, Smrček, Žíla) a také v zahraničních herbářích (DR, HAL, JE – zkratky podle sweetgum.nybg.org/ih/herbarium_list.php). Lokality, uvedené v této práci, jsou podloženy herbářovými doklady a byly revidovány prvním z autorů. Řadu dokladů z Německa revidoval Peter A. Schmidt (Institut für Allgemeine Ökologie und Umweltschutz, Technische Universität Dresden), který je významným monografem středoevropských mateřídoušek.
Použité zkratky:
Th. = ThymusFRO = Thymus pulegioides subsp. carniolicusCHAM = Thymus pulegioides subsp. chamaedrysOTR = Thymus × otrubae = Th. pulegioides subsp. chamaedrys × Th. pulegioides subsp. carniolicus

229
Příroda, Praha, 31: 227–246, 2011
PAN = Thymus pannonicus var. pannonicusALB = Thymus pannonicus var. latifoliusOPI = Thymus × opizii = Th. pulegioides subsp. chamaedrys × Th. pannonicus var. pannonicusPOR = Thymus × porcii = Th. pulegioides subsp. chamaedrys × Th. pannonicus var. latifoliusPLS = Thymus pulcherrimus subsp. sudeticusostatní zkratky, včetně latinských, jsou standardními zkratkami.
V letech 2006–2007 bylo provedeno na lokalitě u Hněvotína, kterou je Národní přírodní památka (NPP) Na skále, plošné mapování mateřídoušek. Mapování bylo provedeno pomocí čtvercové sítě; mapovacím kamenem byl čtverec 1 × 1 metr. Ve čtvercích pak byly zaznamenány všechny přítomné taxony, nebyl zjišťován přesný počet rostlin, jak bylo původně zamýšleno. Důvodem použití této metody je, že trsy mateřídoušek často navzájem prorůstají a není možné bez vyrytí rostlin a jejich oddělení stanovit, o kolik jedinců se jedná. V roce 2010 bylo provedeno plošné mapování v ochranném pásmu, kam se mateřídoušky rozšířily. Jedná se o bývalé intenzivně obhospodařované pole, které bylo za účelem ochrany přírody (zamezení splachů do ZCHÚ) v roce 1999 vykoupeno. Pozemek byl následně ponechán samovolnému vývoji, nebyl osetý žádnou travní směsí, byl pouze pravidelně a intenzivně kosený v rámci managementu ZCHÚ. Postupně se sem začala šířit xerofilní vegetace ze sousedících ploch v NPP. Území bylo pro potřeby mapování (taxonomické inventarizace) rozděleno na sedm dílčích ploch. Plochy i čtvercová síť byla v terénu zafixována, aby se mohlo mapování po letech opakovat, lomové body byly zaměřeny GPS. Výsledná mapa rozšíření jednotlivých taxonů byla zpracována v prostředí ArcView a je uložena v elektronické podobě v archívu AOPK ČR krajského střediska Olomouc (v podobě vrstev shp).
Druhý z autorů prováděl v letech 2008–2010 časově náročné sledování fenologie vybraných taxonů mateřídoušek. Vzhledem k nejběžnějšímu případu hybridizace zájmového taxonu FRO s taxonem CHAM, byly vybrány hlavně tyto dva taxony. Pro doplnění informace byl do fenologického pozorování zařazen jejich kříženec OTR, dále PAN a POR. Na lokalitě bylo k fenologickému pozorování vybráno celkem 34 rostlin. Taxony FRO a CHAM byly vybrány na 12 místech lokality tak, aby tvořily vždy dvojici. Ve dvojicích byly od sebe vzdáleny cca do 0,5 m. To mělo zajistit, aby obě sledované rostliny měly vždy přibližně stejné stanovištní podmínky (půdní substrát, zastínění, expozici atd.). Na dvou místech pak byly ke dvojici rodičovských taxonů FRO a CHAM vybrány i dvě rostliny křížence (OTR). Dále bylo vybráno sedm rostlin PAN a jeden kříženec POR. Všechny vybrané rostliny byly v terénu označeny štítkem s číslem a fenologická pozorování se u nich opakovala po tři roky.
U mateřídoušek je nejdůležitější sledovat fázi kvetení rostlin, tedy dobu produkce pylu. Proto byla fenologická pozorování zúžena pouze na tuto část růstového cyklu. Pozorování byla prováděna v období od května do října, v intervalu jednou týdně. U všech sledovaných taxonů byla zaznamenána fenologická fáze kvetení do tabulky (v programu Excel). Fenologické fáze jsme rozdělili do osmičlenné stupnice (podle zlatník & zvorykin 1932, upraveno): A – květenství s mladými pupeny, B – květenství s poupaty a květy (jeden až pět rozkvetlých květů), C – rozkvétající květenství (šest rozkvetlých květů až 25 % rozkvetlých květů), D – kvetoucí květenství (25–75 % rozkvetlých květů), E – květenství částečně odkvetlé (ještě kvete 10–25 % květů), F – dokvétající květenství (kvete méně jak 10 % květů), G – odkvetlé květenství (nejvýše s jedním a dvěmi dokvétajícími květy), přidána byla kategorie 0 – rostlina bez pupenů.

230
Čáp & Kleinová: Současné poznatky Thymus pulegioides subsp. carniolicus v České republice
Byl proveden průzkum všech potenciálních lokalit výskytu FRO, dříve citovaných jako „Na Babě“ a v Náměšti na Hané. Také byly navštíveny nejbližší recentní lokality FRO v Janově a Vršateckém Podhradie.
Byla provedena karyologická analýza taxonů FRO a CHAM na Katedře botaniky PřF UP Olomouc. Ke karyologické analýze moravských rostlin nemohla být použita klíčící semena, protože rostliny mohly být opyleny pylem rozdílného taxonu, což by ovlivnilo výsledky analýzy. Bylo potřeba analyzovat vrcholové meristémy a kořínky jednotlivých rostlin. Proto bylo nezbytné odebrat jako vzorky část rostlin i s kořeny. Ke karyologické analýze byly odebrány vzorky pouze na lokalitě NPP Na skále, kde je nejbohatší populace taxonu FRO. Odebrané vzorky byly ihned převezeny do laboratoře, kde byly vrcholové meristémy a kořínky ošetřeny chladem a para di-chlor-benzenem (24 hodin) za účelem získání většího počtu mitóz. Takto ošetřený materiál byl fixován ve směsi etanol:kyselina octová (3:1) 15 minut a uložen v 70 % etanolu. Pro karyologickou analýzu byl fixovaný materiál macerován v 1N HCl při teplotě 60 °C. Po promytí vzorků v destilované vodě byl materiál přenesen na sklíčko a ve 45 % kyselině octové byly provedeny roztlaky. Preparát byl zamražen, vysušen a chromozómy byly barveny toluidinovou modří (0,1 % vodní roztok). Po obarvení byly preparáty promyty vodou, usušeny, zakápnuty glycerolem a analyzovány pod mikroskopem. Byly pořízeny fotografie.
Abychom získali dělící se pletiva, bylo snahou převést rostliny také do in vitro podmínek. Vzrostlé vrcholy byly testovány pro převod rostlin do in vitro podmínek. Testovala se koncentrační řada desinfekčního činidla (chloramin; 1 %, 5 %, 8 %) a doba sterilizace (15, 30 minut). Vydesinfikovaná pletiva byla vyseta na Murashige a Skoog médium doplněné o růstové regulátory (0,01 mg/l IBA a 0,01 mg/l BAP).
Souběžně byly prováděny studie průtokovou cytometrií v laboratoři Botanického ústavu AV ČR, v. v. i. v Průhonicích (v roce 2009) a Ústavu experimentální botaniky AV ČR v Olomouci (v roce 2010) a to z lokalit NPP Na skále, Janov a Vršatecké Podhradie (Slovensko).
V roce 2009 byly pro průtokovou cytometrii odebrány na lokalitě NPP Na skále vzorky z 22 rostlin (FRO, CHAM, OTR, OPI a PAN). Jako referenční vzorky byly zařazeny rostliny CHAM také ze tří lokalit z Drahanské vrchoviny – dvě z okolí Myslejovic a jedna od Vincencova. V roce 2010 byly odebrány vzorky celkem z 11 rostlin (FRO, CHAM) a to z lokality NPP Na skále, lokality u Janova a na Vršatci. Odběr vzorků byl proveden tak, že z každé rostliny byly nůžkami odstřiženy dvě až tři větvičky se zelenými listy, vloženy do označených igelitových pytlíků se zipem, zakápnuty kapkou vody, aby nevyschly a ještě v terénu umístěny do přenosného chladicího boxu. Následující den byly vzorky odeslány nebo převezeny k analýze do laboratoře.
Stupeň DNA ploidie byl stanoven z mladých živých listových pletiv (cytometr Partec PA II, rtuťová UV výbojka jako excitační zdroj). Pro izolaci a barvení jader zvolila laboratoř Botanického ústavu AV ČR modifikovanou metodiku podle otto (1990). Přibližně 0,5 cm nepoškozeného listového pletiva analyzované rostliny a přibližně dvojnásobné množství interního standardu (jako vhodný interní standard byly použity diploidní rostliny Bellis perennis 2C = 3,69 pg) bylo homogenizováno ostrou žiletkou spolu s 1 ml chlazeného extrakčního pufru Otto I v Petriho misce. Získaná suspenze byla přefiltrována přes 42 mm nylonový filtr do PE zkumavek a byl přidán 1 ml barvicího roztoku složeného z pufru Otto II, fluorescenčního barviva DAPI (finální koncentrace 4 μg/ml) a antioxidantu ß-mercaptoethanolu (2 μg/ml). Laboratoř Ústavu experimentální botaniky AV ČR v Olomouci zvolila pro izolaci a barvení jader standartní metodiku podle otto (1990), kdy se používají roztoky Otto I a Otto II v poměru 1:2. Výsledné histogramy byly vyhodnoceny programem FloMax. Před vlastním odběrem vzorků ke genetickým analýzám byly ze všech rostlin jako doklad odebrány položky do herbáře (herb. Čáp).

231
Příroda, Praha, 31: 227–246, 2011
V NPP Na skále byly pořízeny tři fytocenologické snímky vegetace s výskytem FRO přímo v ZCHÚ a jeden v ochranném pásmu, kam se mateřídouška začala šířit a kde jsou ještě pozůstatky postagrární vegetace. Jeden snímek byl zapsán na lokalitě u Janova a jeden na lokalitě Vršatecké Podhradie. Fytocenologické snímky byly zapsány za použití sedmičlenné Braun-Blanquetovy stupnice (viz tab. 2).
výsledky
systematické postavení taxonu
V současné době je FRO chápán buď na úrovni druhové – pod jménem Thymus froelichianus Opiz anebo na úrovni poddruhu T. pulegioides subsp. carniolicus. K prvnímu hodnocení se přiklání většina západních studií (např. Machule 1967, neGre 1968, Morales 1986, oberDorfer 1990, lauber & waGner 1998, Morales 2002), ale i některé prameny středoevropské (např. feráková et al. 2001). Středoevropské studie novější doby taxon uvádějí zpravidla na úrovni poddruhu (např. schMiDt 1971, ČáP & ŠíPoŠová 1993, ŠtěPánek & toMŠovic 2000, ČáP 2002). jalas (1974) taxon FRO snížil na úroveň variety, var. vestitus (Lange) Jalas. Na nižší taxonomické úrovni (varieta, forma) bylo publikováno 28 taxonů.
Taxony CHAM a FRO se od sebe odlišují morfologicky (především intenzita odění všech částí rostlin), fytogeograficky a částečně i karyologicky (poslední především na Pyrenejském poloostrově; viz níže v kapitole Karyologie).
Poddruh FRO je nejvíce morfologicky shodný s CHAM. Západoevropský nominátní poddruh subsp. pulegioides má také svůj protějšek se zesíleným oděním. Je velmi pravděpodobné, že jím nebude FRO, ale mohl by jím být var. vestitus (Lange) Jalas. To však potvrdí pouze další studie genetické a také typové položky Langeho. Balkánský poddruh montanus nemá silně ochlupené (některými autory označované jako „paralelní“) formy. Tyto taxonomicko-nomenklatorické otázky musí vyřešit další studie.
Rozpoznání křížence OTR je nesnadné a vyžaduje určitou zkušenost s „nakoukáním“ populací. Je třeba vzít v úvahu šíři variability hustoty odění v jednotlivých částech u FRO. Vyskytují se slaběji oděné rostliny, např. na listových čepelích. Významnější (konstantnější) znaky jsou v odění kalichů, postavení chlupů na hranách lodyh a způsob obrvení okrajů listové čepele (vícebuněčné chlupy, papilky). Snadno by mohlo dojít k záměně za křížence PAN × CHAM, kdy ochlupené rostliny Thymus pannonicus var. pannonicus zkřížené s CHAM mohou mít podobné znaky jako FRO. Přesto se tito kříženci (T. × porcii nm. moravicus Ferd. Weber, pro hybr., nom. illeg.) vyznačují odlišnými kombinacemi znaků: tvar listové čepele včetně řapíku, někdy výskyt drobných stopkatých žlázek v květenství, ne úplně goniotrichní odění květonosných větví po celé délce větve aj. Kříženci T. × „moravicus“ se nachází nezřídka v předhůří Bílých Karpat (fyt. okr. 19), řada lokalit je na střední Moravě i na Brněnsku.
Kříženec CHAM × FRO je znám jen z hněvotínské lokality; je to jeho locus classicus. Jinde v areálu FRO se buď nevyskytuje, anebo nebyl dosud zaznamenán. Společně se vyskytuje CHAM a FRO i na slovenské lokalitě u Vršatce; zde jsme však hybridogenní rostliny nezaznamenali (možná jsme neměli dostatečný časový prostor) a nezaznamenali je ani jiní znalci tohoto rodu, kteří zde botanizovali (F. Weber, Pawlowski). Na Vršatci roste také Thymus pulcherrimus subsp. sudeticus (Lyka) P. Schmidt. Je to jediné místo v areálu taxonů FRO a PLS, kde dlouhodobě oba taxony rostou spolu. PLS kvete dříve než FRO; když FRO rozkvétá, je první již téměř odkvetlý. Přesto PLS jednotlivě rozkvétá v tzv. „druhém kvetení“ a může docházet k přenosu pylu. Kříženec těchto taxonů popsán nebyl, a pokud by vzniknul, tak jedině na vršatecké lokalitě.

232
Čáp & Kleinová: Současné poznatky Thymus pulegioides subsp. carniolicus v České republice
Genetické analýzyKaryologie
Karyologická vyšetření rostlin z NPP Na skále nedopadla příliš dobře. Ve vzorcích odebraných 17. 7. 2009 se nepodařilo zachytit mitózu. Rostliny v laboratoři následně ze-žlout ly a uschly. Čtyři pokusně odebrané řízky zhruba po čtyřech týdnech pustily kořínky, z neznámých důvodů však poměrně záhy po zakořenění taky uschly.
U vzorků z druhého (opakovaného) odběru 1. 9. 2009 se podařilo zachytit malé množství mitóz. To však nedovolovalo důkladnou karyologickou analýzu, neboť chromozómy byly počítány na velmi malém množství mitóz, u kterých nebyly často jednotlivé chromozómy dobře odseparované. I tak se podařilo u dvou vzorků FRO ze vzrostlých vrcholů provést karyologickou analýzu. Počet chromozómů v těchto vzorcích byl odhadnut na 2n = 28.
Převést rostliny do in vitro podmínek a stimulovat jejich růst se nepodařilo. Při silnější desinfekci pletiva nekrotizovala, při nižší koncentraci a kratší době sterilizace pletiva naopak kontaminovala.
U vzorků odebraných 11. 8. 2010 nedovolovalo malé množství mitóz ve vzorcích důkladnou karyologickou analýzu. Vzrostlé vrcholy a kořínky nebyly již ve stavu růstu a tudíž se buňky nedělily. Po mnoha opakování experimentů se podařilo u jednoho vzorku FRO získat mitózu s chromozómy. Vzhledem k minimálnímu počtu mitóz byl počet chromozómů ve vzorku odhadnut na 2n = 56. Tetraploidní počet však nebyl u této rostliny potvrzen průto-kovou cytometrií.
Karyologický rozbor navrhujeme provést znovu. Vzhledem k získaným výsledkům není věrohodný. Pro analýzu bude zřejmě nutné zvolit jiný postup.
Průtoková cytometrie
Poměr intenzity fluorescence jednotlivých rostlin (vzorků) a standardu Bellis perennis (s jednotkovou fluorescencí) vyšel pro FRO 0,354–0,374, CHAM 0,351–0,370, OTR 0,354, OPI 0,351, PAN 0,354. Rozdíly mezi jednotlivými taxony jsou jen malé (max. 2 %). Z výsledků se dá usoudit, že všechny rostliny (vzorky) měly stejnou ploidii.
Průtokovou cytometrii proto není možné využít pro spolehlivé odlišení jednotlivých subspecií ani pro identifikaci potenciálních hybridních jedinců.
AreálItálie: jižní svahy Alp (PiGnati 1982). Rakousko: Kärnten, Tirol, Vorarlberg (lyka 1927). Jižní a severní Švýcarsko (lauber & waGner 1998). Španělsko: především úklony Pyrenejí na severu, ale jednotlivé lokality jsou až k východní části středního Španělska (Morales & GaMarra 1988). Jižní Francie (rouX 1975). Důležité jsou lokality v Německu, odkud se k nám FRO zřejmě dostal (obr. 1). Od jihu, kde je častější, postupuje jednotlivými loka li tami k severu: v severním Bavorsku u Ostheim v. d. Rhön, Elfershausen, Staffelberg, Holheim, Beilngries (schönfelDer 1975), Wolgast, Bielefeld, Halle, Magdeburg, Berlín, Je na, Nordhausen (schMiDt 1968), Frankenalb: Wallersberg (schMiDt & knaPP 1977), Pfalz, Württemberg, ojediněle u Frankfurt a. M., Baden: Kaiserstuhl (lyka 1927), Thüringen Becken und Hügelland (oberDorfer 1990). Ojedinělá lokalita se nachází na Slovensku nad obcí Vršatecké Podhradie (weber 1958, ČáP & ŠíPoŠová 1993). Údaje z Anglie se vztahují k jiným taxonům, např. k Thymus drucei Ronn.
FRO roste na bazických, vzácněji neutrálních až slabě kyselých půdách v travino-bylinných společenstvech. V některých oblastech (mimo naše území) roste i v řídkých lesích dubu pýřitého nebo kaštanovníku.
233
Příroda, Praha, 31: 227–246, 2011
obr. 1. Vybrané lokality Thymus pulegioides subsp. carniolicus v Německu a nástin předpokládaného šíření taxonu do Čech, na Moravu a na Slovensko.Fig. 1. Selected locations of Thymus pulegioides subsp. carniolicus in Germany and an outline of the expected spread of the taxon in Bohemia, Moravia, and Slovakia.
výskyt v České republiceRecentní výskytV současné době roste Thymus pulegioides subsp. carniolicus v České republice na dvou lo kalitách (obr. 2): v NPP Na skále u Hněvotína (fyt. okr. 21a) a na svahu kopce Stříbrník u Janova (fyt. okr. 75). Na druhé uvedené lokalitě byl taxon objeven až v roce 2009 a byly udávány tři trsy, vzdálené od sebe 20 m a 10 m. V roce 2010 zde byl zaznamenán pouze jediný trs ma te řídoušky kraňské. Na „nenalezení“ dvou trsů mohla mít vliv pastva skotu, která na lokalitě probíhá. Jeden trs rostl přímo na stezce, po které dobytek pravidelně chodil. Druhý nezvěstný trs rostl v malém remízku, kam si přes den chodil dobytek lehnout do stínu (Andráško, verb.). V šir ším okolí lokality u Janova roste Thymus pulegioides subsp. chamaedrys. Jediný zjištěný trs ma teřídoušky kraňské roste v okrajové části lučního porostu, u hromadnice se solitérními stromy.
Zaniklé lokalityHerbářově jsou doložena tato naleziště:- Lovosice, Lovoš (1889, R. Schmidt, herb. DR; vidi; fyt. okr. 4b),- Roudnice nad Labem (sine dat. et leg., herb. JE; zřejmě druhá polovina 19. stol.; vidi; fyt. okr. 5b),

234
Čáp & Kleinová: Současné poznatky Thymus pulegioides subsp. carniolicus v České republice
- Kroměříž, les Zámeček (1952, Zavřel, herb. PR; poměrně „záhadný“ nález; nejspíše se jedná o zplanění z vysetých rostlin z okrasných nebo aromatických záhonků; vidi; fyt. okr. 21b),- Slatinice (1921, Josef Otruba, herb. OLM; fyt. okr. 21a); lokalita pravděpodobně zanikla.
obr. 2. Rozšíření Thymys pulegioides subsp. carniolicus v České republice.Fig. 2. Occurrence of Thymys pulegioides subsp. carniolicus in the Czech Republic.
Omylem uváděné lokality
Nejsou všechny podchyceny. Většinou se jedná o determinační záměny za křížence CHAM × PAN (= Thymus × porcii nothovar. moravicus). Příkladem může být Deylův zápis z lokality u nádraží v Náměšti na Hané (Deyl 1996), ve kterém uvádí z dané lokality Thymus froelichianus, T. kosteleckyanus a T. pulegioides. Při kontrole v terénu v roce 2010 jsme zde zaznamenali výskyt CHAM, PAN a ALB a také zmíněného křížence POR.
Provedli jsme revizi dostupných herbářových položek mateřídoušek z této lokality, které Deyl uložil v herbáři OLM. V databázi Vlastivědného muzea v Olomouci je udáváno šest položek sebraných na této lokalitě. Při revizi těchto sběrů byly zjištěny taxony Thymus pannonicus a T. praecox. Na smíšené položce (pořadové číslo 135400), určené sběratelem jako T. froelichianus, byly při revizi zjištěny T. pannonicus a kříženci T. × otrubae a T. × porcii. Bohužel sběr (pořadové číslo 154003), určený sběratelem také jako T. froelichianus, se nepodařilo vzhledem k probíhajícím stavebním úpravám v muzeu a stěhování depozitáře dohledat a položka nemohla být proto revidována.
Vzhledem ke studovanému druhu a předmětné lokalitě v Hněvotíně jsme při revizi herbářových položek v OLM zjistili další dvě zajímavé skutečnosti. Byla zjištěna položka Thymus glabrescens od Hněvotína od Josefa Otruby z roku 1943. Tento taxon jsme při

235
Příroda, Praha, 31: 227–246, 2011
podrobné inventarizaci na lokalitě nenašli. Otruba rostlinu určil jako T. froelichianus. Dále byla zjištěna položka Thymus pulegioides subsp. carniolicus od J. Otruby z roku 1921. Otruba určil tuto rostlinu jako Thymus braunii. Jako lokalizaci uvádí Olomouc: Slatinice. Pořadové číslo sběru 56547. Mateřídoušky v NPP Na skále
Mateřídoušku kraňskou na stepi u Hněvotína zřejmě poprvé nalezl a sebral Podpěra (1905), taxon však rozlišil až F. Weber. Weber byl znalcem této lokality a provedl zde podrobný průzkum mateřídoušek. Uvádí od Hněvotína tyto infraspecifické taxony (pod Thymus froelichianus) (Weberem popsané taxony = nom. illeg.): var. froelichianus, var. allionii, var. murrii, var. laxifoliatus, var. magnifolius (locus classicus), var. suborbiculatus (loc. class.), var. oblongoovatus (loc. class.), var. molliterpilosus (loc. class.), var. villosiusculus (loc. class.), var. olomucensis (loc. class.), var. longepilosus (loc. class.), var. subrigidus (loc. class.), var. duriusculus (loc. class.), var. microphyllus (verosim. loc. class.), var. carniolicus, var. effusoides (loc. class.), var. pseudorepens (loc. class.). Zřejmě všechny tyto variety nemají taxonomické opodstatnění (alespoň ne na této úrovni). Dále zde uvádí křížence, z nichž jeden z rodičů je Thymus pulegioides: Thymus × opizii (loc. class.), Thymus × otrubae (loc. class.), Thymus × spitzneri (loc. class.) - pozn.: tento kříženec je „vymyšlený“, jeden z rodičů (T. pulegioides subsp. montanus) na lokalitě, ani v blízkém okolí, neroste.
Na pruhu výchozů vápence mezi Hněvotínem a Žerůvkami byly pravděpodobně čtyři mikrolokality, kde se FRO vyskytoval. Současný výskyt v NPP Na skále má středové souřadnice ca 49°33‘21.157“N, 17°10‘40.895“E. Ještě v roce 1977 sbíral první z autorů FRO ve vápencovém lomu, který byl zavážen odpadem. Nachází se u silničky Hněvotín – Žerůvky asi 1,63 km JJZ od kostela v Hněvotíně, se středem ca 49°33‘29.165“N, 17°10‘14.567“E. Třetí lokalita se nacházela 1,1 km SSZ od kaple v Žerůvkách. Byla zničena stavbou dálnice; její střed byl asi 49°33‘8.569“N, 17°10‘55.865“E. Čtvrtou lokalitou, již zcela zavezenou a rekultivovanou, byl vápencový lom 670 m S – SSZ od kaple v Žerůvkách se středem ca 49°32‘48.211“N, 17°11‘34.576“E. Celý tento hřeben byl nazýván prvorepublikovými botaniky jako „Baba u Hněvotína“ nebo „Žerůvky“.
Při průzkumu těchto lokalit (mikrolokalit) „Na Babě“ v letech 2008–2009 jsme Thymus pulegioides subsp. carniolicus nalezli pouze v NPP Na skále.
NPP Na skále leží asi 1 km JV od obce Hněvotín. Nadmořská výška lokality je 242–263 m. Geologické podloží je budováno devonskými vápenci. V minulosti sloužily výchozy vápence jako stavební materiál a území sloužilo jako lokální vápencový lom. Vzniklo tak území s členitým povrchem s drobnými výchozy vápenců. Vápence této lokality jsou svým horninovým charakterem a vývojem blízké vápencovému vývoji Moravského krasu. Většinu území (svahy a skalnaté výchozy) pokrývá mělká, kamenitá rendzina modální. Pouze v nejnižších partiích území, středem mělkého údolí, je černozem černická. V okolí ZCHÚ pak převažuje černozem modální (zdroj: Pedologická mapa ČR). Území leží v teplé oblasti T2, která je charakterizovaná dlouhým létem, teplým a suchým, velmi krátkým přechodným obdobím s teplým až mírně teplým jarem a podzimem, krátkou, mírně teplou, suchou až velmi suchou zimou, s velmi krátkým trváním sněhové pokrývky (Quitt 1971).
Při podrobném mapování mateřídoušek pomocí čtvercové sítě byla mapována severovýchodní část NPP, ve které roste Thymus pulegioides subsp. carniolicus spolu s dalšími taxony. Severozápadní část NPP podrobně mapována nebyla, protože zde roste pouze T. pannonicus a od zájmové plochy je oddělena porostem dřevin. Zájmová část lokality je znázorněna na následujícím obrázku (obr. 3).

236
Čáp & Kleinová: Současné poznatky Thymus pulegioides subsp. carniolicus v České republice
obr. 3. Zájmová část NPP Na skále s výskytem Thymus pulegioides subsp. carniolicus, ve které proběhlo podrobné síťové mapování.Fig. 3. Focal area of National Nature Monument Na skále with occurrence of Thymus pulegioides subsp. carniolicus, where detailed network mapping was conducted.
Při inventarizaci byl jako nejčastější taxon zjištěn FRO, v 687 čtvercích, což je více než polovina. Plocha, na které FRO roztroušeně až místy hojně roste, je ca 0,24 ha. Na celé ploše roste společně s CHAM. Ve vlastní NPP roste na SZ orientovaném svahu, ve vykoupeném ochranném pásmu na JV orientovaném. Roste jak v rovném, mírně svažitém terénu se stepní vegetací, tak v mozaice stepní vegetace a křovin (včetně odrostlejších stromů) ve členitém terénu s terénními depresemi a výstupy vápencových skalek a suti.
Výsledky inventarizace, přehled zjištěných taxonů a jejich četnost, jsou shrnuty v následující tabulce (tab. 1). V mapě je pak znázorněno rozšíření jednotlivých taxonů plošně (obr. 4).
Vzhledem k měřítku musela být mapa pro přehlednost zjednodušena a proto se v ní neuvádí rozšíření PAN, které se částečně překrývá s výskytem FRO a CHAM. Není však nutné, protože PAN se jako rodič podílí na hybridizaci s FRO zanedbatelně – bylo zjištěno pouze šest případů. Tito kříženci (OPI) v mapce zakresleni jsou a jsou v místech, kde se na lokalitě potkávají rodičovské taxony FRO a PAN.
237
Příroda, Praha, 31: 227–246, 2011
tab. 1. Taxony zaznamenané v Národní přírodní památce Na skále při síťovém mapování a četnost jejich výskytu.table 1. Taxa recorded in National Natural Monument Na skále in network mapping and frequency of their occurrence.
taxonPočet čtverců s výskytem taxonu Zastoupení v %
(zaokrouhleno)vlastní nPP ochr. pásmo nPP
Thymus pulegioides subsp. carniolicus 647 40 54
Thymus pulegioides subsp. chamaedrys 357 45 32
Thymus pannonicus var. pannonicus 59 – 4,6
Thymus × otrubae 104 7 8,7
Thymus × opizii 6 – 0,5
Thymus × porcii 2 – 0,2
Podle současně přijímané taxonomické klasifikace (např. wisskirchen & haeuPler 1998) patří mateřídoušky zaznamenané při inventarizaci v NPP Na skále k těmto taxonům: Thymus pulegioides L. subsp. chamaedrys (Fries) Gusuleac var. chamaedrysT. pulegioides L. subsp. chamaedrys (Fries) Gusuleac var. parvifolius (Opiz ex Borbás) P. A. SchmidtT. pulegioides L. subsp. carniolicus (Borbás) P. A. SchmidtT. pannonicus All. var. pannonicusT. pannonicus var. latifolius (Besser) Jalas – nebyl zjištěn, ale byly nalezeny dvě rostliny křížence, kde je tento taxon jedním z rodičů T. pulegioides L. subsp. chamaedrys × subsp. carniolicus = T. × otrubae Ferd. Weber, nom. illeg. T. pulegioides subsp. chamaedrys × T. pannonicus var. latifolius = T. × porcii Borbás nothovar. pilisiensis (Borbás) P. A. SchmidtT. pulegioides subsp. chamaedrys × T. pannonicus var. pannonicus = T. × porcii Borbás nothovar. moravicus (Weber, pro hybr.); (doklad v herb. PR, sine dat., leg. Weber)T. pulegioides subsp. carniolicus × T. pannonicus var. pannonicus = T. × opizii Ferd. Weber, nom. illeg.
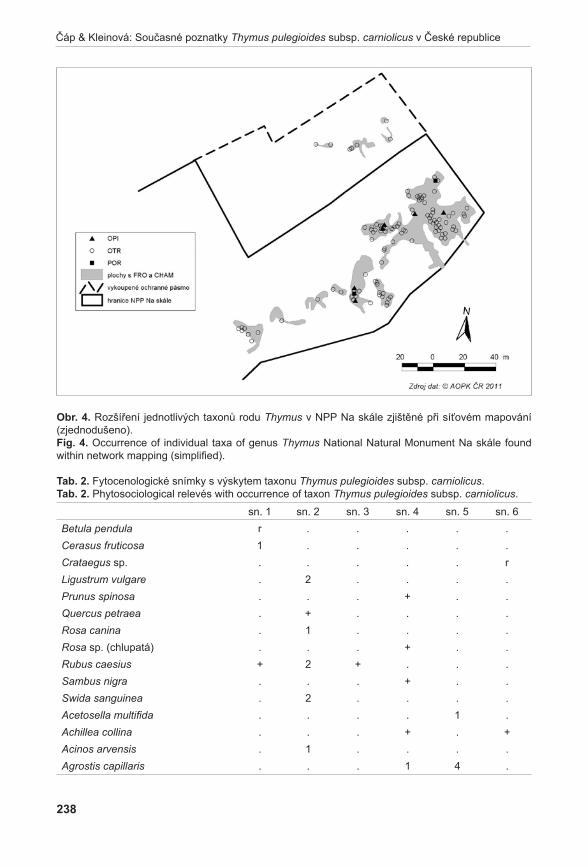
238
Čáp & Kleinová: Současné poznatky Thymus pulegioides subsp. carniolicus v České republice
obr. 4. Rozšíření jednotlivých taxonů rodu Thymus v NPP Na skále zjištěné při síťovém mapování (zjednodušeno).Fig. 4. Occurrence of individual taxa of genus Thymus National Natural Monument Na skále found within network mapping (simplified).
tab. 2. Fytocenologické snímky s výskytem taxonu Thymus pulegioides subsp. carniolicus.tab. 2. Phytosociological relevés with occurrence of taxon Thymus pulegioides subsp. carniolicus.
sn. 1 sn. 2 sn. 3 sn. 4 sn. 5 sn. 6Betula pendula r . . . . .Cerasus fruticosa 1 . . . . .Crataegus sp. . . . . . rLigustrum vulgare . 2 . . . .Prunus spinosa . . . + . .Quercus petraea . + . . . .Rosa canina . 1 . . . .Rosa sp. (chlupatá) . . . + . .Rubus caesius + 2 + . . .Sambus nigra . . . + . .Swida sanguinea . 2 . . . .Acetosella multifida . . . . 1 .Achillea collina . . . + . +Acinos arvensis . 1 . . . .Agrostis capillaris . . . 1 4 .
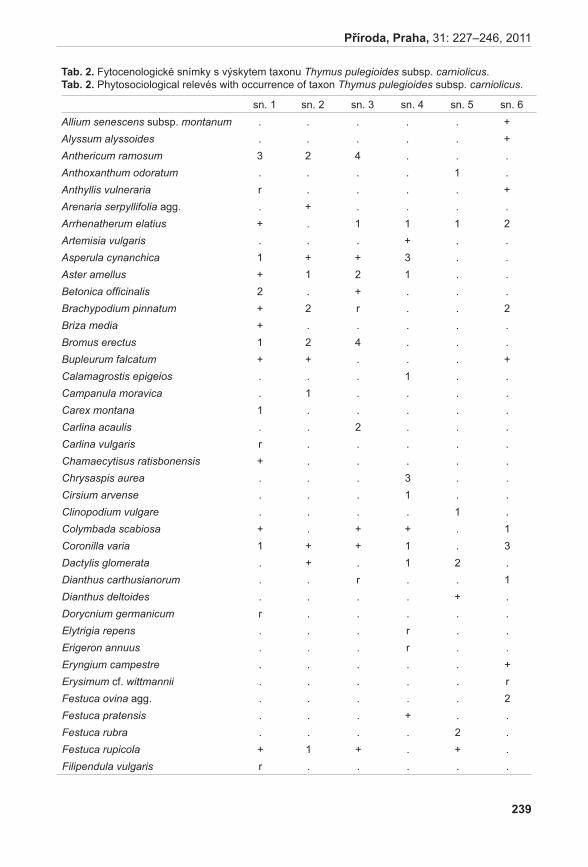
239
Příroda, Praha, 31: 227–246, 2011
tab. 2. Fytocenologické snímky s výskytem taxonu Thymus pulegioides subsp. carniolicus.tab. 2. Phytosociological relevés with occurrence of taxon Thymus pulegioides subsp. carniolicus.
sn. 1 sn. 2 sn. 3 sn. 4 sn. 5 sn. 6Allium senescens subsp. montanum . . . . . +Alyssum alyssoides . . . . . +Anthericum ramosum 3 2 4 . . .Anthoxanthum odoratum . . . . 1 .Anthyllis vulneraria r . . . . +Arenaria serpyllifolia agg. . + . . . .Arrhenatherum elatius + . 1 1 1 2Artemisia vulgaris . . . + . .Asperula cynanchica 1 + + 3 . .Aster amellus + 1 2 1 . .Betonica officinalis 2 . + . . .Brachypodium pinnatum + 2 r . . 2Briza media + . . . . .Bromus erectus 1 2 4 . . .Bupleurum falcatum + + . . . +Calamagrostis epigeios . . . 1 . .Campanula moravica . 1 . . . .Carex montana 1 . . . . .Carlina acaulis . . 2 . . .Carlina vulgaris r . . . . .Chamaecytisus ratisbonensis + . . . . .Chrysaspis aurea . . . 3 . .Cirsium arvense . . . 1 . .Clinopodium vulgare . . . . 1 .Colymbada scabiosa + . + + . 1Coronilla varia 1 + + 1 . 3Dactylis glomerata . + . 1 2 .Dianthus carthusianorum . . r . . 1Dianthus deltoides . . . . + .Dorycnium germanicum r . . . . .Elytrigia repens . . . r . .Erigeron annuus . . . r . .Eryngium campestre . . . . . +Erysimum cf. wittmannii . . . . . rFestuca ovina agg. . . . . . 2Festuca pratensis . . . + . .Festuca rubra . . . . 2 .Festuca rupicola + 1 + . + .Filipendula vulgaris r . . . . .

240
Čáp & Kleinová: Současné poznatky Thymus pulegioides subsp. carniolicus v České republice
tab. 2. Fytocenologické snímky s výskytem taxonu Thymus pulegioides subsp. carniolicus.tab. 2. Phytosociological relevés with occurrence of taxon Thymus pulegioides subsp. carniolicus.
sn. 1 sn. 2 sn. 3 sn. 4 sn. 5 sn. 6Fragaria viridis r + + 2 . .Galium album . . . 2 + .Galium glaucum . r + . . .Galium mollugo . . . 1 . .Galium verum 1 + . . . 1
Gnaphalium sylvaticum . . . . + .
Helianthemum grandiflorum subsp. obscurum . . . . . 2
Hieracium murorum . + + . . .Hieracium pilosella . . . . + .Hieracium umbellatum . + . + . .Hieracium subg. Pilosella . . . . + .Hylotelephium maximum . . . . + .Hypericum perforatum . + . + 1 1Inula hirta r . . . . .Knautia kitaibelii + r + . . .Koeleria pyramidata r . + . . .Lathyrus tuberosus . . + . . .Lembotropis nigricans 2 . 1 . . .Leontodon hispidus . . 1 . . .Linum catharticum + + . . . .Lolium perenne . . . r . .Lotus corniculatus . . 1 . . .Luzula multiflora . . . . + .Medicago falcata . + 1 . . .Medicago lupulina . . . . . 1Peucedanum alsaticum . 1 1 2 . .Peucedanum cervaria 2 . + . . .Picris hieracioides , , , 2 . .Pimpinella saxifraga . . . . + .Plantago lanceolata . . . 1 . .Plantago major . . . + . .Plantago media r . . 1 . .Poa angustifolia . . . 4 . .Poa compressa . 1 . . . .Poa pratensis . . . . + .Polygala vulgaris + . + . . .Potentilla heptaphylla 1 . . . . rPotentilla recta r . . .
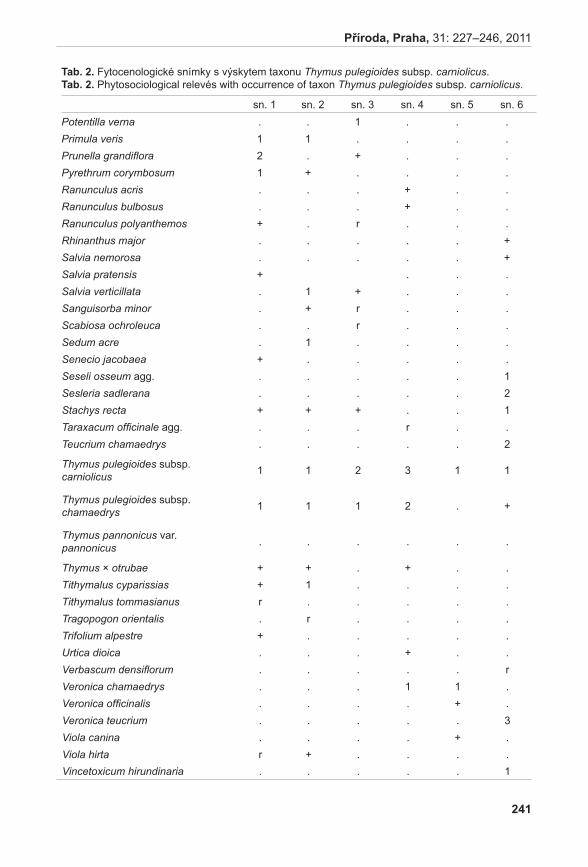
241
Příroda, Praha, 31: 227–246, 2011
tab. 2. Fytocenologické snímky s výskytem taxonu Thymus pulegioides subsp. carniolicus.tab. 2. Phytosociological relevés with occurrence of taxon Thymus pulegioides subsp. carniolicus.
sn. 1 sn. 2 sn. 3 sn. 4 sn. 5 sn. 6Potentilla verna . . 1 . . .Primula veris 1 1 . . . .Prunella grandiflora 2 . + . . .Pyrethrum corymbosum 1 + . . . .Ranunculus acris . . . + . .Ranunculus bulbosus . . . + . .Ranunculus polyanthemos + . r . . .Rhinanthus major . . . . . +Salvia nemorosa . . . . . +Salvia pratensis + . . .Salvia verticillata . 1 + . . .Sanguisorba minor . + r . . .Scabiosa ochroleuca . . r . . .Sedum acre . 1 . . . .Senecio jacobaea + . . . . .Seseli osseum agg. . . . . . 1Sesleria sadlerana . . . . . 2Stachys recta + + + . . 1Taraxacum officinale agg. . . . r . .Teucrium chamaedrys . . . . . 2
Thymus pulegioides subsp. carniolicus 1 1 2 3 1 1
Thymus pulegioides subsp. chamaedrys 1 1 1 2 . +
Thymus pannonicus var. pannonicus . . . . . .
Thymus × otrubae + + . + . .Tithymalus cyparissias + 1 . . . .Tithymalus tommasianus r . . . . .Tragopogon orientalis . r . . . .Trifolium alpestre + . . . . .Urtica dioica . . . + . .Verbascum densiflorum . . . . . rVeronica chamaedrys . . . 1 1 .Veronica officinalis . . . . + .Veronica teucrium . . . . . 3Viola canina . . . . + .Viola hirta r + . . . .Vincetoxicum hirundinaria . . . . . 1

242
Čáp & Kleinová: Současné poznatky Thymus pulegioides subsp. carniolicus v České republice
Snímek č. 1 – loc.: NPP Na skále, 49°33’20.407‘‘N, 17°10’39.467‘‘E; pokryvnost: E3: 0 %, E2: 4 %, E1: 98 %, E0: 30 % (indet.); plocha: 4 × 4 m; svah: 12°; exp. 310°; dat.: 25. 6. 2008; not. Jaroslav Čáp; pozn.: spol. sv. Bromion, facie s dom. Anthericum ramosum.Snímek č. 2 – loc.: NPP Na skále, 49°33’21.204‘‘N, 17°10’43.535‘‘E; pokryvnost: E3: 0 %, E2: 10 %, E1: 60 %, E0: 40 % (indet.); plocha: 5,8 × 3,8 m; svah: 32°; exp. 290°; dat.: 25. 6. 2008; not. Jaroslav Čáp; pozn.: spol. s převahou druhů sv. Bromion v nezapojených porostech na skalkách a jejich římsách (CaCO3).Snímek č. 3 – loc.: NPP Na skále, 49°33’23.239‘‘N, 17°10’45.567‘‘E; pokryvnost: E3: 0 %, E2: 1 %, E1: 85 %, E0: 10 % (indet.); plocha: 4 × 4 m; svah: 10°; exp. 300°; dat.: 25. 6. 2008; not. Jaroslav Čáp; pozn.: spol. s druhy sv. Bromion.Snímek č. 4 – loc.: Ochranné pásmo NPP Na skále (vykoupená část), 49°33’407‘‘N, 17°10’45.712‘‘E; pokryvnost: E3: 0 %, E2: 1 %, E1: 95 %, E0: 3 % (indet.); plocha: 4 × 4 m; svah: 1–2°; exp. 154°; dat.: 4. 8. 2010; not. Jaroslav Čáp; pozn.: sukcesní stádium se zbytky postagrárních lad a s nástupem druhů spol. sv. Bromion.Snímek č. 5 – loc.: Janov, Velká Stříbrná, 50°15’245‘‘N, 17°28’151‘‘E; pokryvnost: E3: 0 %, E2: 10 %, E1: 90 %, E0: 0 %; plocha: 4 × 4 m; svah: 10–12°; exp. 150°; dat.: 5. 8. 2010; not. Jaroslav Čáp; pozn.: nevyhraněná cenóza travního biotopu se sklony ke sv. Violion caninae.Snímek č. 6 – loc.: PR Vršatecká bradlá (SK), 50°15’254‘‘N, 17°28’151‘‘E; pokryvnost: E3: 0 %, E2: 1 %, E1: 90 %, E0: 5 % (indet.); plocha: 4 × 4 m; svah: 25–30°; exp. 220°; dat.: 6. 8. 2010; not. Jaroslav Čáp et Hana Kleinová; pozn.: travnatá cenóza vápencových svahů.
Fenologická pozorováníRozdíly v době kvetení FRO, CHAM a OTR nejsou velké. U sledovaných dvojic převažuje varianta, že FRO vykvetla o něco dříve (o pět až sedm dní) než párová CHAM nebo vykvetly obě ve stejnou dobu. Většinou také odkvetla FRO dříve než CHAM. Kříženec je pak fenologicky mezi oběma rodiči. Rozdíly v průběhu fenologie však byly mezi konkrétními mikrolokalitami. Do hry vstupuje vliv stanoviště (např. zastínění okolní vegetací, expozice ke světovým stranám, skalní podklad), průběh počasí i to, zda mají rostliny oboupohlavné nebo pouze samičí květy. Nastaly proto situace, kdy na jedné mikrolokalitě byla rostlina FRO bez pupenů (kategorie 0) a na vedlejší mikrolokalitě už byla rostlina FRO s částečně odkvetlým květenstvím (kategorie E). To platí i pro rostliny CHAM.
Rozdíly v nástupu jednotlivých fenologických fází a délkou jejich trvání v jednotlivých letech souvisí hlavně s průběhem počasí v daném roce a množstvím srážek zejména v červnu a červenci.
Nástup prvních fenologických fází A-D, tedy od květenství s mladými pupeny po kvetoucí květenství, bylo poměrně rychlé. Většinou se v týdenním intervalu monitoringu ani nepodařilo u jednotlivých rostlin zachytit všechny fenologické fáze. Fáze E-F, tj. květenství částečně odkvetlé až dokvétající, trvalo poměrně déle a s koncem sezóny se dokvétání ještě zpomalovalo a prodlužovalo, což jistě souvisí s relativně chladnějším počasím (studenější noci, ranní rosa, kratší dny apod.).
Při fenologickém pozorování jsme zjistili, že se všechny rodičovské taxony (FRO, CHAM, PAN) a také jejich kříženci, dobou produkce pylu více či méně překrývají, tzn. že alespoň některé z květů rodičovských taxonů kvetou ve stejnou dobu.
Následující tabulka (tab. 3.) shrnuje průběh fenologických fází u sledovaných rostlin. V tabulce není uvedena kategorie 0 (rostliny bez pupenů) a A (květenství s mladými pupeny), protože se nejedná o fázi produkce pylu. Kategorie B a C je v tabulce uvedena společně, protože nástup těchto fází je velmi rychlý a u sledovaných rostlin se hodně překrývá, tabulka je tak přehlednější.
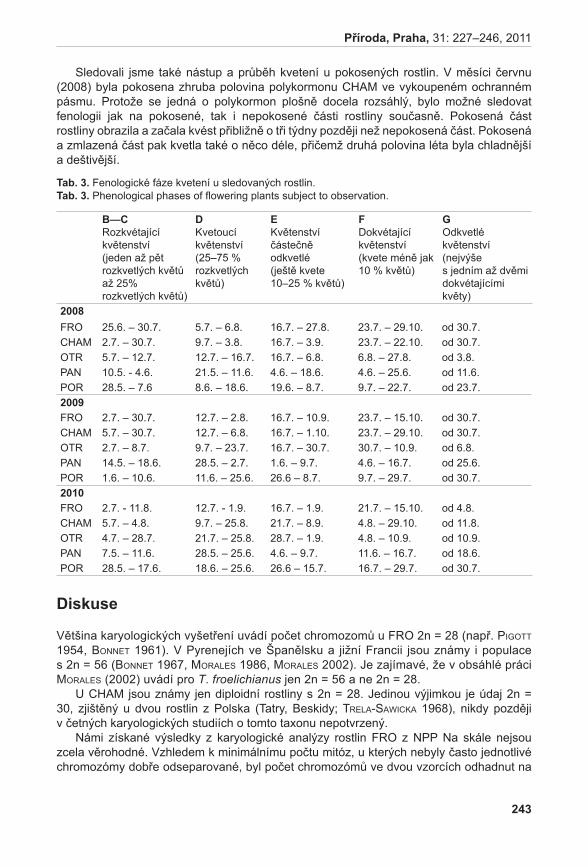
243
Příroda, Praha, 31: 227–246, 2011
Sledovali jsme také nástup a průběh kvetení u pokosených rostlin. V měsíci červnu (2008) byla pokosena zhruba polovina polykormonu CHAM ve vykoupeném ochranném pásmu. Protože se jedná o polykormon plošně docela rozsáhlý, bylo možné sledovat fenologii jak na pokosené, tak i nepokosené části rostliny současně. Pokosená část rostliny obrazila a začala kvést přibližně o tři týdny později než nepokosená část. Pokosená a zmlazená část pak kvetla také o něco déle, přičemž druhá polovina léta byla chladnější a deštivější.
tab. 3. Fenologické fáze kvetení u sledovaných rostlin.tab. 3. Phenological phases of flowering plants subject to observation.
B—c Rozkvétající květenství(jeden až pět rozkvetlých květů až 25% rozkvetlých květů)
dKvetoucí květenství(25–75 % rozkvetlých květů)
EKvětenství částečně odkvetlé(ještě kvete 10–25 % květů)
FDokvétající květenství (kvete méně jak 10 % květů)
GOdkvetlé květenství (nejvýše s jedním až dvěmi dokvétajícími květy)
2008FRO 25.6. – 30.7. 5.7. – 6.8. 16.7. – 27.8. 23.7. – 29.10. od 30.7.CHAM 2.7. – 30.7. 9.7. – 3.8. 16.7. – 3.9. 23.7. – 22.10. od 30.7. OTR 5.7. – 12.7. 12.7. – 16.7. 16.7. – 6.8. 6.8. – 27.8. od 3.8.PAN 10.5. - 4.6. 21.5. – 11.6. 4.6. – 18.6. 4.6. – 25.6. od 11.6.POR 28.5. – 7.6 8.6. – 18.6. 19.6. – 8.7. 9.7. – 22.7. od 23.7.2009FRO 2.7. – 30.7. 12.7. – 2.8. 16.7. – 10.9. 23.7. – 15.10. od 30.7.CHAM 5.7. – 30.7. 12.7. – 6.8. 16.7. – 1.10. 23.7. – 29.10. od 30.7.OTR 2.7. – 8.7. 9.7. – 23.7. 16.7. – 30.7. 30.7. – 10.9. od 6.8.PAN 14.5. – 18.6. 28.5. – 2.7. 1.6. – 9.7. 4.6. – 16.7. od 25.6.POR 1.6. – 10.6. 11.6. – 25.6. 26.6 – 8.7. 9.7. – 29.7. od 30.7.2010FRO 2.7. - 11.8. 12.7. - 1.9. 16.7. – 1.9. 21.7. – 15.10. od 4.8.CHAM 5.7. – 4.8. 9.7. – 25.8. 21.7. – 8.9. 4.8. – 29.10. od 11.8.OTR 4.7. – 28.7. 21.7. – 25.8. 28.7. – 1.9. 4.8. – 10.9. od 10.9.PAN 7.5. – 11.6. 28.5. – 25.6. 4.6. – 9.7. 11.6. – 16.7. od 18.6.POR 28.5. – 17.6. 18.6. – 25.6. 26.6 – 15.7. 16.7. – 29.7. od 30.7.
diskuse
Většina karyologických vyšetření uvádí počet chromozomů u FRO 2n = 28 (např. PiGott 1954, bonnet 1961). V Pyrenejích ve Španělsku a jižní Francii jsou známy i populace s 2n = 56 (bonnet 1967, Morales 1986, Morales 2002). Je zajímavé, že v obsáhlé práci Morales (2002) uvádí pro T. froelichianus jen 2n = 56 a ne 2n = 28.
U CHAM jsou známy jen diploidní rostliny s 2n = 28. Jedinou výjimkou je údaj 2n = 30, zjištěný u dvou rostlin z Polska (Tatry, Beskidy; trela-sawicka 1968), nikdy později v četných karyologických studiích o tomto taxonu nepotvrzený.
Námi získané výsledky z karyologické analýzy rostlin FRO z NPP Na skále nejsou zcela věrohodné. Vzhledem k minimálnímu počtu mitóz, u kterých nebyly často jednotlivé chromozómy dobře odseparované, byl počet chromozómů ve dvou vzorcích odhadnut na

244
Čáp & Kleinová: Současné poznatky Thymus pulegioides subsp. carniolicus v České republice
2n = 28 a v jednom vzorku byl odhadnut na 2n = 56. Karyologický rozbor je potřeba provést znovu a pro analýzu zvolit jiný vhodný postup.
Genofond mateřídoušky kraňské na lokalitě NPP Na skále se vlivem značné vzdálenosti od centra rozšíření tohoto taxonu mohl pozměnit a nemusí již souhlasit s populacemi v jižním Německu či na jižních svazích Alp. To je, mimo jiné, důvodem pokusit se o jeho záchranu na jeho reliktním stanovišti a to nejen do doby jeho důkladného prozkoumání.
ZávěrMateřídouška kraňská je ve střední Evropě velmi vzácnou rostlinou. Na naše území se dostala, pravděpodobně, netypickou cestou pro jiné rostliny – z jihozápadu Evropy obchvatem hradby hraničních hor a průnikem kaňonem Labe do Čech a Slezskou nížinou na Moravu a Slovensko. Dříve se vyskytovala na více lokalitách než v současnosti. Nejpočetnější populace je nyní v NPP Na skále v části, která leží v k. ú. Žerůvky (Olomoucko). Vyskytuje se zde v současnosti zhruba 600 rostlin. Na lokalitě se kříží s CHAM a vzácně i s PAN. Souběžná doba kvetení CHAM a FRO neumožňuje potlačování tlaku pylu CHAM na rostliny FRO tím, že by se v určitou dobu vybrané porosty pokosily. Není zřejmě jiná možnost, než postupná redukce rostlin CHAM ručním vytrháváním jednotlivých trsů. To bude práce zdlouhavá a nebude provedena najednou. Jde o to, snížit počet polykormonů CHAM, případně kříženců OTR.
Pro Hněvotínskou step je mateřídouška kraňská nejdůležitějším taxonem, kterému by měl být podřízen management třeba i na úkor potlačení jiných rostlin, případně živočichů na ně vázaných.
shrnutí Thymus pulegioides L. subsp. carniolicus se recentně vyskytuje v ČR na dvou lokalitách. Populace jsou diploidní s 2n = 28. Rozlišení subsp. chamaedrys a subsp. carniolicus není možné pomocí metody průtokové cytometrie. Totéž platí i pro křížence obou subspecií. Vzhledem k hybridizaci mezi oběma poddruhy může dojít k zániku genofondu subsp. carniolicus. Obě subspecie kvetou na stanovišti NPP Na skále, kde je největší populace subsp. carniolicus v ČR, ve stejnou dobu. Tlak subsp. chamaedrys, která roste na tisícovkách lokalit v ČR, na genofond subsp. carniolicus je zjevný.
summaryAt present, Thymus pulegioides L. subsp. carniolicus is found at two localities in the Czech Republic. The populations are diploid with 2n = 28. The subsp. chamaedrys and subsp. carniolicus cannot be distinguished using the flow cytometry method. The same applies to the hybrids of both subspecies. There is a risk of extinction of the gene pool of the carniolicus subsp. due to hybridisation of the two subspecies. Both subspecies in National Natural Monument Na skále (site with the highest population of carniolicus subsp. in the Czech Republic) produce flowers at the same time. The pressure of chamaedrys subsp., which grows on thousands of localities in the Czech Republic, on the gene pool of carniolicus is evident.

245
Příroda, Praha, 31: 227–246, 2011
PoděkováníAutoři srdečně děkují za spolupráci Janě Rauchové z Botanického ústavu AV ČR, v.v.i., v Průhonicích a Mgr. Janě Čížkové z Ústavu experimentální botaniky AV ČR v Olomouci za provedená měření průtokovou cytometrií, Doc. RNDr. Vladanu Ondřejovi, Ph.D., z katedry botaniky PřF UP Olomouc za karyologická vyšetření, Ing. Jiřímu Danihelkovi, Ph.D., z Ústavu botaniky a zoologie MU v Brně za cenné rady a korektury při sestavování taxonomických pasáží, Ing. Mgr. Michalu Tomáškovi z AOPK ČR za pomoc a rady při zpracování map síťového mapování. Práce byla podpořena projektem MŠMT VaV 2B06178.
literatura bonnet a. l. M. (1961): Contribution a l´étude Caryologique du genra Thymus L. II. A propos de
Thymus nitens Lamotte et de Thymus froelichianus Opiz. – Natur. Monsp. 13: 7–13.bonnet a. l. M. (1967): Contribution a l´étude Caryologique du genra Thymus L. V. A propos de
Thymus alpigenus Kerner, Thymus mastichina L., Thymus polytrichus Kerner et Thymus pulegioides L. – Natur. Monsp. 18: 193–199.
ČáP j. & ŠíPoŠová h. (1993): 31. Thymus L. Dúška (materina dúška). – In: bertová l. & GoliáŠová k. [red.], Flóra Slovenska V/1, pp. 335–367, Veda, Bratislava.
ČáP j. (2002): 27. Thymus L. – mateřídouška. – In: kubát k., hrouDa l., chrtek j. jun., kaPlan z. & ŠtěPánek j. [eds], Klíč ke květeně České republiky, pp. 595–598, Academia, Praha.
Deyl Č. (1996): Hliník a úvoz u železniční stanice Náměšť na Hané. – Ms. [Depon. in: Agentura ochrany přírody a krajiny ČR, středisko Olomouc.]
feráková v., MaGlocký Š. & MarholD k. (2001): Červený zoznam papraďorastov a semenných rastlín Slovenska. – In: baláž, D., MarholD, k. & urban, P. [eds], Červený zoznam rastlín a živočíchov Slovenska, Ochr. Prír. (Banská Bystrica), Supl., 20: 44–77.
jalas j. (1974): Notes on Thymus L. (Labiatae) in Europe III. – Ann. Bot. Fenn. 11: 262–266.lauber k. & waGner G. (1998): Thymus serpyllum L. agg. – In: laber k. & waGner k. [eds.], Flora
Helvetica, 2. Aufl., pp. 882–885, Bern, Stuttgart, Wien.lyka k. (1927): Thymus serpyllum L. – In: heGi G. [ed.], Illustrierte Flora von Mitteleuropa 5/4, pp.
2306–2327, München. Machule M. (1967): Materialen für die Monographie der Gattung Thymus L. – Ms.Morales r. (1986): Notas citotaxonómicas sobre algunos tomillos ibéricos y norteafricanos (Thymus
L., Labiatae). – An. Jard. Bot. Madrid 43 (1): 35–41.Morales r. (2002): The history, botany and taxonomy of the genus Thymus. – In: stahl-biskuP e. &
sáez f. [eds], Thyme. The genus Thymus, pp. 1–43, Taylor and Francis, London and New York.Morales r. & GaMarra r. (1988): Distribution de Thymus sect. Serpyllum en la Península Iberica. –
Lagascalia 15: 457–463.neGre r. (1968): Note sur Thymus serpyllum L. dans la flore pyrénéene. – Collect. Botan. VII (2), 50:
895–916.neuhäuslová z. & kolbek j. (1982): Seznam vyšších rostlin, mechorostů a lišejníků střední Evropy,
užitých v bance geobotanických dat BÚ ČSAV. – Průhonice.oberDorfer e. (1990): Thymian, Quendel, Thymus L. – In: oberDorfer e. [ed.], Pflanzensoziologische
Exkursionsflora, 6. Aufl., pp. 813–813, Ulmer, Stuttgart.otto f. (1990): DAPI staining of fixed cells for high-resolution flow cytometry of nuclear DNA. – In:
crissMan h. a. & Darzynkiewicz z. [eds], Methods in Cell Biology. Vol. 33, pp. 105–110, Academic Press, New York.
PiGnati s. (1982): Thymus L. – In: PiGnati s. [ed.], Flora d´Italia 2, pp. 488–493, Bologna.PiGott c. D. (1954): Species delimitation and radical divergence in British Thymus. – New Phytol. 53:
470–495.Quitt e. (1971): Klimatické oblasti Československa. – Brno.rouX G. (1975): Thymus L. – In: Guinochet M. & De vilMorin r. [eds]: Flore de France, Tome 2, pp.
694–697, CRNS et Doin, Paris.

246
Čáp & Kleinová: Současné poznatky Thymus pulegioides subsp. carniolicus v České republice
schMiDt P. (1968): Beitrag zur Kenntnis der Gattung Thymus L. in Mitteldeutschland. – Hercynia 5: 385–419.
schMiDt P. (1971): Die mitteleuropäischen Arten der Gattung Thymus L. unter besonderer Berücksichtigung von Wuchsform, Verbreitung und soziologischen Verhalten. – Ms. [Depon. in: Fakult. f. Naturwiss. d. Wiss. Rates d. Martin-Luther-Universität, Dresden.]
schMiDt P. & knaPP h. D. (1977): Die Arten der Gattung Thymus L. (Labiatae) im herzynischen Florengebiet. – Wiss. Z. Univ. Halle 26 (2): 71–118.
schönfelDer P. (1975): Zur Unterscheidung der einheimischen Thymus-Sippen und ihrer Verbreitung in Bayern. – Gött. Florist. Rundbr. 9 (3): 70–84.
ŠtěPánek j. & toMŠovic P. (2000): 29. Thymus L. – mateřídouška. – In: slavík b. [ed.], Květena České republiky, Vol. 6, pp. 656–669, Academia, Praha.
trela-sawicka z. (1968): Badania cytologiczne nad rodzajem Thymus L. – Acta biol. Cracov. Bot. 11: 59–69.vozárová M. & sutorý k. (2001): Index herbariorum Reipublicae bohemicae et Reipublicae slovacae.
– Zpr. Čes. Bot. Společ. 36, suppl. 2001/1: 1–95.weber f. (1958): Die Tschechoslowakischen Thymus-Arten und Opiz´s Anteil an deren Erkennung. –
In: kláŠterský i. [ed.], Phillip Maxmilian Opiz und seine Bedeutung für die Pflanzentaxonomie, pp. 159–254, Praha.
wisskirchen r. & haeuPler h. (1998): Standardliste der Farn- und Blütenpflanzen Deutschlands. – Ulmer, Stuttgart.
zlatník a. & zvorykin i. (1932): Pokus o průzkum periodické proměny lesního a lučního stanoviště. – Sborn. Vys. Šk. Zeměd. Brno, Fak. lesn., sign. D19: 1–129.

247
Příroda, Praha, 31: 247–279, 2011
Kriticky ohrožené druhy rostlin České republiky: Jak se odlišují svými biologickými a ekologickými vlastnostmi od hojných druhů?
critically endangered plant species of the czech Republic: can we distinguish them from other plants using their biological and ecological traits?
JarMiLa gabrieLová1, ZuZana MünZbergová1, 2, iveta husáková1, 2 & Jindřich chrtek2
1Katedra botaniky, Přírodovědecká fakulta Univerzity Karlovy, Benátská 2, CZ-128 01 Praha 2; [email protected], [email protected], [email protected]ý ústav Akademie věd ČR, v. v. i., Zámek 1, CZ-252 43 Průhonice; [email protected]
Abstract: Understanding the factors responsible for species rarity is crucial for species effective conservation. One possible approach to obtain information about causes of species rarity is to compare rare and common species. We analysed biological and ecological traits of critically endangered (C1) plant species of the Czech Republic. We compared vegetative, generative and ecological traits of C1 species with three groups of species: (i) closely related common species (which allows phylogenetic correction), (ii) common closely related species sharing the same habitats (i. e. excluding pairs not sharing the same habitat as many differences in species traits can be caused by adaptation to a specific habitat type), and (iii) traits of all plants of the Czech Republic. Information about species traits was mainly obtained from literature and databases. The comparison with closely related common species has shown that C1 species are smaller, flower for shorter periods, and have higher proportions of self-compatibility and higher terminal velocities. C1 species differ also in the mode of dispersion, in their ecology, and distribution. The results based on comparing only species from the same habitat nearly equalled. The results of comparison with the entire flora differed slightly; we found additional differences in the pollination mode and seed mass. The results of all three types of comparison suggest that critically endangered species of the Czech Republic are small and competitively inferior species with some differences in the generative part of their life cycle that occur mainly in open unproductive habitats.Keywords: Central Europe, life-history traits, phylogenetically independent contrasts, rarity, Red List, threatened species
Abstrakt: Pochopení druhové vzácnosti a procesů, které ji podmiňují, je klíčové pro zachování vzácných a ohrožených druhů. Jednou z možných cest, jak získat odpověď na otázku po příčinách druhové vzácnosti, je porovnání vzácných a hojných druhů. Vyznačují se vzácné a ohrožené druhy nějakými specifickými vlastnostmi, které je odlišují od druhů hojných a mohou tak být příčinou jejich vzácnosti a ohrožení? Tuto otázku jsme se snažili zodpovědět v naší práci analýzou biologických a ekologických vlastností kriticky ohrožených druhů rostlin České republiky (kategorie C1 Červeného seznamu, Procházka 2001). Provedli jsme porovnání vegetativních, generativních a ekologických vlastností C1 druhů s vlastnostmi druhů (i) blízce příbuzných hojných (z důvodu fylogenetické korekce), (ii) blízce příbuzných hojných, rostoucích na stejném typu stanoviště (protože řada rozdílů v druhových charakteristikách může být způsobena adaptací na odlišný typ stanoviště) a zároveň (iii) s vlastnostmi druhů celé české květeny. Údaje o druzích byly získány z literatury a databází.

248
Gabrielová et al.: Kriticky ohrožené druhy rostlin, jak se odlišují svými vlastnostmi od hojných druhů?
Ukázalo se, že ve srovnání s druhy blízce příbuznými hojnými dosahují druhy C1 menší výšky, kvetou kratší dobu, je mezi nimi větší zastoupení autokompatibility a mají vyšší hodnoty terminal velocity (horší schopnost šířit se větrem). C1 druhy se rovněž odlišují způsobem disperze, řadou ekologických charakteristik a rozšířením (např. nižší zastoupení c-strategie, vyšší Ellenbergovy indikační hodnoty pro světlo, teplotu, půdní reakci a nižší hodnoty pro dusík, menší areály rozšíření). Výsledky porovnání párů druhů ze stejného typu stanoviště byly obdobné. Srovnání s druhy celé české flóry ukázalo další rozdíly ve způsobu opylení a hmotnosti semen. Pro většinu našich C1 druhů tedy platí, že se jedná o malé, kompetičně slabé druhy s jistými odlišnostmi v generativní fázi životního cyklu, které jsou vázány na otevřená, nezastíněná stanoviště s nízkou produktivitou.Klíčová slova: červený seznam, fylogeneticky nezávislé kontrasty, ohrožené druhy, střední Evropa, vzácnost, životní charakteristiky
Úvod
Porozumění faktorům určujících početnost a prostorové rozšíření organismů stále zůstává jedním z klíčových problémů ekologie (May 1999). Otázky, co podmiňuje relativní početnost druhů ve společenstvu, proč jsou některé druhy vzácné, zatímco jiné běžné a další dokonce invazní, zaměstnávají vědce už léta a doposud na ně nebyla nalezena uspokojivá odpověď. Přitom právě pochopení druhové vzácnosti a procesů, které ji podmiňují, je klíčové pro zachování vzácných a ohrožených druhů.
Jedna z možností, jak zodpovědět otázku příčin druhové vzácnosti, je pokusit se porovnat rozšíření a vlastnosti vzácných druhů s druhy blízce příbuznými hojnými (kunin & Gaston 1997). Existuje celá řada prací srovnávajících populační biologii dvou nebo několika málo blízce příbuzných druhů s podobnou ekologií, které se liší svou početností (např. fieDler 1987, byers & MeaGher 1997, witkowski & laMont 1997, walck et al. 2001, MünzberGová 2005). Čím podrobnější je analýza životního cyklu, tím větší je pravděpodobnost, že skutečně odhalí možné příčiny vzácnosti konkrétního druhu. Na druhou stranu studium populační biologie je velmi náročné a není prakticky možné ho provádět pro všechny vzácné a ohrožené druhy.
Jako alternativa se nabízí hledání vztahu mezi druhovou vzácností a vybranými biologickými charakteristikami, které jsou snadněji měřitelné a o kterých se předpokládá, že jsou korelovány s klíčovými aspekty životního cyklu rostlin (např. disperze, uchycení a vytrvání na stanovišti, weiher et al. 1999). V posledních letech vznikají rozsáhlé databáze životních charakteristik rostlin (např. thoMPson et al. 1997, bonn et al. 2000, klotz et al. 2002, PoschloD et al. 2003, kliMeŠová & kliMeŠ 2006, kleyer et al. 2008), které umožňují použití tohoto přístupu pro velký počet druhů.
Aby bylo možné určit, které vlastnosti jsou typické právě pro skupinu vzácných a ohrožených druhů (a nejsou společné pro většinu druhů naší květeny), je potřeba zjištěné údaje porovnat s nějakým referenčním souborem taxonů. Na prvním místě se nabízí soubor všech druhů české květeny. Takové srovnání je co do množství informací o druhových charakteristikách značně náročné, navíc není ze statistického hlediska zcela korektní. Při porovnání v rámci celé flóry nepředstavují taxony nezávislé body – vývojově příbuznější druhy jsou si svými vlastnostmi bližší než druhy ve fylogenezi vzdálené (freckelton et al. 2002). Výsledky takového srovnání pak poskytnou obecnou informaci o tom, v jakých vlastnostech se kriticky ohrožené druhy odlišují od zbytku květeny. Pokud však fylogenetickou nezávislost druhů zohledníme (např. porovnáním blízce příbuzných druhů, které se liší svou vzácností, cotGreave & PaGel 1997), umožní nám to odhalit konkrétní druhové vlastnosti, které mohou být příčinou vzácnosti a ohrožení. Pomocí

249
Příroda, Praha, 31: 247–279, 2011
fylogenetické korekce totiž v podstatě hledáme evoluční změny, které druh „nechtěně postavily před riziko vyhynutí“ (kunin & Gaston 1997). Metoda porovnání párů blízce příbuzných druhů není jediná možná. Ještě častěji se – zvláště v živočišné ekologii – uplatňuje metoda nezávislých kontrastů (felsenstein 1985). Ve chvíli, kdy nám pro řadu taxonů chybí informace o jejich postavení v rámci fylogenetického stromu a musíme se opírat zejména o taxonomii, je však párové porovnání vhodnější a jednodušší. Nevýhodou je, že se ne vždy podaří najít vhodný pár, čímž se značně snižuje počet stupňů volnosti pro statistické porovnání.
O to, jestli je opravdu nezbytně nutné aplikovat v případě srovnávacích studií fylogenetickou korekci, se stále vedou spory (např. westoby et al. 1995, freckelton et al. 2002). V případě rychle se vyvíjejících znaků a ve chvíli, kdy není k dispozici plně rozřešený fylogenetický strom, může tato metoda vést k nesprávným závěrům (freckelton et al. 2002). Často se proto používají obě metody (tedy fylogenetická korekce i obecné mezidruhové porovnání bez zohlednění příbuzenských vztahů mezi druhy), aby bylo možné výsledky obou typů srovnání porovnat.
„Vzácnost“ a „hojnost“ jsou pojmy relativní a v různých kontextech jsou pojímány odlišným způsobem. Asi nejjednodušší definice zní: „Vzácné druhy se vyznačují nízkou početností a/nebo malým rozsahem areálu“ (Gaston 1994). Pro praktickou aplikaci je ovšem tato definice poněkud vágní, protože chybí jasná kritéria pro stanovení nízké početnosti a malého rozsahu areálu pro konkrétní prostorovou škálu. Každá studie porovnávající charakteristiky životního cyklu vzácných a hojných druhů pojímá definici a kritéria vzácnosti trochu jinak a právě tato nejednotnost by mohla být jedním z důvodů, proč zatím souhrnné studie odhalily tak málo konzistentních rozdílů (pro přehled viz bevill & louDa 1999 a Murray et al. 2002). Murray et al. (2002), kteří shromáždili výsledky 54 studií porovnávajících dohromady celkem 94 charakteristik (z čehož ovšem pouze 30 vlastností bylo objektem zájmu pro více než jednu studii), odhalili pouze jeden konzistentní rozdíl, a to v produkci semen, přičemž vzácné druhy produkovaly ve většině případů méně semen než druhy hojné.
V našem příspěvku se zaměřujeme na kriticky ohrožené druhy1 (kategorie C1 Červeného seznamu cévnatých rostlin České republiky, Procházka 2001). Tato skupina představuje relativně heterogenní směs taxonů, vybraných na základě různých, často subjektivních kritérií. Najdeme zde druhy přirozeně vzácné (např. vázané na speciální stanoviště, která v České republice nejsou plošně příliš zastoupená), druhy ubývající v důsledku negativních dopadů lidské činnosti, dále taxony, jež jsou vzácné a jejichž početnost kolísá v důsledku toho, že jsou na okraji areálu svého rozšíření, a samozřejmě druhy, jež jsou vzácné či ubývají z jiných (často neznámých) příčin. Skupinu kriticky ohrožených druhů jsme zvolili proto, že se jedná o předem definovaný soubor druhů obecně považovaných za vzácné v rámci České republiky, navíc prioritní z hlediska ochrany přírody.
Porovnáním vlastností C1 druhů s vlastnostmi druhů blízce příbuzných, ale co do rozšíření hojných, se snažíme odhalit možné příčiny vzácnosti a ohrožení C1 druhů. Abychom odfiltrovali problém, že řada odlišností v druhových charakteristikách blízce příbuzných druhů může být způsobena adaptací k životu na různém typu stanoviště, provádíme rovněž porovnání podsouboru párů blízce příbuzných druhů rostoucích na stejném typu stanoviště. Protože nás zajímá i otázka, jestli existují nějaké vlastnosti, které by skupinu kriticky ohrožených druhů odlišovaly od zbytku české květeny, provádíme pro vybrané charakteristiky rovněž porovnání C1 druhů s druhy celé české flóry. Z těchto
1) V řadě případů se jedná i o taxonomicky nižší jednotky (subspecie), takže by bylo správné použí-vat termín C1 taxony, pro lepší čitelnost však častěji používáme termín C1 druhy.

250
Gabrielová et al.: Kriticky ohrožené druhy rostlin, jak se odlišují svými vlastnostmi od hojných druhů?
východisek vyplývají následující otázky: (i) Jaké vlastnosti charakterizují kriticky ohrožené (C1) druhy naší květeny? (ii) Jak se vlastnosti C1 druhů odlišují od vlastností druhů blízce příbuzných hojných? (iii) Jak se vlastnosti C1 druhů liší od blízce příbuzných hojných druhů, které rostou na stejném typu stanoviště? (iv) Jak se vlastnosti C1 druhů odlišují od vlastností druhů celé české květeny?
Metodika
výběr blízce příbuzných hojných druhů
Při výběru druhů blízce příbuzných druhům C1, ale co do rozšíření hojných, jsme vycházeli z publikovaných fylogenetických rekonstrukcí příslušných skupin (většinou šlo o rod), díky kterým se stanovil nejblíže příbuzný druh ze stejného rodu, rostoucí na našem území. Pokud taková studie nebyla k dispozici (zhruba v 60 % případů), byla zohledněna v poslední době akceptovaná vnitrodruhová členění, založená většinou na jiných než molekulárních znacích (např. morfologie, počet chromozomů, znaky pylových zrn, sekundární metabolity; J. Chrtek, nepubl. data). Původně měl pojem „hojný druh“ označovat taxon ze stejného rodu, který není uveden na Černém a červeném seznamu cévnatých rostlin ČR (Procházka 2001). Ukázalo se však, že vhodných párů za takové podmínky není dostatek, proto byly nakonec do souboru hojných druhů zahrnuty i druhy kategorie C3 – ohrožené (51 druhů) a C4 – vzácnější taxony vyžadující další pozornost (29 druhů). Celkem bylo takto vytvořeno 206 vhodných párů (příloha 1). Zbylých 277 druhů C1 nemá na našem území hojného blízkého příbuzného z následujících důvodů: (i) blízký příbuzný je sám kriticky či silně ohrožený, případně vyhynulý, (ii) jedná se o monotypický rod, (iii) žádný blízce příbuzný z daného rodu na našem území neroste, (iv) v rámci rodu se jedná o samostatný podrod či sekci, (v) jedná se o skupiny apomiktických taxonů.
sběr dat o vlastnostech druhů
Informace o vlastnostech druhů jsme získali z literatury (ellenberG et al. 1992, kubát et al. 2002, hejný & slavík 1988, 1990, 1992, slavík 1995, 1997, 2000, slavík & ŠtěPánková 2004, ŠtěPánková et al. 2010) a dostupných databází (thoMPson et al. 1997, bonn et al. 2000, klotz et al. 2002, PoschloD et al. 2003, kliMeŠová & kliMeŠ 2006, kleyer et al. 2008). Charakteristiky lze rozdělit do pěti hlavních okruhů (podrobný popis viz příloha 2):• vegetativní znaky: životní forma, délka života, průměrná výška rostliny [m], listová
růžice, vytrvalost listů, anatomie listů, specifická listová plocha (SLA) [mm2/mg], klonalita;
• generativní znaky: období kvetení, typ reprodukce, způsob opylení, inkompatibilita, diklinie (pohlavnost květů), dichogamie (časově nestejné dozrávání prašníků a blizen), odměna za opylení;
• vlastnosti diaspor: hmotnost semen [mg], rozměry semen [mm], tvar semen, terminal velocity (maximální rychlost, které semeno dosáhne při pádu za bezvětří) [m/s], způsob šíření semen, produkce semen, přežívání v půdní bance semen;
• ekologické charakteristiky: ekologická strategie csr (GriMe 1979), urbanita (vyjadřuje afinitu rostlin k urbánním oblastem), hemerobie (měřítko toho, jak přirozená stanoviště rostlina obývá), Ellenbergovy indikační hodnoty;
• rozšíření: obecná informace o stanovišti, výškový stupeň, velikost areálu rozšíření.Hodnoty vlastností všech C1 druhů a jejich podrobný popis jsou dostupné v on-line Da-
tabázi C1 rostlin v rámci datového skladu AOPK.

251
Příroda, Praha, 31: 247–279, 2011
Vybrané druhové vlastnosti (tab. 1) jsme rovněž porovnávali mezi C1 druhy a druhy celé české květeny. Vycházeli jsme přitom z checklistu české flóry (Danihelka et al. 2007), ze kterého jsme vyloučili C1 druhy, vyhynulé druhy a druhy nejasného taxonomického postavení. Získali jsme tak seznam 3 263 druhů a poddruhů.
Je zřejmé, že porovnávané vlastnosti jsou ze své podstaty nesouměřitelné – zejména některé ekologické charakteristiky, které zde prezentujeme jako jednotlivé „vlastnosti“ (ekologická strategie, hemerobie, urbanita), představují komplexní kombinaci celé řady různých faktorů. To je třeba zohlednit při interpretaci výsledků.
vyhodnocení dat
Rozdíly v kvantitativních charakteristikách (např. Ellenbergovy indikační hodnoty, průměrná výška, doba kvetení, hmotnost a rozměry semen) jsme testovali párovým t-testem, případně jeho neparametrickou obdobou – Wilcoxonovým znaménkovým testem (porovnání C1 druhy vs. blízce příbuzné hojné) a jednocestnou analýzou variance nebo Kruskal-Wallisovým testem (porovnání C1 druhy vs. celá flóra). Pro testování rozdílů v kategoriálních vlastnostech (např. vytrvalost listů, typ reprodukce, způsob produkce semen) jsme použili kontingenční tabulku a χ² test. Rozdíly mezi druhy v kategoriálních charakteristikách, které jsou kódovány procentuálně (protože jeden druh se může vyskytovat ve více kategoriích – např. výškový stupeň, způsob opylení), jsme testovali pomocí unimodální přímé mnohorozměrné analýzy CCA (s druhovými vlastnostmi jako „species“ a jednotlivými druhy jako „samples“). Při srovnání s druhy blízce příbuznými hojnými byly páry druhů odečteny jako kovariáty. Statistické analýzy byly provedeny v programu S-PLUS (2000) a CANOCO (ter braak & ŠMilauer 1998).
Abychom odfiltrovali problém, že řada odlišností v druhových charakteristikách blízce příbuzných druhů může být způsobena adaptací k životu na různém typu stanoviště, vyhledali jsme pro druhy C1 a jejich hojné protějšky informaci o stanovišti a vybrali páry druhů C1 a jim blízce příbuzných hojných druhů, které se spolu mohou vyskytovat na stejném typu stanoviště. Výskyt druhů byl procenticky rozkódován do devíti základních typů biotopů (chytrý et al. 2001): vodní toky a nádrže; mokřady a pobřežní vegetace; prameniště a rašeliniště; skály, sutě a jeskyně; alpinské bezlesí; sekundární trávníky a vřesoviště; křoviny; lesy; biotopy silně ovlivněné nebo vytvořené člověkem. Rozdělení jsme provedli na základě zařazení druhů do příslušných fytocenologických jednotek a biotopů podle Moravec et al. (1995) a chytrý et al. (2001). Řada druhů nebyla v těchto publikacích zmíněna, v tom případě jsme na základě převodních tabulek z Katalogu biotopů (chytrý et al. 2001) a údajů o stanovišti z Květeny ČR (hejný & slavík 1988, 1990, 1992, slavík 1995, 1997, 2000, slavík & ŠtěPánková 2004, ŠtěPánková 2010) a Klíče ke Květeně ČR (kubát et al. 2002) zařadili druhy do příslušného biotopu příp. jejich kombinace. Následně jsme vybrali druhy C1 a druhy jim blízce příbuzné hojné, které se vyskytují alespoň částečně ve stejném typu biotopu. V několika případech se druhy mohly společně potkat ve více typech biotopů, pak byl vybrán biotop s větší pravděpodobností společného výskytu. Tímto způsobem jsme dospěli ke 149 párům druhů a pro tento podsoubor jsme provedli stejné statistické testování rozdílů v druhových vlastnostech jako pro všechny páry druhů C1 a druhů jim blízce příbuzných hojných.
Protože provádíme velké množství oddělených porovnání vlastností, které jsou navzájem korelované, vytvořili jsme korelační matici znaků, ve kterých jsme našli sta tis-ticky významné korelace. Korelační matici zde neprezentujeme, protože je příliš rozsáhlá; průkazné korelace se navíc projevily mezi vlastnostmi, u kterých se dá vzájemný vztah předpokládat – především korelace mezi charakteristikami souvisejícími s ekologií a roz-

252
Gabrielová et al.: Kriticky ohrožené druhy rostlin, jak se odlišují svými vlastnostmi od hojných druhů?
šířením (např. mezi Ellenbergovými indikačními hodnotami a hemerobií, urbanitou, typem stanoviště a nadmořskou výškou; mezi csr-strategií a hemerobií apod.). Objevilo se jen málo korelací mezi biologickými vlastnostmi a ekologií (např. mezi terminal velocity a anemochorií, průměrnou výškou rostliny a Ellenbergovými indikačními hodnotami pro půdní reakci a mezi anemogamií a urbanofilií).
V souvislosti s porovnáním C1 druhů s druhy celé české květeny nás zajímala ještě otázka, jestli C1 druhy nejsou v některých čeledích zastoupeny nepoměrně více nebo naopak méně než by odpovídalo náhodě (což by pak mohlo být důvodem případných zjištěných rozdílů mezi C1 druhy a celou českou květenou). Tento problém jsme řešili pomocí randomizačního testu, ve kterém jsme se ptali, zda počet C1 druhů v dané čeledi může být výsledkem náhodného výběru z celého souboru druhů (tj. ze všech druhů české květeny). Konkrétně jsme tedy opakovaně náhodně vybírali 483 druhů ze všech druhů české květeny a v každém kroku jsme zaznamenali, jestli je počet druhů v jednotlivých čeledích tohoto náhodného výběru stejný, vyšší nebo nižší než počet C1 druhů v daných čeledích. Toto náhodné vybírání jsme zopakovali tisíckrát. Pro každou čeleď jsme odvodili empirickou P-hodnotu, která udává pravděpodobnost, se kterou může být počet C1 druhů v jednotlivých čeledích výsledkem náhody a kdy je naopak počet C1 druhů v dané čeledi vyšší nebo nižší, než by odpovídalo náhodě (oboustranný test, α = 0,05).
výsledky
srovnání vlastností c1 druhů s druhy blízce příbuznými hojnými
Ze souboru vegetativních znaků se studované druhy průkazně liší průměrnou výškou – C1 druhy jsou menšího vzrůstu (obr. 1a). Druhy se neliší životní formou, délkou života, distribucí listů na stonku, vytrvalostí listů, listovou anatomií, SLA ani klonalitou (tab. 1).
tab. 1. Rozdíly ve vlastnostech kriticky ohrožených druhů ČR (C1) a druhů blízce příbuzných hojných, blízce příbuzných hojných druhů ze stejného typu habitatu a druhů celé květeny ČR. *** p ≤ 0,001, ** p < 0,01, * p < 0,05, m.s. okrajově signifikantní (p < 0,1), N.S. nesignifikantní; + C1 druhy mají vyšší hodnoty pro daný znak; – C1 druhy mají nižší hodnoty; bez znaménka – signifikantní rozdíly pro faktory; prázdná pole – rozdíly ve vlastnostech netestovány. Test: CCA – unimodální mnohorozměrná přímá analýza, t – párový t-test/jednocestná ANOVA, W – Wilcoxonův znaménkový test/ Kruskal-Wallisův test, χ² – kontingenční tabulka a χ² test. LDD vs. SDD – disperze diaspor na dlouhé vzdálenosti vs. disperze na krátké vzdálenosti.*Klonalita: velké množství znaků excerpovaných z kliMeŠová & kliMeŠ (2006), všechny rozdíly nesignifikantní, proto neukázáno.table 1. Differences in species traits between critically endangered (C1) and common species, common species from the same habitat type and the entire flora of the Czech Republic. *** p ≤ 0.001, ** p < 0.01, * p < 0.05, m.s. marginally significant (p < 0.1), N.S. not significant; + C1 species have higher values of given trait; – C1 species have lower values; without sign – significant differences for factors; empty fields – not tested for differences. Test: CCA – canonical correspondence analysis, t – paired t-test/one-way ANOVA, W – Wilcoxon signed-rank test/ Kruskal-Wallis test, χ² – contingency table with χ² test. LDD vs. SDD – long vs. short distance dispersal.*Clonality: many traits from kliMeŠová & kliMeŠ (2006), all differences not significant, thereby not shown.
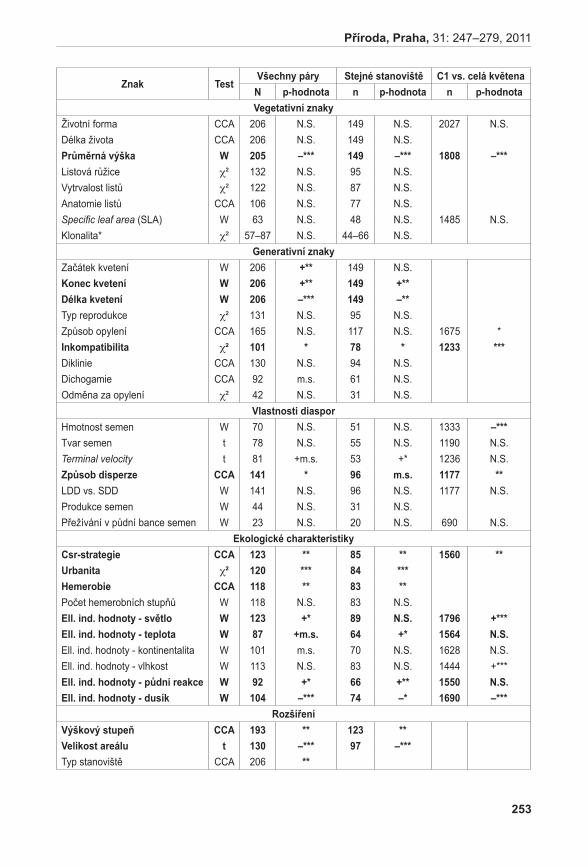
253
Příroda, Praha, 31: 247–279, 2011
Znak testvšechny páry stejné stanoviště c1 vs. celá květena
n p-hodnota n p-hodnota n p-hodnotavegetativní znaky
Životní forma CCA 206 N.S. 149 N.S. 2027 N.S.Délka života CCA 206 N.S. 149 N.S. Průměrná výška W 205 –*** 149 –*** 1808 –***Listová růžice χ² 132 N.S. 95 N.S. Vytrvalost listů χ² 122 N.S. 87 N.S. Anatomie listů CCA 106 N.S. 77 N.S. Specific leaf area (SLA) W 63 N.S. 48 N.S. 1485 N.S.Klonalita* χ² 57–87 N.S. 44–66 N.S.
Generativní znakyZačátek kvetení W 206 +** 149 N.S. Konec kvetení W 206 +** 149 +** délka kvetení W 206 –*** 149 –** Typ reprodukce χ² 131 N.S. 95 N.S. Způsob opylení CCA 165 N.S. 117 N.S. 1675 *inkompatibilita χ² 101 * 78 * 1233 ***Diklinie CCA 130 N.S. 94 N.S. Dichogamie CCA 92 m.s. 61 N.S. Odměna za opylení χ² 42 N.S. 31 N.S.
vlastnosti diasporHmotnost semen W 70 N.S. 51 N.S. 1333 –***Tvar semen t 78 N.S. 55 N.S. 1190 N.S.Terminal velocity t 81 +m.s. 53 +* 1236 N.S.Způsob disperze ccA 141 * 96 m.s. 1177 **LDD vs. SDD W 141 N.S. 96 N.S. 1177 N.S.Produkce semen W 44 N.S. 31 N.S. Přežívání v půdní bance semen W 23 N.S. 20 N.S. 690 N.S.
Ekologické charakteristikycsr-strategie ccA 123 ** 85 ** 1560 **urbanita χ² 120 *** 84 *** Hemerobie ccA 118 ** 83 ** Počet hemerobních stupňů W 118 N.S. 83 N.S. Ell. ind. hodnoty - světlo W 123 +* 89 n.s. 1796 +***Ell. ind. hodnoty - teplota W 87 +m.s. 64 +* 1564 n.s.Ell. ind. hodnoty - kontinentalita W 101 m.s. 70 N.S. 1628 N.S.Ell. ind. hodnoty - vlhkost W 113 N.S. 83 N.S. 1444 +***Ell. ind. hodnoty - půdní reakce W 92 +* 66 +** 1550 n.s.Ell. ind. hodnoty - dusík W 104 –*** 74 –* 1690 –***
Rozšířenívýškový stupeň ccA 193 ** 123 ** velikost areálu t 130 –*** 97 –*** Typ stanoviště CCA 206 **
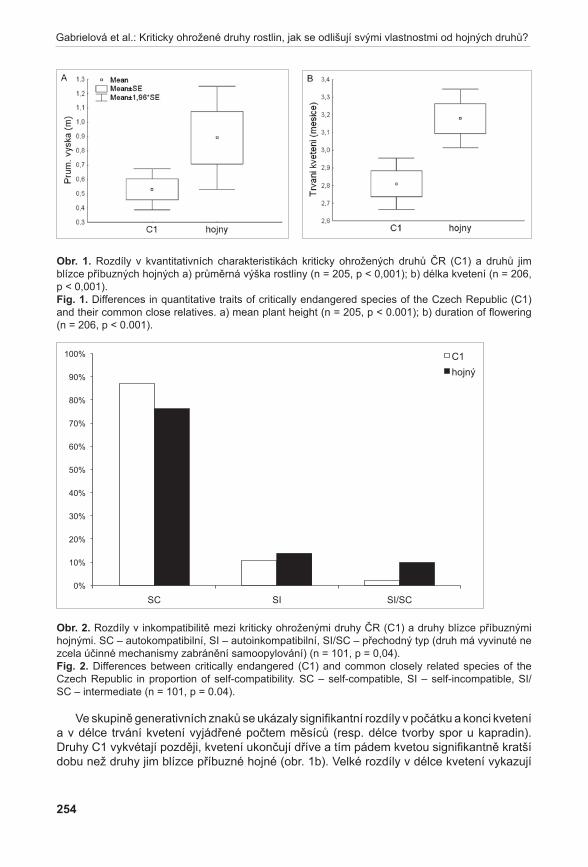
254
Gabrielová et al.: Kriticky ohrožené druhy rostlin, jak se odlišují svými vlastnostmi od hojných druhů?
obr. 1. Rozdíly v kvantitativních charakteristikách kriticky ohrožených druhů ČR (C1) a druhů jim blízce příbuzných hojných a) průměrná výška rostliny (n = 205, p < 0,001); b) délka kvetení (n = 206, p < 0,001).Fig. 1. Differences in quantitative traits of critically endangered species of the Czech Republic (C1) and their common close relatives. a) mean plant height (n = 205, p < 0.001); b) duration of flowering (n = 206, p < 0.001).
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
SC SI SI/SC
C1 hojn
obr. 2. Rozdíly v inkompatibilitě mezi kriticky ohroženými druhy ČR (C1) a druhy blízce příbuznými hojnými. SC – autokompatibilní, SI – autoinkompatibilní, SI/SC – přechodný typ (druh má vyvinuté ne zcela účinné mechanismy zabránění samoopylování) (n = 101, p = 0,04).Fig. 2. Differences between critically endangered (C1) and common closely related species of the Czech Republic in proportion of self-compatibility. SC – self-compatible, SI – self-incompatible, SI/SC – intermediate (n = 101, p = 0.04).
Ve skupině generativních znaků se ukázaly signifikantní rozdíly v počátku a konci kvetení a v délce trvání kvetení vyjádřené počtem měsíců (resp. délce tvorby spor u kapradin). Druhy C1 vykvétají později, kvetení ukončují dříve a tím pádem kvetou signifikantně kratší dobu než druhy jim blízce příbuzné hojné (obr. 1b). Velké rozdíly v délce kvetení vykazují
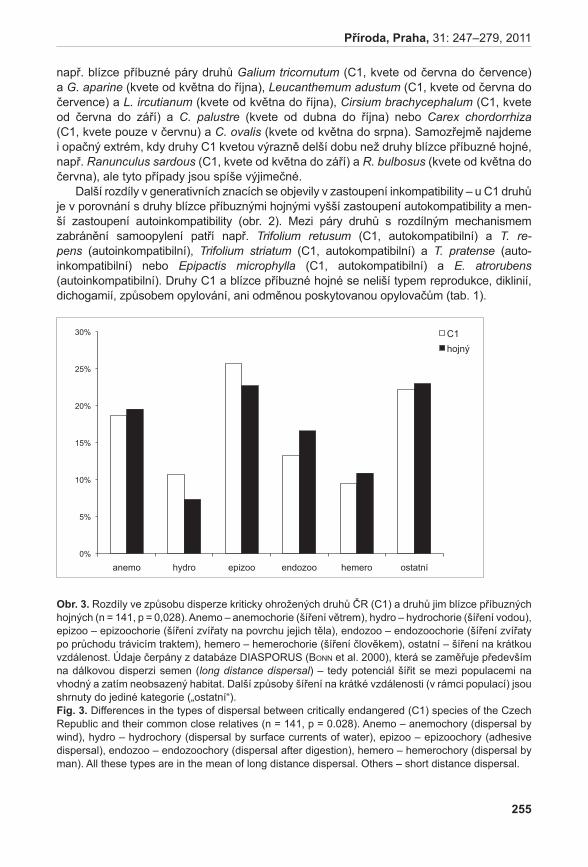
255
Příroda, Praha, 31: 247–279, 2011
např. blízce příbuzné páry druhů Galium tricornutum (C1, kvete od června do července) a G. aparine (kvete od května do října), Leucanthemum adustum (C1, kvete od června do července) a L. ircutianum (kvete od května do října), Cirsium brachycephalum (C1, kvete od června do září) a C. palustre (kvete od dubna do října) nebo Carex chordorrhiza (C1, kvete pouze v červnu) a C. ovalis (kvete od května do srpna). Samozřejmě najdeme i opačný extrém, kdy druhy C1 kvetou výrazně delší dobu než druhy blízce příbuzné hojné, např. Ranunculus sardous (C1, kvete od května do září) a R. bulbosus (kvete od května do června), ale tyto případy jsou spíše výjimečné.
Další rozdíly v generativních znacích se objevily v zastoupení inkompatibility – u C1 druhů je v porovnání s druhy blízce příbuznými hojnými vyšší zastoupení autokompatibility a men-ší zastoupení autoinkompatibility (obr. 2). Mezi páry druhů s rozdílným mechanismem zabránění samoopylení patří např. Trifolium retusum (C1, autokompatibilní) a T. re-pens (autoinkompatibilní), Trifolium striatum (C1, autokompatibilní) a T. pratense (auto-inkompatibilní) nebo Epipactis microphylla (C1, autokompatibilní) a E. atrorubens (autoinkompatibilní). Druhy C1 a blízce příbuzné hojné se neliší typem reprodukce, diklinií, dichogamií, způsobem opylování, ani odměnou poskytovanou opylovačům (tab. 1).
0%
5%
10%
15%
20%
25%
30%
anemo hydro epizoo endozoo hemero ostatní
C1 hojn
obr. 3. Rozdíly ve způsobu disperze kriticky ohrožených druhů ČR (C1) a druhů jim blízce příbuzných hojných (n = 141, p = 0,028). Anemo – anemochorie (šíření větrem), hydro – hydrochorie (šíření vodou), epizoo – epizoochorie (šíření zvířaty na povrchu jejich těla), endozoo – endozoochorie (šíření zvířaty po průchodu trávicím traktem), hemero – hemerochorie (šíření člověkem), ostatní – šíření na krátkou vzdálenost. Údaje čerpány z databáze DIASPORUS (bonn et al. 2000), která se zaměřuje především na dálkovou disperzi semen (long distance dispersal) – tedy potenciál šířit se mezi populacemi na vhodný a zatím neobsazený habitat. Další způsoby šíření na krátké vzdálenosti (v rámci populací) jsou shrnuty do jediné kategorie („ostatní“).Fig. 3. Differences in the types of dispersal between critically endangered (C1) species of the Czech Republic and their common close relatives (n = 141, p = 0.028). Anemo – anemochory (dispersal by wind), hydro – hydrochory (dispersal by surface currents of water), epizoo – epizoochory (adhesive dispersal), endozoo – endozoochory (dispersal after digestion), hemero – hemerochory (dispersal by man). All these types are in the mean of long distance dispersal. Others – short distance dispersal.

256
Gabrielová et al.: Kriticky ohrožené druhy rostlin, jak se odlišují svými vlastnostmi od hojných druhů?
obr. 4. Rozdíly v Ellenbergových indikačních hodnotách mezi kri-ticky ohroženými druhy ČR (C1) a druhy jim blízce příbuznými hoj-nými.a) Ellenbergovy indikační hodnoty pro světlo (n = 123, p = 0,01); b) Ellenbergovy indikační hodnoty pro živiny (n = 104, p < 0,001). c) Rozdíly ve velikosti evropské části areálu rozšíření (n = 130, p < 0,001).Fig. 4. Differences in Ellenberg in di cator values between critically endangered (C1) species of the Czech Republic and their common close relatives.a) Ell. ind. values for light (n = 123, p = 0.01); b) Ell. ind. values for nitrogen (n = 104, p < 0.001). c) Differences in the distribution range (n = 130, p < 0.001).
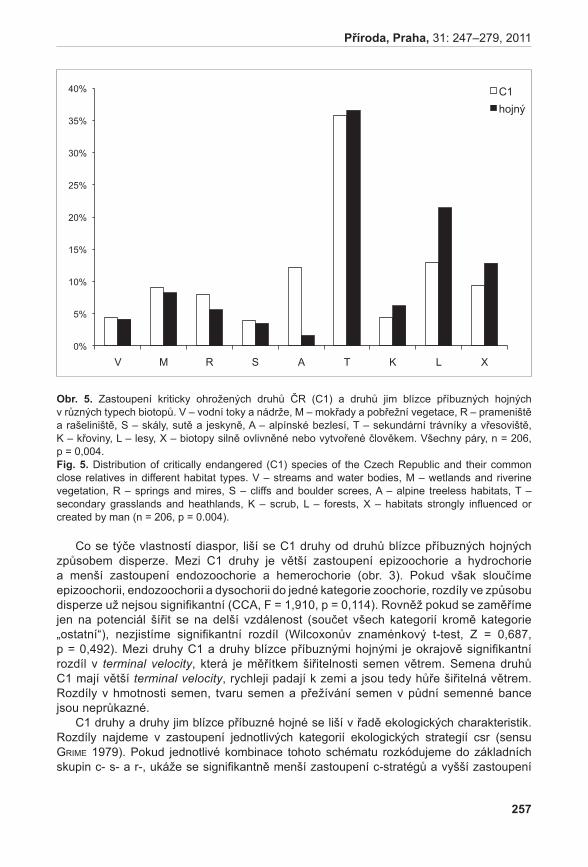
257
Příroda, Praha, 31: 247–279, 2011
0%
5%
10%
15%
20%
25%
30%
35%
40%
V M R S A T K L X
C1 hojn
obr. 5. Zastoupení kriticky ohrožených druhů ČR (C1) a druhů jim blízce příbuzných hojných v různých typech biotopů. V – vodní toky a nádrže, M – mokřady a pobřežní vegetace, R – prameniště a rašeliniště, S – skály, sutě a jeskyně, A – alpínské bezlesí, T – sekundární trávníky a vřesoviště, K – křoviny, L – lesy, X – biotopy silně ovlivněné nebo vytvořené člověkem. Všechny páry, n = 206, p = 0,004.Fig. 5. Distribution of critically endangered (C1) species of the Czech Republic and their common close relatives in different habitat types. V – streams and water bodies, M – wetlands and riverine vegetation, R – springs and mires, S – cliffs and boulder screes, A – alpine treeless habitats, T – secondary grasslands and heathlands, K – scrub, L – forests, X – habitats strongly influenced or created by man (n = 206, p = 0.004).
Co se týče vlastností diaspor, liší se C1 druhy od druhů blízce příbuzných hojných způsobem disperze. Mezi C1 druhy je větší zastoupení epizoochorie a hydrochorie a menší zastoupení endozoochorie a hemerochorie (obr. 3). Pokud však sloučíme epizoochorii, endozoochorii a dysochorii do jedné kategorie zoochorie, rozdíly ve způsobu disperze už nejsou signifikantní (CCA, F = 1,910, p = 0,114). Rovněž pokud se zaměříme jen na potenciál šířit se na delší vzdálenost (součet všech kategorií kromě kategorie „ostatní“), nezjistíme signifikantní rozdíl (Wilcoxonův znaménkový t-test, Z = 0,687, p = 0,492). Mezi druhy C1 a druhy blízce příbuznými hojnými je okrajově signifikantní rozdíl v terminal velocity, která je měřítkem šiřitelnosti semen větrem. Semena druhů C1 mají větší terminal velocity, rychleji padají k zemi a jsou tedy hůře šiřitelná větrem. Rozdíly v hmotnosti semen, tvaru semen a přežívání semen v půdní semenné bance jsou neprůkazné.
C1 druhy a druhy jim blízce příbuzné hojné se liší v řadě ekologických charakteristik. Rozdíly najdeme v zastoupení jednotlivých kategorií ekologických strategií csr (sensu GriMe 1979). Pokud jednotlivé kombinace tohoto schématu rozkódujeme do základních skupin c- s- a r-, ukáže se signifikantně menší zastoupení c-stratégů a vyšší zastoupení
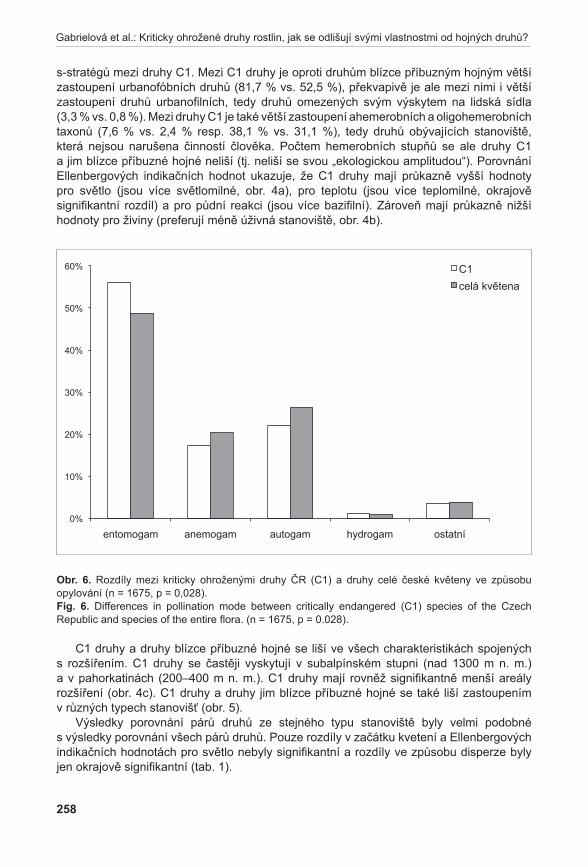
258
Gabrielová et al.: Kriticky ohrožené druhy rostlin, jak se odlišují svými vlastnostmi od hojných druhů?
s-stratégů mezi druhy C1. Mezi C1 druhy je oproti druhům blízce příbuzným hojným větší zastoupení urbanofóbních druhů (81,7 % vs. 52,5 %), překvapivě je ale mezi nimi i větší zastoupení druhů urbanofilních, tedy druhů omezených svým výskytem na lidská sídla (3,3 % vs. 0,8 %). Mezi druhy C1 je také větší zastoupení ahemerobních a oligohemerobních taxonů (7,6 % vs. 2,4 % resp. 38,1 % vs. 31,1 %), tedy druhů obývajících stanoviště, která nejsou narušena činností člověka. Počtem hemerobních stupňů se ale druhy C1 a jim blízce příbuzné hojné neliší (tj. neliší se svou „ekologickou amplitudou“). Porovnání Ellenbergových indikačních hodnot ukazuje, že C1 druhy mají průkazně vyšší hodnoty pro světlo (jsou více světlomilné, obr. 4a), pro teplotu (jsou více teplomilné, okrajově signifikantní rozdíl) a pro půdní reakci (jsou více bazifilní). Zároveň mají průkazně nižší hodnoty pro živiny (preferují méně úživná stanoviště, obr. 4b).
0%
10%
20%
30%
40%
50%
60%
entomogam anemogam autogam hydrogam ostatní
C1 celá kv tena
obr. 6. Rozdíly mezi kriticky ohroženými druhy ČR (C1) a druhy celé české květeny ve způsobu opylování (n = 1675, p = 0,028).Fig. 6. Differences in pollination mode between critically endangered (C1) species of the Czech Republic and species of the entire flora. (n = 1675, p = 0.028).
C1 druhy a druhy blízce příbuzné hojné se liší ve všech charakteristikách spojených s rozšířením. C1 druhy se častěji vyskytují v subalpínském stupni (nad 1300 m n. m.) a v pahorkatinách (200–400 m n. m.). C1 druhy mají rovněž signifikantně menší areály rozšíření (obr. 4c). C1 druhy a druhy jim blízce příbuzné hojné se také liší zastoupením v různých typech stanovišť (obr. 5).
Výsledky porovnání párů druhů ze stejného typu stanoviště byly velmi podobné s výsledky porovnání všech párů druhů. Pouze rozdíly v začátku kvetení a Ellenbergových indikačních hodnotách pro světlo nebyly signifikantní a rozdíly ve způsobu disperze byly jen okrajově signifikantní (tab. 1).

259
Příroda, Praha, 31: 247–279, 2011
Porovnání vlastností druhů c1 a druhů celé české květeny
Porovnání vybraných vlastností druhů C1 a druhů celé české květeny potvrdilo některé již zjištěné skutečnosti (rozdíly v průměrné výšce rostlin, v inkompatibilitě, csr-strategii, Ellenberových indikačních hodnotách pro světlo a živiny). Nově se ukázaly rozdíly ve způsobu opylování (obr. 6). Rozdíly ve způsobu disperze zůstaly průkazné a stejně jako při porovnání s druhy blízce příbuznými hojnými převládá u C1 druhů epizoochorie a hydrochorie. Zatímco však srovnání s druhy blízce příbuznými hojnými ukázalo nepatrně menší zastoupení anemochorie u druhů C1 (18,7 % oproti 19,5 %), při porovnání s druhy celé české květeny bylo zastoupení anemochorie mezi druhy C1 mnohem větší (21,5 % oproti 12,1 %). C1 druhy mají zároveň ve srovnání s druhy celé české květeny signifikantně lehčí semena. Nově se také ukázal rozdíl v Ellenbergových indikačních hodnotách pro vlhkost – druhy C1 je mají vyšší a jsou tedy více vlhkomilné. Rozdíly v hodnotách pro teplotu a půdní reakci byly naopak při tomto porovnání neprůkazné.
C1 druhy jsou zastoupeny v 83 čeledích ze 173 čeledí, které se vyskytují v ČR. V pěti čeledích je zastoupení C1 druhů signifikantně nižší, než by odpovídalo náhodnému samplování z celé české květeny (Alliaceae, Brassicaceae, Onagraceae, Polygonaceae, Rosaceae a Solanaceae). V devíti čeledích je naopak zastoupení C1 druhů signifikantně vyšší, než by odpovídalo náhodě (Aspleniaceae, Asteraceae, Caryophylaceae, Gentianaceae, Isoëtaceae, Lentibulariaceae, Orchidaceae, Orobanchaceae a Santalaceae).
diskuze
Výsledky porovnání vlastností kriticky ohrožených a hojných druhů ukázaly řadu rozdílů v jejich ekologii a rozšíření a jen relativně málo rozdílů v jejich biologických charakteristikách. Z vlastností souvisejících s biologií jsme objevili rozdíly zejména v generativních znacích – C1 druhy začínají kvést později, kvetení ukončují dříve a kvetou tedy kratší dobu než druhy blízce příbuzné hojné. K podobnému zjištění dospěli i severští autoři, zabývající se stejnou problematikou (lahti et al. 1991, Gustafsson 1994). Pokud s délkou trvání kvetení souvisí efektivita a úspěšnost návštěv opylovačů, pak tento fakt vzácné a ohrožené druhy oproti jejich hojnějším protějškům může znevýhodňovat. Na druhou stranu, jak zmiňují lahti et al. (1991) a jak se ukazuje i při pohledu na příklady druhových párů, může být tento jev způsoben metodologickým problémem: málo početné populace a omezený areál vzácných a ohrožených druhů mohou snižovat varianci fenologie kvetení pozorovanou v terénu a pak uváděnou v databázích. Navíc se mezi kriticky ohroženými taxony vyskytuje řada horských druhů, které vzhledem ke klimatickým podmínkám, které panují ve vysokých nadmořských výškách, zákonitě vykvétají později a kvetení ukončují dříve než druhy z nížin.
Zjištěná kratší doba kvetení C1 druhů koresponduje i s další odlišností C1 druhů v reprodukční biologii – vyšší mírou autokompatibility těchto druhů. Vzácné a ohrožené druhy, které tvoří malé a izolované populace, mohou být limitovány ve svém reprodukčním úspěchu při procesu opylení, což se zvláště týká druhů autoinkompatibilních. Opylovači-generalisté efektivně opylují početné populace rostlin, ale při návštěvě druhů s malými a fragmentovanými populacemi přinášejí velké množství nevhodného pylu (kunin & schMiDa 1997). Za této situace je pro rostlinu výhodné vyvinout zvláštní vlastnosti, které by zvýšily kvalitu a kvantitu návštěv opylovačů (nápadná květní morfologie, déle vytrvávající květy), nebo naopak vsadit na vlastnost, která závislost na opylovačích snižuje (autokompatibilita). Pro autokompatibilní rostliny pak už nemá investice do mechanismů zvyšujících pravděpodobnost návštěvy opylovačů takový význam, takže se u nich dá očekávat spíše kratší doba kvetení a méně nápadná květní morfologie (kunin & schMiDa 1997). Zdá se, že významná část C1 druhů

260
Gabrielová et al.: Kriticky ohrožené druhy rostlin, jak se odlišují svými vlastnostmi od hojných druhů?
zvolila právě cestu autokompatibility a zmenšení nákladů na kvetení. Na druhou stranu, dlouhodobé samoopylování blízce příbuzných jedinců v rámci populací může mít negativní vliv na fitness těchto populací v důsledku inbrední deprese (Meffe & carroll 1994).
Srovnání s druhy celé české květeny ukázalo, že mezi druhy C1 je signifikantně větší zastoupení entomogamie a menší zastoupení anemogamie a autogamie. Výsledek koresponduje se závěry dalších studií (kelly & wooDwarD 1996, caDotte & lovett-Doust 2002, PilGriM et al. 2004), které ukazují, že entomogamní druhy mají menší areály rozšíření nebo jsou více ohrožené než druhy anemogamní. Tento jev se označuje jako „náklady za mutualismus“, kdy vazba na zvláštního opylovače omezuje rozsah území, které může druh kolonizovat. Zároveň to znamená větší citlivost entomogamních druhů vůči změnám prostředí a degradaci stanovišť: přizpůsobit se musí nejenom rostlinný druh, ale i jeho mutualistický opylovač (kelly 1996). Fakt, že se při analýze vlastností druhů C1 ukázal signifikantní rozdíl až při porovnání s druhy celé české květeny, by se dal vysvětlit tím, že způsob opylování je zřejmě znak evolučně stabilní, takže se mezi blízce příbuznými druhy neliší.
Zjištěné rozdíly ve vlastnostech diaspor nejsou v rámci obou typů srovnání – druhy C1 vs. blízce příbuzné hojné a druhy C1 vs. celá česká květena – příliš konzistentní. Výsledky porovnání s druhy blízce příbuznými hojnými naznačují, že druhy C1 by mohly být limitovány v procesu šíření (vyšší hodnoty terminal velocity), ovšem při srovnání s druhy celé české flóry vychází trend spíše opačný. Rozdíl v terminal velocity sice není průkazný, C1 druhy však mají ve srovnání s druhy celé české květeny menší hmotnost semen a převažují mezi nimi anemochorní taxony. Tyto výsledky mohou být ovlivněny disproporčně vyšším zastoupením druhů čeledi Orchidaceae (s extrémně lehkými semeny) a Asteraceae (vzhledem k morfologii semen obecně považované za anemochorní taxony) mezi C1 druhy. Na druhou stranu právě pro řadu orchidejí a endemických druhů pampelišek a jestřábníků jsme neměli o vlastnostech semen mnoho údajů, takže do analýz téměř nevstupovaly. Výsledky tedy mohou být důsledkem faktu, že C1 druhy jsou obecně malé, kompetičně slabé taxony, o kterých je známo, že mají lehčí semena, která jsou lépe šiřitelná větrem (např. Peat & fitter 1994). Jiné studie, srovnávající různé způsoby disperze druhů ve vztahu k jejich vzácnosti či ohroženosti, však většinou žádné průkazné rozdíly nezjistily (lahti et al. 1991, kelly 1996, kelly & wooDwarD 1996, heGDe & ellstranD 1999) a rovněž většinou neprokázaly rozdíly v hmotnosti semen (eDwarDs & westoby 1996, eriksson & jakobsson 1998, thoMPson et al. 1999).
Jednou z vlastností, u které by člověk nějaké rozdíly mezi vzácnými a hojnými druhy očekával, je produkce semen. Řada studií na dvojicích druhů vzácný-hojný zjistila, že vzácné druhy produkují méně semen (banks 1980, byers & MeaGher 1997, Pirie et al. 2000, walck 2001) a i autoři studií analyzujících větší soubory taxonů dospívají ke stejným výsledkům (eriksson & jakobsson 1998, caDotte & lovett-Doust 2002, laverGne et al. 2004, PilGriM et al. 2004). V rámci porovnání druhů C1 a jim blízce příbuzných hojných jsme však žádné rozdíly v produkci semen nezjistili. Problém může spočívat v celkově malém počtu a hlavně jisté nespolehlivosti údajů z databáze LEDA pro tuto charakteristiku.
Porovnání ekologických charakteristik ukázalo řadu rozdílů mezi C1 a hojnými druhy, např. v hemerobii a urbanitě. Systém hemerobních stupňů popisuje, jak přirozená stanoviště (z hlediska lidského vlivu) rostlina obývá. Škála hemerobních stupňů je plynulá, jednotlivé druhy se mohou vyskytovat ve více kategoriích, jejichž počet pak odráží rozsah tolerance druhu k lidským zásahům do vegetace (klotz et al. 2002). Asi nepřekvapí, že mezi druhy C1 převažují taxony s nižší mírou tolerance k ovlivnění rostlinných společenstev činností člověka. Rozsahem tolerance (tj. počtem hemerobních stupňů) se však druhy C1 a druhy blízce příbuzné hojné neliší. I mezi druhy C1 tak najdeme některé taxony rostoucí

261
Příroda, Praha, 31: 247–279, 2011
v širokém rozmezí hemerobních stupňů, mezi něž patří např. Hypochaeris glabra, Rumex palustris, Sagina nodosa, Sagina subulata, Viola kitaibeliana či Trifolium striatum. Podobně se při porovnání urbanity ukázalo, že mezi druhy C1 je kromě očekávatelného většího zastoupení druhů urbanofóbních, rostoucích striktně mimo lidská sídla, i větší zastoupení druhů urbanofilních, na lidská sídla úzce vázaných, jako je např. Artemisia scoparia, Bromus squarrosus, Eragrostis pilosa či Herniaria hirsuta.
Ve srovnání s druhy blízce příbuznými hojnými i s druhy celé české květeny dosahují druhy C1 menší průměrné výšky. To je ve shodě s výsledky, které ve svých studiích zjistili heGDe & ellstranD (1999), laverGne et al. (2004) a PilGriM et al. (2004). Zjištěná menší výška druhů C1 naznačuje, že se jedná o druhy s menší schopností kompetice o prostor a světlo. To ostatně potvrzují i výsledky srovnání ekologické strategie, kdy se mezi druhy C1 prokázalo menší zastoupení c-strategie. Druhy C1 zřejmě představují druhy ranějších sukcesních stádií, které jsou později v průběhu sukcese vytlačeny silnějšími konkurenty.
To potvrzují i výsledky porovnání Ellenbergových indikačních hodnot. Porovnání s blízce příbuznými hojnými druhy ukazuje, že C1 druhy se vyskytují častěji na otevřených, teplých, bazických a živinami chudých stanovištích. Mezi páry druhů s výrazně rozdílnými nároky na živiny patří např. Ajuga pyramidalis (C1) a A. reptans, Hypericum pulchrum (C1) a H. hirsutum, Listera cordata (C1) a L. ovata, Nuphar pumila (C1) a N. lutea nebo Torilis arvensis (C1) a T. japonica. Mezi páry druhů se značně odlišnými nároky na půdní reakci patří např. Calamagrostis varia (C1) a C. arundinacea, Carex buxbaumii (C1) a C pilu lifera, Carex rupestris (C1) a C. pauciflora, Juncus sphaerocarpus (C1) a J. bufonius nebo Viola elatior (C1) a V. canina subsp. canina. Porovnání s druhy celé české květeny ukazuje větší zastoupení C1 druhů na bazických, živinami chudých a vlhkých stanovištích. Ačkoli nejsou výsledky obou typů porovnání zcela konzistentní, přesto indikují, že C1 druhy jsou kompetičně slabé taxony nucené růst na extrémnějších typech stanovišť. Otevřená a živinami chudá stanoviště jsou v rámci celé Evropy ohrožená zarůstáním a eutrofizací, což vede k silnému ohrožení druhů na těchto stanovištích (viz např. ratcliffe 1984, fischer & stöcklin 1997, bakker & berenDse 1999).
Porovnání výskytu C1 druhů v různých nadmořských výškách a v různých typech habitatů rovněž ukázalo problém vzácnosti a ohrožení stanovišť, na kterých se C1 druhy vyskytují. Řada C1 druhů (např. Agrostis alpina, Cardamine resedifolia, Carex atrata, Carex rupestris, Galium sudeticum, Gnaphalium supinum, Crepis sibirica, Laserpitium archangelica, Ribes petraeum) je omezena na vysoké nadmořské výšky s výskytem nad horní hranicí lesa, přičemž rozloha těchto biotopů je v České republice velmi omezená. Zajímavý je výrazně menší počet C1 druhů vázaných na lesy a křoviny. To může být způsobeno tím, že lesy a křoviny jsou poměrně rozšířeným a relativně málo ohroženým typem habitatu v rámci ČR (härtel et al. 2009).
C1 druhy mají, ve srovnání s blízce příbuznými druhy, menší areály rozšíření. To ukazuje, že zařazení do červeného seznamu v kategorii C1 koreluje s „přirozenou“ vzácností danou malým areálem rozšíření. Typickými příklady jsou druhy Achillea asplenifolia, Cirsium brachycephalum, Daphne cneorum, Lathyrus heterophyllus, Myosotis stenophylla nebo Trifolium retusum.
limitace práce
V našem příspěvku porovnáváme podobně jako řada autorů před námi vlastnosti druhů excerpované z databází. Tento přístup dovoluje porovnání velkého množství znaků pro velké množství druhů, což umožňuje poměrně robustní statistické testování. Na druhou stranu mohou být informace z databází zatíženy řadou chyb a některé údaje zjištěné

262
Gabrielová et al.: Kriticky ohrožené druhy rostlin, jak se odlišují svými vlastnostmi od hojných druhů?
pro zahraniční populace druhů nemusí zcela odpovídat hodnotám znaků stejných druhů rostoucích v České republice. Škála vlastností dostupná z databází je navíc poněkud omezená a je možné, že rozdíly mezi vzácnými a hojnými druhy se objevují na jemnější škále, než jakou jsou databáze schopny zachytit. Rovněž je důležité zmínit, že rozdíly v jednotlivých vlastnostech druhů nemusí nutně znamenat rozdíly v jejich celkové populační dynamice (MünzberGová 2005). Bylo by proto velmi užitečné spojit výsledky takovéto extenzivní studie porovnávající jednotlivé charakteristiky životního cyklu s mnohem detailnějším porovnáním životního cyklu několika málo vybraných druhů.
Náš datový soubor má tu nevýhodu, že údaje o vlastnostech pro řadu druhů chybí (pro každou vlastnost máme údaje pro trochu odlišný podsoubor druhů). Rovněž provádíme velký počet jednotlivých porovnání znaků, které jsou navzájem korelované. Jistě by bylo velmi zajímavé vyhodnocovat všechny vlastnosti najednou v rámci jedné analýzy, to ovšem bohužel není možné vzhledem k velkému počtu chybějících hodnot a relativně malému počtu druhů s údaji pro všechny vlastnosti.
shrnutí
Srovnání biologických a ekologických vlastností druhů C1 a druhů hojných ukázalo, že možné příčiny ohrožení se skrývají spíše ve stanovištních nárocích druhů než v jejich biologických vlastnostech. Přesto i srovnání biologických vlastností ukázalo zajímavé rozdíly především v generativní fázi životního cyklu. Kriticky ohrožené druhy kvetou kratší dobu, čímž se může snížit úspěšnost jejich opylení, zároveň je však u nich průkazně větší zastoupení autokompatibility, takže případné neúspěchy při procesu opylení mohou být kompenzovány autogamií. Dlouhodobé příbuzenské křížení je však spojeno s rizikem ztrát genetické variability a snížení fitness. Srovnání neodhalilo konzistentní rozdíly ve způsobu disperze; na základě našich údajů se nedá určit, jestli jsou kriticky ohrožené druhy rostlin limitované v procesu disperze.
Druhy C1 dorůstají menší výšky, což svědčí o jejich menší kompetiční schopnosti, která se ukázala i při srovnání ekologických strategií. Mezi druhy C1 je oproti druhům blízce příbuzným hojným větší zastoupení s-stratégů a druhů s menší mírou tolerance k lidským zásahům do vegetace. Rovněž mezi nimi najdeme druhy hůře snášející zastínění a vyskytujících se na málo produktivních stanovištích. Kriticky ohrožené druhy jsou tedy často kompetičně slabé druhy ranějších sukcesních stádií, které jsou v průběhu sukcese přerůstány silnějšími konkurenty. Jsou to také druhy extrémnějších typů stanovišť (otevřená, živinami chudá), která jsou však v rámci celé Evropy ohrožená zarůstáním a eutrofizací, což v důsledku vede i k silnému ohrožení druhů na těchto stanovištích. Zdá se tedy, že u celé řady druhů by stačilo zajistit dostatek vhodných stanovišť, případně důsledněji zavádět vhodná managementová opatření a situace těchto druhů by se mohla zlepšit, protože nejsou omezovány svými biologickými vlastnostmi.
summary
The comparison of biological and ecological traits of citically endangered (C1) and common species has shown that the endangerment and rarity of these species might be rather caused by their habitat requirements than by their biological traits. However, the comparison of biological traits has also shown some important differences – mainly in the generative part of the life cycle. The C1 species flower for shorter periods than common

263
Příroda, Praha, 31: 247–279, 2011
species, which could disadvantage them in the process of pollination. On the other hand, there is a higher proportion of self-compatibility between the C1 species, which could compensate the failure in the process of pollination due to self-fertilization. However, long term inbreeding is connected with a risk of loss of genetic diversity and lowering fitness. The comparison did not show any consistent differences in the mode of dispersion; based on our data, it is not possible to assess whether the C1 species are limited in the process of dispersion.
The C1 species are smaller than common species and thus competitively inferior as shown also by the comparison of csr-strategy. The C1 species are in comparison to their common close relatives more frequently s-strategists and susceptible to human impact in the vegetation. They tend to occur in open and nutrient poor habitats. critically endangered species are thus competitively weak species in early successional stages, i.e. they are being overgrown by stronger competitors in the process of succession. They are also species of extreme habitats (open, nutrient poor), endangered by eutrofication all over Europe, which leads to their strong endangerment in these habitats. It appears that, in case of many C1 species, it would be sufficient to ensure enough suitable habitats or consistently implement appropriate management measures and the situation of these species would be improved as they are not constrained by their biological traits.
Poděkování
Tato práce by se neobešla bez Olivera Tackenberga z Johann-Goethe Universität ve Frankfurtu nad Mohanem, který nám poskytl údaje o terminal velocity některých C1 druhů a zpřístupnil nám svou soukromou databázi druhových vlastností, ve které bylo možné vyhledat charakteristiky pro druhy celé české květeny. Za komentáře k rukopisu děkujeme L. Černé, A. Šlechtové a M. Konvičkovi. Studie byla financována z grantu MŠMT VaV 2B06178, výzkumných záměrů MSM 0021620828 a AV0Z60050516 a grantu 206-08-H049.
literatura
bakker j. P., bekker r. M. & thoMPson k. (2000): From a seed bank database towards a seed database. – Z. Ökologie u. Naturschutz 9: 61–72.
bakker j. P. & berenDse f. (1999): Constraints in the restoration of ecological diversity in grassland and heathland communities. – Trends Ecol. Evol. 14: 63–68.
banks J. A. (1980): The reproductive biology of Erythronium propullans Gray and sympatric populations of E. albidum Nutt. (Liliaceae). – B. Torrey Bot. Club 107: 181–188.
bekker r. M., bakker j. P., GranDin u., kalaMees r., MilberG P., PoscholD P., thoMPson k. & willeMs j. h. (1998): Seed size, shape and vertical distribution in the soil: indicators of seed longevity. – Funct. Ecol. 12: 834–842.
bevill r. l. & louDa s. M. (1999): Comparisons of related rare and common species in the study of plant rarity. – Conserv. Biol. 13: 493–498.
bonn s., PoscholD P. & tackenberG o. (2000): Diasporus – a database for diaspore dispersal - concept and applications in case studies for risk assessment. – Zeitschr. Ökol. Umweltschutz 9: 85–97.
byers D. l. & MeaGher t. r. (1997): A comparison of demographic characteristics in a rare and a common species of Eupatorium. – Ecol. Appl. 7: 519–530.
caDotte M. w. & lovett-Doust j. (2002): Ecological and taxonomic differences between rare and common plants of southwestern Ontario. – Ecoscience 9: 397–406.
chytrý M., kuČera t. & koČí M. [eds] (2001): Katalog biotopů České republiky. – Agentura ochrany přírody a krajiny ČR, Praha.

264
Gabrielová et al.: Kriticky ohrožené druhy rostlin, jak se odlišují svými vlastnostmi od hojných druhů?
cotGrave P. & PaGel M. (1997): Predicting and understanding rarity: the comparative approach. – In: kunin w. e. & Gaston k. j. [eds] (1997), The biology of rarity. Causes and consequences of rare-common differences, pp. 237–261, Chapman & Hall, London.
Danihelka j., chrtek j., kaPlan z. & wilD j. (2007): Seznam jmen cévnatých rostlin použitých v nálezové databázi květeny České republiky (verze 18. 6. 2007). – URL: http://www.ibot.cas.cz/sbirky/sez/index.php (6. 3. 2010).
eDwarDs w. & westoby M. (1996): Reserve mass and dispersal investment in relation to geographic range of plant species: phylogenetically independent contrasts. – J. Biogeogr. 23: 329–338.
ellenberG h., weber h. e., Düll r., wirth v. & werner w. (1992): Zeigerwerte von Pflanzen in Mitteleuropa. 3. durchgesehene Auflage. – Verlag Erich Goltze GmbH & Co KG, Göttingen.
eriksson o. & jakobsson a. (1998): Abundance, distribution and life histories of grassland plants: A comparative study of 81 species. – J. Ecol. 86: 922–933.
felsenstein j. (1985): Phylogenies and the comparative method. – Am. Nat. 125: 2–15.fieDler P. l. (1987): Life history and population dynamics of rare and common marispora lilies
(Calochortus Pursh: Liliaceae). – J. Ecol. 75: 977–995.fischer M. & stöcklin j. (1997): Local extinctions of plants in remnants of extensively used calcareous
grasslands 1950–1985. – Conserv. Biol. 11: 727–737.freckelton r. P., harvey P. h. & PaGel M. (2002): Phylogenetic analysis and comparative data: A test
and review of evidence. – Am. Nat. 160: 712–726.Gaston k. j. (1994): Rarity. – Chapman & Hall, London. GriMe j. P. (1979): Plant strategies and vegetation processes. – John Wiley, Chichester.Gustafsson l. (1994): A comparison of biological characteristics and distribution between Swedish
threatened and non-threatened forest vascular plants. – Ecography 17: 39–49.härtel h., lonČáková j. & hoŠek M. [eds] (2009): Mapování biotopů v České republice. Východiska,
výsledky, perspektivy. – Agentura ochrany přírody a krajiny ČR, Praha. heGDe s. G. & ellstranD n. c. (1999): Life history diferences between rare and common flowering
plant species of California and the British Isles. – Int. J. Plant Sci. 160: 1083–1091.hejný s. & slavík b. [eds] (1988): Květena České socialistické republiky 1. – Academia, Praha.hejný s. & slavík b. [eds] (1990): Květena České republiky 2. – Academia, Praha.hejný s. & slavík b. [eds] (1992): Květena České republiky 3. – Academia, Praha.hultén e. & fries M. (1986): Atlas of North European Vascular Plants: North of the Tropic of Cancer.
– Koeltz Scientific Books, Königstein.kelly c. k. (1996): Identifying plant functional types using floristic data bases: Ecological correlates of
plant range size. – J. Veg. Sci. 7: 417–424.kelly c. k. & wooDwarD f. i. (1996): Ecological correlates of plant range size: taxonomies and
phylogenies in the study of plant commonness and rarity in Great Britain. – Philos. Trans. Roy. Soc., Biol. Sci. 351: 1261–1269.
kleyer r. M., bekker r. M., bakker j., knevel i. c., thoMPson k., sonnenschein M., PoscholD P., van GroenenDael j. M., kliMeŠ l., kliMeŠová j., klotz s., rusch G., herMy M., aDriaens boeDeltje G., bossuyt b., enDels P., GötzenberGer l., hoDGson j. G., jackel a.-k., DannenMann a., kühn i., kunzMann D., ozinGa w., röMerMann c., staDler M., schleGelMilch j., steenDaM h. j., tackenberG o., wilMann b., cornelissen j. h. c., eriksson o., Garnier e., fitter a. & Peco b. (2008): The LEDA Traitbase: a database of plant life-history traits of North West Europe. – J. Ecol. 96: 1266–1274.
klotz s., kühn i. & Durka w. (2002): BIOLFLOR – eine Datenbank mit biologisch-ökologischen Merkmalen zur Flora von Deutschland. – Schriftenreihe für Vegetationskunde 38: 1–334.
kliMeŠová j. & kliMeŠ l. (2006): CLO-PLA3 – database of clonal growth of plants from Central Europe. – URL: http://clopla.butbn.cas.cz (14. 5. 2010).
kubát k., hrouDa l., chrtek j. jun., kaPlan z., kirschner j. & ŠtěPánek j. [eds] (2002): Klíč ke květeně České republiky. – Academia, Praha.
kunin w. e. & Gaston k. j. (1997): The biology of rarity. Causes and consequences of rare-common differences. – Chapman & Hall, London.
kunin w. e. & shMiDa a. (1997): Plant reproductive traits as a function of local, regional, and global abundance. – Conserv. Biol. 11: 183–192.

265
Příroda, Praha, 31: 247–279, 2011
lahti t., keMPPainen e., kurtto a. & uotila P. (1991): Distribution and biological characteristics of threatened vascular plants in Finland. – Biol. Conserv. 55: 299–314.
laverGne s., thoMPson j. D., Garnier e. & Debussche M. (2004): The biology and ecology of narrow endemic and widespread plants: a comparative study of trait variation in 20 congeneric pairs. – Oikos 107: 505–518.
May r. M. (1999): Unanswered questions in ecology. – Philos. Trans. Roy. Soc., Biol. Sci., 354: 1951–1959.Meffe G. k. & carroll r. (1994): Principles of conservation biology. – Sinauer Associates, Inc.Meusel h. & jäGer e. j. (1992): Vergleichende Chorologie der zentraleuropäischen Flora. Text
u. Karten. Bd. 3. - Gustav Fischer Verlag Stuttgart New York.Moravec j., balátová-tuláČková e., blažková D., haDaČ e., hejný s., husák Š., jeník j., kolbek j.,
krahulec f., kroPáČ z., neuhäusl r., rybníČek k., řehořek v. & vicherek j. (1995): Rostlinná společenstva České republiky a jejich ohrožení. Ed. 2. – Severočes. Přír., Příloha 1995: 1–206.
MünzberGová z. (2005): Determinants of species rarity: population growth rates of species sharing the same habitat. – Am. J. Bot. 92: 1987–1994.
Murray b. r., thrall P. h., Gill a. M. & nicotra a. b. (2002): How plant life-history and ecological traits relate to species rarity and commonness at varying spatial scales. – Austral Ecol. 27: 291–310.
Peat h. j. & fitter a. h. (1994): Comparative analyses of ecological characteristics of British angiosperms. – Biol. Rev. 69: 95–115.
PilGriM e. s., crawley M. j., DolPhin k. (2004): Patterns of rarity in the native British flora. – Biol. Conserv. 120: 165–174.
Pirie c. D., walMsley s., inGle r., jiMenez a. P., MaGallanes a. s. & kelly c. k. (2000): Investigations in plant commonness and rarity: A comparison of seed removal patterns in the widespread Jatropha standleyi and the endemic J. chamelensis (Euphorbiaceae). – Biol. J. Linn. Soc. 71: 501–512.
PoschloD P., kleyer M., jackel a. k., DanneMann a. & tackenberG o. (2003): BIOPOP – a database of plant traits and Internet application for nature conservation. – Folia Geobot. 38: 263–271.
Procházka f. [ed.] (2001): Černý a červený seznam cévnatých rostlin České republiky (stav v roce 2000). – Příroda, Praha, 18: 1–166.
ratcliffe D. a. (1984): Post-medieval and recent changes in British vegetation: the culmination of human influence. – New Phytol. 98: 73–100.
slavík b. [ed.] (1995): Květena České republiky 4. – Academia, Praha.slavík b. [ed.] (1997): Květena České republiky 5. – Academia, Praha.slavík b. [ed.] (2000): Květena České republiky 6. – Academia, Praha.slavík b. & ŠtěPánková j. [eds] (2004): Květena České republiky 7. – Academia, Praha.s-Plus (2000): Professional edition for Windows, release 2. (1999) – MathSoft Inc., Needham,
Massachusetts, USA.ŠtěPánková j. [ed.] (2010): Květena České republiky 8. – Academia, Praha.ter braak c. j. & ŠMilauer P. (1998): CANOCO reference manual and user`s guide to Canoco for
Windows: software for canonical community ordination, version 4.5. – Microcomputer Power, Ithaca, New York, USA.
thoMPson k., bakker j. P. & bekker r. M. (1997): Soil seed banks of North-West Europe: methodology, density and longevity. – University Press, Cambridge.
thoMPson k., Gaston k. j. & banD s. r. (1999): Range size, dispersal and niche breadth in the herbaceous flora of central England. – J. Ecol. 87: 150–155.
tutin t. G., heywooD v. h., burGess n. a., valentine D. h., walters s. M., webb D. a. [eds] (1964–1993): Flora Europaea 1–5. – University Press, Cambridge.
walck j. l., baskin j. M. & baskin c. c. (2001): Relative competitive abilities and growth characteristics of narrowly endemic and geographically widespread Solidago species (Asteraceae). – Am. J. Bot. 86: 820–828.
weiher e., van Der werf a., thoMPson k., roDerick M., Garnier e. & eriksson o. (1999): Challenging Theophrastus: a common core list of plant traits for functional ecology. – J. Veg. Sci. 10: 609–620.
westoby M., leishMan M. r. & lorD j. M. (1995): On misinterpreting the „phylogenetic correction“. – J. Ecol. 83: 531–534.
witkowski e. t. f. & laMont b. b. (1997): Does the rare Banksia goodii have inferior vegetative, reproductive or ecological attributes compared with its widespread co-occuring relative B. ardneri? – J. Biogeogr. 24: 469–482.

266
Gabrielová et al.: Kriticky ohrožené druhy rostlin, jak se odlišují svými vlastnostmi od hojných druhů?
Příloha 1. seznam c1 druhů a druhů blízce příbuzných hojnýchC1 druhy – kriticky ohrožené druhy ČR podle Černého a Červeného seznamu cévnatých rostlin ČR (Procházka 2001). Blízce příbuzné hojné – druhy blízce příbuzné C1 druhům, v ČR co do rozšíření hojné (viz Metodika). - C1 druhy nemají žádného blízce příbuzného v ČR, nebo je druh blízce příbuzný hojný rovněž kriticky nebo silně ohrožený. * Druhy ze stejného typu habitatu. Nomenklatura podle kubát et al. 2002.
Appendix 1. list of c1 species and their more common close relativesC1 species – critically endangered species of the Czech Republic according to the Red List of Vascular Plant Species of the Czech Republic (Procházka 2001). Common close relatives – species closely related to C1 species occuring in the Czech Republic (for details on selection see Methods). - C1 species do not have any common close relatives in the Czech Republic or the closely related species is also critically or strongly endangered or extinct. * Species from the same habitat. The nomenclature follows kubát et al. 2002.
c1 Blízce příbuzné hojnéAconitum lycoctonum subsp. vulparia Aconitum lycoctonum subsp. lycoctonumAdenophora liliifolia -Adonis flammea -Agropyron pectinatum Elytrigia repensAgrostemma githago -Agrostis alpina Agrostis caninaAchillea asplenifolia* Achillea pratensis*Aira caryophyllea -Ajuga pyramidalis Ajuga reptans Alchemilla baltica -Alchemilla fissa -Alchemilla glabricaulis -Anacamptis pyramidalis -Androsace septentrionalis -Anemone narcissiflora -Angelica palustris* Angelica sylvestris*Aphanes australis* Aphanes arvensis*Aposeris foetida -Arabis nemorensis* Arabis hirsuta*Arabis sudetica Arabis hirsuta Arenaria grandiflora* Arenaria serpyllifolia*Arnoseris minima -Artemisia pancicii* Artemisia pontica*Artemisia scoparia Artemisia campestris Asplenium adiantum-nigrum -Asplenium adulterinum* Asplenium trichomanes* Asplenium trichomanes subsp. pachyrhachis* Asplenium trichomanes subsp. inexpectans*Aster alpinus Aster amellusAster tripolium subsp. pannonicus* Aster amellus*Astragalus arenarius* Astragalus austriacus* Atriplex rosea* Atriplex tatarica*Batrachium baudotii* Batrachium aquatile*Batrachium penicillatum* Batrachium fluitans*

267
Příroda, Praha, 31: 247–279, 2011
c1 Blízce příbuzné hojnéBeckmannia eruciformis -Betula oycoviensis* Betula pendula*Bolboschoenus glaucus -Bolboschoenus maritimus -Botrychium matricariifolium -Brassica elongata subsp. elongata -Bromus arvensis* Bromus japonicus* Bromus racemosus -Bromus secalinus* Bromus hordeaceus* Bromus squarrosus* Bromus japonicus* Buphthalmum salicifolium -Bupleurum affine* Bupleurum falcatum* Bupleurum longifolium subsp. vapincense -Bupleurum rotundifolium -Bupleurum tenuissimum* Bupleurum falcatum *Calamagrostis phragmitoides Calamagrostis canescens Calamagrostis pseudophragmites Calamagrostis epigejos Calamagrostis stricta* Calamagrostis arundinacea* Calamagrostis varia* Calamagrostis arundinacea*Callitriche hermaphroditica -Callitriche platycarpa* Callitriche stagnalis*Campanula cervicaria* Campanula glomerata*Campanula gelida -Cardamine amara subsp. opicii* Cardamine amara subsp. amara*Cardamine parviflora Cardamine flexuosaCardamine resedifolia Cardamine impatiens Cardaminopsis petraea* Cardaminopsis arenosa*Carex alba* Carex humilis*Carex atrata Carex pilulifera Carex buxbaumii Carex pilulifera Carex capillaris -Carex dioica -Carex hordeistichos -Carex chordorrhiza Carex ovalis Carex pediformis subsp. macroura* Carex pediformis subsp. rhizodes*Carex pseudobrizoides Carex praecox Carex rupestris Carex paucifloraCarex stenophylla* Carex brizoides*Carex vaginata* Carex panicea*Carex viridula subsp. pseudoscandinavica -Carlina acaulis subsp. caulescens* Carlina acaulis subsp. acaulis*Carlina biebersteinii* Carlina vulgaris*Catabrosa aquatica -Centaurea montana subsp. mollis -Centaurea stenolepis* Centaurea pseudophrygia* Centaurium littorale subsp. compressum* Centaurium erythraea*

268
Gabrielová et al.: Kriticky ohrožené druhy rostlin, jak se odlišují svými vlastnostmi od hojných druhů?
c1 Blízce příbuzné hojnéCerastium alsinifolium -Cerastium fontanum* Cerastium holosteoides subsp. triviale*Cerastium tenoreanum* Cerastium brachypetalum*Ceratophyllum submersum* Ceratophyllum demersum*Ceterach officinarum -Cimicifuga europaea -Cirsium brachycephalum Cirsium palustre Cladium mariscus -Cleistogenes serotina -Conioselinum tataricum -Conringia orientalis -Corrigiola littoralis -Cortusa matthioli -Crataegus lindmanii* Crataegus praemonticola*Crepis mollis subsp. mollis Crepis mollis subsp. hieracioidesCrepis pannonica* Crepis biennis*Crepis setosa -Crepis sibirica Crepis paludosa Crocus albiflorus -Crocus heuffelianus -Crypsis aculeata -Cryptogramma crispa -Cuscuta approximata* Cuscuta europaea*Cuscuta lupuliformis -Cyperus flavescens* Cyperus fuscus*Cyperus michelianus* Cyperus fuscus*Cystopteris dickieana* Cystopteris fragilis* Dactylorhiza bohemica -Dactylorhiza traunsteineri subsp. carpatica -Dactylorhiza fuchsii subsp. sooana* Dactylorhiza fuchsii subsp. fuchsii*Dactylorhiza incarnata subsp. serotina -Dactylorhiza maculata subsp. elodes -Dactylorhiza maculata subsp. maculata Dactylorhiza fuchsii subsp. fuchsiiDactylorhiza maculata subsp. transsilvanica* Dactylorhiza fuchsii subsp. fuchsii*Dactylorhiza majalis subsp. turfosa* Dactylorhiza majalis subsp. majalis*Dactylorhiza traunsteineri* Dactylorhiza majalis subsp. majalis*Danthonia alpina* Danthonia decumbens*Daphne cneorum* Daphne mezereum*Dianthus arenarius subsp. bohemicus -
Dianthus carthusianorum subsp. capillifrons* Dianthus carthusianorum subsp. carthusianorum*
Dianthus carthusianorum subsp. sudeticus Dianthus carthusianorum subsp. carthusianorum
Dianthus superbus subsp. superbus -Diphasiastrum oellgaardii Diphasiastrum alpinum Diphasiastrum tristachyum* Diphasiastrum complanatum*Draba nemorosa -

269
Příroda, Praha, 31: 247–279, 2011
c1 Blízce příbuzné hojnéDracocephalum austriacum -Drosera anglica* Drosera rotundifolia*Drosera intermedia* Drosera rotundifolia*Dryopteris cristata* Dryopteris carthusiana* Echium russicum* Echium vulgare*Elatine alsinastrum* Elatine hydropiper* Eleocharis quinqueflora Eleocharis palustris Epipactis greuteri Epipactis helleborine Epipactis helleborine subsp. orbicularis* Epipactis helleborine subsp. helleborine*Epipactis leptochila subsp. neglecta -Epipactis microphylla* Epipactis atrorubensEpipactis nordeniorum* Epipactis helleborine*Epipactis pontica* Epipactis helleborine*Epipactis pseudopurpurata* Epipactis purpurata*Epipactis tallosii* Epipactis helleborine* Epipogium aphyllum -Equisetum variegatum* Equisetum ramosissimum* Eragrostis pilosa* Eragrostis minor* Erica tetralix* Erica carnea* Eriophorum gracile -Eryngium planum* Eryngium campestre* Erysimum repandum -Erythronium dens-canis -Euphorbia salicifolia* Euphorbia esula*Euphorbia seguieriana subsp. minor -Euphrasia micrantha* Euphrasia nemorosa* Euphrasia picta* Euphrasia rostkoviana*Euphrasia slovaca* Euphrasia rostkoviana*Festuca amethystina* Festuca heterophylla*Festuca drymeia* Festuca altissima*Festuca psammophila* Festuca pallens*Festuca trichophylla* Festuca rubra*Festuca vaginata subsp. dominii* Festuca pallens* Festuca versicolor -Filago lutescens* Filago minima* Filago vulgaris* Filago minima*Filipendula ulmaria subsp. picbaueri* Filipendula ulmaria subsp. ulmaria*Gagea bohemica subsp. saxatilis -Galium austriacum* Galium pumilum*Galium sudeticum Galium pumilum Galium tricornutum* Galium aparine*Genista sagittalis -Gentiana punctata -Gentiana verna -Gentianella amarella subsp. amarella -Gentianella campestris subsp. baltica -

270
Gabrielová et al.: Kriticky ohrožené druhy rostlin, jak se odlišují svými vlastnostmi od hojných druhů?
c1 Blízce příbuzné hojnéGentianella germanica subsp. germanica -Gentianella lutescens subsp. carpatica -Gentianella lutescens subsp. lutescens -Gentianella obtusifolia subsp. sturmiana -Gentianella praecox subsp. bohemica -Geranium divaricatum -Gladiolus palustris -Glaucium corniculatum -Glaux maritima -Gnaphalium supinum Gnaphalium sylvaticum Goodyera repens -Groenlandia densa -Gymnadenia conopsea subsp. montana* Gymnadenia conopsea subsp. conopsea*Gymnadenia densiflora Gymnadenia conopsea Gypsophila paniculata -Hackelia deflexa -Hammarbya paludosa -Hedysarum hedysaroides -Heleochloa schoenoides -Helianthemum grandiflorum subsp. grandiflorum Helianthemum obscurum subsp. obscurumHerniaria hirsuta* Herniaria glabra*Hibiscus trionum -Hieracium albinum -Hieracium asperulum -Hieracium blyttianum -Hieracium corconticum -Hieracium engleri -Hieracium fuscoatrum -Hieracium chlorocephalum -Hieracium chrysostyloides -Hieracium inuloides -Hieracium kablikianum -Hieracium kalksburgense -Hieracium luteistylum -Hieracium nigrostylum -Hieracium nivimontis -Hieracium pedunculare -Hieracium pseudalbinum -Hieracium riphaeum -Hieracium rubrum -Hieracium schustleri -Hieracium silesiacum -Hieracium stoloniflorum -Hieracium subortum -Hieracium tortuosum -Hieracium tubulascens -

271
Příroda, Praha, 31: 247–279, 2011
c1 Blízce příbuzné hojnéHieracium villosum -Hierochloë odorata Hierochloë australis Hierochloë repens Hierochloë australis Himantoglossum adriaticum -Hippocrepis comosa -Hippuris vulgaris -Hypericum elegans -Hypericum pulchrum* Hypericum hirsutum*Hypochaeris glabra* Hypochaeris radicata* Chamaecytisus albus* Chamaecytisus austriacus* Chenopodium murale -Chimaphila umbellata -Illecebrum verticillatum -Iris aphylla subsp. fieberi -Iris aphylla subsp. novakii -Isoëtes echinospora -Isoëtes lacustris -Juncus atratus Juncus acutiflorusJuncus capitatus -Juncus gerardii Juncus compressus Juncus sphaerocarpus* Juncus bufonius*Juncus subnodulosus* Juncus articulatus* Juncus tenageia* Juncus bufonius* Juniperus communis subsp. alpina Juniperus communis subsp. communisJurinea cyanoides -Knautia arvensis subsp. pseudolongifolia Knautia arvensis subsp. arvensisKochia prostrata* Kochia scoparia*Lappula semicincta* Lappula squarrosa* Laser trilobum -Laserpitium archangelica Laserpitium prutenicum Lathyrus aphaca* Lathyrus pratensis*Lathyrus heterophyllus* Lathyrus sylvestris*Lathyrus hirsutus -Lathyrus nissolia -Lathyrus palustris* Lathyrus niger*Lathyrus pannonicus subsp. pannonicus -Lathyrus pisiformis* Lathyrus vernus* Leucanthemum adustum* Leucanthemum ircutianum* Leucojum aestivum* Leucojum vernum*Ligularia sibirica -Limodorum abortivum -Linaria arvensis Linaria vulgaris Lindernia procumbens -Linum hirsutum subsp. hirsutum -Liparis loeselii -Listera cordata* Listera ovata*

272
Gabrielová et al.: Kriticky ohrožené druhy rostlin, jak se odlišují svými vlastnostmi od hojných druhů?
c1 Blízce příbuzné hojnéLittorella uniflora -Luronium natans -Luzula spicata* Luzula sudetica* Malaxis monophyllos -Marrubium peregrinum -Marrubium vulgare -Mercurialis ovata* Mercurialis perennis* Minuartia caespitosa -Minuartia corcontica -Minuartia smejkalii -Minuartia viscosa -Misopates orontium -Moneses uniflora -Montia fontana -Myosotis stenophylla* Myosotis sylvatica*Myricaria germanica -Najas minor -Nasturtium microphyllum* Nasturtium officinale* Nigella arvensis -Notholaena marantae -Nuphar pumila* Nuphar lutea* Nymphaea alba -Nymphaea candida -Nymphoides peltata -Onosma arenaria -Ophrys apifera -Ophrys holosericea -Ophrys insectifera -Orchis mascula subsp. mascula -Orchis palustris -Orchis tridentata -Orchis ustulata subsp. aestivalis -Orchis ustulata subsp. ustulata -Ornithogalum pyrenaicum subsp. sphaerocarpum -Orobanche arenaria -Orobanche artemisiae-campestris -Orobanche caesia Orobanche ramosaOrobanche coerulescens -Orobanche picridis -Orobanche purpurea subsp. bohemica Orobanche ramosaOrobanche purpurea subsp. purpurea Orobanche ramosaOrobanche teucrii* Orobanche caryophyllacea*Papaver lecoqii* Papaver dubium*Pedicularis exaltata -Pedicularis sudetica Pedicularis sylvatica Peucedanum carvifolia* Peucedanum alsaticum*

273
Příroda, Praha, 31: 247–279, 2011
c1 Blízce příbuzné hojnéPhyllitis scolopendrium -Phyteuma orbiculare subsp. flexuosum -Phyteuma orbiculare subsp. montanum -Pinguicula vulgaris subsp. bohemica -Pinus × celakovskiorum -Plantago atrata subsp. sudetica Plantago lanceolata Plantago maritima subsp. ciliata -Poa alpina Poa supina Poa crassipes -Poa riphaea -Polycnemum arvense -Polycnemum majus -Polygala amara subsp. brachyptera -Polygala serpyllifolia Polygala vulgaris Potamogeton × angustifolius -Potamogeton polygonifolius -Potamogeton praelongus -Potentilla crantzii subsp. serpentini Potentilla aurea Potentilla patula* Potentilla heptaphylla*Potentilla rupestris -Potentilla sterilis -Primula elatior subsp. corcontica Primula elatior subsp. elatiorPrimula minima -Prunus fruticosa* Prunus avium*Prunus padus subsp. borealis Prunus padus subsp. padusPrunus tenella -Pseudognaphalium luteoalbum -Pseudolysimachion spurium subsp. foliosum* Pseudolysimachion maritimum*Puccinellia limosa Puccinellia distans Pulegium vulgare -Pulicaria dysenterica -Pulicaria vulgaris -Pulsatilla patens -Pulsatilla vernalis -Pyrola chlorantha -Pyrola media* Pyrola minor* Quercus virgiliana* Quercus petraea* Radiola linoides -Ranunculus sardous Ranunculus bulbosus Reseda phyteuma* Reseda lutea*Rhinanthus serotinus subsp. vernalis -Rhodiola rosea -Rhynchospora fusca -Ribes petraeum Ribes alpinum Rubus ambrosius -Rubus amphimalacus -

274
Gabrielová et al.: Kriticky ohrožené druhy rostlin, jak se odlišují svými vlastnostmi od hojných druhů?
c1 Blízce příbuzné hojnéRubus caflischii -Rubus geminatus -Rubus chaerophylloides -Rubus chamaemorus -Rubus lividus -Rubus lusaticus -Rubus pruinosus -Rubus pyramidalis -Rubus scaber -Rubus vestitus -Rumex palustris* Rumex maritimus* Rumex stenophyllus Rumex crispus Sagina nodosa* Sagina procumbens*Sagina subulata* Sagina saginoides*Salix appendiculata -Salix bicolor -Salix lapponum var. daphneola -Salix herbacea -Salix myrsinifolia -Salix myrtilloides -Salix repens* Salix rosmarinifolia* Salvia aethiopis -Salvia austriaca* Salvia nemorosa*Samolus valerandii -Saxifraga oppositifolia -Saxifraga rosacea subsp. steinmannii -Scabiosa lucida subsp. calcicola* Scabiosa ochroleuca*Scorzonera parviflora* Scorzonera humilis* Scrophularia vernalis* Scrophularia nodosa* Sedum villosum Sedum albumSenecio doria -Senecio erucifolius* Senecio jacobaea* Senecio paludosus -Senecio paludosus subsp. paludosus -Serratula lycopifolia* Serratula tinctoria*Scheuchzeria palustris -Schoenus ferrugineus -Schoenus nigricans -Sideritis montana -Silene gallica -Silene rupestris -Silene viscosa -Sorbus graeca Sorbus danubialisSorbus alnifrons -Sorbus gemella -Sorbus hardeggensis -

275
Příroda, Praha, 31: 247–279, 2011
c1 Blízce příbuzné hojnéSorbus quernea -Sorbus rhodanthera -Sorbus sudetica -Spergula pentandra* Spergula morisoni* Spergularia maritima* Spergularia rubra*Spergularia salina* Spergularia rubra*Spiranthes spiralis -Stachys germanica Stachys alpina Stipa borysthenica* Stipa pennata*Stipa eriocaulis* Stipa pulcherrima* Stipa smirnovii -Stipa zalesskii -Taraxacum alpestre -Taraxacum ambrosium -Taraxacum ancoriferum -Taraxacum arachnoideum -Taraxacum arcuatum -Taraxacum argutum -Taraxacum austrinum -Taraxacum bavaricum -Taraxacum bessarabicum -Taraxacum bracteatum -Taraxacum brandenburgicum -Taraxacum cognatum -Taraxacum dentatum -Taraxacum fartoris -Taraxacum huterianum -Taraxacum indigenum -Taraxacum irrigatum -Taraxacum litigiosum -Taraxacum madidum -Taraxacum mendax -Taraxacum paludem-ornans -Taraxacum pauckertianum -Taraxacum pseudobalticum -Taraxacum quaesitum -Taraxacum ranunculus -Taraxacum skalinskanum -Taraxacum subdolum -Taraxacum turfosum -Taraxacum uvidum -Tephroseris aurantiaca -Tephroseris longifolia subsp. moravica* Tephroseris crispa*Thalictrum simplex subsp. galioides* Thalictrum lucidum*Thesium arvense* Thesium linophyllon*Thesium dollineri* Thesium linophyllon*

276
Gabrielová et al.: Kriticky ohrožené druhy rostlin, jak se odlišují svými vlastnostmi od hojných druhů?
c1 Blízce příbuzné hojnéThesium ebracteatum -Thesium rostratum -Thymus carpaticus -Thymus pulegioides subsp. carniolicus* Thymus pulegioides subsp. chamaedrys*Tillaea aquatica -Tofieldia calyculata -Tordylium maximum -Torilis arvensis Torilis japonica Trapa natans -Trifolium badium* Trifolium spadiceum*Trifolium patens* Trifolium campestre* Trifolium retusum* Trifolium repens*Trifolium striatum* Trifolium pratense* Triglochin maritimum -Trigonella monspeliaca -Trichomanes speciosum -Trinia glauca -Typha laxmannii* Typha angustifolia* Typha shuttleworthii* Typha latifolia* Utricularia bremii* Utricularia australis* Utricularia intermedia* Utricularia australis*Utricularia ochroleuca* Utricularia australis*Utricularia vulgaris* Utricularia australis*Valerianella carinata Valerianella locusta Veratrum nigrum* Veratrum album*Verbascum speciosum* Verbascum lychnitis*Veronica bellidioides Veronica serpyllifoliaVeronica opaca* Veronica polita*Vicia oreophila* Vicia cracca*Viola alba* Viola suavis*Viola elatior* Viola canina subsp. canina*Viola kitaibeliana Viola arvensis Viola tricolor subsp. curtisii* Viola tricolor subsp. tricolor*Vitis vinifera subsp. sylvestris -Vulpia bromoides* Vulpia myuros*Wolffia arrhiza -Xanthium strumarium* Xanthium albinum*

277
Příroda, Praha, 31: 247–279, 2011
Příloha 2. Přehled porovnávaných vlastností.označení zdrojů B BIOLFLOR (klotz et al. 2002)BP BIOPOP (PoscholD et al. 2003)C CLO-PLA (kliMeŠová & kliMeŠ 2006)D DIASPORUS (bonn et al. 2000)L LEDA (kleyer at al. 2008)E ellenberG et al. (1992)Kl kubát et al. (2002) Kv hejný & slavík (1988, 1990, 1992), slavík (1995, 1997, 2000), slavík & ŠtěPánková (2004), ŠtěPánková (2010)T thoMPson et al. (1997)
Vegetativní znakyživotní formaKl: fanerofyt, geofyt, hemikryptofyt, hydrofyt, chamaefyt, makrofanerofyt, nanofanerofyt,
terofyt délka životaB, Kv: (i) jednoleté, (ii) dvouleté, (iii) víceleté plodící jednou za život, (iv) víceleté s opakovanou
reprodukcíprůměrná výška rostliny v metrechKl
listová růžiceB: (i) bez růžice, (ii) s částečnou růžicí, (iii) tvoří růžicivytrvalost listůB: (i) krátkověké (listy jarních rostlin, v létě odumírají), (ii) sezónní (listy vytrvávají během
vegetační sezóny, na zimu odumírají), (iii) přezimující (zelené listy vytrvávají přes zimu, na jaře odumírají nebo jsou nahrazeny), (iv) stálezelené
anatomie listůB: sukulentní, skleromorfní, mesomorfní, hygromorfní, helomorfní, hydromorfníspecifická listová plocha (specific leaf area, SLA)L: poměr plochy listu vůči hmotnosti jeho sušiny
v mm²/mgklonalitaC (řada znaků testovaných zvlášť, všechny rozdíly nesignifikantní, proto nevypisováno
samostatně): (i) typ orgánů klonálního růstu (kliMeŠová & kliMeŠ 2006 rozlišují celkem 17 typů, např. horizontální nadzemní výběžek, turion, hlíza, výmladek, rhizom apod., podrobnosti v manuálu k databázi CLO-PLA na http://clopla.butbn.cas.cz/), (ii) přetrvání primárního kořene (přetrvává/nepřetrvává), (iii) délka života genety (roky), (iv) banka pupenů – počet podzemních i nadzemních pupenů v různých výškách, (v) typ banky pupenů (vytrvalá, sezónní, potenciální a kombinace), (vi) role orgánů klonálního růstu (nezbytná, doplňková, regenerativní, žádná), (vii) délka života vegetativního výběžku (1, 2, > 2 roky), (viii) přetrvání spojení mezi mateřskou a dceřinou rametou (1, 2, > 2 roky), (ix) míra klonálního množení – počet dceřiných ramet na mateřskou rametu, (x) laterální šíření (< 0,01, 0,01–0,25, > 0,25 m/rok), (xi) způsob větvení (monopodiální, sympodiální, dichotomické), (xii) způsob odnožování (intra/extravaginální), (xiii) velikost dceřiných ramet (stejně velké/menší než mateřská rostlina), (xiv) generační překryv výběžků
Generativní znakyobdobí kveteníKl: začátek, konec a délka trvání kvetení v rozmezí měsícůtyp reprodukceB: (i) pouze semeny, (ii) většinou semeny – zřídka vegetativně, (iii) semeny a vegetativně,
(iv) většinou vegetativně – zřídka semeny, (v) vegetativnězpůsob opyleníB, Kv: anemogamie, hydrogamie, autogamie, entomogamie, ostatní (geitonogamie,
kleistogamie, pseudokleistogamie)inkompatibilitaB (schopnost vytvoření klíčivých semen po opylení vlastním pylem): autokompatibilní;
autoinkompatibilní, přechodný typdiklinieB (pohlavnost květů – jak jsou v rámci květů/rostliny/a mezi rostlinami rozmístěny samčí
a samičí orgány): (i) oboupohlavné (monoklinické), (ii) jednodomé (monoecické), (iii) dvoudomé (diecické), (iv) gynomonoecické, (v) andromonoecické, (vi) gynodioecické, (vii) androdioecické, (viii) triecické, (ix) samčí sterilní
dichogamieB, Kv (časově nestejné dozrávání prašníků a blizen): proterogynie, homogamie, proterandrieodměna za opyleníB: žádná odměna, nektar, pyl, olej, klamavá odměna

278
Gabrielová et al.: Kriticky ohrožené druhy rostlin, jak se odlišují svými vlastnostmi od hojných druhů?
Vlastnosti diasporhmotnost semene v mgB, BP, D, L
rozměry semenB, Kv: délka, šířka a výška v mmtvar semenB, BP, D, L: vyjádřený vzorcem Vs = ∑ (xi – průměr (x))²/n, kde n = 3, x1 = délka/délka, x2 = výška/
délka a x3 = šířka/délka a Vs pak variance těchto rozměrů, bezrozměrné čísloterminal velocity BP, D, L: maximální rychlost, které semeno dosáhne při pádu za bezvětří v m/szpůsob šíření semenBP, D, L: (i) anemochorie – šíření větrem, (ii) hydrochorie – šíření vodou, (iii)
epizoochorie – šíření zvířaty na povrchu jejich těla, (iv) endozoochrie – šíření zvířaty po průchodu trávicím traktem, (v) hemerochorie – šíření člověkem, (vi) ostatní – šíření na krát kou vzdálenost. Informace čerpány z databáze DIASPORUS (bonn et al. 2000), která uvádí procento citací v literatuře pro určitý typ disperze u daného druhu. Navíc se soustředí především na dálkovou disperzi semen (long distance dispersal) – tedy potenciál šířit se mezi populacemi na vhodný a zatím neobsazený habitat. Další způsoby šíření na krátké vzdálenosti (v rámci populací) jsou shrnuty do jediné kategorie („ostatní“).
produkce semen L: Stanovuje se pěti různými způsoby jako: (i) počet všech semen ramety/jedince, (ii) počet semen v jednom květenství/na jednom stonku vynásobený celkovým počtem květenství/stonků produkovaných rametou, (iii) počet semen na květ/plodenství vynásobený odhadem celkového počtu květů/plodenství, (iv) váha všech semen rostlinného jedince dělená váhou známého počtu semen, (v) váha všech semen produkovaných rostlinou.
přežívání v půdní semenné banceT: Podle schopnosti semen zachovávat si klíčivost při delším pobytu v půdě rozlišují thoMPson et al. (1997) tři typy půdní semenné banky: (i) transientní (semena přetrvávají v půdě méně než jeden rok), (ii) krátkodobě persistentní (semena přetrvávají v půdě 1–5 let) a (iii) dlouhodobě persistentní (semena přetrvávají v půdě pět a více let). Protože se však semena určitého druhu mohou v databázi v závislosti na zdroji a způsobu stanovení přežívání objevit ve všech kategoriích životnosti, zavedli bekker et al. (1998) novou veličinu – longevity index. Ten je vyjádřen vzorcem L = (SP + LP)/(T +SP + LP), kde (SP + LP) je celkový počet záznamů krátkodobě a dlouhodobě persistentních semen a (T + SP + LP) celkový počet záznamů transientních, krátkodobě a dlouhodobě pe rsis-tentních semen pro určitý druh v databázi. Longevity index nabývá hodnot od 0 (striktně transientní) do 1 (striktně persistentní). Podmínkou pro jeho použití jsou alespoň čtyři záznamy v databázi (bakker et al. 2000). Tento požadavek však u řady druhů není splněn, proto se často používá i hodnota maximal longevity type, tedy maximální hodnota přežívání v půdní semenné bance, která se v databázi vyskytuje.
Ekologické charakteristikycsr - strategieB (podle GriMe 1979): c-, s-, r-, cs-, cr-, sr-, csr-stratégurbanitaB (vyjadřuje afinitu rostlin k urbánním oblastem): (i) urbanofóbní – vždy mimo lidská sídla, (ii)
mírně urbanofóbní – většinou mimo lidská sídla, (iii) urbanoneutrální, (iv) mírně urbanofilní – často v osídlených oblastech, (v) urbanofilní – druhy omezené na lidská sídla
hemerobieB (měřítko toho, jak přirozená stanoviště rostlina obývá): (i) ahemerobní – přirozené bez lidského vlivu, (ii) oligohemerobní – slabě ovlivněné činností člověka, (iii) mesohemerobní – mírně ovlivněné činností člověka, (iv) β - euhemerobní – druhy intenzivně obhospodařovaných pastvin, luk a lesů, (v) α - euhemerobní – druhy polních kultur, intenzivně obhospodařovaných lesů se slabě vyvinutou půdou, (vi) polyhemerobní – rosliny obývající stanoviště zcela přeměněná, uměle vytvořená činností člověka
Ellenbergovy indikační hodnotyE: pro světlo, teplotu, kontinentalitu, vlhkost, půdní reakci a dusík
Rozšířeníobecná informace o stanovišti Kl, Kv
výškový stupeň Kl: (i) nížiny – do 200 m n. m., (ii) pahorkatina – 200–400 m n. m., (iii) podhůří – 400–700 m n. m., (iv) hory – 700–1300 m n. m., (v) subalpínský stupeň – nad 1300 m n. m.
velikost areálu rozšíření: Informace o velikosti areálů rozšíření C1 druhů a druhů blízce příbuzných hojných jsme získali pomocí počítačového zpracování publikovaných map areálů (hultén & fries 1986, Meusel & jäGer 1992, tutin et al. 1964–1993). Mapy jsme oscanovali

279
Příroda, Praha, 31: 247–279, 2011
a rektifikovali v programu ArcGIS 9.0 (ESRI, Redlands, CA, USA). Z digitalizovaných map jsme vybrali jen evropské části areálů a stanovili jsme jejich celkovou plochu.
typ stanoviště: Na základě zařazení druhů do příslušných fytocenologických jednotek a biotopů podle Moravec et al. (1995) a chytrý et al. (2001) a na základě obecné informace o stanovišti excerpované z Květeny ČR a Klíče ke Květeně ČR jsme za pomoci převodních tabulek z Katalogu biotopů (chytrý et al. 2001) rozdělili druhy do devíti základních typů biotopů (podle chytrý et al. 2001): vodní toky a nádrže; mokřady a pobřežní vegetace; prameniště a rašeliniště; skály, sutě a jeskyně; alpinské bezlesí; sekundární trávníky a vřesoviště; křoviny; lesy; biotopy silně ovlivněné nebo vytvořené člověkem.

280
Gabrielová et al.: Kriticky ohrožené druhy rostlin, jak se odlišují svými vlastnostmi od hojných druhů?

281
Příroda, Praha, 31: 281–298, 2011
Změny v rozšíření kriticky ohrožených druhů rostlin v České republice na základě síťových mapchanges in the distribution of critically endangered plant species in the czech Republic on the basis of grid maps
Lucie Černá2, JarMiLa gabrieLová1, 2, ZuZana MünZbergová1, 3 & vLastik rybka4
1Katedra botaniky, Přírodovědecká fakulta Univerzity Karlovy, Benátská 2, CZ-128 01 Praha 2; [email protected], [email protected] ochrany přírody a krajiny České republiky, Nuselská 34, CZ-140 00 Praha 4; [email protected]ý ústav Akademie věd České republiky, v. v. i., Zámek 1, CZ-252 43 Průhonice4Botanická zahrada hl. m. Prahy, Nádvorní 134, CZ-171 00 Praha 7; [email protected]
Abstract: For the effective conservation of critically endangered plant species it is important to have detailed information about the changes in their distribution over time. In this study we collected data from literature, herbaria, and flora databases to gain information about historical and actual distribution of 283 selected critically endangered plant species of the Czech Republic. Grid maps of changes in their distribution were reconstructed for different time periods – before 1949, between years 1950 and 1999 and after 2000. The results of the analyzed grid maps show that among the critically endangered species we can find species which have always been rare and have not changed their distribution over time. We can also find species which were relatively abundant in the past but have lost many localities. Some rare species even began to spread. However, most analysed species are decreasing and the decrease was stronger during the second half of the 20th century. The attached grid maps represent basic types of changes in the distribution of this very diverse group of studied species during the last 100 years. The distribution data are stored in a database of characteristics of critically endangered plant species and can be used as a specialised tool for qualified selection and prioritisation in the species conservation.Keywords: critically endangered species, distribution data, grid mapping, rate of decrease, causes of danger
Abstrakt: Efektivní ochrana ohrožených druhů rostlin se neobejde bez kvalitních hodnotících kritérií, mezi které patří i informace o změnách rozšíření ohrožených druhů v čase. V rámci této studie byly rozsáhlou excerpcí odborné literatury, herbářových sbírek a floristických databází získány údaje o historickém a aktuálním rozšíření vybraných 283 kriticky ohrožených druhů rostlin (C1). Pro tyto vybrané C1 druhy byly sestrojeny síťové mapy rozšíření ve třech časových periodách – rozšíření do roku 1949, v letech 1950–1999 a po roce 2000. Z výsledků analýz síťových map vyplývá, že mezi kriticky ohroženými druhy můžeme nalézt druhy, které byly vždy vzácné a početnost jejich populací se v průběhu času neměnila, i druhy, které byly v minulosti relativně hojné a v důsledku různých změn prostředí se dnes ocitly na pokraji vyhynutí. Byly rovněž identifikovány druhy v minulosti zřejmě vzácné, které se v současné době začaly šířit. Většina analyzovaných druhů však spadá do kategorie druhů v různém stupni ubývajících, o něco více pak v průběhu druhé poloviny 20. století. Uvedené příklady síťových map reprezentují základní typy změn rozšíření této

282
Černá et al.: Změny v rozšíření kriticky ohrožených druhů rostlin v České republice
velmi různorodé skupiny druhů v posledních 100 letech. Získaná nálezová data z této práce, uspořádaná v přehledné databázi charakteristik C1 rostlin, mohou do budoucna sloužit jako odborný podklad pro kvalifikovanější rozhodování při prioritizaci druhové ochrany.Klíčová slova: kriticky ohrožené druhy, míra úbytku, nálezová data, příčiny ohrožení, síťové mapování.
Úvod
Pro efektivní ochranu vzácných a ohrožených druhů rostlin je nezbytná nejen znalost jejich aktuálního rozšíření a stavu současných lokalit výskytu, ale i to, k jakým změnám v jejich rozšíření v minulosti docházelo. Došlo u vybraného druhu oproti minulosti k dramatickému poklesu v počtu jeho lokalit? Nebo se jedná o druh, který má konstantně nízký počet lokalit a byl tedy vzácný již v minulosti? Jaké jsou vlastně příčiny případného ústupu vybraného druhu z naší přírody? To jsou otázky, na které bychom měli při sestavování červených seznamů či výběru druhů do záchranných programů umět odpovědět.
K odlišení druhů vzácných od těch skutečně ohrožených (tj. ubývajících) je tedy nutné prozkoumat historii jejich rozšíření a stanovit míru případného úbytku počtu lokalit v čase. Možný pokles lokalit a populací v daném časovém období je dokonce jedním z kritérií IUCN pro klasifikaci ohrožených druhů rostlin (IUCN 2001) – dle těchto kritérií je druh zařazen do kategorie „kriticky ohrožený“, pokud došlo v posledních deseti letech k poklesu jeho populací nejméně o 80 %. Informaci o míře úbytku populací (zejména pokud k rapidnímu snížení početnosti došlo v minulých 50 letech) vyžaduje i jedno z aktuálních kritérií pro výběr druhů do záchranných programů ohrožených druhů v České republice (Marhoul et al. 2003).
K výrazným změnám rozšíření rostlinných (i živočišných) druhů začalo docházet v průběhu 20. století, zejména pak v jeho druhé polovině, a to především kvůli neustále sílícímu tlaku člověka na přírodu a krajinu. Těžba nerostných surovin, rostoucí nároky průmyslu a výstavba dopravní infrastruktury produkující velké množství škodlivých emisí měly za následek rapidní zhoršení kvality životního prostředí. Na ubývání velkého množství dříve běžných druhů se výrazně podepsaly také změny hospodaření v krajině. Ta začala být s nástupem kolektivizace v padesátých letech a s ní spojeným přechodem na velkovýrobu a mechanizaci v zemědělství intenzivně využívána – malá políčka byla spojena v širé lány, mokřadní půdy odvodněny, od pravidelné a mozaikovité seče a pastvy se upustilo. Rychlým tempem tak začala v krajině mizet pestrá mozaika stanovišť, kterou využívaly druhy k migraci: políčka, záhumenky, pastviny, nivní a slatinné louky, písčiny, remízky, meze apod. – to vše hostilo specifickou květenu a množství dnes již vzácných a ohrožených druhů rostlin (např. teplomilné plevele, orchideje).
Od počátku se botanici i amatérští přírodovědci snažili výskyt vzácných druhů rostlin na našem území zaznamenávat – sestavovali soupisy druhů konkrétních území, sepisovali rukopisné práce či publikace zabývající se regionální květenou (vaněČek 1969, Šourek 1969, kubát 1986 a mnoho dalších), zakládali kartotéky (např. Chánova floristická kartotéka jihočeské pobočky ČBS) i herbářové sbírky (muzejní i menší soukromé). Během dlouhodobého floristického výzkumu bylo shromážděno obrovské množství údajů o výskytu rostlin na území ČR v minulosti, avšak komplexní znázornění jejich rozšíření pomocí map chybělo až do vydání prvního dílu Fytokartografických syntéz ČSR (slavík 1986), po kterém v devadesátých letech následoval ještě díl druhý a třetí (slavík 1990 a 1998). Tyto syntézy, znázorňující rozšíření rostlinných druhů na území ČSR/ČR formou síťových map, měly být nutným předstupněm dlouhé roky chystaného Atlasu rozšíření

283
Příroda, Praha, 31: 281–298, 2011
cévnatých rostlin ČSR, který však do dnešní doby nebyl vydán. Z hlediska hodnocení míry případného úbytku ohrožených druhů rostlin je však trochu škoda, že toto dílo rekonstruuje rozšíření druhů za uplynulá dvě století floristického výzkumu a nerozlišuje mezi recentními a historickými nálezy, které jsou tak na mapách znázorněny pohromadě.
V moderní době vznikají za účelem sběru nálezových dat ohrožených druhů elektronické databáze, které umožňují ukládání velkého množství údajů a snadné generování map rozšíření konkrétních druhů. Touto cestou dnes vznikají různé atlasy flóry pro určitá území. Vzhledem k náročnosti zpracování velkého množství dat vznikají častěji atlasy lokálního charakteru, např. pro území národního parku Podyjí (Grulich 1997) či oblast Bílých Karpat (jonGePier & Pechanec 2006). Nové generace map jsou již schopné zachytit dramatické změny v rozšíření vzácných druhů rostlin, ke kterým v posledních desetiletích dochází a oproti fytokartografickým syntézám se tak již mohou zabývat kontrastem historického a aktuálního rozšíření.
Cílem této práce bylo pomocí excerpce odborné literatury, herbářů, floristických databází i konzultací s botaniky získat relevantní údaje o historickém a současném rozšíření skupiny kriticky ohrožených druhů rostlin (kategorie C1, Procházka 2001) z území celé ČR, a to alespoň pro polovinu z nich. Na základě shromážděných dat a vyhodnocení případného úbytku druhů v posledních 100 letech pomocí mapového zobrazení jsme chtěli prozkoumat tuto velmi různorodou skupinu druhů, abychom získali základní představu o tom, jak velká část ze sledovaného souboru druhů je tvořena druhy skutečně ubývajícími, na které bychom měli z hlediska ochrany přírody soustředit naši pozornost nejvíce, a jaká je míra jejich úbytku ve vztahu k jejich vzácnosti.
Metodika
Analýzy byly prováděny na skupině kriticky ohrožených druhů rostlin (dále jen C1) Černého a červeného seznamu cévnatých rostlin ČR (Procházka 2001). Z praktických důvodů jsme z tohoto souboru druhů vyloučili taxonomicky problematické rody Alchemilla, Hieracium, Rubus a Taraxacum. Po vyloučení všech zástupců těchto rodů nám tak v souboru ke studiu zůstalo přes 400 kriticky ohrožených druhů rostlin, pro které měly být nálezové údaje sbírány.
Sběr nálezových dat byl zahájen rozsáhlou excerpcí odborné botanické literatury. Na katedře botaniky PřF UK v Praze byla systematicky procházena a excerpována významná botanická periodika, např. časopis Preslia či Zprávy České (Československé) botanické společnosti, ze kterých byly excerpovány jakékoliv údaje o výskytu C1 rostlin na našem území. Neméně důležitým zdrojem dat byly i regionální sborníky, různé floristické studie, rukopisné práce a diplomové práce (excerpované zejména na PřF MU v Brně) zaměřené na studium konkrétních C1 druhů či květenu určité floristicky zajímavé oblasti. K vyhledávání starších botanických studií a publikací jsme využívali sérii botanických bibliografií Bibliographia botanica Čechoslovaca (Čechica), vydávanou od roku 1952 Botanickým ústavem Akademie věd ČR. Při excerpci byl zapisován každý kladný, lokalizovaný a datovaný nález příslušného C1 druhu v kterémkoli časovém období. Údaje ze základní botanické literatury typu Nová Květena ČSR (Dostál 1989), Květena ČSR/ČR (hejný & slavík 1988, 1990, 1992, slavík 1995, 1997, 2000, slavík & ŠtěPánková 2004, ŠtěPánková 2010), Klíč ke květeně ČR (kubát et al. 2002) a Červená kniha ohrožených a vzácných druhů rostlin ČR (Čeřovský et al. 1999), které byly excerpovány zejména zpočátku projektu, tak mohly být využity jen zčásti, neboť většina údajů o lokalitách výskytu uvedených v těchto publikacích není datována. Každopádně však tyto publikace sloužily

284
Černá et al.: Změny v rozšíření kriticky ohrožených druhů rostlin v České republice
v závěru projektu jako praktické vodítko při hodnocení stavu rozšíření C1 druhů v ČR, neboť obsahovaly důležité informace o mylně uváděných či pochybných lokalitách a informace o současném stavu historických lokalit. S excerpcí literatury současně probíhal sběr dat z botanických materiálů regionálních středisek AOPK ČR a chráněných krajinných oblastí (plány péče, inventarizační průzkumy, mapové podklady v GIS apod.), případně národních parků (botanické materiály KRNAP). Touto rozsáhlou literární excerpcí byly získány tisíce nálezových dat.
Významným zdrojem nálezových dat byl tzv. Dominův materiál deponovaný v Botanickém ústavu AV ČR – rozsáhlé dílo botanika Karla Domina a jeho spolupracovníků obsahující rozpisy floristické literatury a zápisky o nálezech vzácných druhů rostlin z území ČSR do r. 1945. Excerpován byl i materiál navazující na dílo Karla Domina, obsahující výpisky floristické literatury až do roku 1990. V devadesátých letech pak již začala na půdě BÚ AV ČR vznikat elektronická databáze FLDOK (floristická dokumentace, http://florabase.cz/databanka/), shrnující publikované i nepublikované údaje o nálezech cévnatých rostlin, která je dodnes postupně doplňována o další záznamy. Export údajů týkajících se C1 rostlin z databáze FLDOK byl použit jako další významný zdroj dat pro tuto práci. Podařilo se nám rovněž získat nálezová data C1 rostlin z České národní fytocenologické databáze (http://www.sci.muni.cz/botany/vegsci/dbase.php?lang=cz), floristické databáze Bílých Karpat BKFLORA (jonGePier & Pechanec 2006) či nálezové databáze Jihočeské pobočky ČBS (http://jpcbs.prf.jcu.cz/dbjc/index.html). Získáno bylo i několik soukromých databází botaniků – oslovováni byli někteří autoři Květeny ČR a experti na konkrétní C1 druhy či regiony. Důležitým zdrojem údajů o rozšíření C1 druhů byly také regionální kartotéky – např. floristická kartotéka nálezů jižních Čech založená v roce 1980 Václavem Chánem (deponovaná v Jihočeském muzeu) či kartotéka cévnatých rostlin Moravského krasu založená v šedesátých letech 20. stol. Ludmilou Vaněčkovou (deponovaná na správě CHKO Moravský kras).
Byly využity také údaje z Nálezové databáze ochrany přírody (dále jen NDOP), kterou spravuje AOPK ČR. Tato databáze byla spuštěna počátkem roku 2008, tedy v době, kdy už byl projekt téměř v polovině svého řešení. Vznikla spojením menších databází regionálních pracovišť AOPK ČR a zpočátku musela projít rozsáhlou validací nálezových údajů významných druhů živočichů a rostlin, která probíhá prakticky dodnes. Z tohoto důvodu jsme její data, týkající se především aktuálního výskytu C1 druhů, využili až v samotném závěru projektu. Aktuální údaje o rozšíření C1 druhů jsme dále čerpali ze série Additamenta ad floram Reipublicae Bohemicae, která je postupně publikována ve Zprávách České botanické společnosti.
Významným zdrojem údajů o rozšíření rostlin především v minulosti jsou herbářové sbírky, které nám formou herbářových dokladů poskytují hmatatelné důkazy o výskytech vzácných druhů rostlin na území ČR. Před samotnou excerpcí herbářových údajů bylo nejprve nutné rozsáhlý studovaný soubor C1 druhů značně zúžit, a to na druhy, jejichž kompletní excerpce z herbářových sbírek je reálná v časovém horizontu jednoho roku. Nakonec bylo vybráno 67 C1 druhů, pro které se měly údaje v herbářích sbírat. Jedná se např. o několik zástupců čeledí Orchidaceae, Apiaceae, Fabaceae či kriticky ohrožené zástupce čeledi Pyrolaceae, a také např. druhy, o kterých víme, že oproti minulosti významně ustoupily, ale jejich historické rozšíření je málo známé (např. Radiola linoides). Seznam excerpovaných druhů z herbářů uvádíme v Příloze 1.
Za účelem excerpce vybraných 67 C1 druhů byly od konce roku 2009 postupně navštěvovány herbářové sbírky po celé ČR – kromě velkých českých a moravských herbářů (PR, PRC, BRNM, BRNU) byly osobně navštíveny i menší regionální herbáře (OP, OLM, MP, LIT, ROZ, CB, PL, MMI, HR, MJ, OL, SUM, OSM, FMM, ZMT). Z herbářových sbírek,

285
Příroda, Praha, 31: 281–298, 2011
které jsme z časových důvodů nemohli osobně navštívit (GM, OH, KM, KHMS, HOMP, OMJ, VYM, CELM, CESK, MZ, KMKV, CHOM, OVMB, NJM, NZM Valtice, herbářové sbírky muzeí ve Svitavách, Mostě, Ústí nad Labem a Prostějově), nám ve spolupráci s jejich kurátory byly údaje k vybraným druhům elektronicky zasílány (zkratky herbářů podle vozárová & sutorý 2001). S excerpcí byly položky některých C1 druhů v herbářových sbírkách současně revidovány.
Řada údajů získaných z herbářových sbírek nemohla být využita, protože byly neúplné a nepřesné – od velmi zeširoka popsaných lokalit („Böhmen“, „Gesenke“, „Brno – okolí“) po chybějící datum sběru položky či jméno sběratele. Na starších schedách byly české místní názvy lokalit často uvedené v latinském či německém jazyce (navíc psané kurentem), což vyžadovalo následné dohledávání v odborné literatuře (Pfohl 1931, henDrych & jirásek 2002, soják 2007) či konzultace s kurátory sbírek. I přes zmíněné obtíže byly z herbářových sbírek po celé ČR nakonec excerpovány tisíce nálezových údajů.
Pro informaci o rozšíření C1 rostlin na území ČR byly zaznamenávány nálezy od počátku 19. století až po nejaktuálnější údaje. Pro budoucí zobrazení změn v rozšíření vybraných C1 druhů byly stanoveny následující časové periody, které byly následně mezi sebou porovnávány: výskyt druhu do r. 1949, výskyt v letech 1950–1999 a výskyt po roce 2000, který jsme stanovili jako výskyt aktuální.
Excerpovaná data z literatury, kartoték a herbářů byla ukládána nejprve do databáze vytvořené v programu MS Excel, ve které byla průběžně upravována. Kromě běžných úprav dat (např. sjednocení formátu souřadnic, odstranění duplicit apod.) však bylo zásadní mít jednotlivé nálezy lokalizované, aby bylo možné mapy rozšíření zrekonstruovat. Nejběžnějším způsobem zobrazení výskytu rostlinných a živočišných druhů na větším prostorovém měřítku je zakreslení lokalit do polí středoevropské mapovací sítě. Základní pole síťového mapování mají velikost zhruba 12 × 11,1 km a na území ČR se jich nachází celkem 679 (včetně neúplných pohraničních). Pokud byl zaznamenán výskyt daného druhu alespoň na jedné lokalitě uvnitř určitého pole v dané časové vrstvě, získalo toto pole značku výskytu. Velká část sebraných dat byla lokalizována na tato základní pole síťového mapování (případně na jejich čtvrtiny), a to především široce popsané lokality (často u herbářových údajů), které nebylo možné přesněji lokalizovat. Pokud však byla lokalita nálezu podrobněji popsána, snažili jsme se o bližší lokalizaci, a to na katastry (u lokalit definovaných obcí), maloplošně chráněná území (rezervace a přírodní památky) nebo souřadnice v systému WGS-84, či S-JTSK. Při práci jsme se neobešli bez serveru mapy.cz (http://www.mapy.cz), turistických map a především prostředí Nálezové databáze ochrany přírody spravované AOPK ČR (tzv. NDOP, http://portal.nature.cz), kde bylo možné pracovat s mapami ZABAGED® (http://www.cuzk.cz). Pomocí těchto mapových podkladů bylo možné některé nálezy nejen snadno lokalizovat, ale část z nich přímo zakreslit a uložit do databáze jako konkrétní body či polygony (např. rybníky, lesní celky).
Při lokalizaci nálezů jsme naráželi na řadu problémů, se kterými se přirozeně již potýkaly a potýkají podobné studie mapující rozšíření rostlinných druhů. Např. jsme se často setkávali s názvy lokalit, pro které na území ČR existuje více míst se stejným názvem (typicky obce se jménem „Sedlec“, „Břežany“ apod.). V takových případech nám často při hledání správné lokality pomohla publikace Dějiny floristického výzkumu v Čechách (kláŠterský et al. 1982), kde bylo možné dohledat, v jakém území autor nálezu botanizoval. Údaje, které se nám nepodařilo s jistotou lokalizovat, nebyly pro analýzy použity. Dalším problémem byly široce popsané nálezy s lokalizací na rozhraní více síťových polí (např. „Bojanovice – údolí Hlubokého potoka“ na rozhraní polí 7061 a 7062). Takové údaje buď vůbec nebyly lokalizovány a z analýz vyloučeny, nebo byl výskyt vymapován vždy do pole s nejnižším číslem (v uvedeném příkladu tedy do 7061). Nic neobvyklého nebyly

286
Černá et al.: Změny v rozšíření kriticky ohrožených druhů rostlin v České republice
také zaniklé lokality, jako zaniklé obce ve vojenských prostorech, zazemněné rybníky či vytěžené lesní porosty.
Dále jsme museli vyřešit problém s datováním excerpovaných údajů – u nemalé části dat (především u starších herbářových) chybělo datum (rok) nálezu. Bylo tedy nutné některé nálezy uměle datovat. Situaci jsme měli trochu zjednodušenou tím, že nám pro analýzy rozšíření stačilo mít nález datován alespoň do některé ze stanovených časových vrstev. U nedatovaných herbářových údajů do jisté míry napověděly staré schedy, u kterých jsme mohli snadno odhadnout, že spadají jistě do časové periody s nálezem do r. 1949. Někdy napovědělo jméno sběratele a pokud spadalo uvedené rozmezí let, kdy autor podle publikace kláŠterský et al. (1982) žil (popřípadě intenzivně botanizoval), do obou časových vrstev (první a druhá polovina 20. století), byl takový nález evidován v obou těchto časových periodách. Pro nedatované publikované údaje bylo použito jako datum nálezu alespoň datum publikace, a to v případě, kdy bylo jasné, že autor publikace popisuje vlastní nález a neuvádí (necituje) starý nedatovaný nález jiného (většinou staršího) autora. Datované údaje bez autora nálezu byly vyřešeny uvedením „Anonymus“ místo jména sběratele. Nedatované údaje bez autora nálezu byly z analýz vyloučeny, neboť nebylo možné takové nálezy jakkoliv zařadit.
Upravené, lokalizované a datované údaje uložené v Excelové databázi byly ke konci projektu společně s daty získanými z floristických databází importovány do Nálezové databáze ochrany přírody (NDOP). Výhoda této databáze je ta, že umožňuje snadno a rychle zobrazit síťové mapy výskytu druhů v různých časových periodách. NDOP obsahuje mnoho aktuálních nálezů C1 rostlin (např. data z mapování biotopů NATURA 2000). Na druhou stranu tak obrovská databáze čítající 5,8 milionu záznamů o výskytu cévnatých rostlin na území ČR obsahuje také určité procento chyb. Importovaná data v databázi NDOP tak musela zprvu projít zběžnou kontrolou – byly vygenerovány předběžné síťové mapy rozšíření a opraveny či odstraněny zřejmé chyby. Ke kontrole vygenerovaných map jsme využívali výsledky již zmíněných fotokartografických syntéz (slavík 1986, 1990, 1998), se kterými jsme naše mapy porovnávali a mohli tak lépe odhalit na první pohled špatně lokalizované nálezy. Na základě této zběžné kontroly síťových map bylo nakonec vybráno 283 C1 druhů, pro něž jsme měli k dispozici dostatek nálezových dat a tato data bylo možné považovat za spolehlivá.
Analýzy změn rozšíření vybraných C1 druhů spočívaly v porovnání počtu obsazených síťových polí příslušným druhem mezi jednotlivými časovými vrstvami. Stanoven byl tedy počet obsazených polí od r. 1800 do r. 1949 (I), v letech 1950–1999 (II) a po roce 2000 až do současnosti (III, tuto časovou vrstvu jsme považovali za aktuální výskyt). Změnu počtu obsazených polí mezi jednotlivými vrstvami pak charakterizují následující proměnné U1 a U2:
U1 = (I – II)/I - procentuální změna počtu obsazených polí v II. oproti I. časové vrstvě (nabývá záporných hodnot značících, že druh přibyl, až hodnoty 1 značící, že druh zcela vymizel);
U2 = (II – III)/II - procentuální změna počtu obsazených polí v III. oproti II. časové vrstvě (nabývá záporných hodnot značících, že druh přibyl, až hodnoty 1 značící, že druh zcela vymizel).
Pro výsledné hodnoty parametrů U1 a U2 byly nakresleny histogramy zobrazující procentuální zastoupení druhů a rozložení počtů druhů dle jejich změn v rozšíření. Pro parametr U1 bylo zobrazeno jen 268 druhů, neboť byly vyloučeny druhy, pro které jsme neměli data z I. časové vrstvy. Míra úbytku druhů ve vztahu k jejich vzácnosti či hojnosti v minulosti byla zobrazena pomocí bodového grafu s logaritmickým měřítkem.
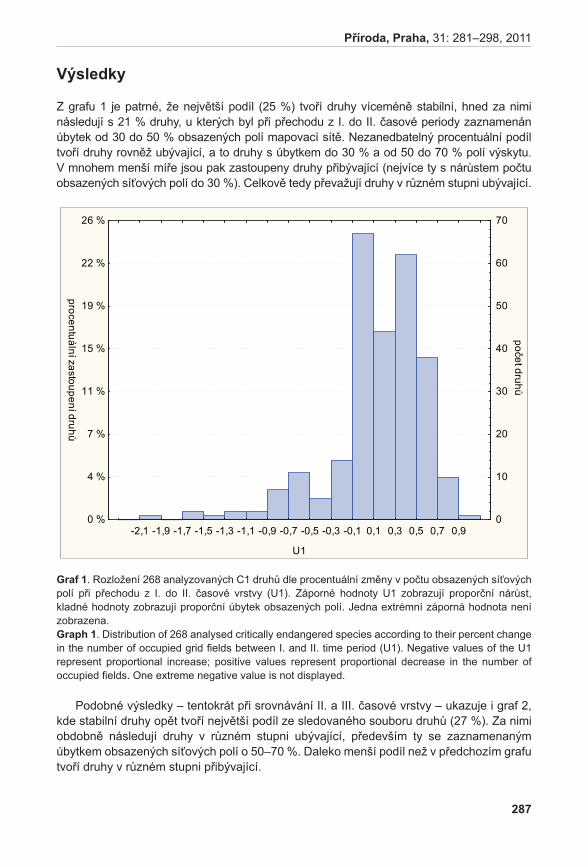
287
Příroda, Praha, 31: 281–298, 2011
výsledky
Z grafu 1 je patrné, že největší podíl (25 %) tvoří druhy víceméně stabilní, hned za nimi následují s 21 % druhy, u kterých byl při přechodu z I. do II. časové periody zaznamenán úbytek od 30 do 50 % obsazených polí mapovací sítě. Nezanedbatelný procentuální podíl tvoří druhy rovněž ubývající, a to druhy s úbytkem do 30 % a od 50 do 70 % polí výskytu. V mnohem menší míře jsou pak zastoupeny druhy přibývající (nejvíce ty s nárůstem počtu obsazených síťových polí do 30 %). Celkově tedy převažují druhy v různém stupni ubývající.
!
Graf 1. Rozložení 268 analyzovaných C1 druhů dle procentuální změny v počtu obsazených síťových polí při přechodu z I. do II. časové vrstvy (U1). Záporné hodnoty U1 zobrazují proporční nárůst, kladné hodnoty zobrazují proporční úbytek obsazených polí. Jedna extrémní záporná hodnota není zobrazena.Graph 1. Distribution of 268 analysed critically endangered species according to their percent change in the number of occupied grid fields between I. and II. time period (U1). Negative values of the U1 represent proportional increase; positive values represent proportional decrease in the number of occupied fields. One extreme negative value is not displayed.
Podobné výsledky – tentokrát při srovnávání II. a III. časové vrstvy – ukazuje i graf 2, kde stabilní druhy opět tvoří největší podíl ze sledovaného souboru druhů (27 %). Za nimi obdobně následují druhy v různém stupni ubývající, především ty se zaznamenaným úbytkem obsazených síťových polí o 50–70 %. Daleko menší podíl než v předchozím grafu tvoří druhy v různém stupni přibývající.
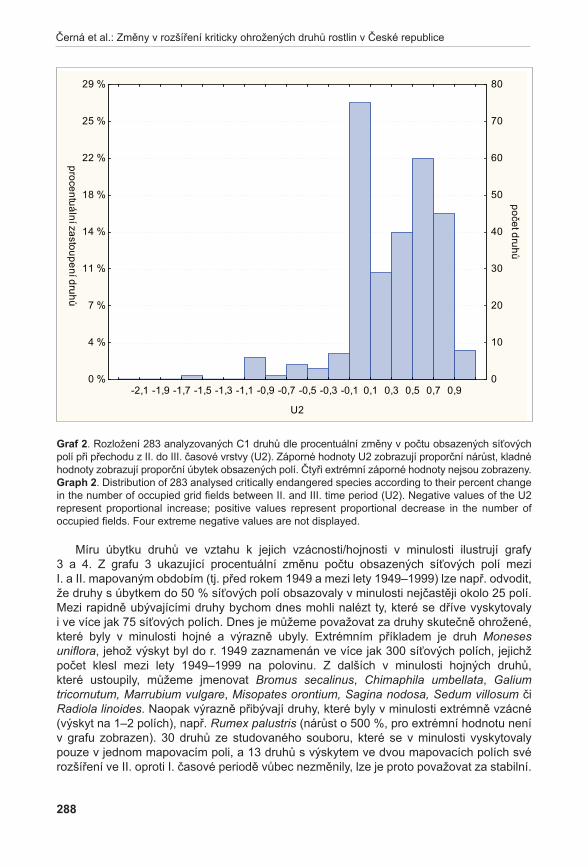
288
Černá et al.: Změny v rozšíření kriticky ohrožených druhů rostlin v České republice
Graf 2. Rozložení 283 analyzovaných C1 druhů dle procentuální změny v počtu obsazených síťových polí při přechodu z II. do III. časové vrstvy (U2). Záporné hodnoty U2 zobrazují proporční nárůst, kladné hodnoty zobrazují proporční úbytek obsazených polí. Čtyři extrémní záporné hodnoty nejsou zobrazeny.Graph 2. Distribution of 283 analysed critically endangered species according to their percent change in the number of occupied grid fields between II. and III. time period (U2). Negative values of the U2 represent proportional increase; positive values represent proportional decrease in the number of occupied fields. Four extreme negative values are not displayed.
Míru úbytku druhů ve vztahu k jejich vzácnosti/hojnosti v minulosti ilustrují grafy 3 a 4. Z grafu 3 ukazující procentuální změnu počtu obsazených síťových polí mezi I. a II. mapovaným obdobím (tj. před rokem 1949 a mezi lety 1949–1999) lze např. odvodit, že druhy s úbytkem do 50 % síťových polí obsazovaly v minulosti nejčastěji okolo 25 polí. Mezi rapidně ubývajícími druhy bychom dnes mohli nalézt ty, které se dříve vyskytovaly i ve více jak 75 síťových polích. Dnes je můžeme považovat za druhy skutečně ohrožené, které byly v minulosti hojné a výrazně ubyly. Extrémním příkladem je druh Moneses uniflora, jehož výskyt byl do r. 1949 zaznamenán ve více jak 300 síťových polích, jejichž počet klesl mezi lety 1949–1999 na polovinu. Z dalších v minulosti hojných druhů, které ustoupily, můžeme jmenovat Bromus secalinus, Chimaphila umbellata, Galium tricornutum, Marrubium vulgare, Misopates orontium, Sagina nodosa, Sedum villosum či Radiola linoides. Naopak výrazně přibývají druhy, které byly v minulosti extrémně vzácné (výskyt na 1–2 polích), např. Rumex palustris (nárůst o 500 %, pro extrémní hodnotu není v grafu zobrazen). 30 druhů ze studovaného souboru, které se v minulosti vyskytovaly pouze v jednom mapovacím poli, a 13 druhů s výskytem ve dvou mapovacích polích své rozšíření ve II. oproti I. časové periodě vůbec nezměnily, lze je proto považovat za stabilní.
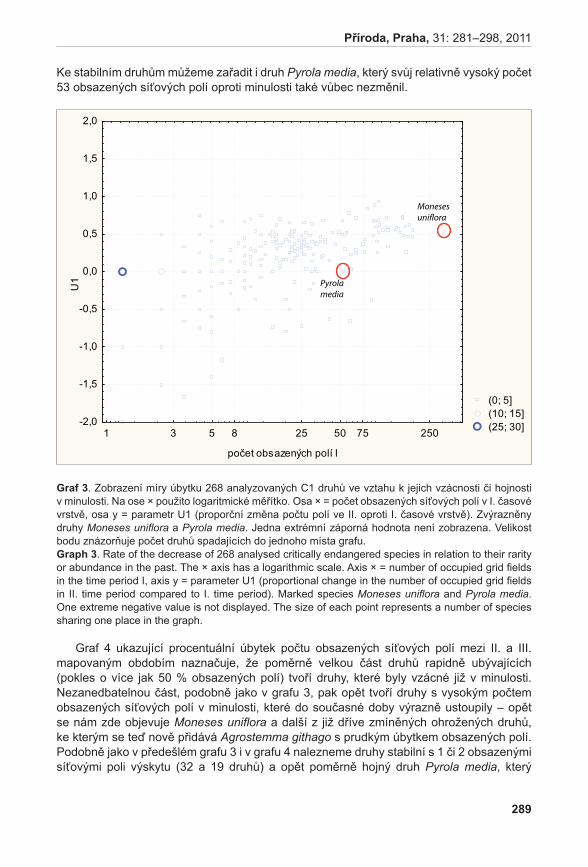
289
Příroda, Praha, 31: 281–298, 2011
Ke stabilním druhům můžeme zařadit i druh Pyrola media, který svůj relativně vysoký počet 53 obsazených síťových polí oproti minulosti také vůbec nezměnil.
"#$%!('!)*+*,!",-.!/0/1!20234)0/!2!3,*4536!-!7087*9:;<!)*=<>!?,*+1!)*24/@@@
Pyrolamedia
Monesesuni�ora
Graf 3. Zobrazení míry úbytku 268 analyzovaných C1 druhů ve vztahu k jejich vzácnosti či hojnosti v minulosti. Na ose × použito logaritmické měřítko. Osa × = počet obsazených síťových polí v I. časové vrstvě, osa y = parametr U1 (proporční změna počtu polí ve II. oproti I. časové vrstvě). Zvýrazněny druhy Moneses uniflora a Pyrola media. Jedna extrémní záporná hodnota není zobrazena. Velikost bodu znázorňuje počet druhů spadajících do jednoho místa grafu. Graph 3. Rate of the decrease of 268 analysed critically endangered species in relation to their rarity or abundance in the past. The × axis has a logarithmic scale. Axis × = number of occupied grid fields in the time period I, axis y = parameter U1 (proportional change in the number of occupied grid fields in II. time period compared to I. time period). Marked species Moneses uniflora and Pyrola media. One extreme negative value is not displayed. The size of each point represents a number of species sharing one place in the graph.
Graf 4 ukazující procentuální úbytek počtu obsazených síťových polí mezi II. a III. mapovaným obdobím naznačuje, že poměrně velkou část druhů rapidně ubývajících (pokles o více jak 50 % obsazených polí) tvoří druhy, které byly vzácné již v minulosti. Nezanedbatelnou část, podobně jako v grafu 3, pak opět tvoří druhy s vysokým počtem obsazených síťových polí v minulosti, které do současné doby výrazně ustoupily – opět se nám zde objevuje Moneses uniflora a další z již dříve zmíněných ohrožených druhů, ke kterým se teď nově přidává Agrostemma githago s prudkým úbytkem obsazených polí. Podobně jako v předešlém grafu 3 i v grafu 4 nalezneme druhy stabilní s 1 či 2 obsazenými síťovými poli výskytu (32 a 19 druhů) a opět poměrně hojný druh Pyrola media, který
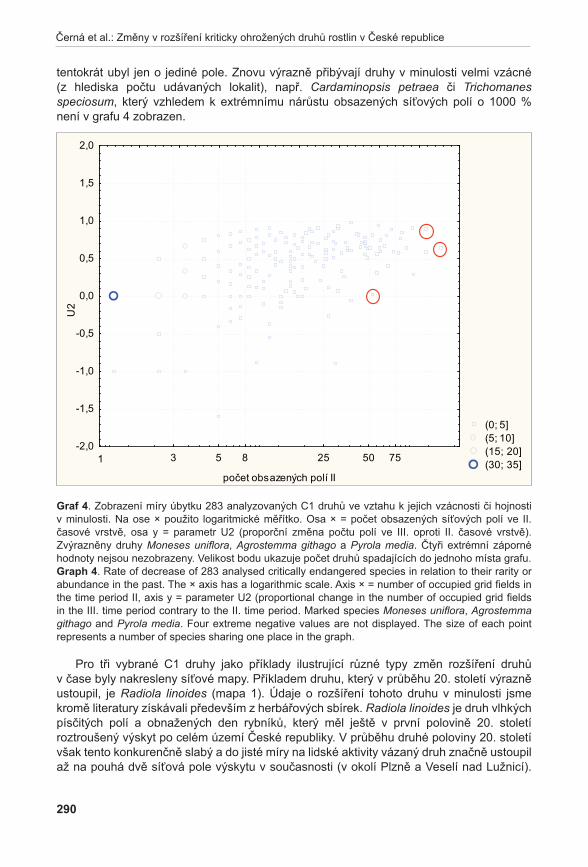
290
Černá et al.: Změny v rozšíření kriticky ohrožených druhů rostlin v České republice
tentokrát ubyl jen o jediné pole. Znovu výrazně přibývají druhy v minulosti velmi vzácné (z hlediska počtu udávaných lokalit), např. Cardaminopsis petraea či Trichomanes speciosum, který vzhledem k extrémnímu nárůstu obsazených síťových polí o 1000 % není v grafu 4 zobrazen.
"#$%!A'!)*+*,!",-.!/0/1!20234)0/!2!3,*4536!-!7087*9:;<!)*=<>!?,*+1!)*24/@@@Graf 4. Zobrazení míry úbytku 283 analyzovaných C1 druhů ve vztahu k jejich vzácnosti či hojnosti v minulosti. Na ose × použito logaritmické měřítko. Osa × = počet obsazených síťových polí ve II. časové vrstvě, osa y = parametr U2 (proporční změna počtu polí ve III. oproti II. časové vrstvě). Zvýrazněny druhy Moneses uniflora, Agrostemma githago a Pyrola media. Čtyři extrémní záporné hodnoty nejsou nezobrazeny. Velikost bodu ukazuje počet druhů spadajících do jednoho místa grafu.Graph 4. Rate of decrease of 283 analysed critically endangered species in relation to their rarity or abundance in the past. The × axis has a logarithmic scale. Axis × = number of occupied grid fields in the time period II, axis y = parameter U2 (proportional change in the number of occupied grid fields in the III. time period contrary to the II. time period. Marked species Moneses uniflora, Agrostemma githago and Pyrola media. Four extreme negative values are not displayed. The size of each point represents a number of species sharing one place in the graph.
Pro tři vybrané C1 druhy jako příklady ilustrující různé typy změn rozšíření druhů v čase byly nakresleny síťové mapy. Příkladem druhu, který v průběhu 20. století výrazně ustoupil, je Radiola linoides (mapa 1). Údaje o rozšíření tohoto druhu v minulosti jsme kromě literatury získávali především z herbářových sbírek. Radiola linoides je druh vlhkých písčitých polí a obnažených den rybníků, který měl ještě v první polovině 20. století roztroušený výskyt po celém území České republiky. V průběhu druhé poloviny 20. století však tento konkurenčně slabý a do jisté míry na lidské aktivity vázaný druh značně ustoupil až na pouhá dvě síťová pole výskytu v současnosti (v okolí Plzně a Veselí nad Lužnicí).

291
Příroda, Praha, 31: 281–298, 2011
K tak silnému ústupu druhu pravděpodobně přispěly změny hospodaření v krajině (např. rušení malých políček, rapidní snížení počtu letněných rybníků).
Jako příklad druhu s relativně stabilním rozšířením uvádíme druh Serratula lycopifolia (mapa 2), který je svým výskytem v České republice vázaný jen na panonskou část jižní Moravy a území Bílých Karpat. Stanovišti jeho výskytu jsou výhřevné travnaté svahy a lesostepní stráně, pro které je vhodné pravidelné jednoroční kosení míst s výskytem druhu a občasné odstraňování náletových dřevin. Tento za květu velmi dekorativní druh je od r. 2000 na svých lokalitách pravidelně monitorován (jedná se o druh soustavy NATURA 2000). O jeho lokality je v současné době dobře pečováno – bělokarpatské populace jsou mimořádně bohaté a nalézají se na plochách mnoha arů (http://www.biomonitoring.cz/).
Posledním druhem, jehož rozšíření v tomto příspěvku ilustrujeme, je Trichomanes speciosum, rovněž druh soustavy NATURA 2000 (mapa 3). Jedná se spíše o speciální případ druhu, jehož počet lokalit v současné době rapidně narůstá. V České republice byl totiž tento druh kapradiny až do devadesátých let minulého století přehlížen, neboť se na našem území vyskytuje jen ve formě vláknitého, nenápadného gametofytu rostoucího ve vlhkých, hlubokých a velmi temných štěrbinách při úpatí skal a jeskyní, který se v našich podmínkách generativně nemnoží (nevytváří sporofyty). Od r. 2001 probíhá intenzivní mapování a monitoring lokalit tohoto druhu, a tak máme v současné době k dispozici kvalitní data o aktuálním stavu jeho rozšíření na území Českého Švýcarska, Dokeska, Kokořínska, Prachovských skal a dalších pískovcových lokalit České křídové tabule.
diskuse
V této práci bylo na základě sestrojených síťových map pro tři časová období analyzováno celkem 283 (resp. 268) C1 druhů. Výsledky ukazují, že v analyzovaném souboru dat spadá většina druhů do kategorie druhů v různém stupni ubývajících. Druhy výrazně ubývající (počet jimi obsazených síťových polí klesl o více než 50 %) se ve větší míře objevují až při zkoumání rozdílů v rozšíření mezi II. (1950–1999) a III. (po r. 2000) časovou periodou, začaly tedy více ubývat v průběhu 20. století. Je pravděpodobné, že velká část druhů tímto úbytkem reagovala na rychlé zhoršení životního prostředí a změny hospodaření v krajině, ke kterým v průběhu 20. století docházelo.
Ve skupině druhů ubývajících můžeme nalézt druhy vzácné již v minulosti, s výskytem jen v několika málo síťových polích. Při hodnocení změn rozšíření u takto vzácných druhů je však třeba mít na paměti, že parametry „U“ představují procentický úbytek obsazených polí a nezohledňují jejich celkový počet. Jedno zaniklé pole výskytu či nově obsazené pole (z důvodu nově nalezené lokality) se pak ve výsledku může projevit jako nárůst či úbytek o sto procent. Tento nedostatek byl zohledněn vynesením parametrů U proti skutečnému počtu obsazených síťových polí v grafech 3 a 4. Mezi ubývajícími druhy ale nalezneme i druhy v minulosti hojné (s vysokým počtem obsazených polí), které do dnešní doby výrazně ustoupily, nejčastěji z důvodu rychlého mizení vhodných stanovišť. Nalezli bychom mezi nimi teplomilné plevele a ruderální archeofyty, druhy vázané na biotopy silně ovlivněné člověkem či druhy vázané na lidské aktivity (Agrostemma githago, Atriplex rosea, Galium tricornutum, Marrubium vulgare a mnoho dalších). Z důvodu razantního úbytku lze takové druhy dnes považovat v rámci ČR za ohrožené.
Poměrně velkou část tvoří druhy stabilní, u nichž se počet obsazených síťových polí vůbec nezměnil nebo změnil jen velmi málo. Většinou se jedná o druhy již v minulosti velmi vzácné, které okupují jednu či několik málo známých lokalit výskytu, např. dru hy al pinského bezlesí (C1 druhy Krkonoš a Hrubého Jeseníku – např. Minuartia
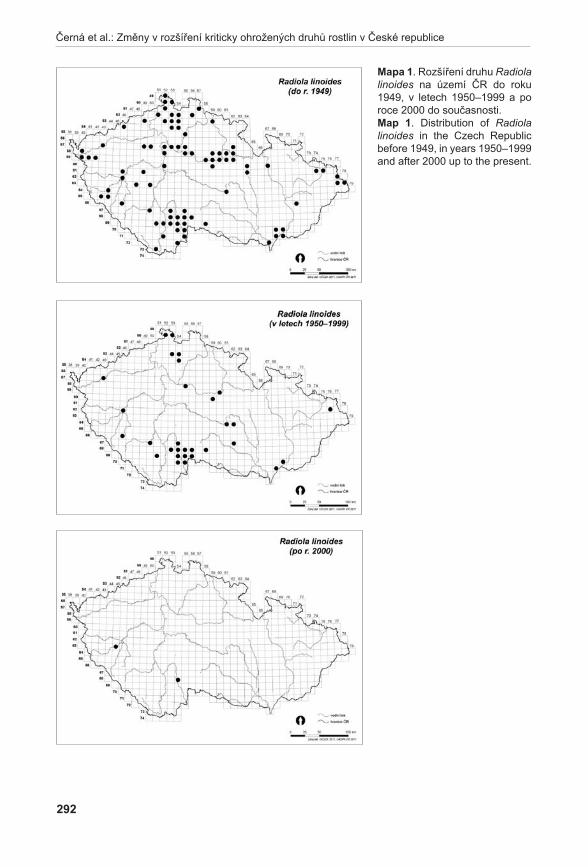
292
Černá et al.: Změny v rozšíření kriticky ohrožených druhů rostlin v České republice
Mapa 1. Rozšíření druhu Radiola linoides na území ČR do roku 1949, v letech 1950–1999 a po roce 2000 do současnosti.Map 1. Distribution of Radiola linoides in the Czech Republic before 1949, in years 1950–1999 and after 2000 up to the present.
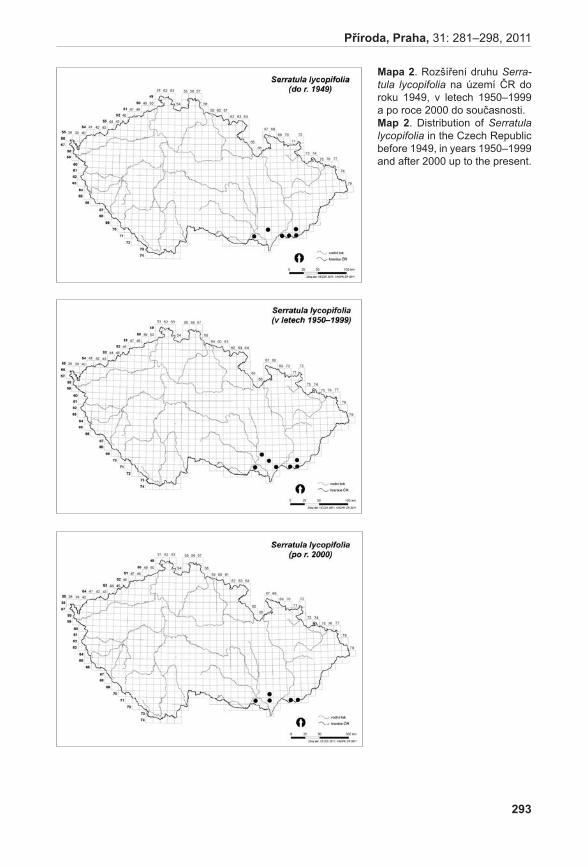
293
Příroda, Praha, 31: 281–298, 2011
Mapa 2. Rozšíření druhu Se rra-tula lycopifolia na území ČR do roku 1949, v letech 1950–1999 a po roce 2000 do současnosti.Map 2. Distribution of Serratula lycopifolia in the Czech Republic before 1949, in years 1950–1999 and after 2000 up to the present.

294
Černá et al.: Změny v rozšíření kriticky ohrožených druhů rostlin v České republice
Mapa 3. Rozšíření druhu Trichomanes speciosum na území ČR v letech 1990–2000 a po roce 2000 do současnosti.Map 3. Distribution of Trichomanes speciosum in the Czech Republic in years 1990–1999 and after 2000 up to the present.
corcontica, Hedysarum hedysaroides), endemity (Dianthus arenarius subsp. bohemicus, Campanula gelida) či druhy, pro které na území ČR existuje jen omezené množství vhodných biotopů (např. Isoëtes lacustris a I. echinospora na dně ledovcových jezer na Šumavě). Vzhledem k výskytu těchto druhů na dobře známých, často chráněných a monitorovaných lokalitách či nedostupných místech (ledovcové kary apod.) by se mohlo zdát jejich zařazení na červený seznam do kategorie kriticky ohrožený poněkud nadhodnocené. Na druhou stranu právě výskyt na jediné lokalitě (či několika málo lokalitách) z nich dělá druhy velmi zranitelné a zařazení takových druhů na červený seznam tak alespoň upozorňuje na jejich jedinečnost. Mezi stabilními druhy příliš neměnícími počty obsazených polí bychom dále mohli nalézt lesní druhy (např. Epipactis microphylla či již v minulosti hojný druh Pyrola media).

295
Příroda, Praha, 31: 281–298, 2011
Druhy, které v čase přibývají, tvoří zvláštní skupinu. V drtivé většině se jedná o druhy, které byly v minulosti uváděny jako velmi vzácné. Z našich analýz je zřejmé, že se často jedná o druhy, které původně nebyly rozlišované (rod Epipactis), byly přehlížené (např. již zmíněný Trichomanes speciosum) či jsou to druhy, které byly v minulosti naprosto běžné, takže jim z hlediska záznamu výskytu nebyla původně věnována tak velká pozornost, a to do té doby, než byl zaznamenán jejich rapidní úbytek. To je případ druhu Agrostemma githago, který dle našich analýz při přechodu z I. do II. časové vrstvy spadá do kategorie druhů přibývajících, zato když srovnáme stav jeho výskytu v druhé polovině 20. století s dnešním (po roce 2000), jedná se o druh, který z přírody České republiky téměř vymizel.
Námi provedené analýzy poskytují základní představu o typech změn v rozšíření vybraných C1 rostlin v čase a zhruba nastiňují procentuální zastoupení druhů v jednotlivých kategoriích. Uvědomujeme si však, že vzhledem k vybraným metodickým postupům sběru a zpracování dat mohou být naše výsledky zkresleny. Již při samotném sběru dat, zvlášť v případě herbářových sběrů, jsme naráželi na problém nedostatku historických údajů u druhů v minulosti naprosto běžných nebo „esteticky nezajímavých“. Lokality takových v minulosti hojných druhů byly velmi široce popsané, občas nebylo možné tyto nálezy do konkrétního pole mapovací sítě jasně zakreslit. Někdy jsme zvolili postup zakreslení široce popsané lokality do síťového pole s nejnižším číslem (viz Metodika), čímž jsme mohli uměle vytvořit nově obsazené pole, jindy musela být nepřesně popsaná lokalita z analýz vyloučena, čímž jsme se možná o některé lokality, které jsme nedokázali blíže specifikovat, připravili. V analýzách pak v kombinaci s aktuálními přesnějšími daty (získanými např. při mapování vegetace) mohlo snadno dojít k tomu, že byly některé druhy chybně vyhodnoceny jako druhy přibývající či ubývající. Zvláštní opatrnosti je třeba dbát při rekonstrukci rozšíření druhů v minulosti velmi vzácných, kde pouhá chyba o jedinou lokalitu může způsobit nežádoucí zkreslení výsledků.
Vzhledem k tomu, že část dat byla na základě zmíněné metodiky uměle datována, mohlo někdy dojít k zařazení nálezu do nesprávné časové vrstvy – na tento problém jsme nejčastěji naráželi při rozhodování o datování nálezu do I. či II. časové vrstvy. Avšak při porovnávání změn rozšíření čistě minulost vs. přítomnost (tj. stav před a po r. 2000) nám případné chybné časové určení výsledky významně nezkresluje. Právě vygenerováním předběžných map a jejich kontrolou před samotnými analýzami jsme se snažili případné chyby odstranit.
Ideální by samozřejmě bylo získat kvalitní nálezová data pro všechny C1 druhy zahrnuté do tohoto projektu a analýzy úbytků druhů rozšířit. V této práci se nám však podařilo získat relevantní údaje pro 283 z nich a i přes výše uvedené nedostatky jsme přesvědčeni, že naše studie nabízí zajímavý pohled na tuto velmi různorodou skupinu kriticky ohrožených druhů rostlin a poskytuje základní představu o jejich chování v prostoru a čase. Následným propojením s biologickými vlastnostmi a ekologickými nároky, které byly studovány v jiných částech projektu (Gabrielová et al. 2011), pak bude možné snáze identifikovat příčiny poklesu populací některých C1 druhů a lépe pochopit jejich reakce na změny životního prostředí.
shrnutí
Na základě sběru nálezových dat z odborné literatury, herbářových sbírek a floristických databází bylo shromážděno velké množství údajů o historickém i současném rozšíření kriticky ohrožených druhů rostlin České republiky. Pomocí síťových map sestrojených pro tři

296
Černá et al.: Změny v rozšíření kriticky ohrožených druhů rostlin v České republice
stanovená časová období byla k 283 vybraným C1 druhům získána informace o změnách jejich rozšíření v průběhu posledních 100 let. Z výsledků analýz síťových map vyplývá, že mezi kriticky ohroženými druhy mají největší zastoupení druhy v různém stupni ubývající, s větší intenzitou ubývání v druhé polovině 20. století. Nalezneme však mezi nimi i druhy v minulosti vzácné s víceméně stabilním počtem obsazených polí v čase i druhy v minulosti hojné, které do současné doby výrazně ustoupily. Druhy v minulosti zřejmě vzácné, které se v současné době začaly šířit, jsou zastoupeny jen okrajově.
Výsledky této práce, přehledně upořádané v Nálezové databázi ochrany přírody i ve veřejně přístupné Databázi charakteristik C1 rostlin (http://portal.nature.cz/c1/rostliny), se stanou nezbytným podkladem pro výběr druhů do záchranných programů a jistě naleznou praktické využití při běžném hodnocení a rozhodování o prioritách ochrany přírody.
summary
Based on data from literature, herbaria, and flora databases we summarized information about historical and actual distribution of critically endangered species of the Czech Republic. Using grid distribution maps for three time periods, we derived information on changes in the distribution of 283 critically endangered species over the last 100 years. The analyses of the grid maps show that most analysed species are decreasing, with a more intensive decrease in the second half of the 20th century. Nevertheless, we can find among them also species rare in the past with a more or less stabile number of occupied grid fields over time as well as species which were relatively abundant in the past but have decreased till the present. The species which were rare in the past but are spreading now are represented marginally.The results of this work are deposited in the database of the Agency for Nature Conservation and Landscape Protection and are also accessible to public (http://portal.nature.cz/c1/rostliny). The information will form a useful background for evaluating priorities for species conservation.
Poděkování
Za konzultace a podnětné připomínky velmi děkujeme Jitce Horáčkové, Vítu Grulichovi, Karlovi Kubátovi, Jiřímu Danihelkovi a Jiřímu Hadincovi. Velký dík patří Milanu Markovi za překlad obtížně čitelných sched. Anně Šmídové děkujeme za koordinaci zpracování Dominova materiálu. Velmi děkujeme též všem botanikům regionálních pracovišť AOPK ČR a správy KRNAP za poskytnuté informace o aktuálních výskytech C1 rostlin, dále osloveným autorům Květeny ČR za poskytnutá nálezová data k vybraným C1 druhům a kurátorům navštívených a oslovených herbářových sbírek za ochotu se do této studie zapojit. V neposlední řadě pak děkujeme všem studentům a brigádníkům za pomoc při náročných úpravách a zpracování nálezových dat a Zdeňkovi Kučerovi za trpělivost a přípravu dat k importu. Za recenzní připomínky děkujeme Petru Petříkovi. Tato studie byla finančně podpořena grantem MŠMT VaV 2B06178. Částečnou podporu poskytly také MSM 0021620828 a AV0Z6005016

297
Příroda, Praha, 31: 281–298, 2011
literatura
Čeřovský j., feráková v., holub j., MaGlocký Š. & Procházka f. [eds] (1999): Červená kniha ohrožených a vzácných druhů rostlin a živočichů ČR a SR. Vol. 5. Vyšší rostliny. – Príroda Bratislava.
Dostál j. (1989): Nová květena ČSSR 1, 2. – Academia, Praha.Gabrielová j., Černá l. & MünzberGová z. (2011): Jak souvisí změny v rozšíření kriticky
ohrožených druhů rostlin České republiky s jejich biologickými vlastnostmi a ekologickými nároky? – Příroda 31: 345–370.
Grulich v. (1997): Atlas rozšíření cévnatých rostlin Národního parku Podyjí. – Masarykova univerzita, Brno.
hejný s. & slavík b. [eds] (1988): Květena České socialistické republiky. Vol. 1. – Academia, Praha. hejný s. & slavík b. [eds] (1990): Květena České republiky. Vol. 2. – Academia, Praha. hejný s. & slavík b. [eds] (1992): Květena České republiky. Vol. 3. – Academia, Praha.henDrych r. & jirásek v. (2002): Příručka pro práci s latinsky psanými herbářovými schedami. –
Zpr. Čes. Bot. Společ., suppl. 2002/2: 1–131.jonGePier j. w.& Pechanec v. (2006): Atlas rozšíření cévnatých rostlin CHKO Bílé Karpaty. – ZO
ČSOP Bílé Karpaty, Veselí nad Moravou.kláŠterský i., hrabětová-uhrová a. & DuDa j. (1982): Dějiny floristického výzkumu v Čechách,
na Moravě a ve Slezsku I a II. – Severočes. Přír., suppl. 1982/1 (1, 2): 1–242.kolektiv autorů (1952–1999): Bibliographia botanica Čechoslovaca (Čechica). – Botanický
ústav ČSAV, Průhonice.kubát k. (1986): Červená kniha vyšších rostlin Severočeského kraje. – Praha.kubát k., hrouDa l., chrtek j. jun., kaPlan z., kirschner j. & ŠtěPánek j. [eds] (2002): Klíč ke
květeně České republiky. – Academia, Praha.iucn (2001): IUCN Red List categories and criteria: Version 3.1. IUCN Species Survival
Commision. – IUCN, Gland, Switzerland and Cambridge, U. K.Marhoul P., brejŠková l., cePáková e. & volf o. (2003): Záchranné programy živočichů v ČR.
Jak dál? – Ochr. Přír., Praha, 58/6: 175–179.Pfohl e. (1931): Orientierungslexicon der Tschechoslowakischen Republik. Dritte Auflage. –
Reichenberg (Liberec).Procházka f. [ed.] (2001): Černý a červený seznam cévnatých rostlin České republiky (stav
v roce 2000). – Příroda, Praha, 18: 1–166.slavík b. (1986): Fytokartografické syntézy ČSR. Vol. 1. – Průhonice.slavík b. (1990): Fytokartografické syntézy ČSR. Vol. 2. – Průhonice.slavík b. (1998): Phytocartographical syntheses of the Czech Republic. Vol. 3. – Praha.slavík b. [ed.] (1995): Květena České republiky. Vol. 4. – Academia, Praha.slavík b. [ed.] (1997): Květena České republiky. Vol. 5. – Academia, Praha.slavík b. [ed.] (2000): Květena České republiky. Vol. 6. – Academia, Praha.slavík b. & ŠtěPánková j. [eds] (2004): Květena České republiky. Vol. 7. – Academia, Praha.soják j. (2007): Latinsko-český botanický slovník. – Zpr. Čes. Bot. Společ. 42, suppl. 2007/1:
1–167.Šourek j. (1969): Květena Krkonoš. Český a polský Krkonošský národní park. – Academia,
Praha.ŠtěPánková j. [ed.] (2010): Květena České republiky. Vol. 8. – Academia, Praha.vaněČek j. (1969): Květena Horažďovicka. Materiál k floristickému výzkumu Horažďovicka. –
Plzeň.vozárová M. & sutorý k. (2001): Index herbariorum Reipublicae bohemicae et Reipublicae
slovacae. – Zpr. Čes. Bot. Společ. 36, suppl. 2001/1: 1–95.

298
Černá et al.: Změny v rozšíření kriticky ohrožených druhů rostlin v České republice
Adenophora liliifoliaAposeris foetidaAster alpinusAstragalus arenariusBupleurum affineBupleurum rotundifoliumBupleurum tenuissimumCeratophyllum submersumCimicifuga europaeaCirsium brachycephalumConioselinum tataricumDaphne cneorumDrosera anglicaDrosera intermediaEchium russicumElatine alsinastrumEpipogium aphyllumEquisetum variegatumEryngium planumGentiana punctataGlaux maritimaHammarbya paludosaHippuris vulgarisChimaphila umbellataJuncus atratusJuncus capitatusJuncus gerardiiJuncus sphaerocarpusJuncus subnodulosusJuncus tenageiaLaserpitium archangelicaLathyrus aphacaLathyrus heterophyllusLathyrus hirsutusLathyrus nissoliaLathyrus palustrisLeucojum aestivumLinaria arvensisLindernia procumbensLinum hirsutum subsp. hirsutumListera cordataLuronium natansMalaxis monophyllosMoneses unifloraNuphar pumilaOrchis palustrisOrchis tridentataPeucedanum carvifoliaPlantago maritima subsp. ciliataPyrola chloranthaPyrola media
Radiola linoidesReseda phyteumaRhodiola roseaSamolus valerandiiScorzonera parvifloraScrophularia vernalisSenecio paludosusSerratula lycopifoliaScheuchzeria palustrisTillaea aquaticaTofieldia calyculataTordylium maximumTrapa natansTriglochin maritimumTrinia glaucaVeratrum nigrum
Příloha 1. seznam druhů excerpovaných v herbářových sbírkách ČR.

299
Příroda, Praha, 31: 299–343, 2011
Jak je to s ohrožením kriticky ohrožených druhů rostlin České republiky v jiných evropských zemích?
critically endangered plant species of the czech Republic: what is the situation of their endangerment in other European countries?
JarMiLa gabrieLová1, tereZa fiaLová1 & ZuZana MünZbergová1, 2
1Katedra botaniky, Přírodovědecká fakulta Univerzity Karlovy, Benátská 2, CZ-128 01 Praha 2; [email protected]; [email protected]; [email protected] 2Botanický ústav Akademie věd České republiky, v. v. i., Zámek 1, CZ-252 43 Průhonice
Abstract: To set priorities in species conservation in a given country it is important to know the situation of the species endangerment in the surrounding countries. From the conservation point of view, more attention should be paid to species largely endangered throughout their distributional range rather than to the species which are endangered in a particular country but still common in the surrounding countries. We investigated the group of critically endangered vascular plant species of the Czech Republic (C1 species) in order to find out what is their conservation status in other European countries. The threat status of 483 C1 species was excerpted from Red Lists and Red Data Books of 31 European countries, the presence or absence data were found in checklists or floras of the respective countries. For each country, we calculated the proportion of C1 species, which are endangered, not endangered (common) or not occuring in the territory. C1 species are relatively common in Italy, France, Ukraine, Croatia, and Romania, however they are considerably endangered in the neighbouring countries – Slovakia, Austria, and Germany. Only 12% of the C1 species (mostly endemic species) are not common in any European country and more than 50% of the C1 species are common at least in seven European countries. The results show that a high number of C1 species occur in other European countries and are quite common there. However, there still remain many species for the conservation of which the Czech Republic is highly responsible.Keywords: Central Europe, endemic species, rare species, red list, risk of extinction, species conservation
Abstrakt: Při výběru druhů určených pro prioritní ochranu v České republice je důležité zohlednit situaci ohrožení těchto druhů v rámci areálu jejich rozšíření. Ochrana druhů, které jsou ohroženy v rámci celého svého areálu, je z hlediska zachování biodiverzity důležitější než ochrana druhů, které jsou v České republice vzácné, ale v ostatních evropských zemích celkem běžné. V našem příspěvku jsme se zaměřili na skupinu kriticky ohrožených druhů rostlin České republiky (kategorie C1 sensu Procházka 2001), pro které jsme zjišťovali, jaká je situace jejich ohrožení v ostatních evropských zemích. Informaci o statutu ohrožení v 31 evropských státech jsme získali z červených seznamů a červených knih daných zemí, přítomnost/nepřítomnost druhu v dané zemi jsme ověřovali pomocí checklistů nebo flór. Pro každý stát jsme stanovili proporci českých C1 druhů, které jsou na jeho území ohrožené, neohrožené (běžné) a které se na jeho území nevyskytují a pro každý druh jsme stanovili v kolika zemích je ohrožený a v kolika zemích běžný. České C1 druhy jsou relativně běžné v Itálii, Francii, Chorvatsku, Rumunsku a na Ukrajině, naopak jsou značně ohrožené ve státech sousedících s Českou republikou – na Slovensku, v Rakousku a Německu. Pouze

300
Gabrielová et al.: Ohrožení C1 rostlin ČR v jiných evropských zemích?
12 % českých C1 druhů (většinou endemické druhy) není běžných v žádné jiné evropské zemi a více než 50 % českých C1 druhů je ohrožených alespoň v sedmi evropských zemích. Z výsledků vyplývá, že poměrně velká část českých C1 druhů se vyskytuje i v jiných evropských zemích a je zde relativně hojná. Přesto zde stále zbývá řada druhů, za jejichž ochranu má Česká republika značnou zodpovědnost.Klíčová slova: červený seznam, druhová ochrana, endemit, riziko vyhynutí, střední Evropa, vzácné druhy nomenklatura: Procházka (2001)
Úvod
V souvislosti se zvětšujícím se počtem druhů ohrožených lidskou činností a ztrátou vhodných stanovišť roste i úsilí tento proces dokumentovat a zvrátit. Užitečné nástroje v druhové ochraně, které mapují pokles biodiverzity a upozorňují na nutnost ochranářských opatření, jsou červené seznamy vzácných a ohrožených druhů. Pro objektivní hodnocení ohrožení druhů podle nebezpečí jejich vyhynutí nebo vyhubení na celosvětové úrovni vypracovala Mezinárodní unie pro ochranu přírody (International Union for Conservation of Nature, IUCN) seznam kategorií a kritérií, na jejichž základě mohou být červené seznamy ohrožených druhů sestavovány. Ohroženost taxonu se odhaduje podle pěti kritérií, která jsou založena na znalostech oblasti výskytu (extent of occurrence), oblasti osídlení (area of occupancy) nebo početnosti populací a jejich známém či očekávaném úbytku v určitém časovém období, případně na znalosti pravděpodobnosti vyhynutí nebo vyhubení ve volné přírodě (hartley & kunin 2003).
Tato kritéria byla vytvořena primárně pro hodnocení ohroženosti na celosvětové úrovni. Pro hodnocení na úrovni nižší než je globální, je nutné daná kritéria poněkud modifikovat (GärDenfors 2001, GärDenfors et al. 2001, iucn 2003, keller et al. 2005). V předběžném hodnocení se nejprve kritéria celosvětového červeného seznamu IUCN (iucn 2001, poslední směrnice pro praktickou aplikaci iucn 2010) použijí na populaci hodnoceného taxonu v dané části světa. Ve druhé fázi je prozkoumána existence a stav konspecifických populací v okolí dané části světa, které mohou ovlivnit riziko extinkce hodnoceného taxonu. Zohlednění stavu okolních populací vede většinou ke snížení kategorie odpovídající celosvětovým kritériím, protože populace v dané části světa může být posilována imigrací z okolních oblastí, čímž se zmenšuje riziko extinkce (iucn 2003).
Objektivní hodnocení na základě jednotných kritérií má tu výhodu, že umožňuje porovnání stupně ohrožení téhož taxonu v různých částech jeho areálu (GärDenfors et al. 2001). Na druhou stranu vyžaduje informace, které nemusí být pro celou řadu druhů v dané části světa dostupné (De GraMMont & cuarón 2005). Primárním záměrem červených seznamů, sestavovaných na základě metodiky IUCN, je navíc pouze objektivně vyhodnotit riziko vyhynutí (vyhubení) taxonu; v případě národních seznamů pak riziko, že druh vyhyne v daném regionu (keller & bollMann 2004). Národní červené seznamy jsou však často sestavovány ještě za jiným účelem – mají sloužit při výběru druhů pro prioritní ochranu (PossinGhaM et al. 2002). Při stanovování priorit druhové ochrany je riziko extinkce pouze jedním z mnoha faktorů, které je třeba zohlednit (roDriGues et al. 2006). Neméně důležitý je globální význam populace taxonu určitého státu a status ohrožení druhu v okolních zemích (GärDenfors 2001). Dále je třeba zohlednit taxonomickou, genetickou či ekologickou jedinečnost taxonu, oporu v legislativě, logistické a ekonomické faktory (typ ochranářského opatření, podloženost adekvátními daty, jeho nutnost a proveditelnost, finanční náklady a případné finanční ztráty jakožto důsledek ochrany) (Miller et al. 2007). Proto si mnoho

301
Příroda, Praha, 31: 299–343, 2011
zemí vytváří své vlastní seznamy vzácných a ohrožených druhů, které jsou založeny na jiných kategoriích a kritériích nebo na základě odhadu odborníků, bez použití nějakého systematického hodnocení.
Rovněž Červený seznam cévnatých rostlin České republiky (holub & Procházka 2000 a jeho upravená verze Procházka 2001, se kterou pracujeme v tomto příspěvku) nebyl sestavován podle metodiky IUCN. Je založen na „středoevropské“ stupnici ohrožení (holub 1996), která je v současné době do značné míry ztotožnitelná s mezinárodně používanými kategoriemi. Rozlišuje následující kategorie: A vyhynulé a nezvěstné taxony (= EX), která je rozdělená na podskupiny A1 vyhynulé, A2 nezvěstné (pravděpodobně vyhynulé) a A3 nejasné případy vyhynulých a nezvěstných, C1 kriticky ohrožené (= CR), C2 silně ohrožené (= EN), C3 ohrožené (= VU), C4a vzácnější taxony vyžadující další pozornost – méně ohrožené (= LR) a C4b vzácnější taxony vyžadující další pozornost – dosud nedostatečně prostudované (DD). Seznam byl sestavován na základě kritérií, která kromě početnosti populací a charakteru ústupu zahrnují i faktory jako úzká vazba na typ stanoviště, biologické vlastnosti taxonu (vitalita, reprodukční poměry, trvání ontogeneze, škůdci, speciální opylovači), jeho atraktivnost a současný stav ochrany nalezišť. Měl by sloužit jako jeden z teoretických podkladů pro aktivní ochranu fytogenofondu a jako základní dokument pro stanovování priorit v druhové ochraně rostlin (Procházka 2001). Nevýhodou je, že obsahuje jen nekomentovaný výčet druhů, takže není jasné, na základě jakých kritérií byl taxon do příslušné kategorie ohrožení zařazen. Zahrnuje 1623 druhů a subspecií, což představuje více než 60 % druhového bohatství souboru české flóry a jen kategorie „kriticky ohrožené“ (C1) obsahuje více než 470 taxonů.
Kategorie C1 zahrnuje taxony, kterým na našem území hrozí akutní riziko vyhynutí. Není ovšem možné věnovat patřičnou pozornost všem těmto druhům. Už podle kritérií, na základě kterých jsou druhy do seznamu zařazovány, je jasné, že se jedná o poměrně heterogenní skupinu. Najdeme zde druhy přirozeně vzácné, druhy ubývající v důsledku negativních dopadů lidské činnosti, taxony, jež jsou vzácné a jejichž početnost kolísá v důsledku toho, že jsou na okraji areálu rozšíření, a samozřejmě druhy, jež jsou vzácné či ubývají z jiných (často neznámých) příčin.
Při výběru druhů určených pro prioritní ochranu by mělo být zohledněno široké spektrum aspektů. Kromě dalšího je důležité znát situaci ohrožení druhu v dalších částech areálu rozšíření. Z hlediska udržení celkové biodiverzity by měla mít přednost ochrana druhů endemických pro daný stát či větší území, jehož je stát součástí, ochrana druhů ohrožených v rámci celého svého areálu rozšíření či ochrana izolovaných arel druhu na okrajích souvislého areálu rozšíření, protože izolace představuje jeden z důležitých speciačních mechanismů a izolované populace se mohou vyvinout v nový druh (schnittler & Günther 1999). Menší pozornost pak může být věnována druhům, které jsou sice v dané zemi vzácné, ale běžné v řadě dalších zemích, ve kterých se vyskytují.
Stanovení druhů zasluhujících zvýšenou pozornost ze strany ochrany přírody není jen záležitost jednotlivých zemí, existuje řada mezinárodně uznávaných dokumentů i nezávazných seznamů, které hodnotí důležitost ochrany druhů z celoevropského hlediska. Jedním z nejstarších dokumentů je Úmluva o ochraně evropských planě rostoucích rostlin, volně žijících živočichů a přírodních stanovišť (council of euroPe 1979), známá též jako Bernská úmluva (Příloha I obsahuje seznam přísně chráněných druhů rostlin, jejichž ochrana vyžaduje celoevropskou spolupráci). Pod záštitou Rady Evropy brzy vznikla další, rozšířená verze seznamu celoevropsky ohrožených druhů (council of euroPe 1983) a zatím poslední verzi seznamu, který už představuje plnohodnotný pokus o evropský červený seznam ohrožených druhů rostlin, zveřejnila Rada Evropy v roce 2001 (council of euroPe 2001). Evropský červený seznam ohrožených druhů rostlin stále čeká na dokončení, původně měl

302
Gabrielová et al.: Ohrožení C1 rostlin ČR v jiných evropských zemích?
být zveřejněn počátkem roku 2011 (http://ec.europa.eu/environment/nature/conservation/species/redlist/vascular_plants.htm). Nezávisle na aktivitě koordinované Radou Evropy vznikl dokument, hodnotící ohrožení evropských rostlin a úlohu botanických zahrad v jejich ochraně, jehož součástí je i seznam evropsky ohrožených druhů, sestavený excerpcí vybraných národních červených seznamů a červených knih (sharrock & jones 2009). Pro střední Evropu sestavili seznam rostlinných druhů vyžadujících prioritní ochranářská opatření schnittler & Günther (1999). Opomenuta by neměla být Směrnice Rady (92/43/EHS) o ochraně přírodních stanovišť volně žijících živočichů a planě rostoucích rostlin, resp. seznamy druhů uvedených v příloze II, IV a V tohoto dokumentu. Červený seznam celosvětově ohrožených druhů rostlin a živočichů sestavuje IUCN, poslední knižní verze vyšla v roce 1998 (walter & Gillett 1998) a v současné době je na internetu k dispozici aktualizovaná elektronická verze (iucn 2011).
V našem příspěvku se zaměřujeme na analýzu situace ohrožení českých C1 druhů v ostatních evropských státech s cílem identifikovat druhy, které jsou z celoevropského hlediska nejohroženější a jejichž ochrana by měla být prioritou. Přitom si klademe následující otázky: 1) Jaký je status ohrožení českých C1 druhů v dalších evropských zemích? 2) V jakých dalších evropských zemích jsou české C1 druhy rovněž ohrožené a Česká republika s nimi tedy sdílí podobné ochranářské priority? V jakých dalších evropských zemích jsou české C1 druhy naopak běžné? 3) Které z našich C1 druhů jsou z celoevropského hlediska ohrožené a které naopak relativně neohrožené? 4) Je nějaký rozdíl v míře celoevropského ohrožení mezi českými C1 druhy zahrnutými a nezahrnutými do Seznamu středoevropských prioritních druhů (schnittler & Günther 1999), resp. chráněnými a nechráněnými podle Vyhlášky MŽP 395/1992 Sb.? 5) Existuje nějaký vztah mezi počtem evropských zemí, ve kterých jsou české C1 druhy ohrožené/běžné a typem stanoviště, na kterém se druhy vyskytují?
Metodika
sběr dat
Pro skupinu kriticky ohrožených druhů (kategorie C1 sensu Procházka 2001) jsme zjišťovali status ohrožení v 31 evropských zemích. Jednalo se o následující státy: Albánie, Belgie, Bulharsko, Dánsko, Estonsko, Finsko, Francie, Chorvatsko, Island, Irsko, Itálie, Německo, Maďarsko, Litva, Lotyšsko, Lucembursko, Malta, Nizozemí, Norsko, Polsko, Portugalsko, Rakousko, Rumunsko, Řecko, Slovensko, Slovinsko, Španělsko, Švédsko, Švýcarsko, Ukrajina a Velká Británie. Status ohrožení jsme vyhledávali v červených seznamech, červených knihách a seznamech druhů chráněných zákonem těchto zemí (viz. příloha 1 a literatura). Upředňostňovali jsme aktuální a úplné verze červených seznamů před staršími verzemi seznamů a červenými knihami. Ne vždy se však podařilo nejaktuálnější seznam získat, např. v případě Itálie jsme excerpovali starší červený seznam (conti et al. 1997), přičemž existuje i novější verze (PiGnatti et al. 2001), která je v současnosti nadále aktualizována. V případě Bulharska je červená kniha a červený seznam, který má být její součástí, stále ve stádiu příprav, proto jsme vycházeli z pracovní verze, která zdaleka není úplná. Rovněž francouzská národní červená kniha z roku 1995 (olivier et al. 1995) je odbornými kruhy považována za zastaralou a neúplnou. Proto Francouzský národní komitét IUCN (UICN Comité Français) spolu s Národním přírodovědeckým muzeem (La Muséum national dxHistoire naturelle) přikročily v roce 2007 k vydávání nového červeného seznamu La Liste rouge des espèces menacées en France, ze kterého byl zatím vydán první díl Orchidées de France métropolitaine (uicn france et al. 2010, J. Čeřovský, in

303
Příroda, Praha, 31: 299–343, 2011
litt.). Také v Itálii se v současné době pracuje na aktualizaci červeného seznamu, ohrožení druhů je vyhodnocováno na základě kritérií IUCN a seznamy nově vyhodnocených taxonů jsou postupně zveřejňovány v periodiku Informatore Botanico Italiano (G. Rossi, in litt.). Z výše uvedených příkladů je zřejmé, že sestavování a zveřejňování červených seznamů a červených knih je permanentní proces, takže jakákoli souhrnná práce v této oblasti velmi rychle zastarává. Excerpce dat pro náš příspěvek byla ukončena ke dni 31. 12. 2009.
Vzhledem k taxonomickým nejasnostem jsme v případě následujících druhů pracovali na druhové úrovni a nerozlišovali jednotlivé subspecie: Gentianella amarella s. l. namísto G. amarella subsp. amarella; Orchis ustulata s. l. namísto O. ustulata subsp. aestivalis a O. ustulata subsp. ustulata; Orobanche purpurea s. l. namísto O. purpurea subsp. bohe-mi ca a O. purpurea subsp. purpurea; Senecio paludosus s. l. namísto S. paludosus subsp. paludosus a některé druhy jsme zcela vyloučili z analýz (Bolboschoenus maritimus subsp. compactus – konkrétní druhy ze skupiny Bolboschoenus maritimus nejsou v různých zdrojích dobře rozlišovány, většinou k dispozici jen údaje o Bolboschoenus maritimus s. l.; Euphrasia picta – v souladu s novějšími poznatky spadá druh E. picta do variability druhu E. rostkoviana a není vylišován jako samostatný taxon, vnitrodruhová variabilita vyžaduje další studium (Smejkal & Dvořáková in slavík 2000); Gymnadenia conopsea subsp. montana – taxonomicko-morfologické hodnocení variability druhu G. conopsea komplikované, holotyp jména G. conopsea subsp. montana ztracen (Procházka et al. in ŠtěPánková 2010); Hieracium kablikianum – pravděpodobně zvláštní morfotyp bez ta xo nomické hodnoty, který představuje přechodný typ mezi skupinami H. fritzei a H. nigritum a vyžaduje další studium (chrtek & MarholD 1998; J. Chrtek, in litt.); Iris aphylla subsp. fieberii, Iris aphylla subsp. novakii – v pojetí Hrouda & Grulich in ŠtěPánková (2010) rozlišovány jen jako morfologické varianty v rámci druhu Iris aphylla; Phyteuma orbiculare subsp. flexuosum, Phyteuma orbiculare subsp. montanum – vnitrodruhová klasifikace druhu Phyteuma orbiculare vychází z monografie schulz (1904), ovšem v současném pojetí nejsou poddruhy většinou rozlišovány (J. Chrtek, in litt.); Plantago maritima subsp. ciliata – Plantago maritima je velmi variabilní druh s mnoha poddruhy, které nejsou v různých zemích dobře rozlišovány; Puccinelia limosa – na území ČR se tento druh s největší pravděpodobností nikdy nevyskytoval, vždy šlo o záměny se druhem P. distans (Moravcová et al. 2001) a Sorbus hardeggensis – dříve považován za druh hybridogenního původu, dnes považován za hybrida. Údaje o endemismu druhů byly doplněny podle Květeny ČR (hejný & slavík 1988, 1990, 1992, slavík 1995, 1997, 2000, slavík & ŠtěPánková 2004, ŠtěPánková et al. 2010) a seznamu M. Gerži (Gerža 2009).
Národní kategorie ohrožení v každé evropské zemi byly převedeny na modifikované kategorie IUCN (iucn 2001): EX – vyhynulý, EX? – pravděpodobně vyhynulý, CR – kriticky ohrožený, EN – ohrožený, VU – zranitelný, NT – téměř ohrožený, R – vzácný, I – nezařazený (druh spadající do některé z kategorií EX, CR, EN, VU nebo R, pro stanovení přesného stupně ohrožení však chybí data), LC/LR – méně ohrožený, DD – chybějící data, NE – nevyhodnocený. Při tomto procesu nebyl použit jednotný postup, protože různé země používají při vytváření vlastních kategorií velmi rozdílné metodiky. V některých případech byl převod na kategorie IUCN dostupný přímo v primárním zdroji. V ostatních případech jsme pracovali s názvy a definicemi národních kategorií dostupných v původním zdroji a stanovili jsme nejbližší kategorii IUCN odpovídající dané národní kategorii.
Rovněž jsme zjišťovali, jestli se taxon v dané zemi vyskytuje a není ohrožen nebo se v zemi nevyskytuje. Data o presenci/absenci druhů jsme primárně vyhledávali v databázi Flora Europaea (http://rbg-web2.rbge.org.uk/FE/fe.html). Tato databáze je ovšem už poměrně zastaralá a obsahuje řadu chyb, proto jsme se snažili informace doplnit z checklistů a v některých případech i aktuálních flór daných zemí (viz příloha 1 a literatura). V případě,

304
Gabrielová et al.: Ohrožení C1 rostlin ČR v jiných evropských zemích?
že checklist nebyl pro danou zemi k dispozici (Albánie, Irsko a Island), využili jsme pouze databázi Flora Europea. Pro druhy nezahrnuté do této databáze (např. některé druhy rodu Hieracium a Taraxacum) jsme použili informaci o areálu rozšíření z Květeny ČR (hejný & slavík 1988, 1990, 1992, slavík 1995, 1997, 2000, slavík & ŠtěPánková 2004, ŠtěPánková et al. 2010). Problém nastal u některých zemí bývalé Jugoslávie (Bosna a Herzegovina, Černá Hora, Makedonie a Srbsko) a bývalého SSSR (Bělorusko, Moldavsko, Rusko), pro které není k dispozici checklist flóry. I když pro většinu z nich (s výjimkou Černé Hory a Makedonie) existují červené seznamy nebo knihy (MaXiMovitch 1993, Postolache 1995, silic 1996, stevanoviČ 1999, anonyMus 2005) nepodařilo se nám pro tyto země získat spolehlivé údaje o prezenci/absenci C1 druhů, protože databáze Flora Europaea nerozlišuje jednotlivé státy dřívější Jugoslávie a SSSR. Dohledání výskytu všech C1 druhů ve flórách těchto zemí (často podobně jako v ČR mnohasvazková díla psaná národním jazykem) přesahovalo rozsah této studie, a proto jsme údaje o ohrožení druhů z těchto zemí nemohli zahrnout do dalších analýz.
Dále jsme zaznamenali, jestli je druh zahrnut do Seznamu středoevropských rostlin vyžadujících prioritní ochranářská opatření (schnittler & Günther 1999), Vyhlášky MŽP 395/1992 Sb. (příloha 3), Bernské úmluvy (council of euroPe 1979), Seznamu vzácných, ohrožených a endemických druhů Evropy (council of euroPe 1983), Směrnice o stanovištích (council of the euroPean coMMunities 1992), Červeného seznamu rostlin IUCN (walter & Gillett 1998), Evropského národního červeného seznamu ohrožených rostlin (council of euroPe 2001) a Seznamu evropských ohrožených druhů (sharrock & jones 2009; viz příloha 2).
Analýza dat
Při analýze dat jsme pracovali se třístupňovou škálou: (i) ohrožený – zahrnuje druhy kategorie CR, EN, VU, NT, R, I; (ii) neohrožený (běžný) – zahrnuje druhy kategorie LC/LR, DD, NE a všechny druhy nezahrnuté do červeného seznamu, ale uvedené na checklistu dané země a (iii) nevyskytuje se – zahrnuje druhy kategorie EX, EX? a všechny druhy, které nejsou uvedené na checklistu dané země. Pro každý druh jsme stanovili počet zemí, ve kterých je druh ohrožen, kde je běžný a kde se nevyskytuje. Abychom zohlednili fakt, že různé země se značně liší svou velikostí, provedli jsme rovněž přepočet na plochu zemí, kdy jsme pro každý druh stanovili celkovou plochu zemí, ve kterých je ohrožen, kde je běžný a kde se nevyskytuje. Oba dva způsoby vyjádření ohroženosti/běžnosti druhu – založené na počtu i na ploše zemí – jsme zahrnuli do následných analýz, ale výsledky byly víceméně podobné, proto prezentujeme vždy jen jednu variantu.
Pro každou zemi jsme vyjádřili počet a podíl počtu ohrožených, běžných a nevyskytujících se druhů, abychom ukázali, ve kterých zemích jsou naše C1 druhy ohrožené a ve kterých běžné. Výsledky jsme zobrazili formou map. Abychom ukázali, které druhy jsou relativně neohrožené (běžné ve velkém počtu zemí) a které jsou naopak ohrožené ve většině zemí svého výskytu, vytvořili jsme kumulativní graf, do kterého jsme vynesli podíl z celkového počtu druhů, běžných v určitém počtu zemí (běžných v 31, 30, 29 atd. zemích). Rovněž jsme vytvořili graf, do kterého jsme vynesli vztah celkové plochy zemí, ve kterých se druh vyskytuje a plochy zemí, ve kterých je druh běžný.
Dále jsme provedli srovnání našich dat s údaji ze studie schnittler & Günther (1999), kteří na základě údajů o celosvětových areálech rozšíření druhů a jejich ohrožení v zemích střední Evropy sestavili Seznam středoevropských druhů cévnatých rostlin vyžadujících prioritní ochranářská opatření (zkráceně Seznam středoevropských prioritních druhů). Jejich vymezení střední Evropy zahrnuje malou část Nizozemí, část Belgie, Švýcarsko,

305
Příroda, Praha, 31: 299–343, 2011
východní Francii, Lucembursko, Německo, Polsko, Českou a Slovenskou republiku, Rakousko a Maďarsko. Zajímalo nás, jestli české C1 druhy zahrnuté do Seznamu středoevropských prioritních druhů jsou více ohrožené v celé Evropě než české C1 druhy nezahrnuté do tohoto seznamu. Tuto otázku jsme testovali dvouvýběrovým t-testem. Podobné porovnání jsme provedli také mezi českými C1 druhy zahrnutými do Vyhlášky MŽP 395/1992 Sb. a českými C1 druhy nechráněnými zákonem ČR. Tímto srovnáním jsme chtěli zjistit, jestli české C1 druhy chráněné v ČR zákonem jsou více ohrožené rovněž z mezinárodního hlediska.
Rovněž nás zajímalo, jestli existuje nějaký vztah mezi typem stanoviště, ve kterém české C1 druhy rostou, a počtem evropských zemí, ve kterých jsou druhy ohrožené/běžné/nevyskytují se. České C1 druhy jsme rozdělili do devíti skupin podle typu stanoviště, ve kterém se převážně vyskytují (vodní toky a nádrže; mokřady a pobřežní vegetace; prameniště a rašeliniště; skály, sutě a jeskyně; alpínské bezlesí; sekundární trávníky a vřesoviště; křoviny; lesy; biotopy silně ovlivněné nebo vytvořené člověkem; kategorie podle Katalogu biotopů, chytrý et al. 2001). Pro druhy z jednotlivých typů stanoviště jsme vytvořili třírozměrný graf s osami znázorňujícími podíl počtu zemí, ve kterých je druh ohrožený, běžný a ve kterých se nevyskytuje (Statistica 7, StatSoft, Tulsa, Oklahoma, USA). Mezi různými typy habitatů jsme porovnali pozici druhů v rámci grafů.
obr. 1. Mapa zobrazující, v jakých zemích jsou kriticky ohrožené druhy ČR rovněž ohrožené. Odstín šedé vyjadřuje podíl českých C1 druhů, které jsou v dané zemi ohrožené, ze všech českých C1 druhů.Fig. 1. Map showing European countries where critically endangered plant species of the Czech Republic (C1 species) are considered as endangered. The tint of grey expresses the proportion from all C1 species, which are endangered in a particular country.

306
Gabrielová et al.: Ohrožení C1 rostlin ČR v jiných evropských zemích?
obr. 2. Mapa zobrazující, v jakých zemích jsou kriticky ohrožené druhy ČR běžné. Odstín šedé vyjadřuje podíl českých C1 druhů, které se v dané zemi vyskytují a nejsou zde ohrožené, ze všech českých C1 druhů.Fig. 2. Map showing European countries where critically endangered plant species of the Czech Republic (C1 species) are present but not considered as endangered. The tint of grey expresses the proportion from all C1 species which are not endangered in a particular country.
výsledky
Status ohrožení českých C1 druhů v jednotlivých evropských zemích je uve den v příloze 3 a rovněž je k dispozici v Databázi C1 rostlin na webo vých stránkách AOPK ČR (http://portal.nature.cz/c1/rostliny/), kde je situace ohrožení v Evropě pro každý druh znázorněna i pomocí mapy. Nejvíce českých C1 druhů (60–70 %) se vyskytuje v sousedních zemích – v Německu, Polsku, Rakousku a na Slovensku. V těchto zemích jsou české C1 druhy rovněž značně ohrožené (30 % ze všech českých C1 druhů je zde ohroženo; obr. 1). Nejméně českých C1 druhů (méně než 20 % z celkového počtu) roste na Maltě, Islandu, v Lucembursku a Irsku. České C1 druhy jsou relativně běžné ve Francii, Chorvatsku, Itálii, Rumunsku a na Ukrajině (40 % ze všech českých C1 druhů je zde běžných; obr. 2).
Pouze 12 % českých C1 druhů není běžných v žádné evropské zemi (obr. 3). Tuto skupinu tvoří převážně endemiti České republiky (viz příloha 3), druhy, jejichž rozšíření je nedokonale známo a jsou rozlišovány pouze v ČR (Dactylorhiza majalis subsp. turfosa), a druhy vyskytující se na malém území, které zahrnuje Českou republiku a jeden nebo
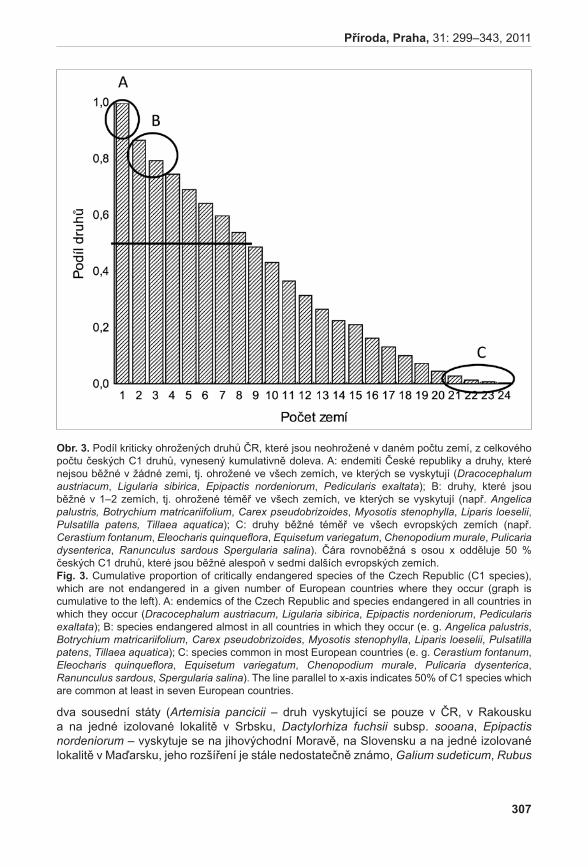
307
Příroda, Praha, 31: 299–343, 2011
obr. 3. Podíl kriticky ohrožených druhů ČR, které jsou neohrožené v daném počtu zemí, z celkového počtu českých C1 druhů, vynesený kumulativně doleva. A: endemiti České republiky a druhy, které nejsou běžné v žádné zemi, tj. ohrožené ve všech zemích, ve kterých se vyskytují (Dracocephalum austriacum, Ligularia sibirica, Epipactis nordeniorum, Pedicularis exaltata); B: druhy, které jsou běžné v 1–2 zemích, tj. ohrožené téměř ve všech zemích, ve kterých se vyskytují (např. Angelica palustris, Botrychium matricariifolium, Carex pseudobrizoides, Myosotis stenophylla, Liparis loeselii, Pulsatilla patens, Tillaea aquatica); C: druhy běžné téměř ve všech evropských zemích (např. Cerastium fontanum, Eleocharis quinqueflora, Equisetum variegatum, Chenopodium murale, Pulicaria dysenterica, Ranunculus sardous Spergularia salina). Čára rovnoběžná s osou x odděluje 50 % českých C1 druhů, které jsou běžné alespoň v sedmi dalších evropských zemích.Fig. 3. Cumulative proportion of critically endangered species of the Czech Republic (C1 species), which are not endangered in a given number of European countries where they occur (graph is cumulative to the left). A: endemics of the Czech Republic and species endangered in all countries in which they occur (Dracocephalum austriacum, Ligularia sibirica, Epipactis nordeniorum, Pedicularis exaltata); B: species endangered almost in all countries in which they occur (e. g. Angelica palustris, Botrychium matricariifolium, Carex pseudobrizoides, Myosotis stenophylla, Liparis loeselii, Pulsatilla patens, Tillaea aquatica); C: species common in most European countries (e. g. Cerastium fontanum, Eleocharis quinqueflora, Equisetum variegatum, Chenopodium murale, Pulicaria dysenterica, Ranunculus sardous, Spergularia salina). The line parallel to x-axis indicates 50% of C1 species which are common at least in seven European countries.
dva sousední státy (Artemisia pancicii – druh vyskytující se pouze v ČR, v Rakousku a na jedné izolované lokalitě v Srbsku, Dactylorhiza fuchsii subsp. sooana, Epipactis nordeniorum – vyskytuje se na jihovýchodní Moravě, na Slovensku a na jedné izolované lokalitě v Maďarsku, jeho rozšíření je stále nedostatečně známo, Galium sudeticum, Rubus
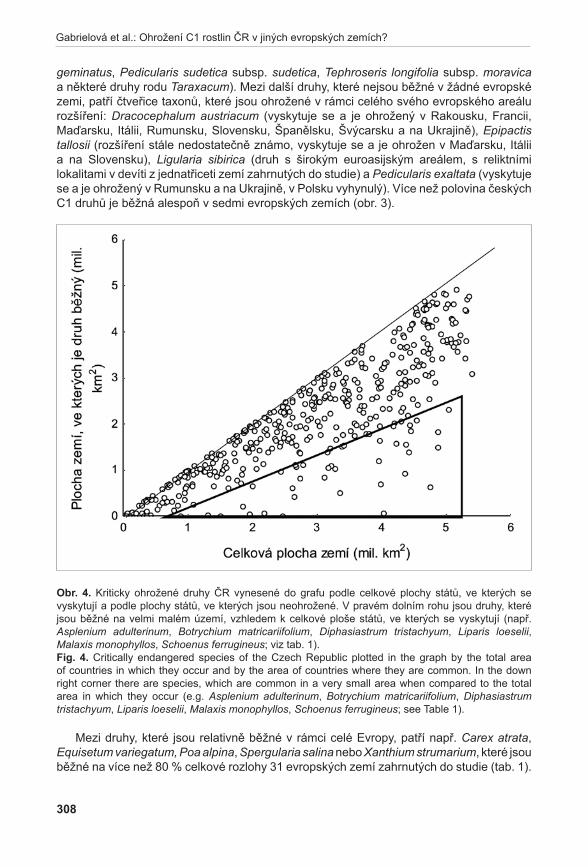
308
Gabrielová et al.: Ohrožení C1 rostlin ČR v jiných evropských zemích?
geminatus, Pedicularis sudetica subsp. sudetica, Tephroseris longifolia subsp. moravica a některé druhy rodu Taraxacum). Mezi další druhy, které nejsou běžné v žádné evropské zemi, patří čtveřice taxonů, které jsou ohrožené v rámci celého svého evropského areálu rozšíření: Dracocephalum austriacum (vyskytuje se a je ohrožený v Rakousku, Francii, Maďarsku, Itálii, Rumunsku, Slovensku, Španělsku, Švýcarsku a na Ukrajině), Epipactis tallosii (rozšíření stále nedostatečně známo, vyskytuje se a je ohrožen v Maďarsku, Itálii a na Slovensku), Ligularia sibirica (druh s širokým euroasijským areálem, s reliktními lokalitami v devíti z jednatřiceti zemí zahrnutých do studie) a Pedicularis exaltata (vyskytuje se a je ohrožený v Rumunsku a na Ukrajině, v Polsku vyhynulý). Více než polovina českých C1 druhů je běžná alespoň v sedmi evropských zemích (obr. 3).
obr. 4. Kriticky ohrožené druhy ČR vynesené do grafu podle celkové plochy států, ve kterých se vyskytují a podle plochy států, ve kterých jsou neohrožené. V pravém dolním rohu jsou druhy, které jsou běžné na velmi malém území, vzhledem k celkové ploše států, ve kterých se vyskytují (např. Asplenium adulterinum, Botrychium matricariifolium, Diphasiastrum tristachyum, Liparis loeselii, Malaxis monophyllos, Schoenus ferrugineus; viz tab. 1).Fig. 4. Critically endangered species of the Czech Republic plotted in the graph by the total area of countries in which they occur and by the area of countries where they are common. In the down right corner there are species, which are common in a very small area when compared to the total area in which they occur (e.g. Asplenium adulterinum, Botrychium matricariifolium, Diphasiastrum tristachyum, Liparis loeselii, Malaxis monophyllos, Schoenus ferrugineus; see Table 1).
Mezi druhy, které jsou relativně běžné v rámci celé Evropy, patří např. Carex atrata, Equisetum variegatum, Poa alpina, Spergularia salina nebo Xanthium strumarium, které jsou běžné na více než 80 % celkové rozlohy 31 evropských zemí zahrnutých do studie (tab. 1).

309
Příroda, Praha, 31: 299–343, 2011
Mezi druhy, které jsou sice široce rozšířené, ale zároveň i vysoce ohrožené (ohrožené na více než 40 % území Evropy) patří např. Botrychium matricariifolium, Drosera anglica, Epipogium aphyllum, Liparis loeselii nebo Schoenus ferrugineus (tab. 1). Abychom odhalili druhy, které jsou značně ohrožené vzhledem k ploše, na které se vyskytují, vynesli jsme do grafu závislost mezi celkovou plochou zemí, ve kterých se druh vyskytuje, a plochou zemí, ve kterých druh není ohrožen (obr. 4). Druhy nacházející se v pravém dolním rohu obr. 4 jsou ty, které jsou ohrožené ve velkém podílu plochy zemí, v nichž se vyskytují. K těmto druhům patří např. Artemisia pancicii, Cirsium brachycephalum, Epipactis tallosii nebo Lathyrus pannonicus subsp. pannonicus (tab. 1).
tab. 1. Příklady kriticky ohrožených druhů ČR, které jsou relativně neohrožené nebo relativně ohrožené v jiných evropských zemích.tab. 1. Examples of critically endangered plant species of the Czech Republic, which are relativelly common or relativelly endangered in other European countries.
Příklady druhů
neohrožené na více než 80 % celkové plochy 31 evropských zemí
Bromus squarrosus, Carex atrata, Cerastium fontanum, Equisetum variegatum, Eragrostis pilosa, Gnaphalium supinum, Juncus gerardii, Lathyrus aphaca, Poa alpina, Pulicaria dysenterica, Pyrola chlorantha, Ranunculus sardous, Sideritis montana, Spergularia salina, Xanthium strumarium
Široce rozšířené, ohrožené na více než 40 % celkové plochy evropských zemí
Botrychium matricariifolium, Diphasiastrum tristachyum, Dracocephalum austriacum, Drosera anglica, Drosera intermedia, Epipogium aphyllum, Eriophorum gracile, Hammarbya paludosa, Liparis loeselii, Malaxis monophyllos, Nymphoides peltata, Ophrys insectifera, Rhynchospora fusca, Scheuchzeria palustris, Schoenus ferrugineus
Relativně ohrožené s ohledem na plochu, na které se vyskytují
Artemisia pancicii, Asplenium adulterinum, Carex pseudobrizoides, Dracocephalum austriacum, Epipactis nordeniorum, Epipactis tallosii, Gentianella campestris subsp. baltica, Laserpitium archangelica, Ligularia sibirica, Liparis loeselii, Pedicularis exaltata, Rubus geminatus, Stipa borysthenica, Stipa smirnovii, Tillaea aquatica
Porovnání mezi českými C1 druhy zahrnutými a nezahrnutými do Seznamu středoevropských prioritních druhů ukázalo signifikantně větší ohrožení českých C1 druhů uvedených v seznamu (obr. 5a). Objevily se ovšem i druhy, které nejsou uvedené v Seznamu středoevropských prioritních druhů a přitom jsou značně ohroženy v rámci celé Evropy, např. Campanula cervicaria, Dianthus superbus subsp. superbus, Isoëtes lacustris, Malaxis monophyllos, Ophrys insectifera, Orchis ustulata nebo Scheuchzeria palustris. Rovněž české C1 druhy v ČR chráněné zákonem jsou ohrožené v signifikantně větším počtu zemí než druhy, které nejsou chráněné zákonem (obr. 5b). Přesto je zde poměrně dost českých C1 druhů, které v ČR nejsou chráněné zákonem, ale jsou ohrožené na relativně velké ploše Evropy, např. Carex pseudobrizoides, Hierochloë odorata, Luronium natans, Minuartia viscosa, Radiola linoides a Typha shuttleworthii. Na druhou stranu najdeme i druhy, které jsou v ČR chráněné zákonem, ale v řadě ostatních evropských zemí jsou běžné, např. Carex atrata, Luzula spicata, Gentiana verna, Aposeris foetida a Veronica bellidioides.
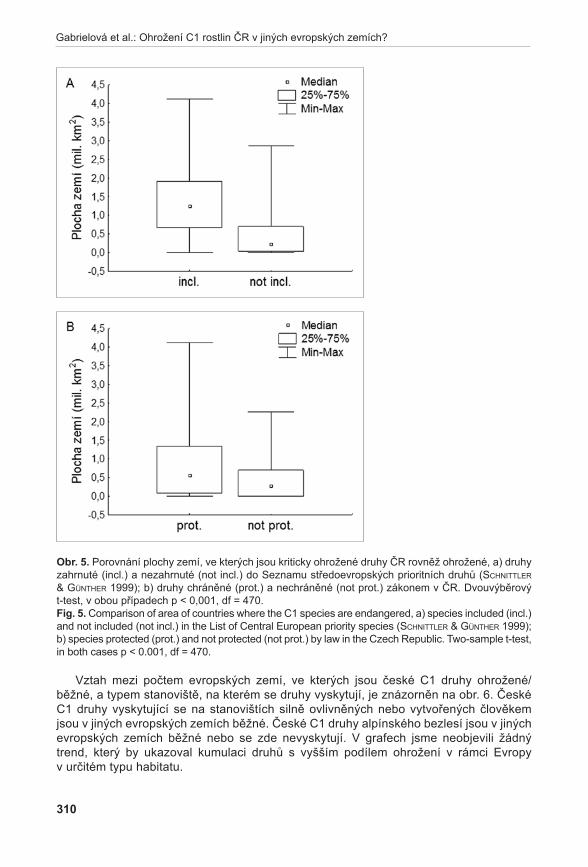
310
Gabrielová et al.: Ohrožení C1 rostlin ČR v jiných evropských zemích?
obr. 5. Porovnání plochy zemí, ve kterých jsou kriticky ohrožené druhy ČR rovněž ohrožené, a) druhy zahrnuté (incl.) a nezahrnuté (not incl.) do Seznamu středoevropských prioritních druhů (schnittler & Günther 1999); b) druhy chráněné (prot.) a nechráněné (not prot.) zákonem v ČR. Dvouvýběrový t-test, v obou případech p < 0,001, df = 470.Fig. 5. Comparison of area of countries where the C1 species are endangered, a) species included (incl.) and not included (not incl.) in the List of Central European priority species (schnittler & Günther 1999); b) species protected (prot.) and not protected (not prot.) by law in the Czech Republic. Two-sam ple t-test, in both cases p < 0.001, df = 470.
Vztah mezi počtem evropských zemí, ve kterých jsou české C1 druhy ohrožené/běžné, a typem stanoviště, na kterém se druhy vyskytují, je znázorněn na obr. 6. České C1 druhy vyskytující se na stanovištích silně ovlivněných nebo vytvořených člověkem jsou v jiných evropských zemích běžné. České C1 druhy alpínského bezlesí jsou v jiných evropských zemích běžné nebo se zde nevyskytují. V grafech jsme neobjevili žádný trend, který by ukazoval kumulaci druhů s vyšším podílem ohrožení v rámci Evropy v určitém typu habitatu.

311
Příroda, Praha, 31: 299–343, 2011
obr. 6. Zastoupení českých C1 druhů podle podílu počtu zemí, ve kterých jsou ohrožené (Ohr.), běžné (Neohr.) a ve kterých se nevyskytují (Nevysk.) s ohledem na typ habitatu, ve kterém se převážně vyskytují.Fig. 6. The distribution of C1 species according to the proportion of the number of countries in which they are endangered (Ohr.), not endangered (Neohr.) and not occuring (Nevysk.) with the respect to the habitat type in which they predominantly occur. Sekundární trávníky – secondary grasslands and heathlands, Biotopy ovlivněné člověkem – habitats strongly influenced or created by man, Alpínské bezlesí – alpine treeless habitats, Lesy – forests, Skály a sutě – cliffs and boulder screes, Vodní toky a nádrže – streams and water bodies, Prameniště a rašeliniště – springs and mires, Mokřady – wetlands and riverine vegetation, Křoviny – scrub.
diskuse
Z výsledků vyplývá, že velký počet českých C1 druhů se vyskytuje v sousedních zemích (Rakousko, Německo, Polsko a Slovensko), kde jsou rovněž značně ohrožené. Česká republika tedy s těmito zeměmi sdílí podobné ochranářské priority, ovšem potenciální „záchranný efekt“ těchto zahraničních populací pro české populace C1 druhů se tím pádem jeví značně omezený. Vysoký počet českých C1 druhů se vyskytuje a je běžných ve Francii, Itálii, Rumunsku a na Ukrajině. V těchto státech má evidentně řada C1 druhů

312
Gabrielová et al.: Ohrožení C1 rostlin ČR v jiných evropských zemích?
centrum svého výskytu a Česká republika pro ně představuje okraj areálu. Značná geografická vzdálenost populací v těchto státech ovšem neumožňuje uvažovat o nich jako o potenciálním zdroji genetického materiálu pro oslabené české populace.
Pokud se zaměříme pouze na státy střední Evropy (Maďarsko, Německo, Polsko, Rakousko, Slovensko a Švýcarsko), můžeme naše výsledky porovnat se závěry studie schnittler & Günther (1999). České C1 druhy zahrnuté do jejich Seznamu středoevropských prioritních druhů si opravdu zasluhují zvýšenou pozornost, protože jsou ohrožené na podstatně větším území než české C1 druhy nezahrnuté do tohoto seznamu. Ovšem je třeba upozornit i na některé „opomenuté“ druhy, které jsou ohrožené v minimálně pěti ze šesti středoevropských států, a přesto nebyly do Seznamu prioritních středoevropských druhů zahrnuty, jako např. Conringia orientalis, Dianthus superbus subsp. superbus, Hippuris vulgaris, Juncus subnodulosus, Orchis ustulata, Radiola linoides nebo Sagina subulata.
Porovnání ohrožení v rámci Evropy mezi druhy chráněnými v ČR zákonem a druhy zákonem nechráněnými ukázalo, že české C1 druhy chráněné zákonem jsou ohrožené na větší ploše evropských zemí a představují tak druhy, které by skutečně měly být ochranářskou prioritou i z mezinárodního úhlu pohledu. To je poměrně zajímavé zjištění, protože při sestavování Vyhlášky možná mohlo být k mezinárodnímu ohrožení taxonů přihlédnuto, ale rozhodně to nebylo nijak důležité kritérium (mnohem větší roli u druhů chráněných zákonem v ČR hraje jejich početnost, stav jejich populací a stanovišť v rámci ČR). Některým druhům, které v současné době nejsou na Vyhlášce MŽP 395/1992 Sb., by možná měla být věnována větší pozornost (např. Adonis flammea, Bupleurum rotundifolium, Conringia orientalis, Galium tricornutum, Orobanche coerulescens, Orobanche picridis, Radiola linoides nebo Sagina subulata), protože tyto druhy jsou ohrožené v rámci celé Evropy. Zčásti se jedná o skupinu teplomilných plevelů, které jsou ohrožené zejména intenzifikací zemědělství a které je obtížné prakticky chránit. Aktualizace vyhlášky, která je v současné době v procesu schvalování na Ministerstvu životního prostředí, už všechny tyto druhy (kromě druhu Galium tricornutum) zahrnuje, a to přinejmenším v kategorii „ohrožené“.
Výsledky ukazují další druhy nebo skupiny druhů, které by neměly být přehlíženy při stanovování ochranářských priorit. Jedná se především o endemity ČR, i když jejich taxonomická hodnota je často poněkud diskutabilní. Přesto představují originální část evoluční historie a jejich extinkce by vedla k ochuzení globální genetické diverzity. Podobnou skupinu druhů, za kterou má Česká republika velkou zodpovědnost, představují úzce rozšířené druhy s centrem areálu rozšíření ve Střední Evropě (např. Artemisia pancicii, Epipactis nordeniorum, Gentianella praecox subsp. bohemica, Rubus geminatus, Pedicularis sudetica nebo Tephroseris longifolia subsp. moravica), případně druhy, jejichž populace v České republice představují izolované arely mimo hlavní část areálu rozšíření (Carex pediformis subsp. macroura, Hieracium subortum, Stipa zaleskii, Taraxacum argutum).
Na druhou stranu analýza rovněž ukázala, že některé druhy, které jsou v ČR velmi vzácné, a proto řazené do kategorie C1 a chráněné zákonem, jsou v jiných evropských zemích poměrně běžné. K těmto druhům patří např. Carex atrata, Juniperus communis subsp. alpina, Luzula spicata nebo Poa alpina, které jsou běžné ve většině států zahrnutých do studie. Jedná se o druhy, které se typicky vyskytují na nějakém vzácném typu stanoviště (v tomto případě alpínské bezlesí), jehož rozsah je v rámci ČR velmi malý, ale v jiných evropských státech je běžnější.
Jako měřítko ohrožení českých C1 druhů v dalších evropských zemích jsme používali počet států a plochu států. Uvědomujeme si, že ani jeden z těchto způsobů není dokonalý a že bychom ideálně měli zohlednit informaci o reálném rozšíření druhu v dané zemi.

313
Příroda, Praha, 31: 299–343, 2011
Bohužel tyto údaje nejsou zdaleka dostupné pro všechny druhy. Tím, že jsme vzali v úvahu rozlohy zemí, jsme se snažili zohlednit skutečnost, že pokud je druh ohrožený např. v Německu, má to trochu jiný význam, než když je ohrožený např. v Lucembursku. Konkrétně v rozlohou velmi malém Lucembursku, kde byl červený seznam sestaven na základě kritérií IUCN a byl vyhodnocován status ohrožení pro všechny druhy flóry, byly jako neohrožené klasifikovány pouze čtyři druhy.
Neméně závažný problém je značná nekonzistence mezi jednotlivými zdroji informací o statutu ohrožení českých C1 druhů v dalších evropských zemích. Některé červené seznamy jsou značně zastaralé, koncept národních kategorií a kritérií se značně liší, odlišuje se také počet a spektrum druhů zahrnutých do červených seznamů, červených knih nebo seznamů druhů chráněných zákonem. Červené knihy typicky obsahují jen výběr části ohrožených druhů dané země; pokud však není k dispozici červený seznam, je to pro nás jediný zdroj informací. Komplikaci představuje rovněž nejednotné taxonomické pojetí různých zemí. Checklisty jsou klíčové pro určení toho, zda je druh v zemi přítomen a neohrožen nebo se v zemi vůbec nevyskytuje. Ovšem i úroveň checklistů se značně liší, navíc pro několik zemí, které nemají checklist, jsme vycházeli z databáze Flora Europaea, která je velmi zastaralá a obsahuje řadu nepřesností. Proto je velmi pravděpodobné, že situace ohrožení v ostatních evropských zemích pro některé české C1 druhy může být podhodnocená nebo naopak nadhodnocená.
I přes tyto nedostatky se však domníváme, že se nám podařilo shromáždit cenné údaje, které mohou napomoci kvalifikovanému rozhodování v ochraně přírody. Pro přímý odhad ohrožení druhů na celoevropské úrovni nemáme dostatek informací a náš přístup tak představuje v podstatě jedinou možnost, jak získat podobné údaje pro poměrně velký počet druhů z rozsáhlého území. Můžeme jen doufat, že k odhadování ohrožení druhů v jednotlivých zemích se v budoucnu široce rozšíří systém stejných porovnatelných kategorií a kritérií (např. systém IUCN).
shrnutí
Na základě údajů o ohrožení českých C1 druhů v dalších evropských zemích jsme schopni identifikovat druhy, za jejichž ochranu má Česká republika velkou zodpovědnost, protože jejich celkové rozšíření je velmi omezené, nebo jsou ohrožené v rámci celého svého evropského areálu rozšíření (endemity ČR; Dracocephalum austriacum, Epipactis tallosii, Ligularia sibirica, Pedicularis exaltata). Rovněž můžeme ukázat druhy, které jsou šířeji rozšířené a v některých státech i běžné, ale přesto jsou vzhledem k celkovému území, na kterém se vyskytují, značně ohrožené a navíc nejsou zahrnuté do žádných mezinárodních konvencí ani seznamů prioritních druhů (např. Campanula cervicaria, Dianthus superbus subsp. superbus, Isoëtes lacustris nebo Scheuchzeria palustris). Na druhou stranu odhalíme i druhy, které jsou v České republice velmi vzácné a chráněné zákonem, ale ve většině dalších evropských zemí jsou běžné – typicky druhy subalpínského stupně (např. Carex atrata, Helianthemum grandiflorum subsp. grandiflorum, Luzula spicata, Poa alpina nebo Veronica bellidioides). Údaje o ohrožení v okolních evropských zemích tak napomohou při stanovení priorit v druhové ochraně cévnatých rostlin v České republice. Pro úplný vhled do problematiky je však samozřejmě potřeba tyto údaje propojit s informacemi nashromážděnými v dalších částech projektu.

314
Gabrielová et al.: Ohrožení C1 rostlin ČR v jiných evropských zemích?
summary
Our data provide a useful insight into setting priorities in species conservation in the Czech Republic. Indeed, data on other aspects of the species will be needed to have realistic understanding of the issue. On the basis of our data, we can identify species for the conservation of which the Czech Republic is highly responsible as they are very narrowly distributed or are endangered in a large part of their distribution range (species endemic in the Czech Republic; Dracocephalum austriacum, Epipactis tallosii, Ligularia sibirica, Pedicularis exaltata). We can also identify species which are more widely distributed and common in some countries but are still relatively highly endangered with the respect to the area in which they occur. These species are also not included in any international convention and even in the List of Central European priority species (e.g. Campanula cervicaria, Dianthus superbus subsp. superbus, Isoëtes lacustris or Scheuchzeria palustris). On the other hand, we can point at species which are very rare and legaly protected in the Czech Republic but not classified as endangered almost in any country where they occur – typically species of high elevations (e.g. Carex atrata, Helianthemum grandiflorum subsp. grandiflorum, Luzula spicata, Poa alpina or Veronica bellidioides). In conclusion, our dataset will provide a valuable contribution to decision-making processes in species conservation.
Poděkování
Děkujeme J. Chrtkovi za pomoc s taxonomickou a nomenklatorickou problematikou druhů a K. Marholdovi za zapůjčení řady užitečných publikací. J. Čeřovskému děkujeme za přínosné komentáře k rukopisu. Studie byla podpořena granty MŠMT VaV 2B06178, 206-08-H049 a výzkumnými záměry MSM 0021620828 a AV0Z60050516.
literatura
aeschiMann D. & heitz c. (2005): Synonymie-Index der Schwezer Flora und der angrenzender Gebiete, 2. Auflage. – Zentrum des Datenverbundnetzes der Schweizer Flora (ZDSF), Genève. URL: http://www.crsf.ch/documents/download/x/ISFS2_20051115.pdf (cit. 5. 2. 2011).
alves j. (2001): Lista de Espécies Autóctones ou Naturalizadas, Ameaçadas, Raras ou com estatuto indeterminado – Baseada na versao de 1996, publicada em: “A conservacao in situ como instrumento de conservacao dinamica da biodiversidade (sementes para um debate)”.
anDrushaitis G. P. [ed.] (1985): Red data of the Latvian SSR. Rare and endangered species of animals and plants. – Zinatne Publishing House, Riga.
anonyMus (2005): Перечень объектов растительного мира , занесенных в Красную Книгу Российской Федерации. – Приложение 1 к приказу МПР России от 25 октября 2005 г. № 289.
assyov b., Petrova a., DiMitrov D. & vassilev r. (2006): Conspectus of the Bulgarian vascular flora. Distribution maps and floristic elements. Third revised and enlarged edition. – Bulgarian Biodiversity Foundation, Sofia.
bunDesaMt für naturschutz: FloraWeb. – URL: http://www.floraweb.de/. (cit. 5. 2. 2011).collinG G. (2005): Red List of the vascular plants of Luxembourg. – Ferrantia 42: 1–77. URL:
http://www.mnhn.lu/recherche/redbook/vascplants/default.htm. (cit. 5. 2. 2011).

315
Příroda, Praha, 31: 299–343, 2011
coMMission for nature conservation of the estonian acaDeMy of sciences (2001-2002): The new Estonian Red List of threatened species. – URL: http://www.zbi.ee/punane/english/index.html. (cit. 5. 2. 2011).
council of euroPe (1979): Convention on the conservation of European wildlife and natural habitats. – Strasbourg.
council of euroPe (1983): List of rare, threatened and endemic plants in Europe. – Nature and Environment Series No 27, Strasbourg.
council of euroPe (2001): European national Red List of threatened vascular plants. Project report and description of the corresponding data base. – T-PVS (2001) 9 Strasbourg.
council of the euroPean coMMunities (1992): Council Directive 92/43/EEC of 21. May 1992 on the conservation of natural habitats and of wild fauna and flora. – Official Journal of the European Communities 35: 7–50.
conti f., abbate G., alessanDrini a. & blasi c. [eds.] (2005): An annotated checklist of the Italian vascular flora. – Palombi Editori.
conti f., Manzi a. & PeDrotti f. (1997): Liste rosse regionali delle piante d‘Italia. – Societá Botanica Italiana. Dipartimento di Botanico ed Ecologia, Univ. degli Studi di Camerino. Camerino.
curtis G. f. & McGouGh h. n. (1988): The Irish Red Data Book 1: Vascular plants. – Wildlife Service Ireland, Dublin.
De GraMMont P. c. & cuarón a. D. (2005): An evaluation of threatened species categorization systems used on the American continent. – Conserv. Biol. 20: 14–27.
finnish environMent institute (2001): Extinct and Threatened vascular plants. – URL: http://www.environment.fi/default.asp?contentid=28900&lan=en) (cit. 5. 2. 2011).
fisher M. a., oswalD k. & aDler w. (2008): Exkursionsflora für Österreich, Lichtenstein und Südtirol, 3. Auflage. – Biologiezentrum der Oberösterreichischen Landesmuseen.
flora euroPaea Database. – Royal Botanic Garden Edinburgh. – URL: http://rbg-web2.rbge.org.uk/FE/fe.html). (cit. 5. 2. 2011).
flora iberica. Plantas vasculares de la Península Ibérica e Islas Baleares. – Real Jardín Botánico. URL: http://www.floraiberica.es/v.2.0/organizacion/planobra.php. (cit. 5. 2. 2011).
GärDenfors u. (2001): Classifying threatened species at national versus global levels. – Trends Ecol. Evol. 16: 511–516.
GärDenfors u., hilton-taylor c., Mace G. & roDríGuez j. P. (2001): The application of IUCN red list criteria at regional levels. – Conserv. Biol. 15: 1206–1212.
GärDenfors u. [ed.]. (2005): Rödlistade arter i Sverige 2005. [The 2005 Red List of Swedish Species]. – ArtDatabanken, SLU, Uppsala.
Gerža M. (2009): Endemismus v České republice. Rostliny – 1. část. – Ochr. Přír. 64/2: 12–15.GuDžinskas z. (1999): Lietuvos induočiai augalai. [Vascular plants of Lithuania]. – Botanikos
instituto leidykla, Vilnius. Guinochet M. & vilMorin r. (1973): Flore de France. Fascicule 1. – Du centre National de la
recherche scientifique, Paris.hartley s. & kunin w. e. (2003): Scale dependency of rarirty, extinction risk and conservation
priority. – Conserv. Biol. 17: 1559–1570.häMet-ahti l., suoMinen j., ulvinen t. & uotila P. (1986): Retkeilykasvio. [Field Flora of Finland].
4th Ed. – Botanical Museum, Finnish Museum of Natural History, Helsinki.hejný s. & slavík b. [eds] (1988): Květena České socialistické republiky. Vol 1. – Academia, Praha.hejný s. & slavík b. [eds] (1990): Květena České republiky. Vol. 2. – Academia, Praha.hejný s. & slavík b. [eds] (1992): Květena České republiky. Vol. 3. – Academia, Praha.holub j. (1996): Stupnice a kriteria ohrožení užívané v Červených seznamech. – Severočes.
Přír., Litoměřice, suppl. 9: 7–14.holub j. & Procházka f. (2000): Red list of the flora of the Czech Republic (state in the year
2000). – Preslia, Praha, 72: 187–230.

316
Gabrielová et al.: Ohrožení C1 rostlin ČR v jiných evropských zemích?
cheffinGs c. f. & farrell l. [eds] (2005): The vascular plant Red Data List for Great Britain. – Species Status 7: 1–116. Joint Nature Conservation Committee, Peterborough.
chrtek j. & MarholD k. (1998): Taxonomy of the Hieracium fritzei Group (Asteraceae) in the Sudeten Mtd and the West Carpathians. (Studies in Hieracium sect. alpina II.). – Phyton (Horn. Austria) 37: 181–217.
chytrý M., kuČera t. & koČí M. [eds] (2001): Katalog biotopů České republiky. – Agentura ochrany přírody a krajiny České republiky, Praha.
inGaDóttir á., einarsson e., kristinsson h. & eGilsson k. [eds] (1996): Valisti 1. Plöntur. Red Data List 1. Plants. – Icelandic Natural History Institute, Reykjavik.
iucn (2001): IUCN Red List Categories and Criteria: Version 3.1. – IUCN Species Survival Commission. IUCN, Gland, Switzerland and Cambridge, U.K.
iucn (2003): Guidelines for application of IUCN Red List Criteria at regional levels: Version 3.0. – IUCN Species Survival Commission. IUCN, Gland, Switzerland and Cambridge, U.K.
iucn (2010): IUCN Standards and Petitions Subcommittee. 2010. Guidelines for using the IUCN Red List Categories and Criteria. Version 8.1. Prepared by the Standards and Petitions Subcommittee in March 2010. – URL: http://intranet.iucn.org/webfiles/doc/SSC/RedList/RedListGuidelines.pdf (cit. 26. 9. 2011).
IUCN (2011): IUCN Red List of Threatened Species. Version 2011.1. – URL: http://www.iucnredlist.org (cit. 26. 9. 2011).
kÅlÅs j. a., viken Å. & bakken t. [eds] (2006): Norsk Rødliste 2006. [2006 Norwegian Red List]. – Artsdatabanken, Norway. URL: http://www.artsdatabanken.no/Article.aspx?m=246&amid=6696 (cit. 5. 2. 2011).
karlsson t. (2004): Checklista över Nordens kärlväxter. Version 2004-01-19. – Naturhistoriska riksmuseet. http://www2.nrm.se/fbo/chk/chk3.htm (cit. 5. 2. 2011).
kazMierczakowa r. & zarzycki k. [eds] (2001): Polska Czerwona Księga Roślin, ed. 2. Pteridophytes and Flowering Plants. – Instytut Botaniki im. W. Szafera, Polska Akademia Nauk, Kraków.
keller v. & bollMann k. (2004): From Red List to species of conservation concern. – Conserv. Biol. 18: 1636–1644.
keller v., zbinDen n., schMiD h. & volet b. (2005): A case study in applying the IUCN regional guidelines for national Red Lists and justifications for their modification. – Conserv. Biol. 19: 1827–1834.
király G. [ed.] (2007): Vöros Lista. A magyarországi edényes flóra veszélyeztetett fajai. [Red list of vascular flora of Hungary]. – Saját kiadás, Sopron.
király G. [ed.] (2009): Új magyar füvészkönyv. Magyarország hajtásos növényei.Határozókulcsok. [New Hungarian Herbal. The vascular plants of Hungary. identification key]. – Aggteleki Nemzeti Park Igazgatóság, Jósvafö.
kukk t. (1999): Eesti taimestik. [Vascular plant flora of Estonia.]. – Teaduste Akadeemia Kirjastus, Tartu-Tallinn.
laMbinon j., Del vosalle l. & DuviGneauD j. (2008): Nouvelle flore de la Belgique, du Grand-Duché de Luxembourg, du Nord de la France et des Régions voisines (Ptéridophytes et Spermatophytes). Cinquieme Édition. – Edition du Jardin botanique national de Belgique.
liD j. & liD D. t. (2005): Norsk flora. 7. utgåve ved Reidar Elven. – Det Norske Samlaget, Oslo.lietuvos resPublikos aPlinkos Ministro (2003): Lietuvos raudonąją knygą įrašytų saugomų
gyvūnų, augalų ir grybų rūšių sąrašas [The red data book of animals, plants and mushrooms of Lithuania]. 2003 m. spalio 13 d. Nr. 504, Vilnius. – URL: http://www3.lrs.lt/pls/inter3/dokpaieska.showdoc_e?p_id=219902 (cit. 5. 2. 2011).
MarholD k. & hinDák f. (1997): Zoznam nižších a vyšších rastlín Slovenska. – Veda, vydavateľstvo SAV. URL: http://ibot.sav.sk/checklist/ (cit. 5. 2. 2011).

317
Příroda, Praha, 31: 299–343, 2011
MarholD k., Mártonfi P., Mereďa P. jun., Mráz P., hoDálová i., kolník M., kuČera j., lihová j., Mrázová v., Perný M. & valko i. (2007): Karyologická databáza papraďorastov a semenných rastlín Slovenska. Verzia 1.0 (Posledná aktualizácia 31.10.2007). – URL: http://www.chromosomes.sav.sk/ (cit. 5.2.2011).
MaXiMovitch D. a. [ed.] (1993): Cyrvonaja Kniga Respubliki Belarus. – Minsk.van Der MeijDen r., oDé b., Groen l. G., witte M. & bal D. (2000): Bedreigde en kwetsbare
vaatplanten in Nederland. Basisrapport met voorstel vor de Rode Lijst. [Redlist of vascular plants of Netherland]. – Gorteria 26, Nationaal Herbarium Nederland, Leiden en Stichting FLORON.
MifsuD s. (2002/2010): Wild plants of the Mediterranean Islands of Malta. – URL: http://www.maltawildplants.com/SearchIndex.php (cit. 5. 2. 2011).
Miller r. M., roDríGuez j. P., aniskowicz-fowler t., baMbaraDeniya ch., boles r., eaton M. a., GärDenfors u., keller v., Molur s., walker s. & Pollock c. (2007): National threatened species listing based on IUCN criteria and regional guidelines: Current status and future perspectives. – Conserv. Biol. 21: 684–696.
Mirek z., Piękoś-Mirkowa h., zając a. & zając M. (2002): Flowering plants and pteridophytes of Poland. – A checklist. [Krytyczna lista roslin naczyniowych Polski.] – W. Szafer Institute of Botany, Polish Academy of Sciences, Kraków.
Mirek z., zarzycki k., wojewoDa w. & szeląG z. [eds] (2006): Red List of plants and fungi in Poland. – Kraków.
Moravcová l., jarolíMová v. & zákravský P. (2001): Morphological differences and chromosome numbers in Puccinellia distans and P. limosa populations from Central Europe. – Preslia 73: 161–172.
Moreno j. c. [ed.] (2008): Lista Roja 2008 de la flora vascular española. – Dirección General de Medio Natural y Política Forestal (Ministerio de Medio Ambiente, y Medio Rural y Marino, y Sociedad Española de Biología de la Conservación de Plantas), Madrid. URL: http://www.conservacionvegetal.org/PDF/Lista_Roja_Flora%202008.pdf (cit. 5. 2. 2011).
Moser D., GyGaX a., bäuMler b., wyler n. & Palese r. (2002): Rote Liste der gefährdeten Farn- und Blütenpflanzen der Schweiz. – Bundesamt für Umwelt, Wald und Landschaft, Bern; Zentrum des Datenverbundnetzes der Schweizer Flora, Chambésy; Conservatoire et Jardin botaniques de la Ville de Genève, Chambésy. BUWAL-Reihe „Vollzug Umwelt“. URL: http://www.crsf.ch/documents/download/d/d_rl2002_pflanzen.pdf (cit. 5. 2. 2011).
Mosyakin s. l. & feDoronchuk M. M. (1999): Vascular plants of Ukraine. A nomenclatural checklist. – National Academy of Sciences of Ukraine. M. G. Khologny Institute of Botany.
niklfelD h. (1999): Rote Listen gefährdeter Pflanzen Österreichs. 2. Auflage. – Grüne Reihe des Bundesministeriums für Gesundheit und Umweltschutz, Band 5, Wien.
nikoli t. [ed.] (2010): Flora Croatica Database, On-Line. – Department of Botany, Faculty of Science, University of Zagreb. URL: http://hirc.botanic.hr/fcd (cit. 5. 2. 2011).
nyáráDy e.i. & savulescu t. [eds] (1952–1976): Flora Republicii Socialiste România. Vol. I–XIII. – Editura Academiei Republicii Socialiste România. Bucuresti.
olivier l., GallanD j.-P., nMaurin h. & rouX P.-P. [eds] (1995): Livre Rouge de la Flore Menacé de France, I. Espéces Prioritaires. – Museum National d’Histoire Naturelle, Conservatoire Botanique National de Porquerolles & Ministère de l’Environnement, Paris.
oltean M., neGrean G., PoPescu a., roMan n., Dihoru G., sanDa v. & Mihailescu s. (1994): Lista rosie a plantelor superioare din România. [Red list of higher plants of Romania.] – Institutul de Biologie, Academia Româna.
Phitos D., striD a., snoGeruP s. & Greuter w. [eds] (1995): The Red Data Book of rare and threatened plants of Greece. – WWF-Greece, Athens.
PiGnatti s., MeneGoni P. & Giacanelli v. [eds] (2001). Liste rosse e blu della flora italiana. – ANPA, Collana Stato dell’Ambiente 1. Alcagraf srl Roma.
PossinGhaM h. P., anDelMan s. j., burGMan M. a., MeDellín r. a., Master l. l. & keith D. a. (2002): Limits to the use of threatened species lists. – Trends Ecol. Evol. 17: 503–507.

318
Gabrielová et al.: Ohrožení C1 rostlin ČR v jiných evropských zemích?
Postolache G. (1995): Vegetatia Republicii Moldova. – Academia de stiinte a Republicii Moldova, Institul de Botanica. Chisinau.
Pravilnik o uvrstitvi oGroženih rastlinskih in živalskih vrst v rDeČi seznaM (Uradni list Republike Slovenije, št. 82/2002 z dne 24. 9. 2002). Priloga 1: Rdeči seznam praprotnic in semenk (Pteridophyta & Spermatophyta). – URL: http://www.uradni-list.si/1/content?id=38615 (cit. 5. 2. 2011).
Procházka f. [ed.] (2001): Černý a červený seznam cévnatých rostlin České republiky (stav v roce 2000). – Příroda, Praha 18: 1–166.
roDriGues a. s. l., PilGriM j. D., laMoreuX j. f., hoffMann M. & brooks t. M. (2006): The value of the IUCN Red List for conservation. – Trends Ecol. Evol. 21: 71–76.
saintenoy-siMon j., barbier y., Delescaille l.-M., Dufrêne M., Gathoye j.-l. & verté P. (2006): Première liste des espèces rares, menacées et protégées de la Région Wallonne (Ptéridophytes et Spermatophytes). [First list of rare, endangered and protected species of Wallonia (Pteridophytes and Spermatophytes).] Version 1 (7/3/2006). – URL: http://biodiversite.wallonie.be/especes/ecologie/plantes/listerouge/ (cit. 5. 2. 2011).
sharrock s. & jones M. (2009): Conserving Europexs threatended plants: Progress towards Target 8 of the Global Strategy for Plant Conservation. – Botanic Gardens Conservation International, Richmond, U.K.
shelyaG-sosonko yu. r. [ed.] (1996): The Red Data Book of Ukraine. Vegetable Kingdom. Ukrainska Enciklopedia, Kyiv. – URL: http://mail.menr.gov.ua/publ/redbook/redbook.php?action=zmist2 (cit. 5. 2. 2011).
scheMbri P.j. & sultana j. (1989): Red Data Book for the Maltese Islands. – Environment Division Ministry of Education.
schnittler M. & Günther k.-f. (1999): Central European vascular plants requiring priority conservation measures – an analysis from national Red Lists and distribution maps. – Biodivers. Conserv. 8: 891–925.
schulz r. (1904): Monographie der Gattung Phyteuma. – Geisenheim am Rhein.silic c. (1996): Spisak biljnih vrsta (Pteridohpyta i Spermatophyta) za crvenu knjigu Bosne
i Hercegovine. – Zemaljski Museum of Bosnia Herzegovina, Sarajevo.slavík b. [ed.] (1995): Květena České republiky. Vol. 4. – Academia, Praha.slavík b. [ed.] (1997): Květena České republiky. Vol. 5. – Academia, Praha.slavík b. [ed.] (2000): Květena České republiky. Vol. 6. – Academia, Praha.slavík b. & ŠtěPánková j. [eds] (2004): Květena České republiky. Vol. 7. – Academia, Praha.stace c. a., ellis r. G., kent D. h. & Mccosh D. j. (2003): Vice-county census catalogue of the
vascular plants of Great Britain, the Isle of Man and the Channel Islands edited by published by the Botanical Society of the British Isles, London. – URL: http://www.botanicalkeys.co.uk/flora/vccc/index.html (cit. 5. 2. 2011).
stanDaarDlijst neDerlanDse flora 2003. – URL: http://www.floron.nl/ (cit. 5. 2. 2011).stevanoviČ v. b. [ed.] (1999): Crvena knjiga flore Srbije 1. Isčezli i ugroženi taksoni. – Ministarstvo
za životnu sredinu RS, Beograd.stoltze M. & Pihl s. [eds] (1998): Rødliste 1997 over planter og dyr i Danmark [Red List 1997 of
plants and animals in Denmark]. Miljø- og Energiministeriet, Danmarks Miljøundersøgelser og Skov- og Naturstyrelsen. [Ministry of Environment and Energy, National Environmental Research Institute, National Forest and Nature Agency]. – URL: http://www.sns.dk/1pdf/rodlis.pdf (cit. 5. 2. 2011).
striD a. & tan k. [eds] (1997): Flora Hellenica. Vol. 1. – Koeltz Scientific Books, Königstein.striD a. & tan k. [eds] (2002): Flora Hellenica. Vol. 2. – A. R. Gantner Verlag. Ruggell.ŠtěPánková j. [ed.] (2010): Květena České republiky. Vol. 8. – Academia, Praha.tabaka l. v., Gavrilova G. b. & fatare i. ya. (1988): Flora sosudistych rastenij Latvijskoj SSR.
[Flora of vascular plants of the Latvian SSR]. – Zinatne publishing house, Riga.tela botanica. Le réseau de la botanique francophone. – URL: http://www.tela-botanica.org/
(cit. 5. 2. 2011).

319
Příroda, Praha, 31: 299–343, 2011
trPin D. (1995): Register flore Slovenije. Praprotnice in cvetnice. [Register of the flora of Slovenia. Ferns and vascular plants]. – Ljubljana: Znanstvenoraziskovalni center SAZU.
uicn france, Mnhn, fcbn & sfo (2010): La Liste rouge des espéces menacées en France. Chapitre Orchidées de France métropolitaine. – Paris, France.
van lanDuyt w., vanhecke l. & hoste i. (2006). Rode Lijst van de vaatplanten van Vlaanderen en het Brussels Hoofdstedelijk Gewest. [Red list of vascular plants of Flanders and region of Brussel]. – In: van lanDuyt w., hoste i., vanhecke l., van Den breMt P., vercruysse e. & De beer D. [eds], Atlas van de Flora van Vlaanderen en het Brussels Gewest. INBO en Nationale Plantentuin van België, Brussel. URL: http://www.inbo.be/content/page.asp?pid=BEL_VLA_SOO_rodelijst (cit. 5. 2. 2011).
vanGjeli j., ruci b. & Mullaj a. (1995): Libri i kuq. Bimët e kërcënuara e të rralla të Shqipërisë. [Red Book. Threatened and rare plants species of Albania.] – Tiranë.
velchev v., kozuharov s., bonDev i., kuzManov b. & Markova M. [eds] (1984): Red Data Book of the Peoplexs Republic of Bulgaria: Volume 1, Plants. – Publishing House of the Bulgarian Academy of Sciences, Sofia, Bulgaria.
vlaDiMirov v. (2008): List of vascular plants that will be included in the new edition of the Bulgarian Red Data Book. Vol. 1. Plants and Fungi. – Ms.
vyhláŠka 395/1992 Sb. ministerstva životního prostředí České republiky ze dne 11. června 1992, kterou se provádějí některá ustanovení zákona České národní rady č. 114/1992 Sb., o ochraně přírody a krajiny. – URL: http://www.mzp.cz/www/platnalegislativa.nsf/ (cit. 5. 2. 2011).
walter k. s. & Gillett h. j. [eds] (1998): 1997 IUCN Red List of threatened plants. Compiled by the World Conservation Monitoring Centre. – IUCN - The World Conservation Union, Gland, Switzerland and Cambridge, U.K.
winD P. & Pihl s. [eds] (2004): Danish vascular plant species. – In: winD P. & Pihl s. [eds], The Danish Red List. The National Environmental Research Institute, Aarhus University. URL: http://redlist.dmu.dk (updated April 2010) (cit. 5. 2. 2011).

320
Gabrielová et al.: Ohrožení C1 rostlin ČR v jiných evropských zemích?
Příloha 1. Seznam zemí, jejich červených seznamů (červených knih a seznamů druhů chráněných zákonem) a checklistů.Appendix 1. List of countries, their redlists (red data books, lists of species protected by law) and checklists.
stát Červený seznam checklist/Flóra
Albánie vanGjeli et al. (1995) flora euroPaea Database
Belgie saintenoy-siMon et al. (2006); van lanDuyt et al. (2006) laMbinon et al. (2008)
Bulharsko velchev et al. (1984); vlaDiMirov (2008) assyov et al. (2006)
dánsko stoltze & Pihl (1998) winD & Pihl (2004)
Estonsko coMMission for nature conservation of the estonian acaDeMy of sciences (2001–2002) kukk (1999)
Finsko finnish environMent institute (2001) häMet-ahti et al. (1986)
Francie olivier et al. (1995), uicn france 2010 tela botanica, Guinochet & vilMorin (1973)
chorvatsko nikoli (2010) nikoli (2010)
irsko curtis & McGouGh (1988) flora euroPaea Database
island inGaDóttir et al. (1996) flora euroPaea Database
itálie conti et al. (1997) conti et al. (2005)
litva anDrushaitis (1985) tabaka et al. (1988)
lotyšsko lietuvos resPublikos aPlinkos Ministro (2003) GuDžinskas (1999)
lucembursko collinG (2005) collinG (2005)
Maďarsko király (2007) király (2009)
Malta scheMbri & sultana (1989) MifsuD (2002/2010)
německo bunDesaMt für naturschutz: florawebbunDesaMt für naturschutz: floraweb
nizozemí MeijDen et al. (2000) stanDaarDlijst neDerlanDse flora 2003
norsko kÅlÅs et al. (2006) liD & liD (2005)
Polsko kazMierczakowa & zarzycki (2001), Mirek et al. (2006) Mirek et al. (2002)
Portugalsko alves (2001) flora iberica
Rakousko niklfelD (1999) fisher et al. (2008)
Rumunsko oltean et al. (1994) nyáráDy & savulescu (1952–1976)
Řecko Phitos et al. (1995) striD & tan (1997, 2002)
slovensko MarholD & hinDák (1997) MarholD et al. (2007)
slovinsko Pravilnik o uvrstitvi oGroženih rastlinskih in živalskih vrst v rDeČi seznaM
trPin (1995)
Španělsko Moreno (2008) flora iberica
Švédsko GärDenfors (2005) karlsson (2004)
Švýcarsko Moser et al. (2002) aeschiMann & heitz (2005)
ukrajina shelyaG-sosonko (1996) Mosyakin & feDoronchuk (1999)
velká Británie cheffinGs & farrell (2005) stace et al. (2003)
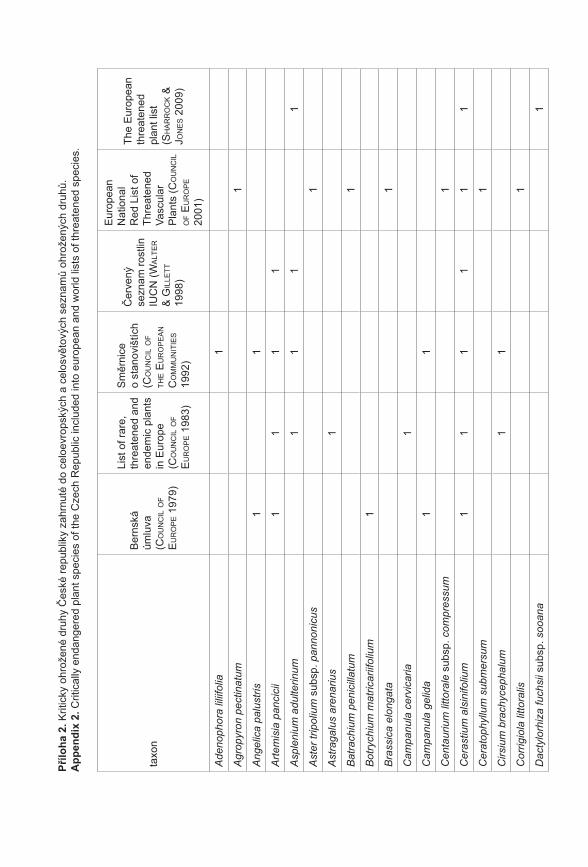
321
Příroda, Praha, 31: xx–xxx, 2011Př
íloha
2. K
ritic
ky o
hrož
ené
druh
y Č
eské
repu
blik
y za
hrnu
té d
o ce
loev
rops
kých
a c
elos
věto
vých
sez
nam
ů oh
rože
ných
dru
hů.
App
endi
x 2.
Crit
ical
ly e
ndan
gere
d pl
ant s
peci
es o
f the
Cze
ch R
epub
lic in
clud
ed in
to e
urop
ean
and
wor
ld li
sts
of th
reat
ened
spe
cies
.
taxo
n
Ber
nská
úm
luva
(c
ou
nc
il o
f e
ur
oP
e 1
979)
List
of r
are,
th
reat
ened
and
en
dem
ic p
lant
s in
Eur
ope
(co
un
cil
of
eu
ro
Pe 1
983)
Sm
ěrni
ce
o st
anov
ištíc
h (c
ou
nc
il o
f th
e e
ur
oP
ea
n
co
MM
un
itie
s
1992
)
Čer
vený
se
znam
rost
lin
IUC
N (w
alt
er
& G
ille
tt
1998
)
Eur
opea
n N
atio
nal
Red
Lis
t of
Thre
aten
ed
Vasc
ular
P
lant
s (c
ou
nc
il
of
eu
ro
Pe
2001
)
The
Eur
opea
n th
reat
ened
pl
ant l
ist
( sh
ar
ro
ck &
jo
ne
s 2
009)
Ade
noph
ora
liliif
olia
1
Agr
opyr
on p
ectin
atum
1
Ang
elic
a pa
lust
ris
11
Arte
mis
ia p
anci
cii
11
11
Asp
leni
um a
dulte
rinum
1
11
1
Ast
er tr
ipol
ium
sub
sp. p
anno
nicu
s 1
Ast
raga
lus
aren
ariu
s 1
Bat
rach
ium
pen
icill
atum
1
Bot
rych
ium
mat
ricar
iifol
ium
1
Bra
ssic
a el
onga
ta
1
Cam
panu
la c
ervi
caria
1
Cam
panu
la g
elid
a 1
1
Cen
taur
ium
litto
rale
sub
sp. c
ompr
essu
m
1
Cer
astiu
m a
lsin
ifoliu
m
11
11
11
Cer
atop
hyllu
m s
ubm
ersu
m
1
Cirs
ium
bra
chyc
epha
lum
1
1
Cor
rigio
la li
ttora
lis
1
Dac
tylo
rhiz
a fu
chsi
i sub
sp. s
ooan
a 1
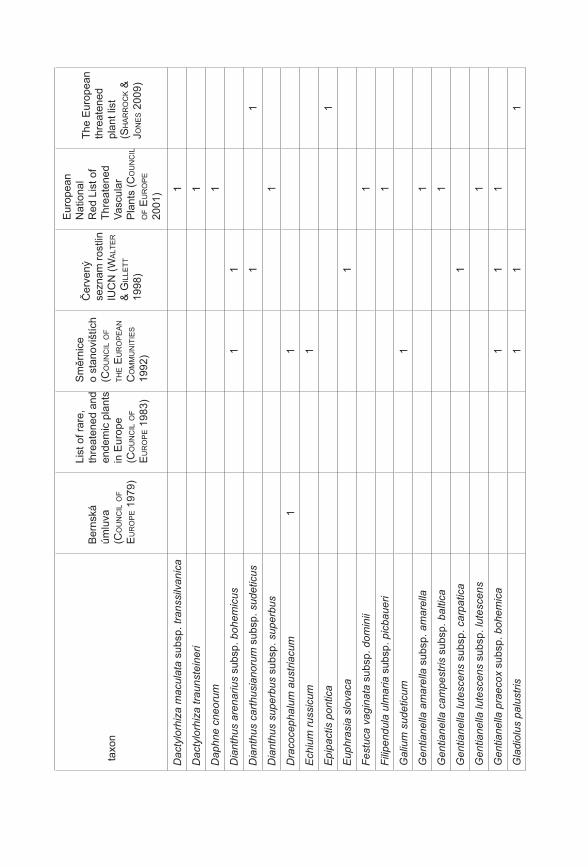
322
Liška: Česká jména lišejníků
taxo
n
Ber
nská
úm
luva
( c
ou
nc
il o
f e
ur
oP
e 1
979)
List
of r
are,
th
reat
ened
and
en
dem
ic p
lant
s in
Eur
ope
( co
un
cil
of
eu
ro
Pe 1
983)
Sm
ěrni
ce
o st
anov
ištíc
h ( c
ou
nc
il o
f th
e e
ur
oP
ea
n
co
MM
un
itie
s
1992
)
Čer
vený
se
znam
rost
lin
IUC
N ( w
alt
er
& G
ille
tt
1998
)
Eur
opea
n N
atio
nal
Red
Lis
t of
Thre
aten
ed
Vasc
ular
P
lant
s (c
ou
nc
il
of
eu
ro
Pe
2001
)
The
Eur
opea
n th
reat
ened
pl
ant l
ist
( sh
ar
ro
ck &
j o
ne
s 2
009)
Dac
tylo
rhiz
a m
acul
ata
subs
p. tr
anss
ilvan
ica
1
Dac
tylo
rhiz
a tra
unst
eine
ri 1
Dap
hne
cneo
rum
1
Dia
nthu
s ar
enar
ius
subs
p. b
ohem
icus
1
1
Dia
nthu
s ca
rthus
iano
rum
sub
sp. s
udet
icus
1
1
Dia
nthu
s su
perb
us s
ubsp
. sup
erbu
s 1
Dra
coce
phal
um a
ustri
acum
1
1
Ech
ium
russ
icum
1
Epi
pact
is p
ontic
a 1
Eup
hras
ia s
lova
ca
1
Fest
uca
vagi
nata
sub
sp. d
omin
ii 1
Filip
endu
la u
lmar
ia s
ubsp
. pic
baue
ri1
Gal
ium
sud
etic
um
1
Gen
tiane
lla a
mar
ella
sub
sp. a
mar
ella
1
Gen
tiane
lla c
ampe
stris
sub
sp. b
altic
a 1
Gen
tiane
lla lu
tesc
ens
subs
p. c
arpa
tica
1
Gen
tiane
lla lu
tesc
ens
subs
p. lu
tesc
ens
1
Gen
tiane
lla p
raec
ox s
ubsp
. boh
emic
a 1
11
Gla
diol
us p
alus
tris
11
1
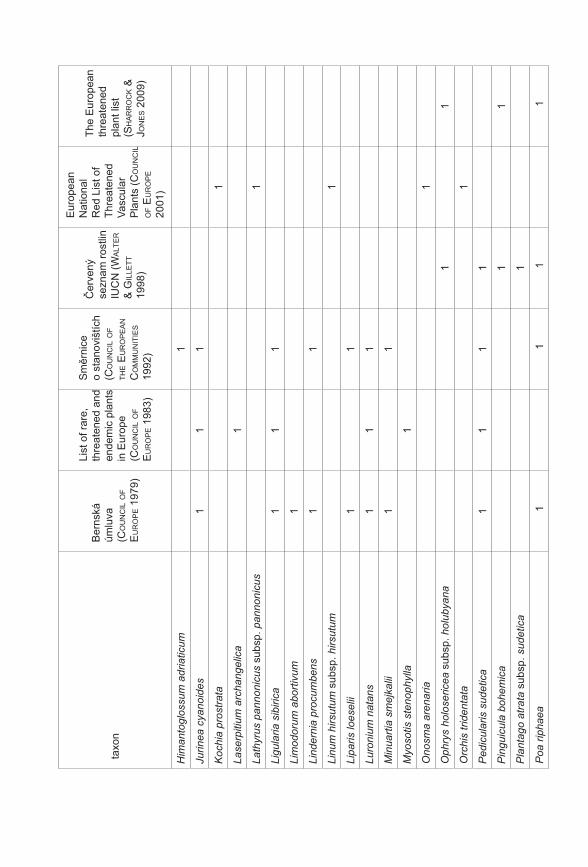
323
Příroda, Praha, 31: xx–xxx, 2011
taxo
n
Ber
nská
úm
luva
(c
ou
nc
il o
f e
ur
oP
e 1
979)
List
of r
are,
th
reat
ened
and
en
dem
ic p
lant
s in
Eur
ope
( co
un
cil
of
eu
ro
Pe 1
983)
Sm
ěrni
ce
o st
anov
ištíc
h (c
ou
nc
il o
f th
e e
ur
oP
ea
n
co
MM
un
itie
s
1992
)
Čer
vený
se
znam
rost
lin
IUC
N ( w
alt
er
& G
ille
tt
1998
)
Eur
opea
n N
atio
nal
Red
Lis
t of
Thre
aten
ed
Vasc
ular
P
lant
s ( c
ou
nc
il
of
eu
ro
Pe
2001
)
The
Eur
opea
n th
reat
ened
pl
ant l
ist
( sh
ar
ro
ck &
jo
ne
s 2
009)
Him
anto
glos
sum
adr
iatic
um
1
Jurin
ea c
yano
ides
1
11
Koc
hia
pros
trata
1
Lase
rpiti
um a
rcha
ngel
ica
1
Lath
yrus
pan
noni
cus
subs
p. p
anno
nicu
s 1
Ligu
laria
sib
irica
1
11
Lim
odor
um a
borti
vum
1
Lind
erni
a pr
ocum
bens
1
1
Linu
m h
irsut
um s
ubsp
. hirs
utum
1
Lipa
ris lo
esel
ii 1
1
Luro
nium
nat
ans
11
1
Min
uarti
a sm
ejka
lii
11
Myo
sotis
ste
noph
ylla
1
Ono
sma
aren
aria
1
Oph
rys
holo
seric
ea s
ubsp
. hol
ubya
na
11
Orc
his
tride
ntat
a 1
Ped
icul
aris
sud
etic
a 1
11
1
Pin
guic
ula
bohe
mic
a 1
1
Pla
ntag
o at
rata
sub
sp. s
udet
ica
1
Poa
riph
aea
11
11
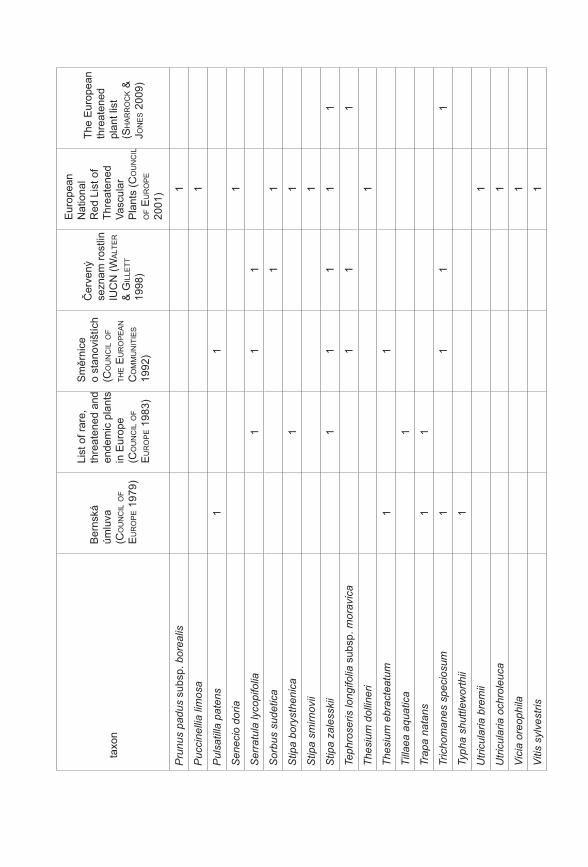
324
Liška: Česká jména lišejníků
taxo
n
Ber
nská
úm
luva
(c
ou
nc
il o
f e
ur
oP
e 1
979)
List
of r
are,
th
reat
ened
and
en
dem
ic p
lant
s in
Eur
ope
( co
un
cil
of
eu
ro
Pe 1
983)
Sm
ěrni
ce
o st
anov
ištíc
h (c
ou
nc
il o
f th
e e
ur
oP
ea
n
co
MM
un
itie
s
1992
)
Čer
vený
se
znam
rost
lin
IUC
N ( w
alt
er
& G
ille
tt
1998
)
Eur
opea
n N
atio
nal
Red
Lis
t of
Thre
aten
ed
Vasc
ular
P
lant
s (c
ou
nc
il
of
eu
ro
Pe
2001
)
The
Eur
opea
n th
reat
ened
pl
ant l
ist
( sh
ar
ro
ck &
jo
ne
s 2
009)
Pru
nus
padu
s su
bsp.
bor
ealis
1
Puc
cine
llia
limos
a 1
Pul
satil
la p
aten
s 1
1
Sen
ecio
dor
ia
1
Ser
ratu
la ly
copi
folia
1
11
Sor
bus
sude
tica
11
Stip
a bo
ryst
heni
ca
11
Stip
a sm
irnov
ii 1
Stip
a za
less
kii
11
11
1
Teph
rose
ris lo
ngifo
lia s
ubsp
. mor
avic
a 1
11
Thes
ium
dol
liner
i 1
Thes
ium
ebr
acte
atum
1
1
Tilla
ea a
quat
ica
1
Trap
a na
tans
1
1
Tric
hom
anes
spe
cios
um
11
11
Typh
a sh
uttle
wor
thii
1
Utri
cula
ria b
rem
ii 1
Utri
cula
ria o
chro
leuc
a 1
Vici
a or
eoph
ila
1
Vitis
syl
vest
ris
1

325
Příroda, Praha, 31: xx–xxx, 2011Př
íloha
3. K
ritic
ky o
hrož
ené
druh
y Č
R a
jejic
h oh
rože
ní v
dal
ších
evr
opsk
ých
zem
ích.
AL
– A
lbán
ie, B
– B
elgi
e, B
G –
Bul
hars
ko, D
K –
Dán
sko,
ES
T –
Est
onsk
o, F
IN –
Fin
sko,
F –
Fra
ncie
, HR
– C
horv
atsk
o, IR
L –
Irsko
, IS
– Is
land
, I –
Itá
lie, L
T –
Litv
a, L
V –
Lot
yšsk
o, L
– L
ucem
burs
ko, H
– M
aďar
sko,
M –
Mal
ta, D
– N
ěmec
ko, N
L –
Niz
ozem
í, N
– N
orsk
o, P
L –
Pol
sko,
P –
Por
tuga
lsko
, A
– R
akou
sko,
RO
– R
umun
sko,
GR
– Ř
ecko
, SK
– S
love
nsko
, SLO
– S
lovi
nsko
, E –
Špa
něls
ko, S
WE
– Š
véds
ko, C
H –
Švý
cars
ko, U
A –
Ukr
ajin
a,
UK
– V
elká
Brit
ánie
.E
X –
vyh
ynul
ý, E
X?
– pr
avdě
podo
bně
vyhy
nulý
, C
R –
krit
icky
ohr
ožen
ý, E
N –
ohr
ožen
ý, V
U –
zra
nite
lný,
NT
– té
měř
ohr
ožen
ý, R
– v
zácn
ý, I
–
neza
řaze
ný (d
ruh
spad
ajíc
í do
někt
eré
z ka
tego
rií E
X, C
R, E
N, V
U n
ebo
R, p
ro s
tano
vení
pře
snéh
o st
upně
ohr
ožen
í vša
k ch
ybí d
ata)
, LC
/LR
– m
éně
ohro
žený
, DD
– c
hybě
jící d
ata,
NE
– n
evyh
odno
cený
, ? –
výs
kyt p
ochy
bný,
§ –
dru
h ch
ráně
ný z
ákon
em d
ane
zem
ě, 1
– d
ruh
se v
zem
i vys
kytu
je, j
e bě
žný,
prá
zdné
– d
ruh
se v
zem
i nev
ysky
tuje
E –
end
emit
Čes
ké re
publ
iky,
§1,
§2,
§3
– dr
uh c
hrán
ěný
záko
nem
v Č
R (V
yhlá
ška
MŽP
395
/199
2 S
b.):
§1 –
krit
icky
ohr
ožen
ý, §
2 –
siln
ě oh
rože
ný,
§3 –
ohr
ožen
ý, C
E –
dru
hy z
ahrn
uté
do d
o S
ezna
mu
stře
doev
rops
kých
rost
lin v
yžad
ujíc
ích
prio
ritní
och
raná
řská
opa
tření
(sc
hn
ittl
er &
Gü
nth
er 1
999)
.
App
endi
x 3.
Crit
ical
ly e
ndan
gere
d sp
ecie
s of
the
Cze
ch R
epub
lic a
nd th
eir e
ndan
germ
ent i
n ot
her E
urop
ean
coun
tries
. A
L –
Alb
ania
, B –
Bel
gium
, BG
– B
ulga
ria, D
K –
Den
mar
k, E
ST
– E
ston
ia, F
IN –
Fin
land
, F –
Fra
nce,
HR
– C
roat
ia, I
RL
– Ire
land
, IS
– Ic
elan
d, I
– Ita
ly, L
T –
Lith
uani
a, L
V –
Lat
via,
L –
Lux
embo
urg,
H –
Hun
gary
, M –
Mal
ta, D
– G
erm
any,
NL
– N
ethe
rland
s, N
– N
orw
ay, P
L –
Pol
and,
P –
Por
tuga
l, A
– A
ustri
a, R
O –
Rom
ania
, GR
– G
reec
e, S
K –
Slo
vaki
a, S
LO –
Slo
veni
a, E
– S
pain
, SW
E –
Sw
eden
, CH
– S
witz
erla
nd, U
A –
Ukr
aine
, UK
– U
nite
d K
ingd
om o
f Gre
at B
ritai
n an
d N
orth
ern
Irela
nd.
EX
– e
xtin
ct, E
X?
– pr
obab
ly e
xtin
ct, C
R –
crit
ical
ly e
ndan
gere
d, E
N –
end
ange
red,
VU
– v
ulne
rabl
e, N
T –
near
thre
aten
ed, R
– ra
re, I
– in
dete
rmin
ate
(spe
cies
kno
wn
to b
e ex
tinct
, end
ange
red,
vul
nera
ble
or r
are,
but
who
se d
egre
e of
bei
ng e
ndan
gere
d ca
nnot
be
spec
ified
due
to in
suffi
cien
t dat
a),
LC/L
R –
leas
t con
cern
/low
er ri
sk, D
D –
dat
a de
ficie
nt, N
E –
not
eva
luat
ed, ?
– o
ccur
renc
e do
ubtfu
l, §
– sp
ecie
s pr
otec
ted
by th
e la
w, 1
– p
rese
nt, n
ot
enda
nger
ed in
the
coun
try, e
mpt
y –
abse
nt in
the
coun
tryE
– s
peci
es e
ndem
ic to
the
Cze
ch R
epub
lic, §
1, §
2, §
3 –
spec
ies
prot
ecte
d by
the
law
in th
e C
zech
Rep
ublic
(Dec
ree
of th
e M
inis
try o
f the
Env
ironm
ent
No.
395
/199
2 C
oll.)
: §1
– cr
itica
lly e
ndan
gere
d, §
2 –
stro
ngly
end
ange
red,
§3
– en
dang
ered
, CE
– s
peci
es re
cord
ed in
the
List
of C
entra
l Eur
opea
n va
scul
ar p
lant
s re
quiri
ng p
riorit
y co
nser
vatio
n m
easu
res
(sc
hn
ittl
er &
Gü
nth
er 1
999)
.
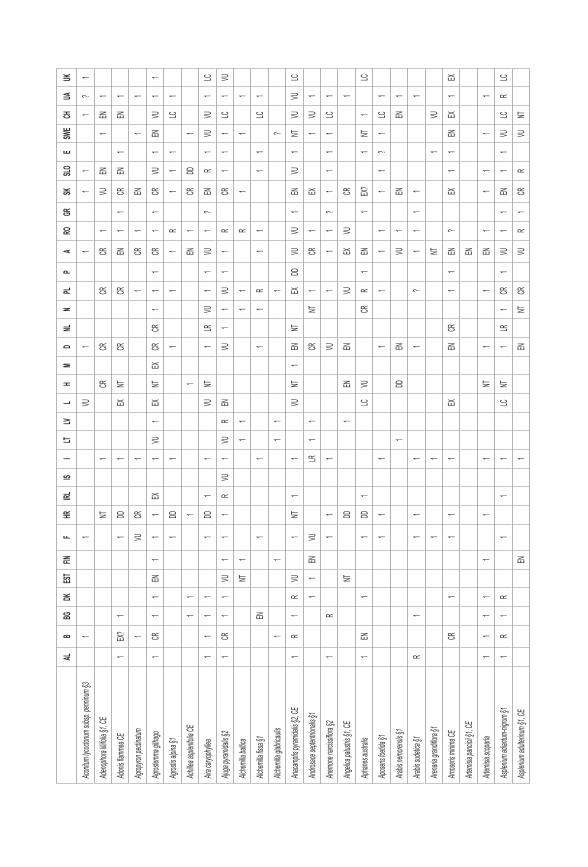
326
Liška: Česká jména lišejníků
AlB
BGdK
Est
FinF
HRiRl
isi
ltlv
lH
Md
nln
PlP
ARo
GRsK
slo
EsW
EcH
uAuK
Aconitu
m lyco
ctonum
subsp
. penni
num §3
11
VU1
11
11
?1
Adeno
phora l
iliifolia
§1, CE
NT1
CRCR
CRCR
1VU
EN1
EN1
Adoni
s flam
mea C
E1
EX?
11
DD1
EXNT
CRCR
EN1
1CR
EN1
EN1
Agrop
yron p
ectina
tum
1VU
CR1
1CR
1EN
11
Agros
temma
githag
o 1
CR1
1EN
11
1EX
1VU
1EX
NTEX
CRCR
11
1CR
11
CRVU
1EN
VU1
1
Agros
tis alp
ina §1
1DD
11
11
R1
11
LC1
Achille
a aspl
enifoli
a CE
11
11
EN1
CRDD
1
Aira c
aryoph
yllea
11
11
1DD
11
VUNT
1LR
VU1
1VU
1?
ENR
1VU
VU1
LC
Ajuga
pyram
idalis
§21
CR1
1VU
11
1R
VU1
VUR
ENVU
11
VU1
1R
CR1
11
LC1
VU
Alchem
illa ba
ltica
NT1
11
11
R1
11
Alchem
illa fis
sa §1
EN1
11
1R
11
11
LC1
Alchem
illa gla
bricaul
is1
11
11
?
Anaca
mptis
pyram
idalis
§2, CE
1R
1R
VU1
NT1
1VU
NT1
ENNT
EXDD
VUVU
1EN
VU1
NTVU
VULC
Andro
sace s
eptent
rionalis
§11
1EN
VULR
11
CRNT
1CR
1EX
1VU
1
Anem
one na
rcissifl
ora §2
1R
11
1VU
11
1?
11
11
LC1
Angel
ica pa
lustris
§1, C
ENT
DD1
ENEN
VUEX
VUCR
1
Aphan
es aus
tralis
1EN
11
DD1
LCVU
CRR
1EN
1EX
?1
NT1
LC
Apose
ris foe
tida §1
11
11
11
11
1?
1LC
1
Arabis
nemo
rensis
§11
DDEN
VU1
ENEN
1
Arabis
sudet
ica §1
R1
11
11
?1
11
11
Arenar
ia gran
diflora
§11
1NT
1VU
Arnose
ris min
ima CE
CR1
11
1EX
ENCR
11
EN?
EX1
1EN
EX1
EX
Artem
isia pa
ncicii
§1, CE
EN
Artem
isia sc
oparia
11
11
11
1NT
11
EN1
11
11
Asplen
ium ad
iantum
-nigrum
§11
R1
R1
11
LCNT
1LR
1CR
1VU
11
EN1
1VU
LCR
LC
Asplen
ium ad
ulterinu
m §1, C
EEN
1EN
NTCR
VUR
1CR
RVU
NT
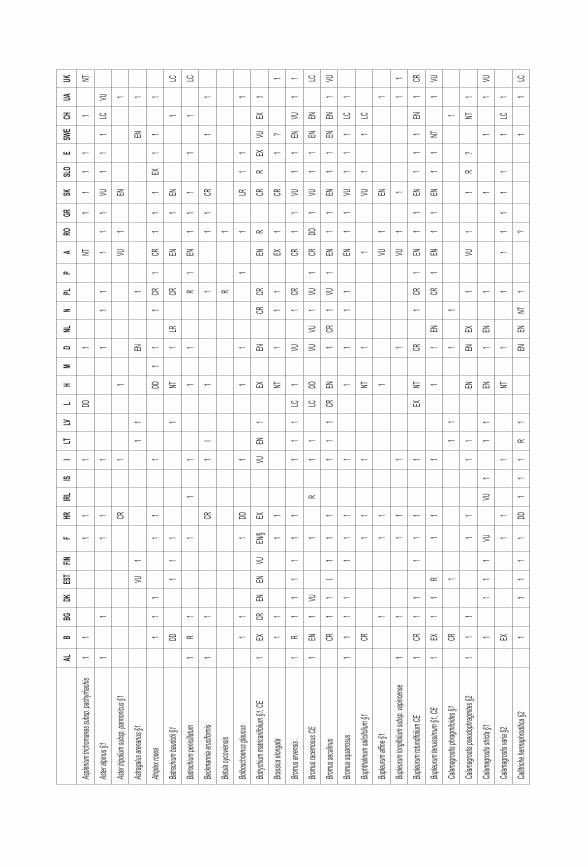
327
Příroda, Praha, 31: xx–xxx, 2011
AlB
BGdK
Est
FinF
HRiRl
isi
ltlv
lH
Md
nln
PlP
ARo
GRsK
slo
EsW
EcH
uAuK
Asplen
ium tric
homane
s subs
p. pach
yrhach
is 1
11
11
DD1
NT1
11
11
NT
Aster a
lpinus
§11
11
11
11
11
11
VU1
11
LCVU
Aster t
ripolium
subsp
. panno
nicus
§1CR
11
VU1
EN1
Astrag
alus a
renariu
s §1
VU1
11
EN1
EN1
Atriple
x rosea
1
11
11
1DD
11
1CR
1CR
11
1EX
11
1
Batrac
hium b
audotii
§1DD
11
11
NT1
LRCR
EN1
EN1
LC
Batrac
hium p
enicilla
tum
1R
11
11
11
R1
EN1
11
11
LC
Beckm
annia e
rucifor
mis
11
CR1
I1
11
1CR
11
Betula
oycov
iensis
R
1
Bolbo
schoen
us gla
ucus
11
1DD
11
11
1LR
11
1
Botryc
hium m
atricar
iifolium
§1, C
E1
EXCR
ENEN
VUEN
/§EX
VUEN
1EX
ENCR
CREN
RCR
REX
VUEX
1
Brassi
ca elo
ngata
11
11
NT1
11
EX1
CR1
?1
Bromu
s arve
nsis
1R
11
11
11
11
1LC
1VU
1CR
CR1
1VU
11
ENVU
11
Bromu
s racem
osus C
E1
EN1
VU1
R1
1LC
DDVU
VU1
VU1
CRDD
1VU
11
ENEN
LC
Bromu
s seca
linus
CR1
1I
11
11
11
CREN
1CR
1VU
1EN
11
EN1
1EN
EN1
VU
Bromu
s squa
rrosus
1
11
11
11
11
11
1EN
11
VU1
11
LC1
Bupht
halmu
m salic
ifolium
§1CR
11
1NT
11
VU1
1LC
Buple
urum a
ffine §
11
11
1VU
1EN
1
Buple
urum l
ongifol
ium su
bsp. va
pincen
se 1
11
11
1VU
11
11
Buple
urum r
otundi
folium
CE1
CR1
11
11
1EX
NTCR
1CR
1EN
11
EN1
11
EN1
CR
Buple
urum t
enuiss
imum §
1, CE
1EX
11
R1
11
11
ENCR
1EN
11
EN1
1NT
1VU
Calam
agrost
is phra
gmitoi
des §1
CR1
11
11
1
Calam
agrost
is pseu
dophra
gmites
§21
11
11
11
ENEN
EX1
VU1
1R
?NT
1
Calam
agrost
is stric
ta §1
11
11
VUVU
11
1EN
1EN
11
11
VU
Calam
agrost
is varia
§2EX
11
1NT
11
11
11
11
LC1
Callitr
iche h
ermaph
roditic
a §2
11
11
1DD
11
1R
1EN
ENNT
1?
11
LC
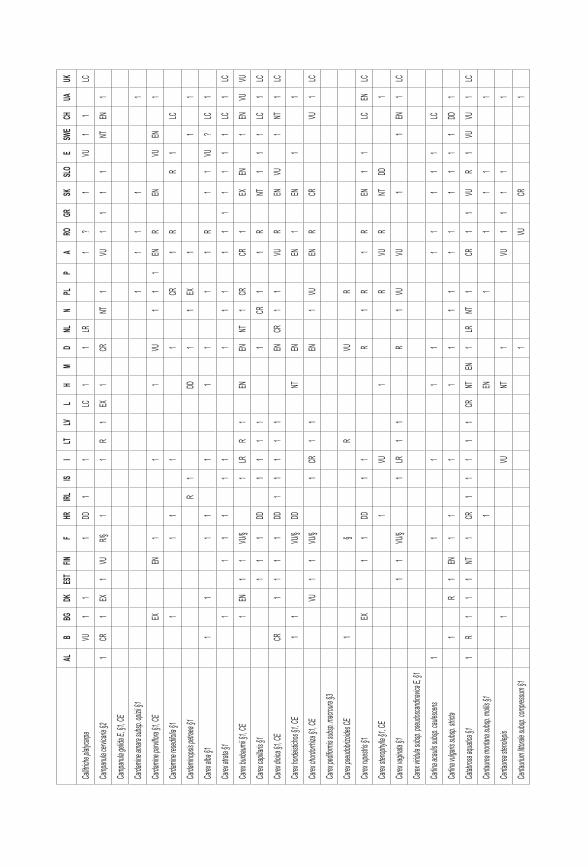
328
Liška: Česká jména lišejníků
AlB
BGdK
Est
FinF
HRiRl
isi
ltlv
lH
Md
nln
PlP
ARo
GRsK
slo
EsW
EcH
uAuK
Callitr
iche p
latycar
pa VU
11
1DD
11
LC1
1LR
1?
1VU
11
LC
Camp
anula c
ervica
ria §2
1CR
1EX
1VU
R/§1
1R
1EX
1CR
NT1
VU1
11
1NT
EN1
Camp
anula g
elida E
, §1, C
E
Carda
mine a
mara s
ubsp. o
pizii §
11
11
11
Carda
mine p
arviflo
ra §1, C
EEX
EN1
11
VU1
11
ENR
ENVU
EN1
Carda
mine re
sedifol
ia §1
11
11
1CR
1R
R1
LC
Carda
minops
is petra
ea §1
R1
DD1
1EX
11
1
Carex
alba §
11
11
11
11
11
R1
1VU
?LC
1
Carex
atrata
§11
11
11
11
11
11
11
11
1LC
1LC
Carex
buxba
umii §
1, CE
1EN
11
VU/§
1LR
R1
ENEN
NT1
CRCR
1EX
EN1
ENVU
VU
Carex
capill
aris §1
11
1DD
11
11
1CR
11
RNT
11
1LC
1LC
Carex
dioica
§1, C
ECR
11
11
DD1
11
11
ENCR
11
VUR
ENVU
1NT
1LC
Carex
horde
isticho
s §1, C
E1
1VU
/§DD
NTEN
EN1
EN1
1
Carex
chord
orrhiz
a §1, C
EVU
11
VU/§
1CR
11
EN1
VUEN
RCR
VU1
LC
Carex
pedifo
rmis s
ubsp. m
acrour
a §3
Carex
pseud
obrizo
ides C
E1
§R
VUR
Carex
rupest
ris §1
EX1
1DD
11
R1
R1
REN
11
LCEN
LC
Carex
stenop
hylla §
1, CE
1VU
1R
VUR
NTDD
1
Carex
vagin
ata §1
11
VU/§
1LR
11
R1
VUVU
11
EN1
LC
Carex
viridu
la subs
p. pseu
doscan
dinavi
ca E, §
1
Carlin
a acau
lis sub
sp. ca
ulesce
ns 1
11
11
11
11
1LC
Carlin
a vulg
aris su
bsp. st
ricta
1R
1EN
11
11
11
11
11
11
1DD
1
Catab
rosa a
quatica
§11
R1
11
NT1
CR1
11
11
CRNT
EN1
LRNT
1CR
11
VUR
1VU
VU1
LC
Centa
urea m
ontana
subsp
. molli
s §1
1EN
11
11
1
Centa
urea s
tenole
pis
1VU
NT1
VU1
11
11
Centa
urium l
ittorale
subsp
. comp
ressum
§11
VUCR
1
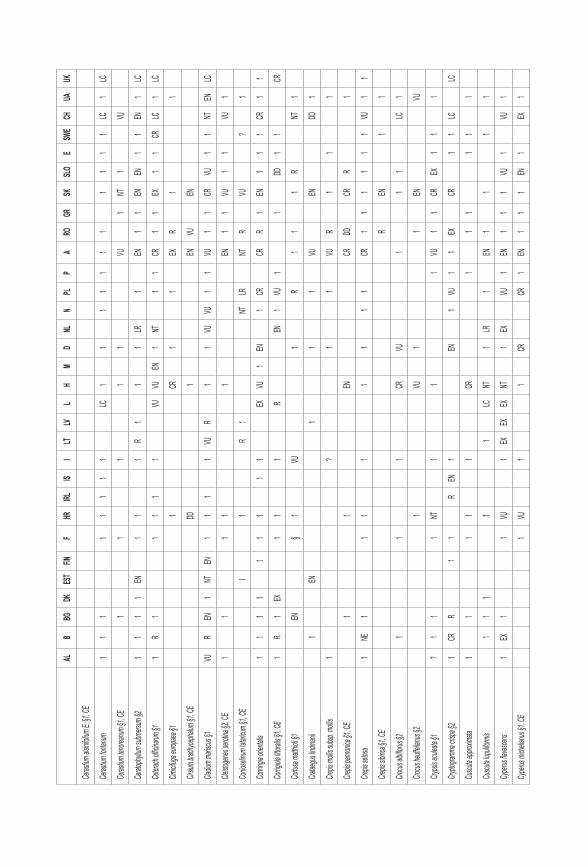
329
Příroda, Praha, 31: xx–xxx, 2011
AlB
BGdK
Est
FinF
HRiRl
isi
ltlv
lH
Md
nln
PlP
ARo
GRsK
slo
EsW
EcH
uAuK
Ceras
tium als
inifoliu
m E, §1
, CE
Ceras
tium fon
tanum
1
11
11
11
1LC
11
11
11
11
11
1LC
1LC
Ceras
tium ten
oreanu
m §1, C
E1
11
11
VU1
NT1
VU
Cerato
phyllum
subm
ersum
§21
11
1EN
11
1R
11
1LR
1EN
11
ENEN
11
EN1
LC
Cetera
ch offi
cinaru
m §1
1R
11
11
1VU
VUEN
1NT
11
CR1
1EX
11
CRLC
1LC
Cimicif
uga eu
ropaea
§11
CR1
1EX
R1
1
Cirsiu
m brac
hyceph
alum §
1, CE
DD1
ENVU
EN
Cladiu
m maris
cus §1
VUR
EN1
NTEN
11
11
VUR
11
VUVU
11
VU1
1CR
VU1
1NT
ENLC
Cleisto
genes
serotin
a §2, C
E1
11
11
EN1
1VU
11
VU1
Conio
selinu
m tata
ricum §
1, CE
I1
R1
NTLR
NTR
VU?
1
Conrin
gia ori
entalis
1
11
11
11
11
EXVU
1EN
1CR
CRR
1EN
11
1CR
11
Corrig
iola litt
oralis
§1, CE
1R
1EX
11
1R
EN1
VU1
1DD
11
CR
Cortus
a matth
ioli §1
EN§
1VU
1R
11
1R
NT1
Cratae
gus lin
dmani
i 1
EN1
11
VUEN
DD1
Crepis
mollis
subsp.
mollis
1?
11
VUR
11
1
Crepis
panno
nica §
1, CE
11
ENCR
DDCR
R1
Crepis
setos
a 1
NE1
11
11
11
1CR
11
11
11
VU1
1
Crepis
sibiric
a §1, C
ER
EN1
1
Crocus
albiflo
rus §2
11
1CR
VU1
11
LC1
Crocus
heuffe
lianus
§21
VU1
1EN
VU
Crypsi
s acul
eata §
1?
11
1NT
11
1VU
11
CREX
11
1
Crypto
gramm
a crisp
a §2
1CR
R1
1R
EN1
EN1
VU1
1EX
CR1
1LC
LC
Cuscu
ta appr
oxima
ta 1
11
11
CR1
11
11
1
Cuscu
ta lupu
liformi
s 1
11
11
LCNT
1LR
1EN
11
11
Cyper
us flav
escens
1
EX1
1VU
1EX
EXEX
NT1
EXVU
1EN
11
1VU
1VU
1
Cyper
us mic
helian
us §1,
CE1
VU1
1CR
CR1
EN1
11
EN1
EX1
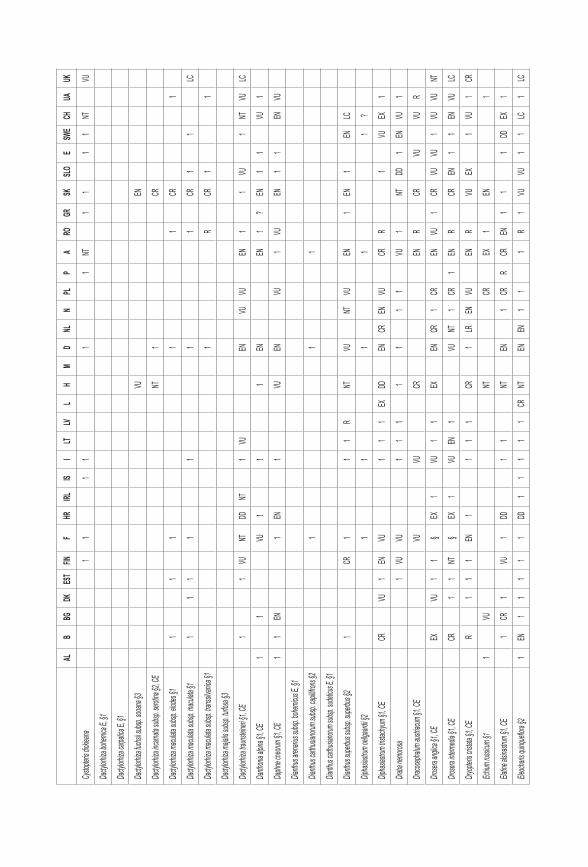
330
Liška: Česká jména lišejníků
AlB
BGdK
Est
FinF
HRiRl
isi
ltlv
lH
Md
nln
PlP
ARo
GRsK
slo
EsW
EcH
uAuK
Cysto
pteris
dickie
ana
11
11
11
NT1
11
1NT
VU
Dacty
lorhiza
bohem
ica E,
§1
Dacty
lorhiza
carpa
tica E,
§1
Dacty
lorhiza
fuchsi
i subsp
. sooan
a §3
VUEN
Dacty
lorhiza
incarn
ata su
bsp. se
rotina
§2, CE
NT1
CR
Dacty
lorhiza
macul
ata su
bsp. el
odes §
11
11
11
CR1
Dacty
lorhiza
macul
ata su
bsp. m
aculata
§11
11
11
11
CR1
1LC
Dacty
lorhiza
macul
ata su
bsp. tra
nssilva
nica §
11
RCR
11
Dacty
lorhiza
majali
s subs
p. turfo
sa §3
Dacty
lorhiza
trauns
teineri
§1, CE
11
VUNT
DDNT
1VU
ENVU
VUEN
11
VU1
NTVU
LC
Danth
onia a
lpina §
1, CE
11
VU1
11
ENEN
1?
EN1
1VU
1
Daphn
e cneo
rum §1
, CE
11
EN1
EN1
VUEN
VU1
VUEN
11
ENVU
Dianth
us are
narius
subsp
. bohem
icus E
, §1
Dianth
us car
thusia
norum
subsp
. capill
ifrons
§21
11
Dianth
us car
thusia
norum
subsp
. sudet
icus E
, §1
Dianth
us sup
erbus
subsp.
super
bus §2
1CR
11
1R
NTVU
NTVU
EN1
EN1
ENLC
Diphas
iastrum
oellga
ardii §
21
11
11
?
Diphas
iastrum
tristac
hyum §
1, CE
CRVU
1EN
VU1
11
EXDD
ENCR
ENVU
CRR
1VU
EX1
Draba
nemoro
sa 1
VUVU
11
11
11
1VU
1NT
DD1
ENVU
1
Dracoc
ephalu
m aust
riacum
§1, C
EVU
VUCR
ENR
CRVU
VUR
Droser
a angl
ica §1
, CE
EXVU
11
§EX
1VU
11
EXEN
CR1
CREN
VU1
CRVU
VU1
VUVU
NT
Droser
a interm
edia §
1, CE
CR1
1NT
§EX
1VU
EN1
VUNT
1CR
1EN
RCR
EN1
1EN
VULC
Dryopt
eris cri
stata §
1, CE
R1
11
EN1
11
1CR
1LR
ENVU
ENR
VUEX
1VU
1CR
Echium
russic
um §1
1VU
NTCR
EX1
EN1
Elatine
alsina
strum §
1, CE
1CR
1VU
1DD
11
NTEN
1CR
RCR
EN1
11
DDEX
1
Eleoch
aris qu
inquefl
ora §2
1EN
11
11
1DD
11
11
1CR
NTEN
EN1
11
R1
VUVU
11
LC1
LC
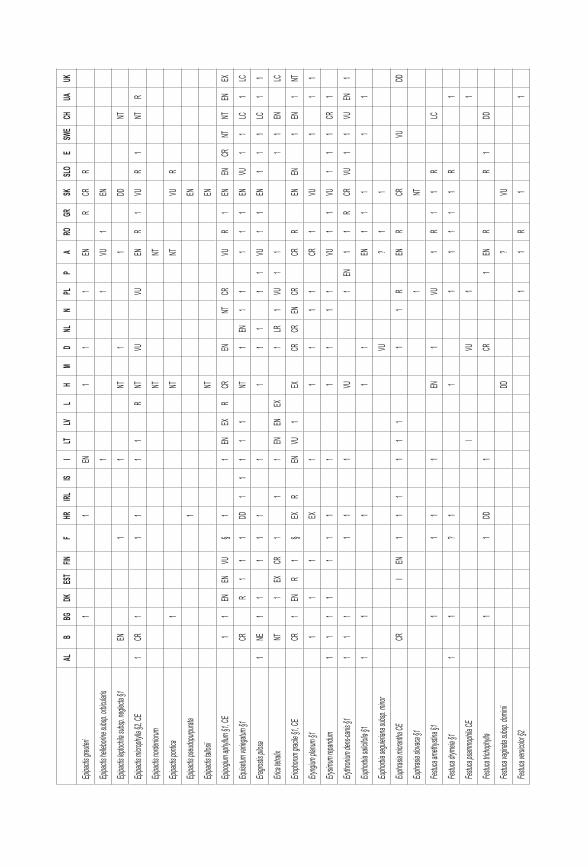
331
Příroda, Praha, 31: xx–xxx, 2011
AlB
BGdK
Est
FinF
HRiRl
isi
ltlv
lH
Md
nln
PlP
ARo
GRsK
slo
EsW
EcH
uAuK
Epipa
ctis gre
uteri
11
EN1
11
ENR
CRR
Epipa
ctis he
lleborin
e subs
p. orbic
ularis
11
VU1
EN
Epipa
ctis lep
tochila
subsp
. negle
cta §1
EN1
1NT
11
DDNT
Epipa
ctis mi
crophy
lla §2,
CE1
CR1
11
11
RNT
VUVU
ENR
1VU
R1
NTR
Epipa
ctis no
rdenio
rum
NTNT
Epipa
ctis po
ntica
1NT
NTVU
R
Epipa
ctis ps
eudopu
rpurata
1
EN
Epipa
ctis tal
losii
NTEN
Epipo
gium a
phyllum
§1, C
E1
1EN
ENVU
§1
1EN
EXR
CREN
NTCR
VUR
1EN
ENCR
NTNT
ENEX
Equis
etum v
ariegat
um §1
CRR
11
1DD
11
11
1NT
1EN
11
11
1EN
VU1
1LC
1LC
Eragro
stis pil
osa
1NE
11
11
11
11
11
1VU
11
EN1
11
LC1
1
Erica
tetralix
NT
1EX
CR1
11
ENEN
EX1
LR1
VU1
11
1EN
LC
Erioph
orum g
racile
§1, CE
CR1
ENR
1§
EXR
ENVU
1EX
CRCR
ENCR
CRR
ENEN
1EN
1NT
Eryngi
um pla
num §1
11
1EX
11
11
1CR
1VU
11
1
Erysim
um rep
andum
1
11
11
11
11
11
1VU
11
VU1
11
CR1
Erythro
nium d
ens-ca
nis §1
11
11
11
VU1
EN1
1R
CRVU
11
VUEN
1
Eupho
rbia sa
licifolia
§11
11
11
EN1
11
11
Eupho
rbia se
guieria
na sub
sp. mi
nor
VU?
11
Euphr
asia m
icranth
a CE
CRI
EN1
11
11
11
1R
ENR
CRVU
DD
Euphr
asia s
lovaca
§11
NT
Festuc
a ame
thystin
a §1
11
11
EN1
VU1
R1
1R
LC
Festuc
a drym
eia §1
11
?1
11
11
11
R1
Festuc
a psam
mophi
la CE
IVU
11
Festuc
a trich
ophylla
1
1DD
1CR
1EN
RR
1DD
Festuc
a vagi
nata s
ubsp. d
ominii
DD
?VU
Festuc
a vers
icolor
§21
1R
11
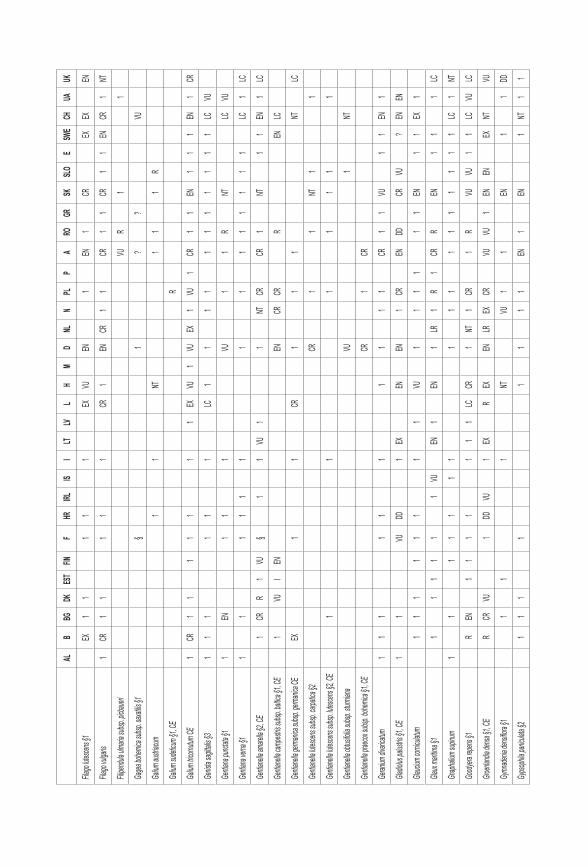
332
Liška: Česká jména lišejníků
AlB
BGdK
Est
FinF
HRiRl
isi
ltlv
lH
Md
nln
PlP
ARo
GRsK
slo
EsW
EcH
uAuK
Filago
lutesce
ns §1
EX1
11
11
EXVU
EN1
EN1
CREX
EXEN
Filago
vulgar
is 1
CR1
11
11
CR1
ENCR
11
CR1
1CR
11
ENCR
1NT
Filipen
dula u
lmaria
subsp.
picbau
eriVU
R1
1
Gagea
bohem
ica su
bsp. sa
xatilis
§1§
1?
?VU
Galium
austri
acum
11
NT1
11
R
Galium
sudet
icum §
1, CE
R
Galium
tricorn
utum C
E1
CR1
11
11
11
EXVU
1VU
EX1
VU1
CR1
1EN
11
1EN
1CR
Genis
ta sagi
ttalis §
31
11
11
1LC
11
11
11
11
11
1LC
VU
Gentia
na pun
ctata §
11
EN1
11
VU1
1R
NTLC
VU
Gentia
na ver
na §1
11
11
11
11
11
11
11
LC1
LC
Gentia
nella a
marell
a §2, C
E1
CRR
1VU
§1
1VU
11
NTCR
CR1
NT1
1EN
1LC
Gentia
nella c
ampes
tris su
bsp. ba
ltica §
1, CE
1VU
IEN
ENCR
CRR
ENLC
Gentia
nella g
ermani
ca sub
sp. ge
rmanic
a CE
EX1
1CR
11
1NT
LC
Gentia
nella l
utesce
ns sub
sp. ca
rpatica
§2CR
11
NT1
1
Gentia
nella l
utesce
ns sub
sp. lut
escens
§2, C
E1
11
11
11
Gentia
nella o
btusifo
lia sub
sp. stu
rmiana
VU
1NT
Gentia
nella p
raecox
subsp
. bohem
ica §1
, CE
CR1
CR
Geran
ium div
aricatu
m 1
11
11
11
11
1CR
11
VU1
1EN
1
Gladio
lus pa
lustris
§1, C
E1
1VU
DD1
EXEN
EN1
CREN
DDCR
VU?
ENEN
Glauci
um co
rnicula
tum
11
11
11
11
VU1
11
11
1EN
11
EX1
Glaux
maritim
a §1
11
11
11
VUEN
1EN
1LR
1R
1CR
REN
11
1LC
Gnaph
alium s
upinum
1
11
11
11
11
11
11
11
11
LC1
NT
Goody
era rep
ens §1
REN
11
11
11
1LC
CR1
NT1
CR1
RVU
VU1
1LC
VULC
Groenl
andia d
ensa §
1, CE
RCR
VU1
DDVU
1EX
REX
ENLR
EXCR
VUVU
1EN
ENEX
NTVU
Gymn
adenia
densi
flora §
11
11
NTVU
11
EN1
1DD
Gypso
phila p
anicul
ata §2
11
11
11
11
EN1
EN1
NT1
1
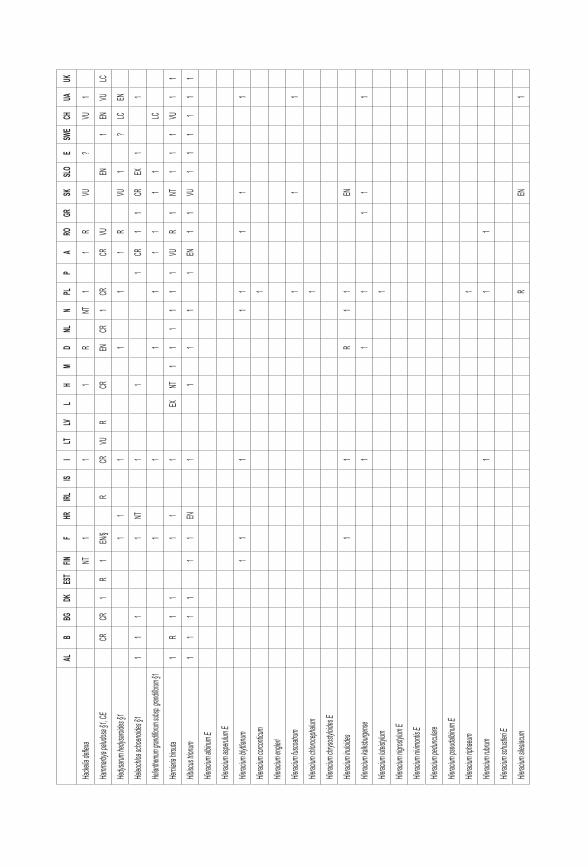
333
Příroda, Praha, 31: xx–xxx, 2011
AlB
BGdK
Est
FinF
HRiRl
isi
ltlv
lH
Md
nln
PlP
ARo
GRsK
slo
EsW
EcH
uAuK
Hacke
lia defl
exa
NT1
11
RNT
11
RVU
?VU
1
Hamm
arbya
paludo
sa §1,
CECR
CR1
R1
EN/§
RCR
VUR
CREN
CR1
CRCR
VUEN
1EN
VULC
Hedys
arum h
edysar
oides
§11
11
11
1R
VU1
?LC
EN
Heleo
chloa
schoen
oides
§11
11
1NT
11
1CR
11
CREX
11
Helian
themu
m gran
diflorum
subsp
. grand
iflorum
§11
11
11
11
1LC
Hernia
ria hirs
uta
1R
11
11
1EX
NT1
11
11
1VU
R1
NT1
11
VU1
1
Hibisc
us trio
num
11
11
11
EN1
11
11
EN1
1VU
11
11
11
Hierac
ium alb
inum E
Hierac
ium as
perulu
m E
Hierac
ium bly
ttianum
1
11
11
11
1
Hierac
ium co
rcontic
um
1
Hierac
ium en
gleri
Hierac
ium fus
coatrum
11
1
Hierac
ium ch
lorocep
halum
1
Hierac
ium ch
rysost
yloide
s E
Hierac
ium inu
loides
11
R1
1EN
Hierac
ium ka
lksbur
gense
11
11
11
Hierac
ium lut
eistylu
m 1
Hierac
ium nig
rostylu
m E
Hierac
ium niv
imontis
E
Hierac
ium pe
duncul
are
Hierac
ium ps
eudalb
inum E
Hierac
ium rip
haeum
1
Hierac
ium rub
rum
11
1
Hierac
ium sc
hustler
i E
Hierac
ium sil
esiacu
m R
EN1
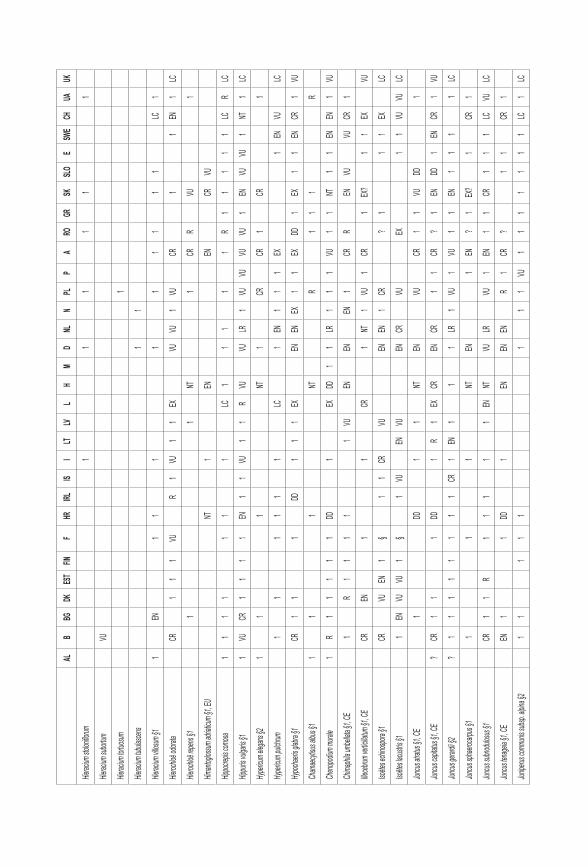
334
Liška: Česká jména lišejníků
AlB
BGdK
Est
FinF
HRiRl
isi
ltlv
lH
Md
nln
PlP
ARo
GRsK
slo
EsW
EcH
uAuK
Hierac
ium sto
loniflo
rum
11
11
11
Hierac
ium su
bortum
VU
Hierac
ium tor
tuosum
1
Hierac
ium tub
ulasce
ns1
1
Hierac
ium vil
losum
§11
EN1
11
11
11
11
LC1
Hieroc
hloë o
dorata
CR
11
1VU
R1
VU1
1EX
VUVU
1VU
CR1
1EN
1LC
Hieroc
hloë re
pens §
11
1NT
1CR
RVU
1
Himant
ogloss
um ad
riaticu
m §1, E
UNT
1EN
ENCR
VU
Hippoc
repis c
omosa
1
11
11
11
LC1
11
11
R1
11
11
LCR
LC
Hippur
is vulg
aris §1
1VU
CR1
11
1EN
11
VU1
1R
VUVU
LR1
VUVU
VUVU
1EN
VUVU
1NT
1LC
Hyper
icum e
legans
§21
11
NT1
CRCR
1CR
1
Hyper
icum p
ulchru
m 1
11
11
1LC
1EN
11
1EX
1EN
VULC
Hypoc
haeris
glabra
§1CR
11
1DD
11
1EX
ENEN
EX1
1EX
DD1
EX1
1EN
CR1
VU
Cham
aecytis
us alb
us §1
11
1NT
R1
11
R
Cheno
podium
murale
1
R1
11
11
DD1
EXDD
11
LR1
11
VU1
1NT
11
ENEN
1VU
Chima
phila u
mbella
ta §1, C
E1
R1
11
11
VUEN
ENEN
1CR
REN
VUVU
CR1
Illeceb
rum ve
rticilla
tum §1
, CE
CREN
11
CR1
NT1
VU1
CR1
EX?
11
EXVU
Isoëte
s echi
nospor
a §1
CRVU
EN1
§1
1CR
VUEN
EN1
CR?
11
1EX
LC
Isoëte
s lacus
tris §1
1EN
VUVU
1§
1VU
ENVU
ENCR
VUEX
11
VUVU
LC
Juncus
atratu
s §1, C
E1
DD1
1NT
ENVU
CR1
1VU
DD1
Juncus
capita
tus §1
, CE
?CR
11
1DD
1R
1EX
CREN
CR1
1CR
?1
ENDD
1EN
CR1
VU
Juncus
gerar
dii §2
?1
11
11
11
1CR
1EN
11
1LR
1VU
1VU
11
EN1
11
1LC
Juncus
sphae
rocarp
us §1
11
1NT
EN1
EN?
1EX
?1
CR1
Juncus
subno
dulosu
s §1
CR1
1R
11
11
1EN
NTVU
LRVU
1EN
11
CR1
11
LCVU
LC
Juncus
tenage
ia §1, C
EEN
11
DD1
ENEN
ENR
1CR
?1
1CR
1
Junipe
rus co
mmuni
s subs
p. alpin
a §2
11
11
11
11
VU1
11
11
11
LC1
LC

335
Příroda, Praha, 31: xx–xxx, 2011
AlB
BGdK
Est
FinF
HRiRl
isi
ltlv
lH
Md
nln
PlP
ARo
GRsK
slo
EsW
EcH
uAuK
Jurine
a cyan
oides
§1, CE
EN1
Knaut
ia arve
nsis s
ubsp. p
seudol
ongifol
ia E, §1
Kochi
a pros
trata §
11
11
1NT
EN1
11
11
Lappul
a sem
icincta
1
11
1
Laser t
rilobum
§11
11
VU/§
1LR
11
VU1
?NT
1
Laserp
itium a
rchang
elica §
1CR
1R
R1
1
Lathyr
us aph
aca
1R
11
11
11
1LC
NT1
1CR
11
11
11
EN1
11
CR1
VU
Lathyr
us het
erophy
llus §2
11
1EN
1VU
1LC
1
Lathyr
us hirs
utus
1R
11
11
1CR
1EN
11
11
EN1
1VU
11
1CR
11
Lathyr
us nis
solia
1R
11
11
1CR
1EN
VU1
REN
11
VU1
11
CR1
LC
Lathyr
us pal
ustris
§11
ENCR
11
11
DDNT
11
11
NT1
LREN
VUEN
1EN
1VU
1NT
Lathyr
us pan
nonicu
s subs
p. pann
onicus
§1DD
ENEN
CR
Lathyr
us pis
iformis
§11
R1
EN1
RCR
VU?
1
Leucan
themu
m adus
tum
11
1DD
11
NT1
1LC
1
Leucoj
um ae
stivum
§1, C
E1
ENEN
11
11
1NT
1VU
1EN
VU1
VUVU
11
VUVU
LC
Ligula
ria sib
irica §
1, CE
CRVU
VU/§
CREN
EXCR
ENR
VU
Limodo
rum ab
ortivum
§11
CRR
11
1EX
NTCR
1EN
R1
ENVU
1NT
EN
Linaria
arvens
is 1
EX1
11
1EX
EXCR
EX1
EX?
1EX
1DD
Linder
nia pro
cumben
s §1, C
E1
CR1
VUVU
1EN
VUR
ENVU
CRVU
1EX
1
Linum
hirsut
um su
bsp. hi
rsutum
§11
11
1R
EN1
1VU
11
Liparis
loesel
ii §1, C
ECR
EXVU
REN
VU/§
DDEN
VUVU
CREN
ENEX
CREN
RCR
ENNT
VUVU
EN
Listera
corda
ta §1
11
1NT
11
11
11
VU1
11
NT1
VU1
REN
VU1
1LC
VULC
Litorell
a unifl
ora §1
, CE
RVU
1§
DD1
1VU
1EN
EN1
VU1
CRVU
1EN
LC
Luroni
um na
tans C
EVU
VU§
DD1
11
VUVU
CREX
ENEN
LC
Luzula
spica
ta §1
11
11
11
11
11
11
1LC
1LC
Malax
is mono
phyllos
§1R
VUDD
VUVU
REX
1CR
VU1
REN
VUVU
VUVU
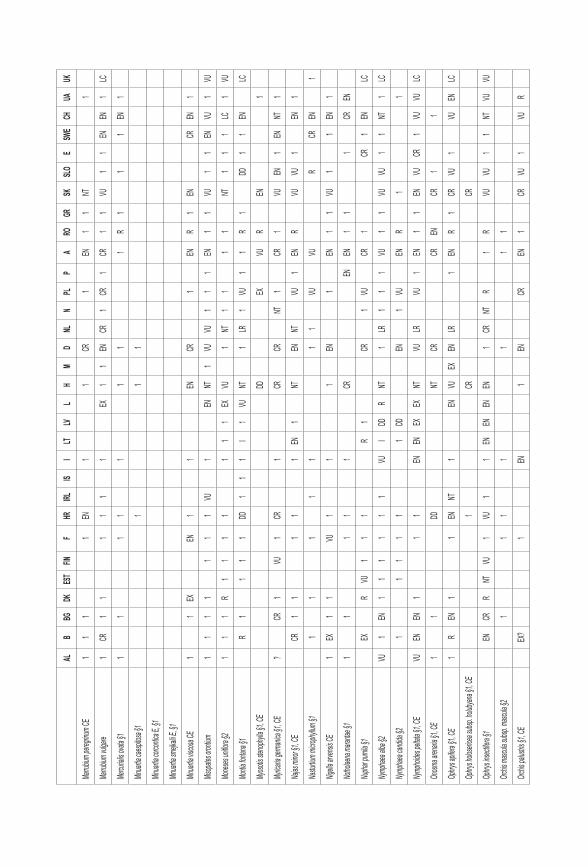
336
Liška: Česká jména lišejníků
AlB
BGdK
Est
FinF
HRiRl
isi
ltlv
lH
Md
nln
PlP
ARo
GRsK
slo
EsW
EcH
uAuK
Marru
bium p
eregrin
um CE
11
11
EN1
1CR
1EN
11
NT1
Marru
bium v
ulgare
1
CR1
11
11
1EX
11
ENCR
1CR
1CR
11
VU1
1EN
EN1
LC
Mercu
rialis o
vata §
11
11
11
11
1R
11
1EN
1
Minuar
tia cae
spitos
a §1
11
1
Minuar
tia cor
contica
E, §1
Minuar
tia sm
ejkalii
E, §1
Minuar
tia vis
cosa C
E1
1EX
EN1
1EN
CR1
ENR
1EN
CREN
1
Misopa
tes oro
ntium
11
11
11
1VU
1EN
NT1
VUVU
11
1EN
11
VU1
1EN
VU1
VU
Mones
es uni
flora §
21
11
R1
11
11
11
EXVU
1NT
11
11
NT1
11
LC1
VU
Montia
fontan
a §1
R1
11
1DD
11
1I
1VU
NT1
LR1
VU1
1R
1DD
11
ENLC
Myoso
tis ste
nophyl
la §1, C
EDD
EXVU
REN
1
Myrica
ria ger
manic
a §1, C
E?
CR1
VU1
CR1
CRCR
NT1
CR1
VUEN
1EN
NT1
Najas
minor
§1, C
ECR
11
11
1EN
1NT
ENNT
VU1
ENR
VUVU
1EN
1
Nastu
rtium m
icrophy
llum §1
11
11
11
1VU
VUR
CREN
1
Nigella
arvens
is CE
1EX
11
VU1
11
EN1
EN1
1VU
11
EN1
Notho
laena
maran
tae §1
11
11
1CR
ENEN
11
1CR
EN
Nupha
r pumil
a §1
EXR
VU1
11
R1
CR1
VUCR
1CR
1EN
LC
Nymp
haea a
lba §2
VU1
EN1
11
11
1VU
IDD
RNT
1LR
11
1VU
11
VUVU
11
NT1
LC
Nymp
haea c
andida
§21
11
11
1DD
EN1
VUEN
R1
1
Nymp
hoides
peltat
a §1, C
EVU
ENEN
11
1EN
ENEX
EXNT
VULR
VU1
EN1
1EN
VUCR
1VU
VULC
Onosm
a aren
aria §1
, CE
11
DDNT
CRCR
ENCR
11
Ophry
s apife
ra §1, C
E1
REN
11
ENNT
1EN
VUEX
ENLR
1EN
R1
CRVU
1VU
ENLC
Ophry
s holo
serice
a subs
p. holu
byana
§1, CE
1CR
CR
Ophry
s insec
tifera §
1EN
CRR
NTVU
1VU
11
ENEN
ENEN
1CR
NTR
1R
VUVU
11
NTVU
VU
Orchis
mascu
la subs
p. masc
ula §2
11
11
11
1
Orchis
palus
tris §1
, CE
EX?
1EN
1EN
CREN
1CR
VU1
VUR
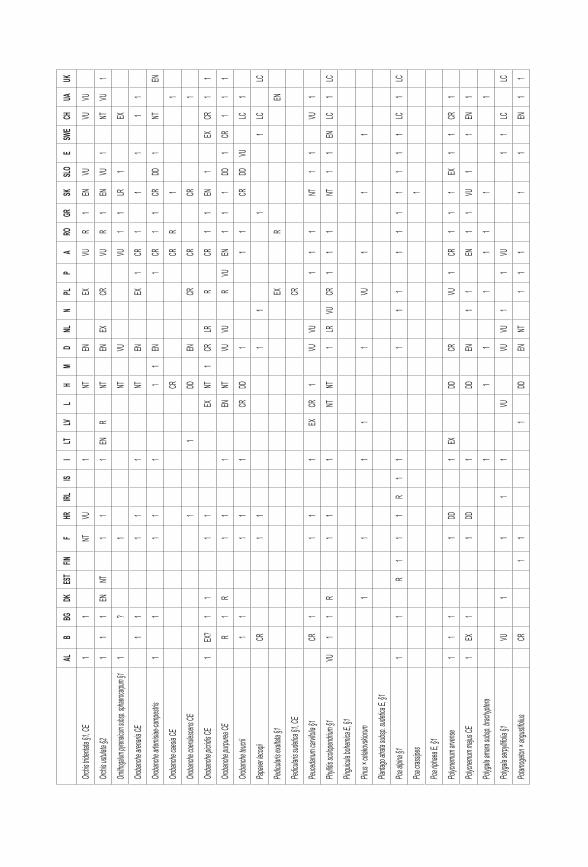
337
Příroda, Praha, 31: xx–xxx, 2011
AlB
BGdK
Est
FinF
HRiRl
isi
ltlv
lH
Md
nln
PlP
ARo
GRsK
slo
EsW
EcH
uAuK
Orchis
trident
ata §1
, CE
11
NTVU
1NT
ENEX
VUR
1EN
VUVU
VU
Orchis
ustula
ta §2
11
1EN
NT1
11
ENR
NTEN
EXCR
VUR
1EN
VU1
NTVU
1
Ornitho
galum p
yrenaic
um sub
sp. sph
aeroca
rpum §
11
?1
NTVU
VU1
1LR
1EX
Oroban
che are
naria C
E1
11
11
NTEN
EX1
CR1
11
11
Oroban
che art
emisia
e-cam
pestris
1
11
11
11
EN1
CR1
1CR
DD1
NTEN
Oroban
che ca
esia C
ECR
CRR
11
Oroban
che co
erules
cens C
E1
1DD
ENCR
CRCR
1
Oroban
che pic
ridis C
E1
EX?
11
11
EXNT
1CR
LRR
CR1
1EN
1EX
CR1
1
Oroban
che pu
rpurea
CER
1R
11
1EN
NTVU
VUR
VUEN
11
1DD
1CR
11
1
Oroban
che teu
crii
11
11
1CR
DD1
11
CRDD
VULC
1
Papav
er leco
qii CR
11
11
11
LCLC
Pedic
ularis
exaltat
a §1
EXR
EN
Pedic
ularis
sudetic
a §1, C
ECR
Peuce
danum
carvif
olia §1
CR1
11
1EX
CR1
VUVU
11
1NT
11
VU1
Phyllit
is scol
opendr
ium §1
VU1
1R
11
1NT
NT1
LRVU
CR1
11
NT1
1EN
LC1
LC
Pingui
cula b
ohemic
a E, §1
Pinus
× cela
kovski
orum
11
11
1VU
11
1
Planta
go atra
ta subs
p. sude
tica E,
§1
Poa a
lpina §
11
1R
11
1R
11
11
11
11
11
11
LC1
LC
Poa c
rassip
es 1
Poa ri
phaea
E, §1
Polyc
nemum
arvens
e 1
11
1DD
1EX
DDCR
VU1
CR1
11
EX1
1CR
1
Polyc
nemum
majus
CE1
EX1
1DD
1DD
EN1
1EN
11
VU1
1EN
1
Polyg
ala am
ara su
bsp. br
achypt
era
11
11
11
11
Polyg
ala se
rpyllifo
lia §1
VU1
11
1VU
VUVU
11
VU1
1LC
LC
Potam
ogeton
× angu
stifoliu
s CR
11
1DD
ENNT
11
11
1EN
11
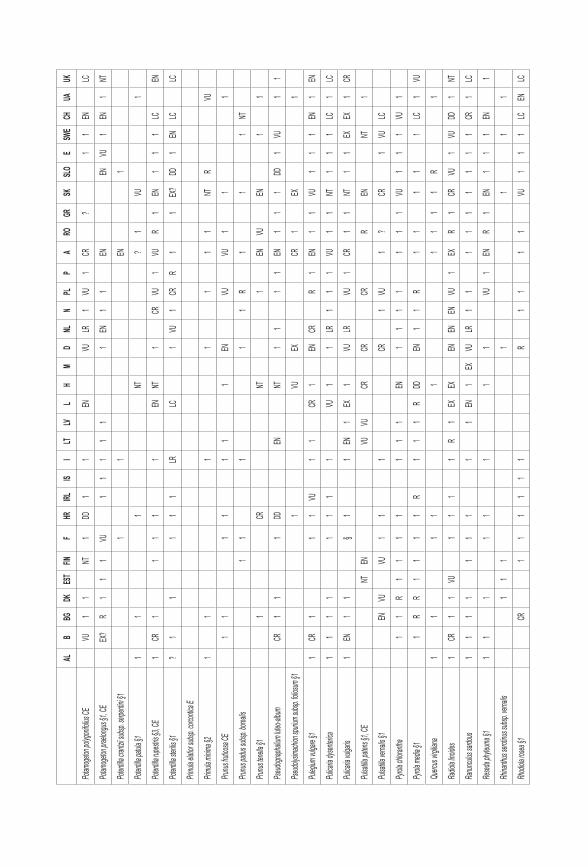
338
Liška: Česká jména lišejníků
AlB
BGdK
Est
FinF
HRiRl
isi
ltlv
lH
Md
nln
PlP
ARo
GRsK
slo
EsW
EcH
uAuK
Potam
ogeton
polyg
onifoli
us CE
VU1
1NT
1DD
11
ENVU
LR1
VU1
CR?
11
ENLC
Potam
ogeton
praelo
ngus §
1, CE
EX?
R1
11
VU1
11
11
1EN
11
ENEN
VU1
EN1
NT
Poten
tilla cra
ntzii s
ubsp. s
erpent
ini §1
11
EN1
Poten
tilla pa
tula §1
11
1NT
?1
VU1
Poten
tilla rup
estris
§3, CE
1CR
11
11
1EN
NT1
CRVU
1VU
R1
EN1
11
LCEN
Poten
tilla ste
rilis §1
?1
11
11
LRLC
1VU
1CR
R1
1EX
?DD
1EN
LCLC
Primu
la elati
or subs
p. corc
ontica
E
Primu
la minim
a §2
11
11
11
1NT
RVU
Prunus
frutico
sa CE
11
11
11
1EN
VUVU
11
1
Prunus
padus
subsp
. borea
lis 1
11
11
R1
11
NT
Prunus
tenella
§11
CRNT
1EN
VUEN
11
Pseudo
gnapha
lium lut
eo-alb
um
CR1
11
DDEN
NT1
11
1EN
11
1DD
1VU
11
Pseudo
lysima
chion
spurium
subsp
. folios
um §1
1VU
EXCR
1EX
1
Puleg
ium vu
lgare §
11
CR1
11
VU1
1CR
1EN
CRR
1EN
11
VU1
11
EN1
EN
Pulica
ria dys
enteric
a 1
11
11
11
1VU
11
LR1
11
VU1
1NT
11
1LC
1LC
Pulica
ria vul
garis
1EN
11
§1
1EN
1EX
1VU
LRVU
1CR
11
NT1
1EX
EX1
CR
Pulsa
tilla pa
tens §
1, CE
NTEN
VUVU
CRCR
CRR
ENNT
1
Pulsa
tilla ve
rnalis
§1EN
VUVU
11
1CR
1VU
1?
CR1
VULC
Pyrola
chlora
ntha
11
R1
11
11
11
EN1
11
11
11
VU1
11
VU1
Pyrola
media
§11
RR
11
11
R1
11
RDD
EN1
1R
11
11
1LC
1VU
Querc
us virg
iliana
11
11
11
11
1R
1
Radio
la lino
ides
1CR
11
VU1
11
1R
1EX
EXEN
ENEN
VU1
EXR
1CR
VU1
VUDD
1NT
Ranun
culus
sardou
s 1
11
11
11
11
EN1
EXVU
LR1
11
11
11
11
CR1
LC
Resed
a phyt
euma §
11
11
11
11
1VU
1EN
R1
EN1
11
EN1
Rhina
nthus
serotin
us sub
sp. ve
rnalis
11
11
11
1
Rhodi
ola ros
ea §1
CR1
11
11
1R
11
11
VU1
11
LCEN
LC
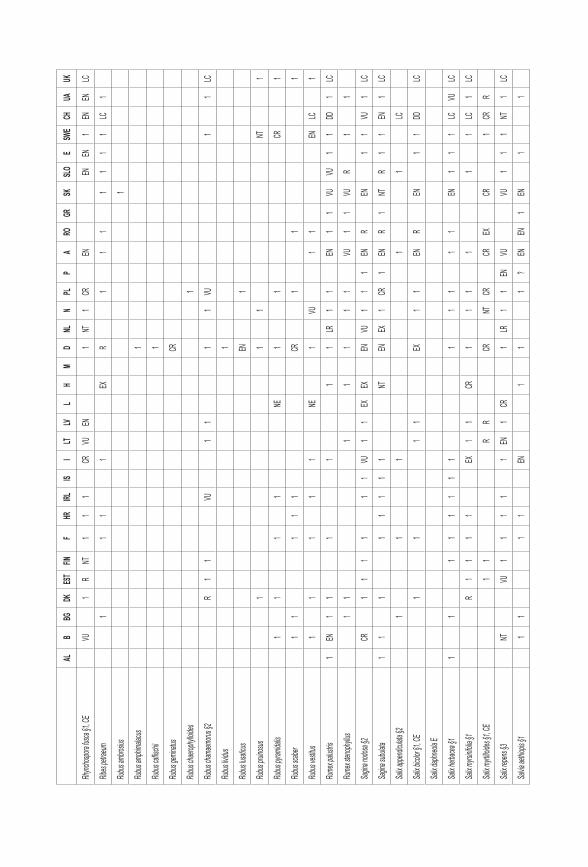
339
Příroda, Praha, 31: xx–xxx, 2011
AlB
BGdK
Est
FinF
HRiRl
isi
ltlv
lH
Md
nln
PlP
ARo
GRsK
slo
EsW
EcH
uAuK
Rhync
hospor
a fusca
§1, C
EVU
1R
NT1
11
CRVU
EN1
NT1
CREN
ENEN
1EN
ENLC
Ribes
petrae
um
11
11
EXR
11
11
11
1LC
1
Rubus
ambro
sius
1
Rubus
amphi
malac
us 1
Rubus
caflis
chii
1
Rubus
gemin
atus
CR
Rubus
chaer
ophyllo
ides
1
Rubus
cham
aemoru
s §2
R1
1VU
11
11
VU1
1LC
Rubus
lividu
s 1
Rubus
lusatic
us EN
1
Rubus
pruino
sus1
11
NT1
Rubus
pyram
idalis
11
11
NE1
1CR
1
Rubus
scabe
r 1
11
11
CR1
11
Rubus
vestit
us 1
11
11
NE1
VU1
1EN
LC1
Rume
x palu
stris
1EN
11
11
11
LR1
1EN
11
VUVU
11
DD1
LC
Rume
x sten
ophyllu
s 1
11
11
11
VU1
1VU
R1
1
Sagin
a nodo
sa §2
CR1
11
11
1VU
11
EXEX
ENVU
11
1EN
REN
11
VU1
LC
Sagin
a subu
lata
11
11
11
11
NTEN
EX1
CR1
ENR
1NT
R1
1EN
1LC
Salix
append
iculata
§21
11
11
LC
Salix
bicolo
r §1, C
E1
11
1EX
11
ENR
EN1
1DD
LC
Salix
daphne
ola E
Salix
herbac
ea §1
11
11
11
11
11
11
1EN
11
1LC
VULC
Salix
myrsin
ifolia §
1R
11
11
EX1
1CR
11
11
11
LC1
LC
Salix
myrtill
oides
§1, CE
11
RR
CRNT
CRCR
EXCR
1CR
R
Salix
repens
§3NT
VU1
11
11
EN1
CR1
LR1
1EN
VUVU
11
1NT
1LC
Salvia
aethio
pis §1
11
11
EN1
11
?EN
EN1
EN1
1
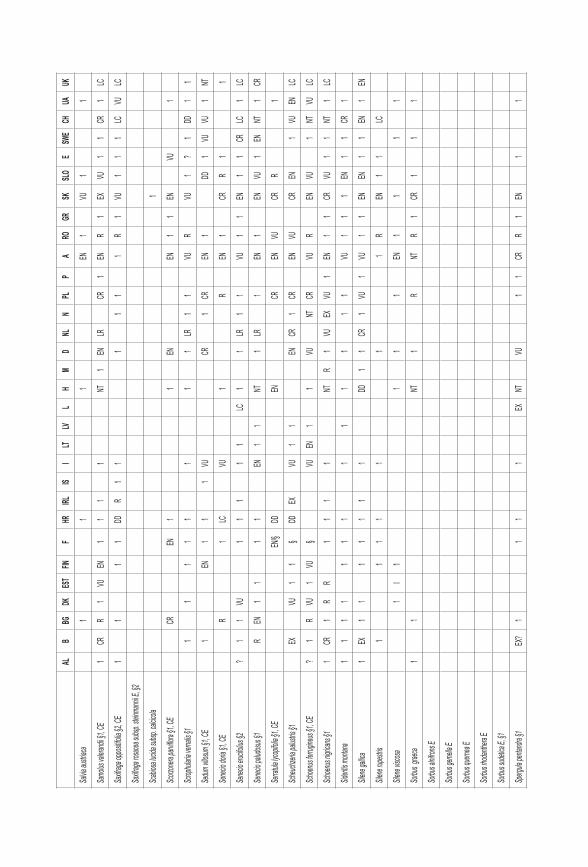
340
Liška: Česká jména lišejníků
AlB
BGdK
Est
FinF
HRiRl
isi
ltlv
lH
Md
nln
PlP
ARo
GRsK
slo
EsW
EcH
uAuK
Salvia
austri
aca
11
1EN
1VU
11
Samo
lus va
lerandi
i §1, C
E1
CRR
1VU
EN1
11
1NT
1EN
LRCR
1EN
R1
EXVU
11
CR1
LC
Saxifr
aga op
positifo
lia §2,
CE1
11
1DD
R1
11
11
1R
1VU
11
1LC
VULC
Saxifr
aga ros
acea s
ubsp. s
teinma
nnii E,
§2
Scabio
sa luc
ida su
bsp. ca
lcicola
1
Scorzo
nera p
arviflo
ra §1, C
ECR
EN1
1EN
EN1
1EN
VU1
Scroph
ularia
vernal
is §1
11
11
11
11
LR1
1VU
RVU
1?
1DD
11
Sedum
villos
um §1
, CE
1EN
11
1VU
CR1
CREN
1DD
1VU
VU1
NT
Senec
io doria
§1, C
ER
1LC
VU1
REN
1CR
R1
1
Senec
io eruc
ifolius
§2?
11
VU1
11
11
LC1
1LR
11
VU1
1EN
11
CRLC
1LC
Senec
io palu
dosus
§1R
EN1
11
1EN
11
NT1
LR1
EN1
ENVU
1EN
NT1
CR
Serra
tula lyc
opifoli
a §1, C
EEN
/§DD
ENCR
ENVU
CRR
1
Scheuc
hzeria
palust
ris §1
EXVU
11
§DD
EXVU
11
ENCR
1CR
ENVU
CREN
1VU
ENLC
Schoen
us ferr
ugineu
s §1, C
E?
1R
VU1
VU§
VUEN
11
VUNT
CRVU
REN
VU1
NTVU
LC
Schoen
us nig
ricans
§11
CR1
RR
11
11
NTR
1VU
EXVU
1EN
11
CRVU
11
NT1
LC
Siderit
is mont
ana
11
11
11
11
11
11
1VU
11
1EN
11
CR1
Silene
gallica
1
EX1
11
11
11
DD1
1CR
1VU
1VU
11
ENEN
11
EN1
EN
Silene
rupest
ris 1
11
11
11
REN
11
LC
Silene
viscos
a 1
I1
11
1EN
11
11
Sorbu
s grae
ca 1
1NT
1R
NTR
1CR
11
1
Sorbu
s alnif
rons E
Sorbu
s gem
ella E
Sorbu
s quer
nea E
Sorbu
s rhoda
nthera
E
Sorbu
s sude
tica E,
§1
Sperg
ula pe
ntandr
a §1
EX?
11
11
EXNT
VU1
1CR
R1
EN1
1
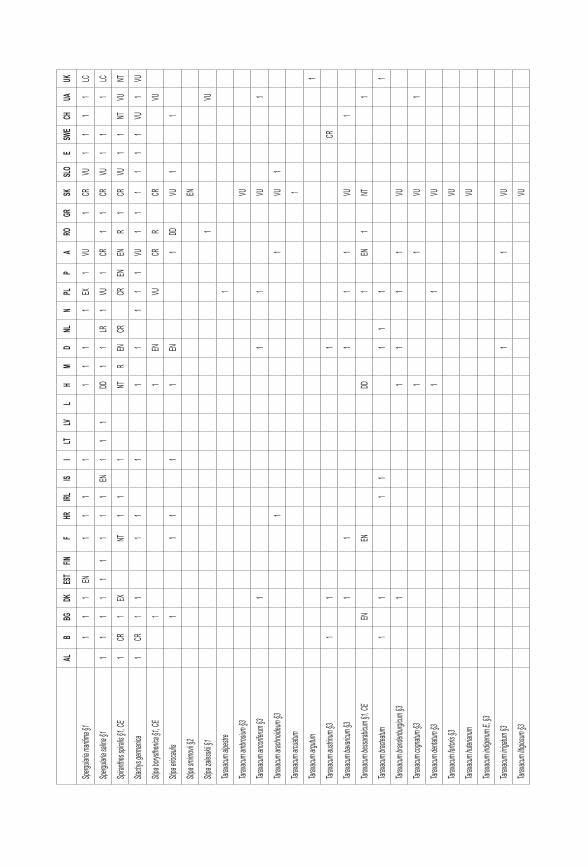
341
Příroda, Praha, 31: xx–xxx, 2011
AlB
BGdK
Est
FinF
HRiRl
isi
ltlv
lH
Md
nln
PlP
ARo
GRsK
slo
EsW
EcH
uAuK
Sperg
ularia
maritim
a §1
11
1EN
11
11
11
11
EX1
VU1
CRVU
11
11
LC
Sperg
ularia
salina
§11
11
11
11
11
EN1
11
DD1
1LR
1VU
1CR
11
CRVU
11
1LC
Spiran
thes s
piralis
§1, CE
1CR
1EX
NT1
11
NTR
ENCR
CREN
ENR
1CR
VU1
1NT
VUNT
Stachy
s germ
anica
1CR
11
11
11
11
11
VU1
11
11
1VU
1VU
Stipa b
orysth
enica
§1, CE
11
ENVU
CRR
CRVU
Stipa e
riocaul
is 1
11
11
EN1
DDVU
11
Stipa s
mirnov
ii §2
EN
Stipa z
alessk
ii §1
1VU
Taraxa
cum alp
estre
1
Taraxa
cum am
brosiu
m §3
VU
Taraxa
cum an
corifer
um §3
11
1VU
1
Taraxa
cum ara
chnoid
eum §3
11
VU1
Taraxa
cum arc
uatum
1
Taraxa
cum arg
utum
1
Taraxa
cum au
strinum
§31
11
CR
Taraxa
cum ba
varicu
m §3
11
11
1VU
1
Taraxa
cum be
ssarab
icum §
1, CE
ENEN
DD1
EN1
NT1
Taraxa
cum bra
cteatu
m 1
11
11
11
1
Taraxa
cum bra
ndenbu
rgicum
§31
11
11
VU
Taraxa
cum co
gnatum
§31
1VU
1
Taraxa
cum de
ntatum
§31
1VU
Taraxa
cum far
toris §
3VU
Taraxa
cum hu
terianu
m VU
Taraxa
cum ind
igenum
E, §3
Taraxa
cum irri
gatum
§31
1VU
Taraxa
cum liti
giosum
§3VU
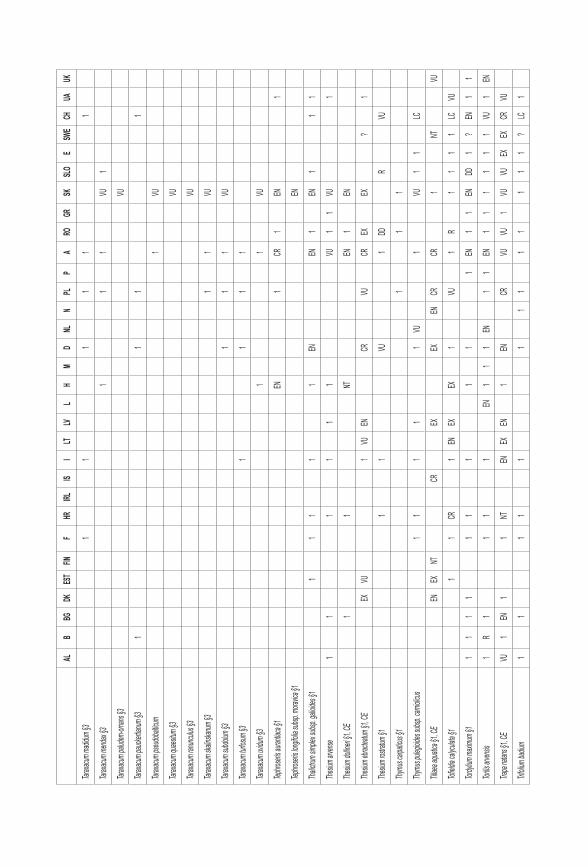
342
Liška: Česká jména lišejníků
AlB
BGdK
Est
FinF
HRiRl
isi
ltlv
lH
Md
nln
PlP
ARo
GRsK
slo
EsW
EcH
uAuK
Taraxa
cum ma
didum
§31
11
11
1
Taraxa
cum me
ndax §
31
11
VU1
Taraxa
cum pa
ludem
-ornan
s §3
VU
Taraxa
cum pa
uckerti
anum §
31
11
1
Taraxa
cum ps
eudoba
lticum
1
VU
Taraxa
cum qu
aesitum
§3VU
Taraxa
cum ran
unculu
s §3
VU
Taraxa
cum sk
alinska
num §3
11
VU
Taraxa
cum su
bdolum
§31
11
VU
Taraxa
cum tur
fosum
§31
11
1
Taraxa
cum uv
idum §
31
1VU
Tephro
seris a
urantia
ca §1
EN1
CR1
EN1
Tephro
seris l
ongifol
ia subs
p. mora
vica §
1EN
Thalict
rum sim
plex s
ubsp. g
alioide
s §1
11
11
1EN
EN1
EN1
11
Thesiu
m arve
nse
11
11
11
VU1
1VU
1
Thesiu
m dolli
neri §1
, CE
11
NTEN
1EN
Thesiu
m ebra
cteatu
m §1, C
EEX
VU1
VUEN
CRVU
CREX
EX?
1
Thesiu
m rost
ratum §
11
1VU
1DD
RVU
Thymu
s carp
aticus
§11
11
Thymu
s pule
gioide
s subs
p. carn
iolicus
1
11
11
VU1
VU1
1LC
Tillaea
aquat
ica §1
, CE
ENEX
NTCR
EXEX
ENCR
CR1
NTVU
Tofield
ia caly
culata
§11
1CR
1EN
EXEX
1VU
1R
11
11
LCVU
Tordyl
ium ma
ximum
§11
11
11
11
11
1EN
11
ENDD
1?
EN1
1
Torilis
arvens
is 1
R1
11
1EN
11
1EN
11
EN1
11
11
1VU
1EN
Trapa
natans
§1, C
EVU
1EN
11
NTEN
EXEN
1EN
CRVU
VU1
VUVU
EXEX
CRVU
Trifoliu
m badi
um
11
11
11
11
11
11
1?
LC1
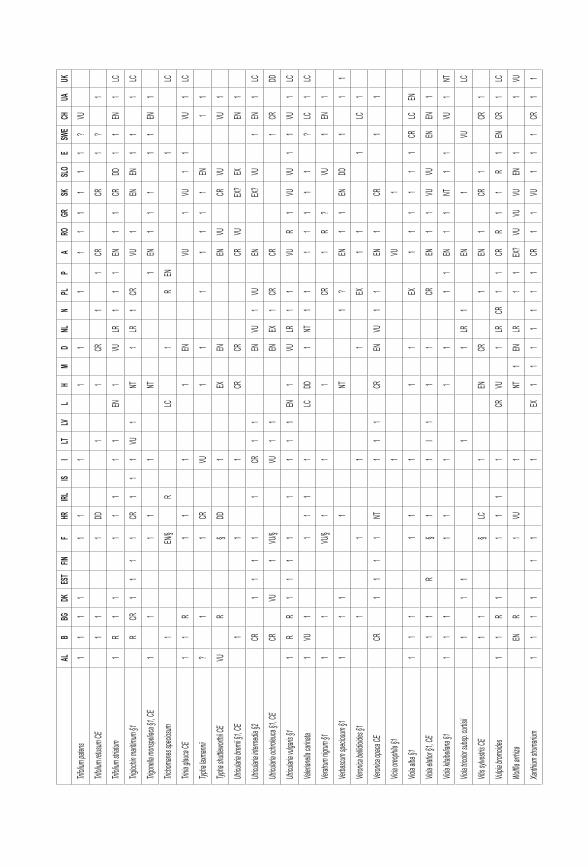
343
Příroda, Praha, 31: xx–xxx, 2011
AlB
BGdK
Est
FinF
HRiRl
isi
ltlv
lH
Md
nln
PlP
ARo
GRsK
slo
EsW
EcH
uAuK
Trifoliu
m pate
ns 1
11
11
11
11
11
11
11
1?
VU
Trifoliu
m retu
sum CE
11
1DD
11
CR1
1CR
CR1
?1
Trifoliu
m stria
tum
1R
11
11
11
1EN
1VU
LR1
11
EN1
1CR
DD1
1EN
1LC
Trigloc
hin ma
ritimum
§1R
CR1
11
1CR
11
1VU
1NT
1LR
1CR
VU1
ENEN
11
1LC
Trigone
lla mo
nspelia
ca §1,
CE1
11
11
NT1
EN1
11
11
EN1
Trichom
anes s
pecios
um
1EN
/§R
LC1
REN
1LC
Trinia g
lauca
CE1
1R
11
11
ENVU
1VU
11
VU1
LC
Typha
laxma
nnii
?1
1CR
VU1
11
11
11
EN1
1
Typha
shuttle
worthi
i CE
VUR
§DD
1EX
ENEN
VUCR
VUVU
1
Utricu
laria b
remii §
1, CE
11
1CR
CRCR
VUEX
?EX
EN1
Utricu
laria in
termedi
a §2
CR1
11
11
CR1
1EN
VU1
VUEN
EX?
VU1
EN1
LC
Utricu
laria o
chrole
uca §1
, CE
CRVU
1VU
/§VU
11
ENEX
1CR
CR1
CRDD
Utricu
laria v
ulgaris
§11
RR
11
11
11
11
EN1
VULR
11
VUR
1VU
VU1
1VU
1LC
Valeria
nella c
arinata
1
VU1
11
11
LCDD
1NT
11
11
11
1?
LC1
LC
Veratru
m nigru
m §1
11
VU/§
11
1CR
1R
?VU
1EN
1
Verbas
cum sp
eciosu
m §1
11
11
NT1
?EN
11
ENDD
11
1
Veroni
ca bel
lidioid
es §1
11
11
EX1
11
LC1
Veroni
ca opa
ca CE
CR1
11
1NT
11
1CR
ENVU
11
EN1
CR1
1
Vicia o
reophi
la §1
1VU
1
Viola a
lba §1
11
11
11
11
EX1
11
11
1CR
LCEN
Viola e
latior §
1, CE
11
R§
11
I1
11
CREN
11
VUVU
ENEN
1
Viola k
itaibel
iana §
11
11
11
11
11
1EN
11
NT1
1VU
1NT
Viola t
ricolor
subsp.
curtis
ii 1
11
11
LR1
EN1
VULC
Vitis s
ylvest
ris CE
11
§LC
1EN
CR1
EN1
CR1
CR1
Vulpia
bromo
ides
11
R1
11
11
CRVU
1LR
CR1
1CR
R1
1R
1EN
CR1
LC
Wolffia
arrhiz
a EN
R1
VU1
NT1
ENLR
11
EX?
VUVU
VUEN
11
VU
Xanth
ium str
umariu
m1
11
11
11
EX1
11
11
11
CR1
1VU
11
1CR
11

344
Liška: Česká jména lišejníků

345
Příroda, Praha, 31: 345–370, 2011
Jak souvisí změny v rozšíření kriticky ohrožených druhů rostlin České republiky s jejich biologickými vlastnostmi a ekologickými nároky?
How are changes in distribution of critically endangered plant species of the czech Republic connected with their biological and ecological traits?
JarMiLa gabrieLová1, 2, Lucie Černá2 & ZuZana MünZbergová1, 3
1 Katedra botaniky, Přírodovědecká fakulta Univerzity Karlovy, Benátská 2, CZ-128 01 Praha 2; [email protected]; [email protected] Agentura ochrany přírody a krajiny České republiky, Nuselská 34, CZ-140 00 Praha 4; [email protected] Botanický ústav Akademie věd České republiky, v. v. i., Zámek 1, CZ-252 43 Průhonice
Abstract: The number of species threatened with extinction due to human activities is increasing and it is hardly possible to reverse this trend and prevent further reduction of abundance of all endangered species. It is thus necessary to set priorities, to define groups of species with similar traits and possible causes of endangerment and to specify common conservation measures for these groups of species. We examine the relationship between the rate of decline of critically endangered vascular plant species of the Czech Republic (C1 species) and their biological and ecological traits. We also assess priorities for the conservation of these species. We work with the information about the historical and current number of localities (number of grid cells on distribution maps). On the basis of the information, we define the rate of species decline and the rate of change in occurrence within the map grid cells (this parameter differentiates between species steadily occurring within the same map grid cells and species that previously occurred in other map grid cells than at present). Within the analyzed set of 185 C1 species we identify several subgroups based on their rate of change and rate of decline, two of which are well characterized by their biological and ecological traits. The first is a group of massively declining species, which change their occurrence within the map grid cells during the time. This group comprises mainly annual species occurring on habitats strongly influenced or created by man, species with high requirements for nutrients and species whose seeds are dispersed by man. The second group comprises non-declining, stable species – species of alpine treeless habitats, forest species, phanerophytes and anemochorous species. The massively declining species, which tend to occur in different map grid cells during the time, are more endangered than non-declining stable species. However, there are currently no appropriate mechanisms for their conservation.Keywords: grid mapping, life-history traits, rarity, rate of decline, Red List
Abstrakt: Počet druhů ohrožených vyhynutím v důsledku činnosti člověka stále narůstá a není v silách ochrany přírody tento trend zvrátit a zabránit dalšímu snižování početnosti všech ohrožených druhů. Je potřeba stanovit priority, vymezit v rámci rozsáhlého souboru ohrožených druhů skupiny s podobnými vlastnostmi a možnými příčinami ohrožení a pro tyto skupiny pak najít společná ochranářská opatření. V našem příspěvku zkoumáme, jak souvisí míra ústupu kriticky ohrožených druhů rostlin České republiky (C1 druhy) s jejich biologickými

346
Gabrielová et al.: Jak souvisí změny v rozšíření C1 rostlin ČR s jejich vlastnostmi?
a ekologickými vlastnostmi a rovněž zhodnocujeme priority pro ochranu těchto druhů. Vycházíme z informací o historickém a současném počtu lokalit (polí síťového mapování), na základě kterých definujeme míru úbytku druhu a míru obměny osídlení mapových polí (tento parametr odlišuje druhy stabilně se vyskytující v rámci stejných mapových polí a druhy, které se v minulosti vyskytovaly v jiných mapových polích než v současnosti). V rámci souboru 185 analyzovaných C1 druhů vymezujeme několik podskupin, z nichž dvě jsou dobře charakterizované biologickými vlastnostmi a ekologickými nároky. Jedná se o skupinu druhů masivně ubývajících, obměňujících lokality svého výskytu, která je tvořena zejména jednoletými druhy vázanými na biotopy ovlivněnými nebo vytvořenými člověkem, druhy s vysokými nároky na živiny a druhy, jejichž semena jsou šířena člověkem. Druhou skupinu představují neubývající stabilní druhy, které vytrvávají na lokalitách svého výskytu, tedy zejména druhy alpínského bezlesí a lesů, fanerofyty a anemochorní druhy. Druhy masivně ubývající, obměňující lokality svého výskytu, jsou více ohrožené než druhy se stabilním počtem stále stejných lokalit, v současné době však neexistují vhodné mechanismy jejich ochrany.Klíčová slova: červený seznam, druhové charakteristiky, míra úbytku, síťové mapování, vzácnost
Úvod
V průběhu minulých dvou století se tvář naší krajiny změnila k nepoznání. Tradiční hospodaření udržující mozaiku nejrůznějších maloplošných biotopů ustoupilo intenzivnímu obhospodařování rozsáhlých polních kultur a lesních plantáží (konviČka et al. 2005). Změna způsobu hospodaření, přímé ničení nebo degradace a fragmentace stanovišť vedly k ústupu řady rostlinných a živočišných druhů, které byly dříve hojné. Přesto nemůžeme mluvit o všeobecném úbytku biodiverzity, protože ta se naopak spíše zvyšuje – nástupem nových, někdy i invazních druhů, které se u nás dříve nevyskytovaly. Tyto nepůvodní druhy pak často dále vytlačují „starousedlíky“, jejichž situace se stává kritickou. Jednou z priorit ochrany přírody je zabránit dalšímu snižování početnosti ohrožených druhů či dokonce jejich úplnému vymizení.
Za v různé míře ohrožené je v České republice (ČR) považováno 1623 druhů a poddruhů cévnatých rostlin, což představuje více než 60 % druhového bohatství české flóry (Procházka 2001). V kategorii s nejvyšším stupněm ohrožení „kriticky ohrožené“ (C1) najdeme více než 470 druhů, kterým by ze strany ochrany přírody měla být věnována mimořádná pozornost, protože jim často hrozí akutní riziko vyhynutí. Kromě běžných opatření jako je legislativní ochrana, vymezování chráněných území a zabezpečování potřebného managementu, může být v případě některých zvláště ohrožených druhů přistoupeno k vypracování záchranného programu. Tyto programy jsou ovšem poměrně nákladné a potenciálních objektů pro jejich realizaci je příliš mnoho. Je potřeba stanovit priority, což je možné jen na základě znalosti biologie a ekologie druhu, stanovištních nároků, areálu rozšíření, ohrožení v okolních zemích a aktuálního a historického rozšíření v rámci ČR (Master 1991, keller & bollMann 2004, sutherlanD et al. 2004, Pärtel et al. 2005).
Znalost ohrožení druhu v okolních evropských zemích, informace o areálu rozšíření a pozice ČR v rámci tohoto areálu napomáhají stanovení priorit tím, že hodnotí „odpovědnost“ České republiky za ochranu místních populací druhu. Z hlediska udržení celkové biodiverzity by měla mít přednost ochrana druhů endemických pro daný stát či větší území, jehož je stát součástí, ochrana druhů ohrožených v rámci celého svého areálu a případně ochrana

347
Příroda, Praha, 31: 345–370, 2011
reliktních populací a izolovaných arel druhu, protože izolace představuje jeden z důležitých speciačních mechanismů a izolované populace se mohou vyvinout v nový druh (schnittler & Günther 1999). Naopak druhům, pro které ČR představuje hranici areálu rozšíření a je těžké je chránit, protože jejich populace často kolísají vlivem náhodných výkyvů klimatu, může být věnována menší pozornost, pokud jsou v ostatních částech areálu ještě hojné.
Na základě informací o aktuálním a historickém rozšíření druhu na našem území můžeme rozlišit druhy přirozeně vzácné, osidlující specifické habitaty a dlouhodobě přežívající i při malé početnosti populací, od druhů skutečně ohrožených, které se dříve vyskytovaly mnohem hojněji a v posledních desetiletích významně ustoupily. Získat spolehlivé údaje o tom, jak byl druh rozšířen v minulosti, není zcela triviální, protože o jeho výskytu můžeme usuzovat jen nepřímo – především na základě herbářových materiálů a literárních zdrojů. Na základě těchto pramenů dostaneme spíš než obraz skutečného historického rozšíření obrázek o tom, jak aktivně se botanizovalo v různých časových periodách, která místa botanici s oblibou navštěvovali a které druhy tam rádi sbírali. Navíc se tímto způsobem nedá příliš přesně zmapovat rozšíření druhů, které byly v minulosti všeobecně hojné, protože jim ze strany botaniků nebyla věnována větší pozornost, případně je popis lokalit jejich výskytu příliš široký, např. DoMin (1942) „Agrostemma githago L. V obilí v celém kraji, c. 200–380, obecná; je hojnější než chrpa a stračka.“. Naopak druhy, které nikdy nebyly příliš hojné, byly často sbírány a zaznamenávány nepoměrně častěji, protože představovaly botanicky zajímavý „úlovek“. Žádná lepší metoda však zatím neexistuje a díky přesně lokalizovaným a datovaným herbářovým položkám a spolehlivým literárním údajům si můžeme udělat alespoň částečnou představu o tom, jak byl daný druh v různých předchozích časových periodách hojný (shaffer et al. 1998, rich & karran 2006).
Znalost biologických vlastností a ekologických nároků druhů je důležitá, protože může úzce souviset s jejich vzácností a ohrožením. Existuje řada studií porovnávajících vzácné a běžné druhy ve snaze najít charakteristiky, které vzácné druhy odlišují a mohou tak být příčinou jejich vzácnosti (pro přehled viz kunin & Gaston 1997, bevill & louDa 1999, Murray et al. 2002, Gabrielová et al. 2011). Ač se většina těchto prací snaží o zobecnění, přeci jen jsou v mnoha aspektech nesrovnatelné a zřejmě proto bylo zatím objeveno jen málo konzistentních rozdílů v charakteristikách vzácných a běžných druhů. I přesto však druhové vlastnosti zůstávají důležitým vysvětlujícím faktorem při zkoumání míry úbytku a ohrožení druhů.
walker & Preston (2006) studovali vlastnosti vyhynulých rostlin dvou anglických hrabství a zjistili, že druhy kompetičně slabé (malé, stres-tolerantní druhy), rostoucí na otevřených stanovištích s extrémními hodnotami pro půdní reakci a vlhkost, mají mnohem větší pravděpodobnost vyhynutí než by odpovídalo náhodě. Na druhou stranu se jim nepodařilo odhalit jasnou souvislost mezi rizikem vyhynutí a schopností disperze nebo způsobem reprodukce. farnsworth & oGurcak (2008) se zaměřili na oblast zhruba dvojnásobku České republiky na severovýchodě Spojených států a zkoumali, jak míra úbytku rostlinných druhů souvisí s jejich životními charakteristikami. Zjistili, že signifikantně více než ostatní skupiny druhů definovaných v jejich studii ubývají druhy entomogamní, druhy, jejichž semena se šíří na krátkou vzdálenost, druhy nížin a druhy, které v dané oblasti dosahují severního okraje areálu rozšíření. Podobnou problematikou, ale na jiné skupině organismů, se zabývali Dennis et al. (2004), kteří studovali, jak souvisí životní historie, morfologické a fyziologické vlastnosti motýlů s ekologickou strategií jejich hostitelských rostlin. Ukázalo se, že motýli s uzavřenými populacemi, které jsou součástí metapopulací, druhy s malou mobilitou, s kratším obdobím letové aktivity, pomalým vývojem a specializací na hostitelskou rostlinu nebo biotop, jsou spojeni především s hostitelskými rostlinami typu s-stratégů a jsou více ohrožené současnými změnami prostředí. Pavlíková & konviČka (in press), kteří podrobně

348
Gabrielová et al.: Jak souvisí změny v rozšíření C1 rostlin ČR s jejich vlastnostmi?
analyzovali charakteristiky životního cyklu velkých nočních motýlů, dospěli k závěru, že druhy trávníků a světlých lesů jsou více ohrožené než druhy uzavřených lesů.
Stále tedy přetrvává snaha nalézt v rámci rozsáhlé skupiny vzácných a ohrožených druhů nějaké společné jmenovatele, vymezit podskupiny s podobnými vlastnostmi a možnými příčinami ohrožení a pro tyto podskupiny pak najít případná společná ochranářská opatření. V našem příspěvku zkoumáme souvislost míry ústupu kriticky ohrožených druhů cévnatých rostlin a jejich biologických a ekologických vlastností.*)
Metodika
vztah druhových vlastností a změny počtu osídlených mapových polí a jejich obměny
Do analýz jsme zahrnuli 190 C1 druhů, pro které máme údaje pro všechny potřebné vlastnosti. Pracovali jsme s následujícími druhovými charakteristikami: životní forma, délka života, průměrná výška, období kvetení, způsob opylení, způsob šíření semen, Ellenbergovy indikační hodnoty, typ stanoviště, výškový stupeň a informace o ohrožení v ostatních evropských zemích (seznam zkratek v Příloze 1, popis jednotlivých vlastností viz Gabrielová et al. 2011, Příloha 2). Dále jsme měli k dispozici údaje o počtu polí síťového mapování, ve kterých se druh vyskytuje/vyskytoval během tří časových intervalů: do roku 1949, v letech 1950–1999 a po roce 2000 (počet mapových polí v tomto intervalu považujeme za aktuální výskyt). Do analýz vstupovaly následující hodnoty: průměr z celkového počtu mapových polí v prvním (do roku 1949) a druhém (1950–1999) časovém intervalu, celkový počet mapových polí v současnosti (třetí časový interval – po roku 2000), počet kontinuálně stabilních mapových polí – na těchto mapových polích se druh vyskytoval ve všech časových intervalech.
Díky těmto hodnotám jsme získali dvě veličiny (i) proměnnou charakterizující změnu počtu polí síťového mapování v současnosti oproti předchozím dvěma časovým intervalům (U) a (ii) proměnnou charakterizující obměnu osídlení mapových polí druhem (O). Parametr U je definován jako: U = 1-[III/průměr (I+II)], přičemž I = celkový počet polí síťového mapování v prvním časovém intervalu, II = celkový počet polí síťového mapování ve druhém časovém intervalu, III = celkový počet polí síťového mapování ve třetím časovém intervalu. Dle hodnoty tohoto parametru byly druhy rozděleny do tří kategorií:
hodnota U kategorie popis
0,8–1 mas. ubyldruh masivně ubývající – počet mapových polí ve třetím oproti průměrnému počtu v prvním a druhém časovém intervalu se snížil o více než 80 %
0,2–0,8 část. ubyldruh částečně ubývající – počet mapových polí ve třetím oproti průměrnému počtu v prvním a druhém časovém intervalu se snížil o 20–80 %
-0,2–0,2 neubyldruh neubývající – počet mapových polí ve třetím časovém intervalu oproti průměrnému počtu v prvním a druhém časovém intervalu přibližně stejný (± 20 %)
*) Pozn.: V řadě případů se jedná i o taxonomicky nižší jednotky (subspecie), takže by bylo správné používat termín C1 taxony, pro lepší čitelnost však většinou používáme termín C1 druhy.
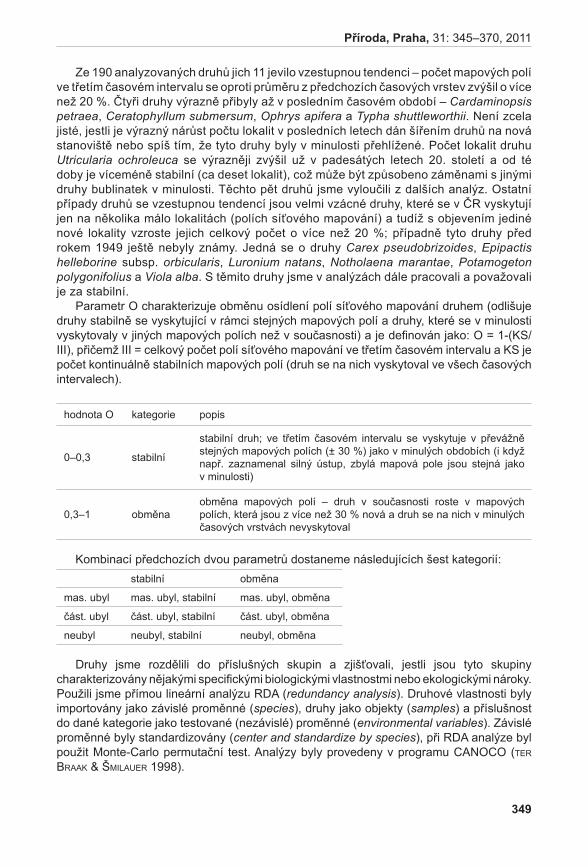
349
Příroda, Praha, 31: 345–370, 2011
Ze 190 analyzovaných druhů jich 11 jevilo vzestupnou tendenci – počet mapových polí ve třetím časovém intervalu se oproti průměru z předchozích časových vrstev zvýšil o více než 20 %. Čtyři druhy výrazně přibyly až v posledním časovém období – Cardaminopsis petraea, Ceratophyllum submersum, Ophrys apifera a Typha shuttleworthii. Není zcela jisté, jestli je výrazný nárůst počtu lokalit v posledních letech dán šířením druhů na nová stanoviště nebo spíš tím, že tyto druhy byly v minulosti přehlížené. Počet lokalit druhu Utricularia ochroleuca se výrazněji zvýšil už v padesátých letech 20. století a od té doby je víceméně stabilní (ca deset lokalit), což může být způsobeno záměnami s jinými druhy bublinatek v minulosti. Těchto pět druhů jsme vyloučili z dalších analýz. Ostatní případy druhů se vzestupnou tendencí jsou velmi vzácné druhy, které se v ČR vyskytují jen na několika málo lokalitách (polích síťového mapování) a tudíž s objevením jediné nové lokality vzroste jejich celkový počet o více než 20 %; případně tyto druhy před rokem 1949 ještě nebyly známy. Jedná se o druhy Carex pseudobrizoides, Epipactis helleborine subsp. orbicularis, Luronium natans, Notholaena marantae, Potamogeton polygonifolius a Viola alba. S těmito druhy jsme v analýzách dále pracovali a považovali je za stabilní.
Parametr O charakterizuje obměnu osídlení polí síťového mapování druhem (odlišuje druhy stabilně se vyskytující v rámci stejných mapových polí a druhy, které se v minulosti vyskytovaly v jiných mapových polích než v současnosti) a je definován jako: O = 1-(KS/III), přičemž III = celkový počet polí síťového mapování ve třetím časovém intervalu a KS je počet kontinuálně stabilních mapových polí (druh se na nich vyskytoval ve všech časových intervalech).
hodnota O kategorie popis
0–0,3 stabilní
stabilní druh; ve třetím časovém intervalu se vyskytuje v převážně stejných mapových polích (± 30 %) jako v minulých obdobích (i když např. zaznamenal silný ústup, zbylá mapová pole jsou stejná jako v minulosti)
0,3–1 obměnaobměna mapových polí – druh v současnosti roste v mapových polích, která jsou z více než 30 % nová a druh se na nich v minulých časových vrstvách nevyskytoval
Kombinací předchozích dvou parametrů dostaneme následujících šest kategorií:stabilní obměna
mas. ubyl mas. ubyl, stabilní mas. ubyl, obměna
část. ubyl část. ubyl, stabilní část. ubyl, obměna
neubyl neubyl, stabilní neubyl, obměna
Druhy jsme rozdělili do příslušných skupin a zjišťovali, jestli jsou tyto skupiny charakterizovány nějakými specifickými biologickými vlastnostmi nebo ekologickými nároky. Použili jsme přímou lineární analýzu RDA (redundancy analysis). Druhové vlastnosti byly importovány jako závislé proměnné (species), druhy jako objekty (samples) a příslušnost do dané kategorie jako testované (nezávislé) proměnné (environmental variables). Závislé proměnné byly standardizovány (center and standardize by species), při RDA analýze byl použit Monte-Carlo permutační test. Analýzy byly provedeny v programu CANOCO (ter braak & ŠMilauer 1998).

350
Gabrielová et al.: Jak souvisí změny v rozšíření C1 rostlin ČR s jejich vlastnostmi?
Dále jsme testovali vztah druhových vlastností a parametrů U (resp. III/průměr (I+II)) a O (resp. III/KS) pomocí zobecněných lineárních modelů (GLM) v programu S-PLUS 2000 Professional (S-PLUS 2000). U parametrů U (resp. III/průměr (I+II)) a O (resp. III/KS) předpokládáme gamma rozdělení. Prováděli jsme oddělené testy pro každou vlastnost zvlášť. V případě kategoriálních charakteristik, kódovaných procentuálně (jeden druh se může vyskytovat ve více kategoriích – např. výškový stupeň, způsob opylení), jsme testovali vliv všech hladin dané charakteristiky dohromady. Pro každou vlastnost jsme stanovili p-hodnotu a procento vysvětlené variability.
Zhodnocení priorit pro ochranu c1 druhů na základě údajů o historickém a současném rozšíření v ČR a ohrožení v ostatních evropských zemích
Na základě údajů o změnách počtu lokalit oproti předchozím dvěma časovým intervalům (parametr U), obměny osídlení mapových polí druhem (parametr O) a situaci ohrožení v ostatních evropských zemích (parametr E) můžeme předběžně zhodnotit míru ohrožení druhů a vybrat druhy, které jsou z těchto hledisek nejohroženější.
Relativní ohrožení v ostatních evropských zemích (parametr E) byl definován jako poměr plochy zemí, ve kterých je druh ohrožen, k celkové ploše zemí, ve kterých se druh vyskytuje. Na základě tohoto parametru jsme druhy rozdělili do následujících kategorií:
hodnota E kategorie popis
0–0,5 Evr. neohrožený druh ohrožený na méně než 50 % plochy zemí, ve kterých se vyskytuje
0,5–1 Evr. ohrožený druh ohrožený na více než 50 % plochy zemí, ve kterých se vyskytuje
Do hodnocení jsme zahrnuli 285 druhů, pro které máme potřebné informace. Z tohoto souboru jsme vyloučili endemity a subendemity (14 druhů), které představují zvláštní skupinu druhů, za jejichž ochranu má ČR mezinárodní odpovědnost a automaticky by tak měly spadat mezi prioritní taxony. Ve všech případech se jedná o druhy s velmi malým počtem lokalit (většinou 1–2 mapová pole), v rámci kterých je ale jejich výskyt víceméně stabilizovaný. U pěti druhů je mezinárodní odpovědnost deklarována příslušnými dokumenty (druhy přílohy č. IV Směrnice rady 92/43/EHS a druhy přílohy č. I Bernské úmluvy – Campanula gelida, Cerastium alsinifolium, Dianthus arenarius subsp. bohemicus, Galium sudeticum, Poa riphaea). Zbylé druhy (Carex viridula subsp. pseudoscandinavica, Dactylorhiza bohemica, Dianthus carthusianorum subsp. sudeticus, Knautia arvensis subsp. pseudolongifolia, Minuartia corcontica, Plantago atrata subsp. sudetica, Primula elatior subsp. corcontica, Saxifraga rosacea subsp. steinmannii, Sorbus sudetica) jsou až na dva případy endemity Vysokých Sudet a jejich ochrana je víceméně zaručena přísnou územní ochranou jejich stanovišť.
Druhy jsme na základě parametrů U a O rozdělili do skupin zmíněných v předchozí kapitole. Parametr U pracuje s procentickým úbytkem lokalit a nezohledňuje jejich celkový počet. Přitom je rozdíl, jestli se druh v minulosti vyskytoval na 100 lokalitách a nyní je jen na 50, nebo pokud rostl na dvou lokalitách a nyní je jen na jedné. V obou případech jde o pokles počtu lokalit o 50 %, ale v prvním případě je pokles početnosti mnohem markantnější. Proto jsme pro hodnocení druhů vzácných, které se v průběhu všech časových intervalů vyskytovaly v průměru v méně než šesti mapových polích, přijali dodatečné kritérium, které odlišilo druhy, u kterých došlo k poklesu počtu lokalit pouze o jedno až dvě mapová
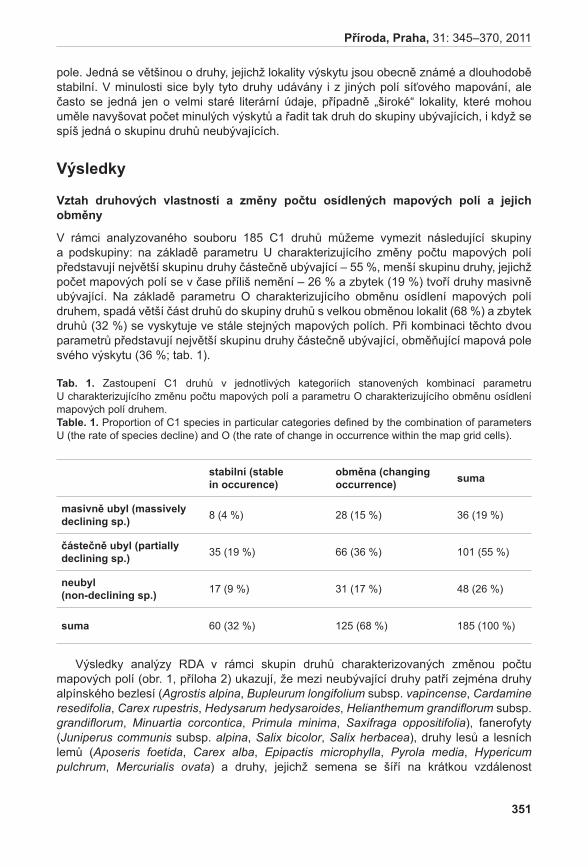
351
Příroda, Praha, 31: 345–370, 2011
pole. Jedná se většinou o druhy, jejichž lokality výskytu jsou obecně známé a dlouhodobě stabilní. V minulosti sice byly tyto druhy udávány i z jiných polí síťového mapování, ale často se jedná jen o velmi staré literární údaje, případně „široké“ lokality, které mohou uměle navyšovat počet minulých výskytů a řadit tak druh do skupiny ubývajících, i když se spíš jedná o skupinu druhů neubývajících.
výsledky
vztah druhových vlastností a změny počtu osídlených mapových polí a jejich obměny
V rámci analyzovaného souboru 185 C1 druhů můžeme vymezit následující skupiny a podskupiny: na základě parametru U charakterizujícího změny počtu mapových polí představují největší skupinu druhy částečně ubývající – 55 %, menší skupinu druhy, jejichž počet mapových polí se v čase příliš nemění – 26 % a zbytek (19 %) tvoří druhy masivně ubývající. Na základě parametru O charakterizujícího obměnu osídlení mapových polí druhem, spadá větší část druhů do skupiny druhů s velkou obměnou lokalit (68 %) a zbytek druhů (32 %) se vyskytuje ve stále stejných mapových polích. Při kombinaci těchto dvou parametrů představují největší skupinu druhy částečně ubývající, obměňující mapová pole svého výskytu (36 %; tab. 1).
tab. 1. Zastoupení C1 druhů v jednotlivých kategoriích stanovených kombinací parametru U charakterizujícího změnu počtu mapových polí a parametru O charakterizujícího obměnu osídlení mapových polí druhem.table. 1. Proportion of C1 species in particular categories defined by the combination of parameters U (the rate of species decline) and O (the rate of change in occurrence within the map grid cells).
stabilní (stable in occurence)
obměna (changing occurrence) suma
masivně ubyl (massively declining sp.) 8 (4 %) 28 (15 %) 36 (19 %)
částečně ubyl (partially declining sp.) 35 (19 %) 66 (36 %) 101 (55 %)
neubyl (non-declining sp.) 17 (9 %) 31 (17 %) 48 (26 %)
suma 60 (32 %) 125 (68 %) 185 (100 %)
Výsledky analýzy RDA v rámci skupin druhů charakterizovaných změnou počtu mapových polí (obr. 1, příloha 2) ukazují, že mezi neubývající druhy patří zejména druhy alpínského bezlesí (Agrostis alpina, Bupleurum longifolium subsp. vapincense, Cardamine resedifolia, Carex rupestris, Hedysarum hedysaroides, Helianthemum grandiflorum subsp. grandiflorum, Minuartia corcontica, Primula minima, Saxifraga oppositifolia), fanerofyty (Juniperus communis subsp. alpina, Salix bicolor, Salix herbacea), druhy lesů a lesních lemů (Aposeris foetida, Carex alba, Epipactis microphylla, Pyrola media, Hypericum pulchrum, Mercurialis ovata) a druhy, jejichž semena se šíří na krátkou vzdálenost
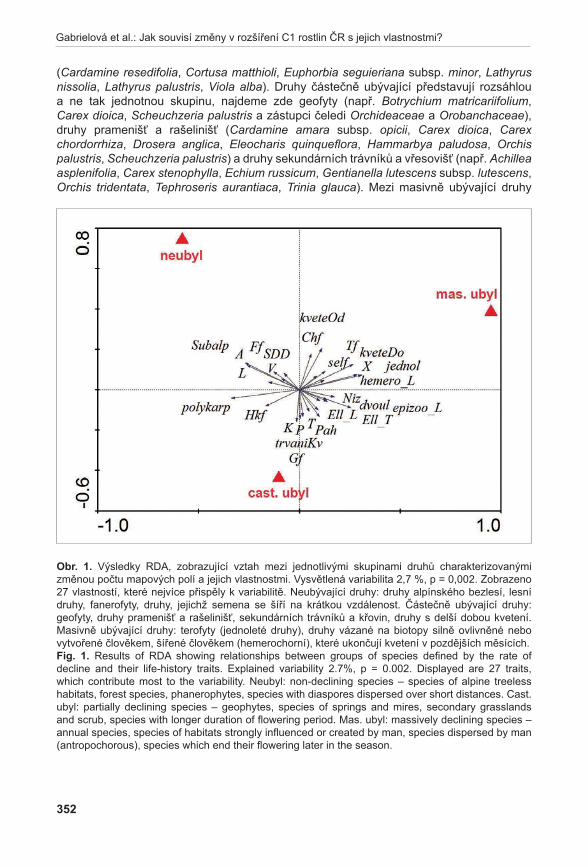
352
Gabrielová et al.: Jak souvisí změny v rozšíření C1 rostlin ČR s jejich vlastnostmi?
(Cardamine resedifolia, Cortusa matthioli, Euphorbia seguieriana subsp. minor, Lathyrus nissolia, Lathyrus palustris, Viola alba). Druhy částečně ubývající představují rozsáhlou a ne tak jednotnou skupinu, najdeme zde geofyty (např. Botrychium matricariifolium, Carex dioica, Scheuchzeria palustris a zástupci čeledi Orchideaceae a Orobanchaceae), druhy pramenišť a rašelinišť (Cardamine amara subsp. opicii, Carex dioica, Carex chordorrhiza, Drosera anglica, Eleocharis quinqueflora, Hammarbya paludosa, Orchis palustris, Scheuchzeria palustris) a druhy sekundárních trávníků a vřesovišť (např. Achillea asplenifolia, Carex stenophylla, Echium russicum, Gentianella lutescens subsp. lutescens, Orchis tridentata, Tephroseris aurantiaca, Trinia glauca). Mezi masivně ubývající druhy
obr. 1. Výsledky RDA, zobrazující vztah mezi jednotlivými skupinami druhů charakterizovanými změnou počtu mapových polí a jejich vlastnostmi. Vysvětlená variabilita 2,7 %, p = 0,002. Zobrazeno 27 vlastností, které nejvíce přispěly k variabilitě. Neubývající druhy: druhy alpínského bezlesí, lesní druhy, fanerofyty, druhy, jejichž semena se šíří na krátkou vzdálenost. Částečně ubývající druhy: geofyty, druhy pramenišť a rašelinišť, sekundárních trávníků a křovin, druhy s delší dobou kvetení. Masivně ubývající druhy: terofyty (jednoleté druhy), druhy vázané na biotopy silně ovlivněné nebo vytvořené člověkem, šířené člověkem (hemerochorní), které ukončují kvetení v pozdějších měsících.Fig. 1. Results of RDA showing relationships between groups of species defined by the rate of decline and their life-history traits. Explained variability 2.7%, p = 0.002. Displayed are 27 traits, which contribute most to the variability. Neubyl: non-declining species – species of alpine treeless habitats, forest species, phanerophytes, species with diaspores dispersed over short distances. Cast. ubyl: partially declining species – geophytes, species of springs and mires, secondary grasslands and scrub, species with longer duration of flowering period. Mas. ubyl: massively declining species – annual species, species of habitats strongly influenced or created by man, species dispersed by man (antropochorous), species which end their flowering later in the season.
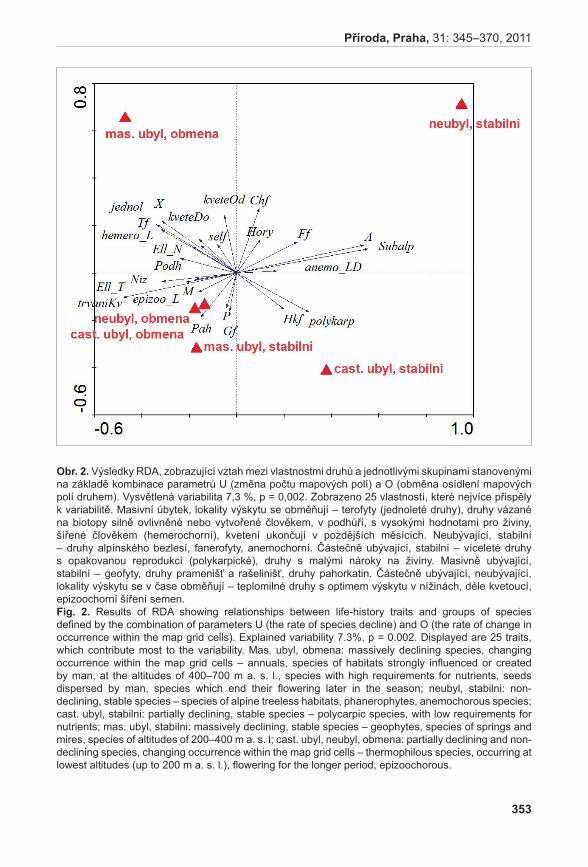
353
Příroda, Praha, 31: 345–370, 2011
obr. 2. Výsledky RDA, zobrazující vztah mezi vlastnostmi druhů a jednotlivými skupinami stanovenými na základě kombinace parametrů U (změna počtu mapových polí) a O (obměna osídlení mapových polí druhem). Vysvětlená variabilita 7,3 %, p = 0,002. Zobrazeno 25 vlastností, které nejvíce přispěly k variabilitě. Masivní úbytek, lokality výskytu se obměňují – terofyty (jednoleté druhy), druhy vázané na biotopy silně ovlivněné nebo vytvořené člověkem, v podhůří, s vysokými hodnotami pro živiny, šířené člověkem (hemerochorní), kvetení ukončují v pozdějších měsících. Neubývající, stabilní – druhy alpínského bezlesí, fanerofyty, anemochorní. Částečně ubývající, stabilní – víceleté druhy s opakovanou reprodukcí (polykarpické), druhy s malými nároky na živiny. Masivně ubývající, stabilní – geofyty, druhy pramenišť a rašelinišť, druhy pahorkatin. Částečně ubývající, neubývající, lokality výskytu se v čase obměňují – teplomilné druhy s optimem výskytu v nížinách, déle kvetoucí, epizoochorní šíření semen.Fig. 2. Results of RDA showing relationships between life-history traits and groups of species defined by the combination of parameters U (the rate of species decline) and O (the rate of change in occurrence within the map grid cells). Explained variability 7.3%, p = 0.002. Displayed are 25 traits, which contribute most to the variability. Mas. ubyl, obmena: massively declining species, changing occurrence within the map grid cells – annuals, species of habitats strongly influenced or created by man, at the altitudes of 400–700 m a. s. l., species with high requirements for nutrients, seeds dispersed by man, species which end their flowering later in the season; neubyl, stabilni: non-declining, stable species – species of alpine treeless habitats, phanerophytes, anemochorous species; cast. ubyl, stabilni: partially declining, stable species – polycarpic species, with low requirements for nutrients; mas. ubyl, stabilni: massively declining, stable species – geophytes, species of springs and mires, species of altitudes of 200–400 m a. s. l; cast. ubyl, neubyl, obmena: partially declining and non-declining species, changing occurrence within the map grid cells – thermophilous species, occurring at lowest altitudes (up to 200 m a. s. l.), flowering for the longer period, epizoochorous.

354
Gabrielová et al.: Jak souvisí změny v rozšíření C1 rostlin ČR s jejich vlastnostmi?
patří zejména terofyty (monokarpické druhy) – Androsace septentrionalis, Centaurium littorale subsp. compressum, Cyperus flavescens, Gentianella campestris subsp. baltica, Gentianella germanica subsp. germanica, Glaucium corniculatum, Chenopodium murale, Pseudognaphalium luteo-album, Pulicaria vulgaris, Radiola linoides, druhy vázané na biotopy silně ovlivněné nebo vytvořené člověkem (Agrostemma githago, Arnoseris minima, Atriplex rosea, Bromus arvensis, Bromus secalinus, Bromus squarrosus, Galium tricornutum, Chenopodium murale, Marrubium vulgare, Xanthium strumarium) nebo druhy alespoň částečně šířené činností člověka (hemerochorní) (Agrostemma githago, Arnoseris minima, Bromus arvensis, Bromus secalinus, Pulegium vulgare).
Výsledek analýzy RDA v rámci skupin vymezených na základě kombinace parametrů U (změna počtu mapových polí) a O (obměna osídlení mapových polí druhem) (obr. 2, příloha 2) potvrzuje a doplňuje poznatky získané v předchozí analýze. Mezi druhy masivně ubývající, které se v čase objevují v různých polích síťového mapování, patří zejména terofyty (jednoleté druhy), vázané na biotopy silně ovlivněné nebo vytvořené člověkem, s optimem výskytu v podhůří, s vysokými nároky na živiny, šířené člověkem (hemerochorní). Tyto vlastnosti dobře charakterizují plevele extenzivních polních kultur, které v poslední době značně ubyly vlivem intenzifikace zemědělství (Agrostemma githago, Bromus arvensis, Bromus racemosus, Bromus secalinus, Galium tricornutum, Glaucium corniculatum), případně i další ruderální archeofyty, které v poslední době ustoupily v důsledku změn v uspořádání synantropních stanovišť a z dalších, ne zcela objasněných příčin (Atriplex rosea, Chenopodium murale, Marrubium vulgare, Xanthium strumarium). Rovněž sem patří druhy vázané na vlhké obnažené substráty (např. letněná dna rybníků), případně i mírně zasolené (Centaurium littorale subsp. compressum, Elatine alsinastrum, Pseudognaphalium luteo-album, Pulegium vulgare, Pulicaria vulgaris, Radiola linoides).
Další dobře definovanou skupinu druhů představují druhy neubývající, stabilní (setrvávající na svých původních lokalitách), tedy druhy alpínského bezlesí, fanerofyty, často anemochorní. Jedná se o druhy zmíněné již výše – Agrostis alpina, Bupleurum longifolium subsp. vapincense, Hedysarum hedysaroides, Helianthemum grandiflorum subsp. grandiflorum, Juniperus communis subsp. alpina, Minuartia corcontica, Primula minima, Salix bicolor, Salix herbacea, Saxifraga oppositifolia. Druhy částečně ubývající, stabilní jsou charakterizovány jako víceleté druhy s opakovanou reprodukcí, s malými nároky na živiny (např. Adenophora liliifolia, Carex capillaris, Hammarbya paludosa, Jurinea cyanoides, Orchis palustris, Orchis tridentata, Pulsatilla patens, Pulsatilla vernalis, Schoenus ferrugineus, Schoenus nigricans, Thesium rostratum). Druhy masivně ubývající, stabilní nejsou zcela dobře definované, patří mezi ně osm druhů spíše vlhčích stanovišť s optimem výskytu v pahorkatinách (např. Cyperus flavescens, Gentiana verna, Gentianella campestris subsp. baltica, Potamogeton praelongus, Scorzonera parviflora, Tofieldia calyculata).
Zbylé dvě skupiny druhů částečně ubývajících či neubývajících, jejichž lokality výskytu se v čase obměňují, jsou převážně teplomilné druhy s optimem výskytu v nížinách, déle kvetoucí, se semeny šířenými epizoochorně. Jedná se např. o teplomilné plevele, jejichž ústup oproti minulosti nebyl tak dramatický (Adonis flammea, Bupleurum rotundifolium, Conringia orientalis, Erysimum repandum, Misopates orontium, Nigella arvensis, Papaver lecoqii, Ranunculus sardous), případně druhy slatin a slanisek (Bupleurum tenuissimum, Cladium mariscus, Glaux maritima, Juncus atratus, Juncus subnodulosus, Pulicaria dysenterica, Samolus valerandii) a řada vodních druhů (Groenlandia densa, Hippuris vulgaris, Najas minor, Nuphar pumila, Potamogeton × angustifolius, Trapa natans, Utricularia intermedia, Utricularia bremii, Wolffia arrhiza).
Výsledky analýzy GLM (tab. 2) ukazují, že úbytek druhu průkazně závisí na délce života rostlin, Ellenbergových indikačních hodnotách pro teplotu a okrajově signifikantně na typu
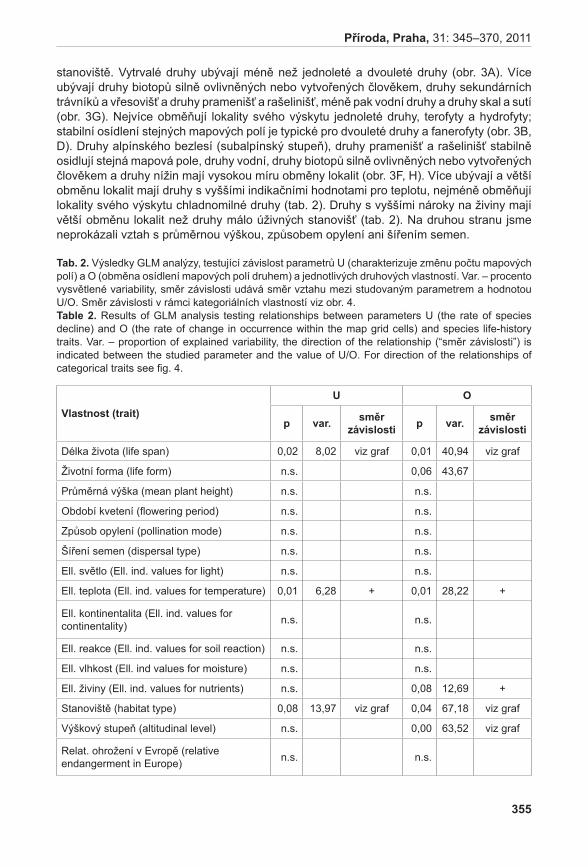
355
Příroda, Praha, 31: 345–370, 2011
stanoviště. Vytrvalé druhy ubývají méně než jednoleté a dvouleté druhy (obr. 3A). Více ubývají druhy biotopů silně ovlivněných nebo vytvořených člověkem, druhy sekundárních trávníků a vřesovišť a druhy pramenišť a rašelinišť, méně pak vodní druhy a druhy skal a sutí (obr. 3G). Nejvíce obměňují lokality svého výskytu jednoleté druhy, terofyty a hydrofyty; stabilní osídlení stejných mapových polí je typické pro dvouleté druhy a fanerofyty (obr. 3B, D). Druhy alpínského bezlesí (subalpínský stupeň), druhy pramenišť a rašelinišť stabilně osidlují stejná mapová pole, druhy vodní, druhy biotopů silně ovlivněných nebo vytvořených člověkem a druhy nížin mají vysokou míru obměny lokalit (obr. 3F, H). Více ubývají a větší obměnu lokalit mají druhy s vyššími indikačními hodnotami pro teplotu, nejméně obměňují lokality svého výskytu chladnomilné druhy (tab. 2). Druhy s vyššími nároky na živiny mají větší obměnu lokalit než druhy málo úživných stanovišť (tab. 2). Na druhou stranu jsme neprokázali vztah s průměrnou výškou, způsobem opylení ani šířením semen.
tab. 2. Výsledky GLM analýzy, testující závislost parametrů U (charakterizuje změnu počtu mapových polí) a O (obměna osídlení mapových polí druhem) a jednotlivých druhových vlastností. Var. – procento vysvětlené variability, směr závislosti udává směr vztahu mezi studovaným parametrem a hodnotou U/O. Směr závislosti v rámci kategoriálních vlastností viz obr. 4.table 2. Results of GLM analysis testing relationships between parameters U (the rate of species decline) and O (the rate of change in occurrence within the map grid cells) and species life-history traits. Var. – proportion of explained variability, the direction of the relationship (“směr závislosti”) is indicated between the studied parameter and the value of U/O. For direction of the relationships of categorical traits see fig. 4.
vlastnost (trait)u o
p var. směr závislosti p var. směr
závislosti
Délka života (life span) 0,02 8,02 viz graf 0,01 40,94 viz graf
Životní forma (life form) n.s. 0,06 43,67
Průměrná výška (mean plant height) n.s. n.s.
Období kvetení (flowering period) n.s. n.s.
Způsob opylení (pollination mode) n.s. n.s.
Šíření semen (dispersal type) n.s. n.s.
Ell. světlo (Ell. ind. values for light) n.s. n.s.
Ell. teplota (Ell. ind. values for temperature) 0,01 6,28 + 0,01 28,22 +
Ell. kontinentalita (Ell. ind. values for continentality) n.s. n.s.
Ell. reakce (Ell. ind. values for soil reaction) n.s. n.s.
Ell. vlhkost (Ell. ind values for moisture) n.s. n.s.
Ell. živiny (Ell. ind. values for nutrients) n.s. 0,08 12,69 +
Stanoviště (habitat type) 0,08 13,97 viz graf 0,04 67,18 viz graf
Výškový stupeň (altitudinal level) n.s. 0,00 63,52 viz graf
Relat. ohrožení v Evropě (relative endangerment in Europe) n.s. n.s.
356
Gabrielová et al.: Jak souvisí změny v rozšíření C1 rostlin ČR s jejich vlastnostmi?

357
Příroda, Praha, 31: 345–370, 2011
obr. 3. Závislost parametrů U (změna počtu mapových polí) a O (obměna osídlení mapových polí druhem) na druhových charakteristikách, jejichž vliv na parametry vyšel průkazně. Závislost úbytku (přibývání) druhů A) na délce života – více ubývají jednoleté a dvouleté druhy; C) na životní formě (neprůkazná závislost); E) na výškovém stupni (neprůkazná závislost); G) na typu stanoviště – nejvíce ubývají druhy biotopů silně ovlivněných nebo vytvořených člověkem (X) (okrajově průkazná závislost). Závislost obměny osídlení mapových polí druhem B) na délce života – nejvíce obměňují lokality svého výskytu jednoleté druhy; D) na životní formě – nejvíce stabilní jsou fanerofyty, nejvíce obměňují lokality svého výskytu terofyty a hydrofyty (okrajově průkazná závislost); F) na výškovém stupni – nejméně obměňují lokality svého výskytu druhy subalpínského stupně, nejvíce druhy podhůří; H) na typu stanoviště – nejvíce obměňují lokality svého výskytu druhy biotopů silně ovlivněných nebo vytvořených člověkem (X), nejvíce stabilní jsou druhy alpínského bezlesí (Alp).Fig. 3. Relationship between parameters U (the rate of species decline = “úbytek”) and O (the rate of change in occurrence within the map grid cells = “obměna”) and the species life-history traits, with significant effect on U and O. Relationship between species decline (U) and A) life span – annuals (“jednolete”) and biennials (“dvoulete”) are declining more than polycarpic species (“vytrvale”); C) life form (not significant); E) altitudinal level (not significant); G) habitat type – species of habitats strongly influenced or created by man (X) are declining the most (marginally significant). Relationship between the change in occurrence within the map grid cells (O) and B) life span – annuals (“jednolete”) are changing map grid cells the most; D) life form – phanerophytes are most stable, terophytes and hydrophytes are changing the map grid cells the most (marginally significant); F) altitudinal level – the most stable are species of altitudes above 1300 m a. s. l., species of altitudes of 400–700 m a. s. l. are changing map grid cells the most; H) the habitat type – species of habitats strongly influenced or created by man (X) are changing map grid cells the most, most stable are species of alpine treeless habitats (Alp). (For explanation of abbreviations see Appendix 1).
Zhodnocení priorit pro ochranu c1 druhů na základě údajů o historickém a současném rozšíření v ČR a ohrožení v ostatních evropských zemích
Jako nejohroženější se podle naší klasifikace jeví skupina druhů masově ubývajících, v Evropě ohrožených. Do této skupiny spadá pět hodnocených druhů: Gentianella campestris subsp. baltica, Campanula cervicaria, Himantoglossum adriaticum, Scorzonera parviflora a Spiranthes spiralis. Himantoglossum adriaticum patří do skupiny druhů, které byly na našem území i v minulosti extrémně vzácné a lokality v ČR jsou na severním okraji jeho areálu. Gentianella campestris subsp. baltica a Scorzonera parviflora patří mezi stabilní druhy, které se však dnes vyskytují již jen na zlomku svých původních lokalit. Campanula cervicaria a Spiranthes spiralis patří mezi druhy, které lokality svého výskytu spíše obměňují.
Skupina druhů částečně ubývajících, v Evropě rovněž ohrožených, zahrnuje 30 druhů, kterým by rovněž měla být věnována zvýšená pozornost. I v rámci nich lze rozlišit dvě podskupiny z hlediska obměny osídlení mapových polí druhem. První skupinou jsou druhy spíše obměňující lokality výskytu (15 druhů). Z těchto druhů patří dva mezi druhy na našem území vždy vzácné (označeny zkratkou vz. za názvem taxonu): Anacamptis pyramidalis, Botrychium matricariifolium, Carex buxbaumii, Carex chordorrhiza, Drosera anglica, Epipogium aphyllum, Eriophorum gracile, Gladiolus palustris, Groenlandia densa (vz.), Littorella uniflora, Malaxis monophyllos, Scheuchzeria palustris, Tillaea aquatica, Thesium ebracteatum (vz.), Trapa natans. Druhou skupinu představují druhy rostoucí převážně na lokalitách, na kterých se vyskytovaly i v minulosti (15 druhů, vzácné druhy označené vz.): Angelica palustris (vz.), Artemisia pancicii (vz.), Carex stenophylla, Cirsium brachycephalum (vz.), Hammarbya paludosa, Lathyrus pannonicus subsp. pannonicus (vz.), Laserpitium archangelica (vz.), Onosma arenaria (vz.), Ophrys insectifera, Orchis palustris, Orchis tridentata, Pedicularis exaltata (vz.), Pulsatilla patens, Rhynchospora fusca (vz.), Schoenus ferrugineus (vz.).

358
Gabrielová et al.: Jak souvisí změny v rozšíření C1 rostlin ČR s jejich vlastnostmi?
Informace o tom, jestli se druhy vyskytují spíše na lokalitách, na kterých rostly i v minulosti, nebo mají tendenci lokality svého výskytu obměňovat, je poměrně důležitá. Jako ohroženější se totiž jeví druhy, jejichž lokality se v průběhu času obměňují. Ochrana těchto druhů prostřednictvím ochrany jejich stanovišť nemusí být úspěšná – jejich výskyt je neprediktabilní a je pro ně normální, že lokality jejich výskytu zanikají a zase se objevují nové. Na druhou stranu ochrana druhé skupiny druhů, které mají tendenci vytrvávat na svých historických lokalitách, je mnohem méně problematická, protože lokality jejich výskytu jsou obecně známé a je pravděpodobné, že ochranou těchto lokalit (a případně vhodným managementem) docílíme i zachování těchto druhů v naší přírodě.
Je těžké určit, jaká skupina druhů by měla mít z hlediska ochrany přírody prioritu. Na jednu stranu je zřejmé, že za stejné časové a finanční úsilí věnované na záchranu jednoho druhu s nestabilním a neprediktabilním výskytem by bylo možné účinně chránit mnohem větší počet druhů „stabilních“ (např. druhů alpínského bezlesí). Ochrana druhů obměňujících lokality svého výskytu, vázaných na přechodná stanoviště, je nesnadnější, má méně jisté výsledky, vyžaduje více zdrojů, pečlivé plánování a odehrává se na širší škále. Rezignací v tomto směru bychom však ztratili celou řadu mobilních a zároveň specializovaných taxonů (např. i většinu druhů hmyzu), které jsou vázané na určité krajinné procesy a dynamickou obměnu stanovišť.
diskuse
Z několika podskupin vymezených v rámci C1 druhů na základě parametrů U – změna počtu mapových polí v jednotlivých časových vrstvách a O – obměna osídlení mapových polí druhem jsou dvě podskupiny dobře charakterizovány biologickými vlastnostmi a ekologickými nároky. Jedná se o skupinu druhů masivně ubývajících, obměňujících v čase lokality svého výskytu, která je tvořena především jednoletými druhy (terofyty), vázanými na biotopy, které jsou silně ovlivněné nebo vytvořené člověkem, druhy rostoucími v podhůří, s vysokými nároky na živiny a šířené člověkem (hemerochorní). Tyto vlastnosti dobře popisují dříve zcela běžné plevele a synantropní druhy, které z naší krajiny vymizely v důsledku intenzifikace zemědělství a ústupu od tradičního způsobu hospodaření. Druhou skupinu představují druhy neubývající, se stabilním výskytem na stejných lokalitách – převážně druhy alpínského bezlesí, které v ČR obývají několik málo vhodných lokalit v Krkonoších a Hrubém Jeseníku. Situace těchto druhů by při zachování územní ochrany měla být i nadále stabilizovaná, potenciální ohrožení mohou představovat snad jen výraznější klimatické změny.
Obecně se zdá, že biologické vlastnosti druhů s mírou jejich úbytku a mírou obměny lokalit souvisí mnohem méně než jejich stanovištní nároky. Ve výsledcích analýzy RDA byly sice různé skupiny charakterizovány i některými biologickými vlastnostmi – životní formou, délkou života, způsobem šíření semen nebo délkou trvání kvetení, tyto závislosti jsou ovšem často obtížně interpretovatelné. Při testování vztahu druhových vlastností a parametrů U a O pomocí zobecněných lineárních modelů vyšla průkazná závislost už jen u délky života resp. životní formy. Přitom výsledky porovnání vlastností druhů C1 a druhů blízce příbuzných hojných (Gabrielová et al. 2011) naznačují, že určitou roli by mohly hrát i vlastnosti jako průměrná výška nebo způsob opylení. Vliv těchto vlastností na míru úbytku druhu jsme však neprokázali, zato se opět potvrdil vliv stanoviště, které druh obývá, výškového stupně nebo nároků na teplotu či živiny.

359
Příroda, Praha, 31: 345–370, 2011
Stanovištní nároky druhů jsou v našich analýzách zachyceny jen velmi hrubě – rozdělení do devíti základních typů habitatů zdaleka nepostihuje rozmanitou škálu biotopů, které C1 druhy obývají. Zejména kategorie sekundární trávníky a vřesoviště je velmi široká, zahrnuje mnoho různých typů travních porostů od naprosto běžných ovsíkových nebo vlhkých pcháčových luk až po velmi vzácné a ohrožené kontinentální zaplavované louky, písčiny nebo slaniska. Na druhou stranu, i tato hrubá kategorizace potvrzuje trendy zjištěné i u jiných skupin organismů – ptáků a motýlů (reif et al. 2008, Pavlíková & konviČka in press). Kromě již zmiňovaných druhů alpínského bezlesí, které neubývají a vytrvávají na stále stejných lokalitách, jsou relativně stabilní – co do počtu lokalit – i lesní druhy. Vedle druhů stanovišť silně ovlivněných nebo vytvořených člověkem poměrně silně ubývají i druhy sekundárních trávníků a vřesovišť a druhy pramenišť a rašelinišť. Jak ukazují např. Pavlíková & konviČka (in press), i v rámci skupiny velkých nočních motýlů jsou více ohrožené druhy vázané na bezlesí, případně na otevřené řídké lesy než druhy uzavřených, hustých lesů. reif et al. (2008) v úvodu svého článku potvrzují obecný trend, že populace lesních druhů ptáků spíše narůstají, zatímco druhy vázané na zemědělsky obhospodařované biotopy spíše ubývají. Studie je ovšem zaměřena na závislosti v jemněji vymezených typech stanovišť, takže ukazuje, že i v rámci lesních druhů se najdou skupiny ubývající (vázané na horské jehličnaté lesy) a naopak mezi druhy zemědělské krajiny druhy přibývající.
Je jasné, že prezentované výsledky jsou do značné míry ovlivněny spolehlivostí údajů o úbytku a obměně mapových polí, definicí parametrů U a O a vymezením kategorií na základě těchto parametrů. Údaje o polích síťového mapování jsou zatím spíše předběžného charakteru, ale jsme přesvědčeni, že i přes veškeré nedostatky a komplikace (viz Černá et al. 2011) určitě podchycují alespoň základní trend ve změnách počtu lokalit – úbytek, ± stabilní počet mapových polí a přibývání – i v jejich obměně, tedy jestli má druh spíš tendenci zůstávat na svých historických lokalitách nebo osidlovat lokality nové.
Určitě má také smysl tyto dva parametry odlišovat, protože každý vypovídá o něčem jiném a může být vodítkem při rozhodování o možnostech praktické ochrany druhu. Úbytek počtu lokalit je úzce spjat s mírou ohrožení druhu – druhy masivně ubývající jsou více ohrožené než druhy vždy vzácné, ale co do počtu lokalit relativně stabilní. Úbytek počtu lokalit také zřejmě více souvisí se změnami v krajině, s mizením vhodných biotopů. Na druhou stranu obměna mapových polí popisuje jakousi dynamiku druhu v krajině na velkém prostorovém měřítku a mohla by souviset spíše s biologickými charakteristikami, jako je schopnost šíření. Dalo by se očekávat, že druhy s větší obměnou lokalit budou přizpůsobeny k šíření na delší vzdálenosti a bude se tedy jednat převážně o druhy anemochorní, zoochorní nebo hemerochorní a naopak druhy stabilní, vytrvávající na svých lokalitách, budou přizpůsobeny k šíření na kratší vzdálenost. Jenže stejně tak dobře může obměna lokalit souviset i s typem stanoviště, protože existují habitaty stabilní – např. právě alpínské bezlesí a lesy – nebo habitaty nestabilní (raná sukcesí stádia na nově vytvořených plochách po disturbanci, obnažená dna rybníků, štěrkové náplavy, extenzivní polní kultury) a druhy osidlující tato stanoviště sice musí mít vyvinuty patřičné mechanismy přizpůsobení se jejich dynamice, tyto mechanismy však nutně nemusí souviset s námi použitou klasifikací šíření semen. Každopádně je ochrana druhů s velkou obměnou lokalit problematická, protože jejich výskyt je nepředvídatelný, vázaný na stanoviště přechodného charakteru, která postupem času zanikají v důsledku přirozeného vývoje. Jedinou cestou udržení těchto druhů v naší krajině je zajistit, aby měly i v budoucnu dostatek potenciálně vhodných biotopů. Při omezených zdrojích se tak jako schůdnější jeví ochrana druhů stabilních, protože u nich je velká pravděpodobnost, že při zachování dosavadního charakteru biotopu buď pouhým nezasahováním nebo naopak vhodným managementem, vytrvají na svém stanovišti i nadále. Avšak ani na ochranu druhů s nestabilním výskytem bychom rozhodně neměli rezignovat.

360
Gabrielová et al.: Jak souvisí změny v rozšíření C1 rostlin ČR s jejich vlastnostmi?
shrnutí
V rámci souboru 185 analyzovaných C1 druhů jsme na základě míry úbytku druhu a obměny osídlení mapových polí druhem vymezili šest podskupin druhů s podobnými biologickými vlastnostmi a ekologickými nároky. Z těchto šesti podskupin jsou nejlépe charakterizované dvě podskupiny druhů: 1) druhy masivně ubývající, obměňující lokality svého výskytu – většinou jednoleté druhy vázané na biotopy ovlivněné nebo vytvořené člověkem, druhy s vysokými hodnotami pro živiny a druhy, jejichž semena jsou šířena člověkem. Tyto vlastnosti dobře popisují plevele extenzivních polních kultur, které v poslední době značně ubyly vlivem intenzifikace zemědělství a další ruderální archeofyty, které zmizely v důsledku změn v uspořádání synantropních stanovišť. 2) Stabilní druhy, vytrvávající na lokalitách svého výskytu – druhy alpínského bezlesí a lesů, fanerofyty a anemochorní druhy. Další dvě podskupiny druhů – druhy částečně ubývající a druhy stabilní, obě obměňující lokality svého výskytu, jsou si ve svých charakteristikách velmi blízké – jde převážně o teplomilné druhy s optimem výskytu v nížinách, déle kvetoucí, se semeny šířenými epizoochorně. Jedná se tedy opět o řadu teplomilných plevelů, jejichž úbytek oproti minulosti nebyl tak masivní, případně druhy slanisek a řadu vodních druhů. Míra úbytku druhů průkazně souvisí s délkou jejich života, s Ellenbergovými indikačními hodnotami pro teplotu a typem stanoviště. Se stejnými vlastnostmi průkazně souvisí i míra obměny lokalit, která je navíc závislá na životní formě druhu, Ellenbergových indikačních hodnotách pro živiny a na výškovém stupni. Vztah s dalšími charakteristikami, které hrají roli při odlišení vzácných a běžných druhů – např. s průměrnou výškou, délkou trvání kvetení, způsobem opylení nebo způsobem šíření semen, se nepotvrdil. Tento fakt svědčí o tom, že úbytek druhů a obměna jejich lokalit souvisí hlavně s jejich stanovištními nároky a s dalšími faktory, které se nám nepodařilo podchytit. Z hlediska ochrany přírody jsou důležité zejména vlastní údaje o míře úbytku druhu, obměně jeho lokalit a případně dalších parametrech, které už v současné době jsou nebo v brzké době budou o studovaných druzích dostupné – situace ohrožení v okolních zemích, velikost areálů rozšíření, pozice ČR v rámci těchto areálů nebo míra obsazenosti vhodných stanovišť. Na základě všech těchto údajů bude možné kvalifikovaně zhodnotit skutečnou míru ohrožení druhů a vybírat druhy pro prioritní ochranu.
summary
Based on the information about the rate of species decline and the rate of change in occurrence within the map grid cells, we defined six groups of species with similar biological and ecological traits within the analysed set of 185 critically endangered plant species. From the six groups, two are well characterised by their traits. The first group are massively declining species, which tend to occur in different map grid cells during the time. They are mostly annual species occurring on habitats strongly influenced or created by man, species with high requirements for nutrients and species with seeds dispersed by man. These traits are typical for weeds of extensive fields that have recently considerably decreased due to intensification of agriculture and other ruderal archaeophytes which disappeared as a result of changes in the habitats strongly influenced by human activities. The second group comprises non-declining, stable species, i.e. species of alpine treeless habitats and forests, phanerophytes, anemochorous species. The other two groups of species – partially declining or non-declining species, changing occurrence within the map grid cells during the time – are very similar in their traits. They are mostly thermophilic species,

361
Příroda, Praha, 31: 345–370, 2011
occurring in lowlands, flowering for a longer period, with adhesive dispersion of their seeds (epizoochorous). This is again characteristic for thermophilous weeds, whose decline was not as rapid as in the past, or for species of salt marshes and some aquatic species. The rate of species decline is significantly connected with their life span, Ellenberg indicator values for temperature, and the habitat type. The rate of change in occurrence within the map grid cells during the time is significantly connected with equal traits and, in addition, also with the Ellenberg indicator values for nutrients and with the same altitudinal level. The relationship with other traits that play a role in distinguishing rare and common species – such as mean height, duration of flowering, pollination mode or dispersal type – was not confirmed. This fact suggests that the decline of species and the change in their occurrence within the map grid cells is related primarily to their habitat requirements, and other factors that we failed to capture. The data most important for practical species conservation are mainly those providing information on the species decline and change in occurrence within the map grid cells. Along with the information on the state of endangerment in the other European countries, the size of the distribution range, the position of the Czech Republic within this range, and the extent of occupancy of available habitats, these data will provide sufficient background for a qualified assessment of the real degree of endangerment of the species and for selecting priority species for conservation.
Poděkování
Za pomoc se sběrem a zpracováním dat pro tento příspěvek děkujeme T. Fialové, Z. Kučerovi, V. Rybkovi a mnoha dalším nejmenovaným pomocníkům, kteří větší či menší měrou přispěli k excerpci a lokalizování nálezových dat. Za komentáře k rukopisu děkujeme T. Minárikové, M. Konvičkovi a účastníkům doktorandského posemináře na Katedře botaniky PřF UK. Studie byla financována z grantu MŠMT VaV 2B06178 a částečně podpořena grantem 206–08–H049, výzkumnými záměry MSM 0021620828 a AV0Z60050516.
literatura
bevill r. l. & louDa s. M. (1999): Comparisons of related rare and common species in the study of plant rarity. – Conserv. Biol. 13: 493–498.
Černá l., Gabrielová j. MünzberGová z. & rybka v. (2011): Změny v rozšíření kriticky ohrožených druhů rostlin v České republice na základě síťových map. – Příroda, Praha, 31: 281–298.
Denis r. l. h., hoDGson j. G., Grenyer r., shreeve t. G. & roy D. b. (2004): Host plants and butterfly biology. Do host-plant strategie drive butterfly status? – Ecol. Entomol. 29: 12–26.
DoMin k. (1942): Prodromus květeny mšenské. – Acta Bot. Bohem. 13 (1941): 1–236.farnsworth e. j. & oGurcak D. e. (2008): Functional groups of rare plants differ in levels of imperilment.
– Am. J. Bot. 95: 943–953.Gabrielová j., MünzberGová z., husáková i. & chrtek j. (2011): Kriticky ohrožené druhy rostlin ČR: Jak
se odlišují svými biologickými a ekologickými vlastnostmi od hojných druhů? – Příroda, Praha, 31: 247–279.
keller v. & bollMann k. (2004): From red list to species of conservation concern. – Conserv. Biol. 18: 1636–1644.
konviČka M., beneŠ j. & Čížek l. (2005): Ohrožený hmyz nelesních stanovišť: ochrana a management. – Sagittaria, Olomouc.
kunin w. e. & Gaston k. j. (1997): The biology of rarity. Causes and consequenses of rare-common differences. – Chapman & Hall, London.
Master l. l. (1991): Assessing threats and setting priorities for conservation. – Conserv. Biol. 5: 559–563.

362
Gabrielová et al.: Jak souvisí změny v rozšíření C1 rostlin ČR s jejich vlastnostmi?
Murray b. r., thrall P. h., Gill a. M. & nicotra a. b. (2002): How plant life-history and ecological traits relate to species rarity and commonness at varying spatial scales. – Austral Ecol. 27: 291–310.
Pavlíková a. & konviČka M. (in press): An ecological classification of Central European macromoths: habitat associations and conservation status returned from life history attributes. – J. Insect Conserv.
Pärtel M., kalaMees r., reier ü., tuvi e.-l., roosaluste e., vellak a. & zobel M. (2005): Grouping and priorization of vascular plant species for conservation: combining natural rarity and management. – Biol. Conserv. 123: 271–278.
Procházka f. [ed.] (2001): Černý a červený seznam cévnatých rostlin České republiky (stav v roce 2000). – Příroda, Praha, 18: 1–166.
rich t. c. G. & karran a. b. (2006): Floristic changes in the British Isles: comparison of techniques for assessing changes in frequency of plants with time. – Bot. J. Linn. Soc. 152: 279–301.
reif j., storch D., voříŠek P., Šťastný k. & bejČek v. (2008): Bird-habitat associations predict population trends in central European forest and farmland birds. – Biodivers. Conserv. 17: 3307–3319.
s−Plus (2000): Professional edition for Windows, release 2. − MathSoft, Inc., Massachusetts.shaffer h. b., fisher r. n. & DaviDson c. (1998): The role of natural history collections in documenting
species declines. – Trends Ecol. Evol. 13: 27–30.schnittler M. & Günther k.-f. (1999): Central European vascular plants requiring priority conservation
measures – an analysis from national Red Lists and distribution maps. – Biodivers. Conserv. 8: 891–925.
sutherlanD w. j., Pullin a. s., DolMan P. M. & kniGht t. M. (2004): The need for evidence-based conservation. – Trends Ecol. Evol. 19: 305–308.
ter braak c. j. & ŠMilauer P. (1998): CANOCO reference manual and user`s guide to Canoco for Windows: software for canonical community ordination, version 4.5. – Microcomputer Power, Ithaca, New York, USA.
walker k. j. & Preston ch. D. (2006): Ecological predictors of extinction risk in the flora of lowland England, UK. – Biodivers. Conserv. 15: 1913–1942.
363
Příroda, Praha, 31: 345–370, 2011
Příloha 1. Přehled zkratek analyzovaných vlastností.Appendix 1. Overview of abbreviations of analysed traits.
zkratka (abbreviation) vysvětlení explanation
% ohr. z vyskytu
podíl plochy evropských států, ve kterých je druh ohrožen, z celkové plochy evropských států, ve kterých se druh vyskytuje
proportion of area of European countries, in which a species is endangered, from the whole area of European countries in which a species occur
A typ stanoviště: alpinské bezlesí habitat type: alpine treeless habitats
anemo opylení: anemogamie (opylení větrem) pollination mode: anemogamy
anemo_LDD šíření semen: anemochorie (šíření větrem) dispersal type: anemochory
dvoul délka života: dvouleté rostliny life span: biennial
Ell_C Ellenbergovy indikační hodnoty pro kontinentalitu
Ellenberg indicator values for continentality
Ell_L Ellenbergovy indikační hodnoty pro světlo Ellenberg indicator values for light
Ell_M Ellenbergovy indikační hodnoty pro vlhkost Ellenberg indicator values for moisture
Ell_N Ellenbergovy indikační hodnoty pro živiny Ellenberg indicator values for nutrients
Ell_R Ellenbergovy indikační hodnoty pro půdní reakci
Ellenberg indicator values for soil reaction
Ell_T Ellenbergovy indikační hodnoty pro teplotu
Ellenberg indicator values for temperature
endozoo_LDD šíření semen: endozoochorie (šíření zvířaty po průchodu trávicím traktem) dispersal type: endozoochory
entomo opylení: entomogamie (opylení hmyzem) pollination mode: entomogamy
epizoo_LDD šíření semen: epizoochorie (šíření zvířaty na povrchu jejich těla) dispersal type: epizoochory
Ff životní forma fanerofyt life history: phanerophyte
Gf životní forma geofyt life history: geophyte
hemero_LDD šíření semen: hemerochorie (šíření člověkem) dispersal type: antropochory
Hf životní forma hydrofyt life history: hydrophyte
Hkf životní forma hemikryptofyt life history: hemicryptophyte
Hory výškový stupeň: hory (700–1300 m n. m.) altitudinal level: 700–1300 m a. s. l.

364
Gabrielová et al.: Jak souvisí změny v rozšíření C1 rostlin ČR s jejich vlastnostmi?
Příloha 1. PokračováníAppendix 1. Continue
zkratka (abbreviation) vysvětlení explanation
hydro_LDD šíření semen: hydrochorie (šíření vodou) dispersal type: hydrochory
Chf životní forma chamaefyt life history: chamaephyte
jednol délka života: jednoleté rostliny life span: annual
K typ stanoviště: křoviny habitat type: scrub
KveteDo konec kvetení (měsíc) ending of flowering (month)
KveteOd začátek kvetení (měsíc) beginning of flowering (month)
L typ stanoviště: lesy habitat type: forests
M typ stanoviště: mokřady a pobřežní vegetace
habitat type: wetlands and riverine vegetation
Niz výškový stupeň: nížiny (do 200 m n. m.) altitudinal level: under 200 m a. s. l.
ostatní opylení: geitonogamie, kleistogamie, pseudokleistogamie
pollination mode: geitonogamy, cleistogamy, pseudocleistogamy
P typ stanoviště: prameniště a rašeliniště habitat type: springs and mires
Pah výškový stupeň: pahorkatiny (200–400 m n. m.) altitudinal level: 200–400 m a. s. l.
Podh výškový stupeň: podhůří (400–700 m n. m.) altitudinal level: 400–700 m a. s. l.
polykarp délka života: polykarpické rostliny life span: polycarpic
prum_vyska průměrná výška (m) mean plant height (m)
S typ stanoviště: skály, sutě a jeskyně habitat type: cliffs and boulder screes
SDD šíření semen: šíření na krátkou vzdálenost dispersal type: short distance dispersal
self opylení: autogamie (samoopylení) pollination mode: autogamy
Subalp výškový stupeň: subalpínský (nad 1300 m n. m.) altitudinal level: above 1300 m a. s. l.
T typ stanoviště: sekundární trávníky a vřesoviště
habitat type: secondary grasslands and heathlands
Tf životní forma terofyt life history: terophyte
trvaniKv délka trvání kvetení v rozmezí měsíců duration of flowering (months)
V typ stanoviště: vodní toky a nádrže habitat type: streams and water bodies

365
Příroda, Praha, 31: 345–370, 2011
Příloha 1. PokračováníAppendix 1. Continue
zkratka (abbreviation) vysvětlení explanation
V typ stanoviště: vodní toky a nádrže habitat type: streams and water bodies
water opylení: hydrogamie (opylení vodou) pollination mode: hydrogamy
X typ stanoviště: biotopy silně ovlivněné nebo vytvořené člověkem
habitat type: habitats strongly influenced or created by man
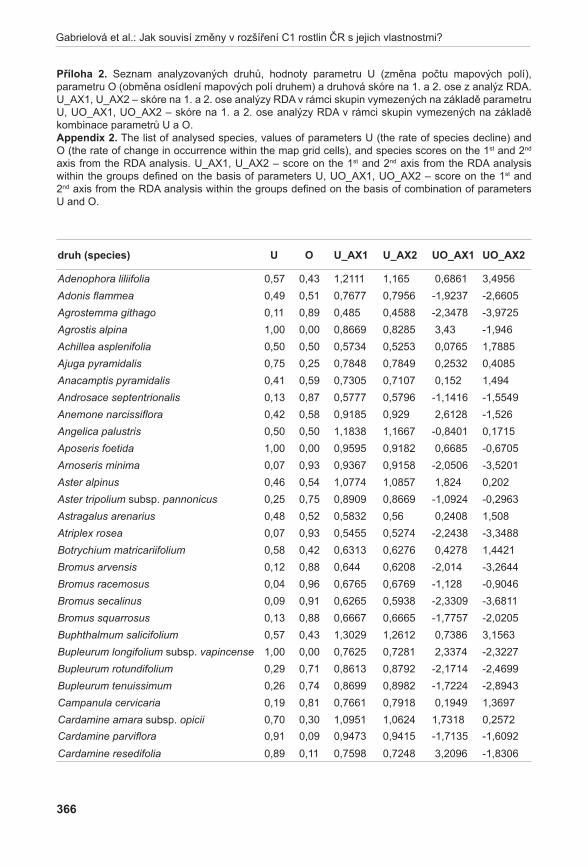
366
Gabrielová et al.: Jak souvisí změny v rozšíření C1 rostlin ČR s jejich vlastnostmi?
Příloha 2. Seznam analyzovaných druhů, hodnoty parametru U (změna počtu mapových polí), parametru O (obměna osídlení mapových polí druhem) a druhová skóre na 1. a 2. ose z analýz RDA. U_AX1, U_AX2 – skóre na 1. a 2. ose analýzy RDA v rámci skupin vymezených na základě parametru U, UO_AX1, UO_AX2 – skóre na 1. a 2. ose analýzy RDA v rámci skupin vymezených na základě kombinace parametrů U a O.Appendix 2. The list of analysed species, values of parameters U (the rate of species decline) and O (the rate of change in occurrence within the map grid cells), and species scores on the 1st and 2nd axis from the RDA analysis. U_AX1, U_AX2 – score on the 1st and 2nd axis from the RDA analysis within the groups defined on the basis of parameters U, UO_AX1, UO_AX2 – score on the 1st and 2nd axis from the RDA analysis within the groups defined on the basis of combination of parameters U and O.
druh (species) u o u_AX1 u_AX2 uo_AX1 uo_AX2
Adenophora liliifolia 0,57 0,43 1,2111 1,165 0,6861 3,4956Adonis flammea 0,49 0,51 0,7677 0,7956 -1,9237 -2,6605Agrostemma githago 0,11 0,89 0,485 0,4588 -2,3478 -3,9725Agrostis alpina 1,00 0,00 0,8669 0,8285 3,43 -1,946Achillea asplenifolia 0,50 0,50 0,5734 0,5253 0,0765 1,7885Ajuga pyramidalis 0,75 0,25 0,7848 0,7849 0,2532 0,4085Anacamptis pyramidalis 0,41 0,59 0,7305 0,7107 0,152 1,494Androsace septentrionalis 0,13 0,87 0,5777 0,5796 -1,1416 -1,5549Anemone narcissiflora 0,42 0,58 0,9185 0,929 2,6128 -1,526Angelica palustris 0,50 0,50 1,1838 1,1667 -0,8401 0,1715Aposeris foetida 1,00 0,00 0,9595 0,9182 0,6685 -0,6705Arnoseris minima 0,07 0,93 0,9367 0,9158 -2,0506 -3,5201Aster alpinus 0,46 0,54 1,0774 1,0857 1,824 0,202Aster tripolium subsp. pannonicus 0,25 0,75 0,8909 0,8669 -1,0924 -0,2963Astragalus arenarius 0,48 0,52 0,5832 0,56 0,2408 1,508Atriplex rosea 0,07 0,93 0,5455 0,5274 -2,2438 -3,3488Botrychium matricariifolium 0,58 0,42 0,6313 0,6276 0,4278 1,4421Bromus arvensis 0,12 0,88 0,644 0,6208 -2,014 -3,2644Bromus racemosus 0,04 0,96 0,6765 0,6769 -1,128 -0,9046Bromus secalinus 0,09 0,91 0,6265 0,5938 -2,3309 -3,6811Bromus squarrosus 0,13 0,88 0,6667 0,6665 -1,7757 -2,0205Buphthalmum salicifolium 0,57 0,43 1,3029 1,2612 0,7386 3,1563Bupleurum longifolium subsp. vapincense 1,00 0,00 0,7625 0,7281 2,3374 -2,3227Bupleurum rotundifolium 0,29 0,71 0,8613 0,8792 -2,1714 -2,4699Bupleurum tenuissimum 0,26 0,74 0,8699 0,8982 -1,7224 -2,8943Campanula cervicaria 0,19 0,81 0,7661 0,7918 0,1949 1,3697Cardamine amara subsp. opicii 0,70 0,30 1,0951 1,0624 1,7318 0,2572Cardamine parviflora 0,91 0,09 0,9473 0,9415 -1,7135 -1,6092Cardamine resedifolia 0,89 0,11 0,7598 0,7248 3,2096 -1,8306
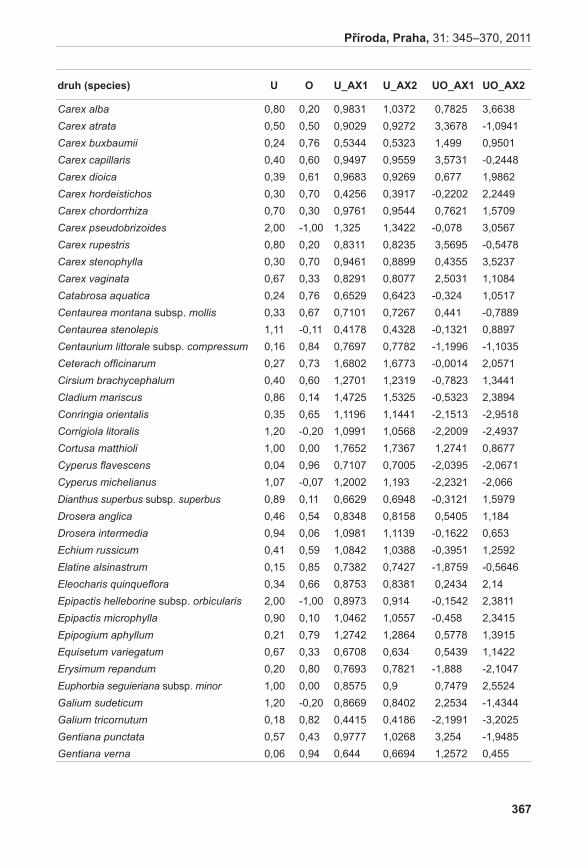
367
Příroda, Praha, 31: 345–370, 2011
druh (species) u o u_AX1 u_AX2 uo_AX1 uo_AX2
Carex alba 0,80 0,20 0,9831 1,0372 0,7825 3,6638Carex atrata 0,50 0,50 0,9029 0,9272 3,3678 -1,0941Carex buxbaumii 0,24 0,76 0,5344 0,5323 1,499 0,9501Carex capillaris 0,40 0,60 0,9497 0,9559 3,5731 -0,2448Carex dioica 0,39 0,61 0,9683 0,9269 0,677 1,9862Carex hordeistichos 0,30 0,70 0,4256 0,3917 -0,2202 2,2449Carex chordorrhiza 0,70 0,30 0,9761 0,9544 0,7621 1,5709Carex pseudobrizoides 2,00 -1,00 1,325 1,3422 -0,078 3,0567Carex rupestris 0,80 0,20 0,8311 0,8235 3,5695 -0,5478Carex stenophylla 0,30 0,70 0,9461 0,8899 0,4355 3,5237Carex vaginata 0,67 0,33 0,8291 0,8077 2,5031 1,1084Catabrosa aquatica 0,24 0,76 0,6529 0,6423 -0,324 1,0517Centaurea montana subsp. mollis 0,33 0,67 0,7101 0,7267 0,441 -0,7889Centaurea stenolepis 1,11 -0,11 0,4178 0,4328 -0,1321 0,8897Centaurium littorale subsp. compressum 0,16 0,84 0,7697 0,7782 -1,1996 -1,1035Ceterach officinarum 0,27 0,73 1,6802 1,6773 -0,0014 2,0571Cirsium brachycephalum 0,40 0,60 1,2701 1,2319 -0,7823 1,3441Cladium mariscus 0,86 0,14 1,4725 1,5325 -0,5323 2,3894Conringia orientalis 0,35 0,65 1,1196 1,1441 -2,1513 -2,9518Corrigiola litoralis 1,20 -0,20 1,0991 1,0568 -2,2009 -2,4937Cortusa matthioli 1,00 0,00 1,7652 1,7367 1,2741 0,8677Cyperus flavescens 0,04 0,96 0,7107 0,7005 -2,0395 -2,0671Cyperus michelianus 1,07 -0,07 1,2002 1,193 -2,2321 -2,066Dianthus superbus subsp. superbus 0,89 0,11 0,6629 0,6948 -0,3121 1,5979Drosera anglica 0,46 0,54 0,8348 0,8158 0,5405 1,184Drosera intermedia 0,94 0,06 1,0981 1,1139 -0,1622 0,653Echium russicum 0,41 0,59 1,0842 1,0388 -0,3951 1,2592Elatine alsinastrum 0,15 0,85 0,7382 0,7427 -1,8759 -0,5646Eleocharis quinqueflora 0,34 0,66 0,8753 0,8381 0,2434 2,14Epipactis helleborine subsp. orbicularis 2,00 -1,00 0,8973 0,914 -0,1542 2,3811Epipactis microphylla 0,90 0,10 1,0462 1,0557 -0,458 2,3415Epipogium aphyllum 0,21 0,79 1,2742 1,2864 0,5778 1,3915Equisetum variegatum 0,67 0,33 0,6708 0,634 0,5439 1,1422Erysimum repandum 0,20 0,80 0,7693 0,7821 -1,888 -2,1047Euphorbia seguieriana subsp. minor 1,00 0,00 0,8575 0,9 0,7479 2,5524Galium sudeticum 1,20 -0,20 0,8669 0,8402 2,2534 -1,4344Galium tricornutum 0,18 0,82 0,4415 0,4186 -2,1991 -3,2025Gentiana punctata 0,57 0,43 0,9777 1,0268 3,254 -1,9485Gentiana verna 0,06 0,94 0,644 0,6694 1,2572 0,455
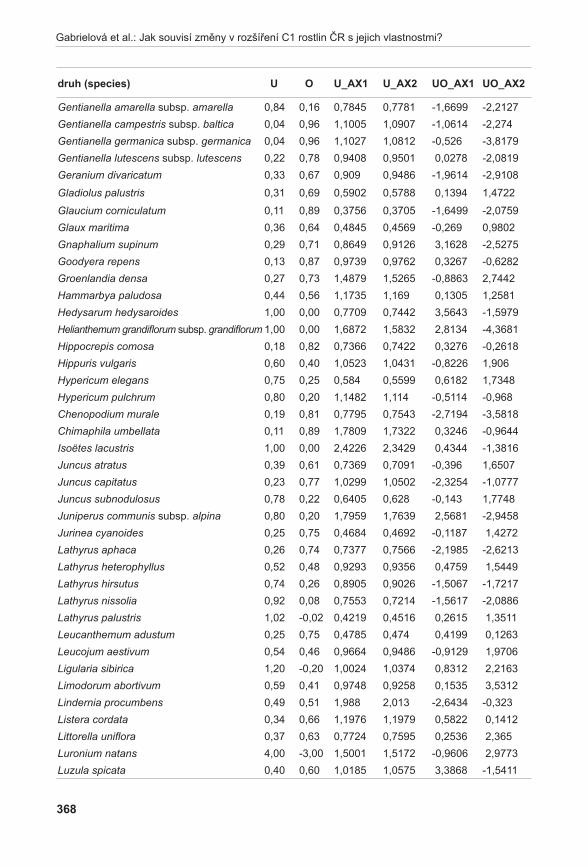
368
Gabrielová et al.: Jak souvisí změny v rozšíření C1 rostlin ČR s jejich vlastnostmi?
druh (species) u o u_AX1 u_AX2 uo_AX1 uo_AX2
Gentianella amarella subsp. amarella 0,84 0,16 0,7845 0,7781 -1,6699 -2,2127Gentianella campestris subsp. baltica 0,04 0,96 1,1005 1,0907 -1,0614 -2,274Gentianella germanica subsp. germanica 0,04 0,96 1,1027 1,0812 -0,526 -3,8179Gentianella lutescens subsp. lutescens 0,22 0,78 0,9408 0,9501 0,0278 -2,0819Geranium divaricatum 0,33 0,67 0,909 0,9486 -1,9614 -2,9108
Gladiolus palustris 0,31 0,69 0,5902 0,5788 0,1394 1,4722
Glaucium corniculatum 0,11 0,89 0,3756 0,3705 -1,6499 -2,0759Glaux maritima 0,36 0,64 0,4845 0,4569 -0,269 0,9802Gnaphalium supinum 0,29 0,71 0,8649 0,9126 3,1628 -2,5275Goodyera repens 0,13 0,87 0,9739 0,9762 0,3267 -0,6282Groenlandia densa 0,27 0,73 1,4879 1,5265 -0,8863 2,7442Hammarbya paludosa 0,44 0,56 1,1735 1,169 0,1305 1,2581Hedysarum hedysaroides 1,00 0,00 0,7709 0,7442 3,5643 -1,5979Helianthemum grandiflorum subsp. grandiflorum 1,00 0,00 1,6872 1,5832 2,8134 -4,3681Hippocrepis comosa 0,18 0,82 0,7366 0,7422 0,3276 -0,2618Hippuris vulgaris 0,60 0,40 1,0523 1,0431 -0,8226 1,906Hypericum elegans 0,75 0,25 0,584 0,5599 0,6182 1,7348Hypericum pulchrum 0,80 0,20 1,1482 1,114 -0,5114 -0,968Chenopodium murale 0,19 0,81 0,7795 0,7543 -2,7194 -3,5818Chimaphila umbellata 0,11 0,89 1,7809 1,7322 0,3246 -0,9644Isoëtes lacustris 1,00 0,00 2,4226 2,3429 0,4344 -1,3816Juncus atratus 0,39 0,61 0,7369 0,7091 -0,396 1,6507Juncus capitatus 0,23 0,77 1,0299 1,0502 -2,3254 -1,0777Juncus subnodulosus 0,78 0,22 0,6405 0,628 -0,143 1,7748Juniperus communis subsp. alpina 0,80 0,20 1,7959 1,7639 2,5681 -2,9458Jurinea cyanoides 0,25 0,75 0,4684 0,4692 -0,1187 1,4272Lathyrus aphaca 0,26 0,74 0,7377 0,7566 -2,1985 -2,6213Lathyrus heterophyllus 0,52 0,48 0,9293 0,9356 0,4759 1,5449Lathyrus hirsutus 0,74 0,26 0,8905 0,9026 -1,5067 -1,7217Lathyrus nissolia 0,92 0,08 0,7553 0,7214 -1,5617 -2,0886Lathyrus palustris 1,02 -0,02 0,4219 0,4516 0,2615 1,3511Leucanthemum adustum 0,25 0,75 0,4785 0,474 0,4199 0,1263Leucojum aestivum 0,54 0,46 0,9664 0,9486 -0,9129 1,9706Ligularia sibirica 1,20 -0,20 1,0024 1,0374 0,8312 2,2163Limodorum abortivum 0,59 0,41 0,9748 0,9258 0,1535 3,5312Lindernia procumbens 0,49 0,51 1,988 2,013 -2,6434 -0,323Listera cordata 0,34 0,66 1,1976 1,1979 0,5822 0,1412Littorella uniflora 0,37 0,63 0,7724 0,7595 0,2536 2,365Luronium natans 4,00 -3,00 1,5001 1,5172 -0,9606 2,9773Luzula spicata 0,40 0,60 1,0185 1,0575 3,3868 -1,5411
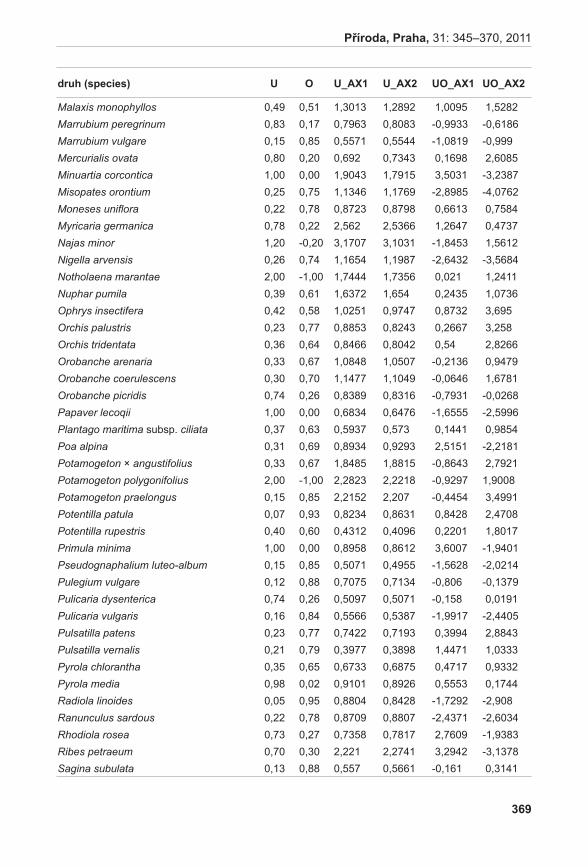
369
Příroda, Praha, 31: 345–370, 2011
druh (species) u o u_AX1 u_AX2 uo_AX1 uo_AX2
Malaxis monophyllos 0,49 0,51 1,3013 1,2892 1,0095 1,5282Marrubium peregrinum 0,83 0,17 0,7963 0,8083 -0,9933 -0,6186Marrubium vulgare 0,15 0,85 0,5571 0,5544 -1,0819 -0,999Mercurialis ovata 0,80 0,20 0,692 0,7343 0,1698 2,6085Minuartia corcontica 1,00 0,00 1,9043 1,7915 3,5031 -3,2387Misopates orontium 0,25 0,75 1,1346 1,1769 -2,8985 -4,0762Moneses uniflora 0,22 0,78 0,8723 0,8798 0,6613 0,7584Myricaria germanica 0,78 0,22 2,562 2,5366 1,2647 0,4737Najas minor 1,20 -0,20 3,1707 3,1031 -1,8453 1,5612Nigella arvensis 0,26 0,74 1,1654 1,1987 -2,6432 -3,5684Notholaena marantae 2,00 -1,00 1,7444 1,7356 0,021 1,2411Nuphar pumila 0,39 0,61 1,6372 1,654 0,2435 1,0736Ophrys insectifera 0,42 0,58 1,0251 0,9747 0,8732 3,695Orchis palustris 0,23 0,77 0,8853 0,8243 0,2667 3,258Orchis tridentata 0,36 0,64 0,8466 0,8042 0,54 2,8266Orobanche arenaria 0,33 0,67 1,0848 1,0507 -0,2136 0,9479Orobanche coerulescens 0,30 0,70 1,1477 1,1049 -0,0646 1,6781Orobanche picridis 0,74 0,26 0,8389 0,8316 -0,7931 -0,0268Papaver lecoqii 1,00 0,00 0,6834 0,6476 -1,6555 -2,5996Plantago maritima subsp. ciliata 0,37 0,63 0,5937 0,573 0,1441 0,9854Poa alpina 0,31 0,69 0,8934 0,9293 2,5151 -2,2181Potamogeton × angustifolius 0,33 0,67 1,8485 1,8815 -0,8643 2,7921Potamogeton polygonifolius 2,00 -1,00 2,2823 2,2218 -0,9297 1,9008Potamogeton praelongus 0,15 0,85 2,2152 2,207 -0,4454 3,4991Potentilla patula 0,07 0,93 0,8234 0,8631 0,8428 2,4708Potentilla rupestris 0,40 0,60 0,4312 0,4096 0,2201 1,8017Primula minima 1,00 0,00 0,8958 0,8612 3,6007 -1,9401Pseudognaphalium luteo-album 0,15 0,85 0,5071 0,4955 -1,5628 -2,0214Pulegium vulgare 0,12 0,88 0,7075 0,7134 -0,806 -0,1379Pulicaria dysenterica 0,74 0,26 0,5097 0,5071 -0,158 0,0191Pulicaria vulgaris 0,16 0,84 0,5566 0,5387 -1,9917 -2,4405Pulsatilla patens 0,23 0,77 0,7422 0,7193 0,3994 2,8843Pulsatilla vernalis 0,21 0,79 0,3977 0,3898 1,4471 1,0333Pyrola chlorantha 0,35 0,65 0,6733 0,6875 0,4717 0,9332Pyrola media 0,98 0,02 0,9101 0,8926 0,5553 0,1744Radiola linoides 0,05 0,95 0,8804 0,8428 -1,7292 -2,908 Ranunculus sardous 0,22 0,78 0,8709 0,8807 -2,4371 -2,6034Rhodiola rosea 0,73 0,27 0,7358 0,7817 2,7609 -1,9383Ribes petraeum 0,70 0,30 2,221 2,2741 3,2942 -3,1378Sagina subulata 0,13 0,88 0,557 0,5661 -0,161 0,3141
370
Gabrielová et al.: Jak souvisí změny v rozšíření C1 rostlin ČR s jejich vlastnostmi?
druh (species) u o u_AX1 u_AX2 uo_AX1 uo_AX2
Salix bicolor 1,00 0,00 1,7404 1,6916 4,1182 -2,5342Salix herbacea 1,00 0,00 1,5726 1,5009 4,1614 -3,5241Samolus valerandii 0,45 0,55 0,6772 0,6743 -0,5027 0,4488Saxifraga oppositifolia 1,00 0,00 0,8206 0,8083 3,5339 -0,7763Scorzonera parviflora 0,08 0,92 0,8389 0,8744 -0,017 2,459Scrophularia vernalis 0,78 0,22 0,8987 0,8887 -0,0487 1,8285Sedum villosum 0,06 0,94 1,087 1,0713 -0,3084 -2,2684Senecio doria 0,55 0,45 0,7799 0,7784 0,1559 0,2605Serratula lycopifolia 0,83 0,17 0,6575 0,7225 0,3775 2,3098Scheuchzeria palustris 0,64 0,36 1,1992 1,1531 -0,0789 2,1802Schoenus ferrugineus 0,50 0,50 0,5559 0,5205 0,4212 2,7123Schoenus nigricans 0,22 0,78 0,4637 0,428 0,0398 2,2409Spergularia maritima 0,48 0,52 0,5999 0,5934 -0,4072 0,0074Spergularia salina 0,40 0,60 0,6851 0,6858 -1,7007 -1,1234Spiranthes spiralis 0,03 0,97 0,9045 0,9092 -0,0806 -0,0752Tephroseris aurantiaca 0,57 0,43 0,4726 0,4632 0,7267 0,9036Thesium ebracteatum 0,50 0,50 0,601 0,594 0,1288 1,3987Thesium rostratum 0,33 0,67 0,67 0,6605 0,8551 2,6243Thymus pulegioides subsp. carniolicus 0,80 0,20 1,6671 1,6198 0,0396 -1,8724Tillaea aquatica 0,68 0,32 1,3423 1,3579 -2,0379 -0,0698Tofieldia calyculata 0,12 0,88 0,7923 0,8264 0,0774 1,946Trapa natans 0,60 0,40 2,3088 2,323 -2,2449 1,7856Trifolium badium 1,00 0,00 0,7782 0,8021 1,501 -2,0383Trifolium retusum 0,23 0,77 0,8887 0,8855 -1,5932 -1,4871Trifolium striatum 0,31 0,69 0,8042 0,7925 -1,2636 -1,0648Triglochin maritimum 0,07 0,93 0,6996 0,7241 -0,1569 1,5877Trinia glauca 0,35 0,65 1,2577 1,2016 0,5689 2,1915Utricularia bremii 1,00 0,00 1,4487 1,4499 -0,6749 1,9945Utricularia intermedia 0,60 0,40 1,2475 1,2643 -0,4061 1,8361Veronica bellidioides 0,33 0,67 1,0663 1,1133 3,0843 -2,8189Veronica opaca 0,28 0,72 1,5801 1,5891 -2,6499 -2,6579Viola alba 1,20 -0,20 1,1673 1,192 -0,3154 1,7791Viola elatior 0,40 0,60 0,8193 0,8043 -0,1024 2,6238Viola kitaibeliana 1,00 0,00 1,1262 1,1448 -1,2564 -1,0737Vulpia bromoides 0,11 0,89 0,8455 0,8615 -1,308 0,0759Wolffia arrhiza 0,91 0,09 1,4941 1,5085 -0,5167 2,3774Xanthium strumarium 0,19 0,81 0,5865 0,5689 -2,6598 -3,3641

Poznámky

Poznámky

















