Page 1
Využití molekulární cytogenetiky pro analýzu
dynamiky genomu a evoluce pohlavních
chromozómů u nižších obratlovců
Alexandr Sember
Ústav živočišné fyziologie a genetiky, AV ČR, v.v.i, Liběchov
Page 2
Cytogenetika v post-genomové době
Page 3
Laboratoř genetiky ryb
Page 4
Cytogenetika, evoluce a biodiverzita
74 ?
Diploid individuals: 2n=50
Page 5
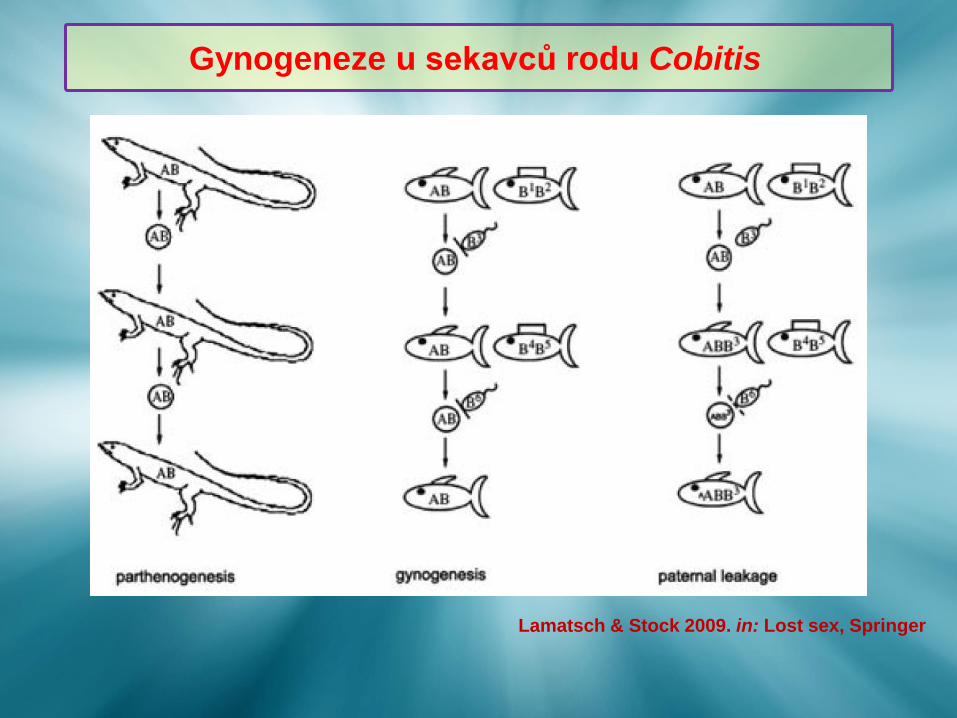
Gynogeneze u sekavců rodu Cobitis
Lamatsch & Stock 2009. in: Lost sex, Springer
Page 6
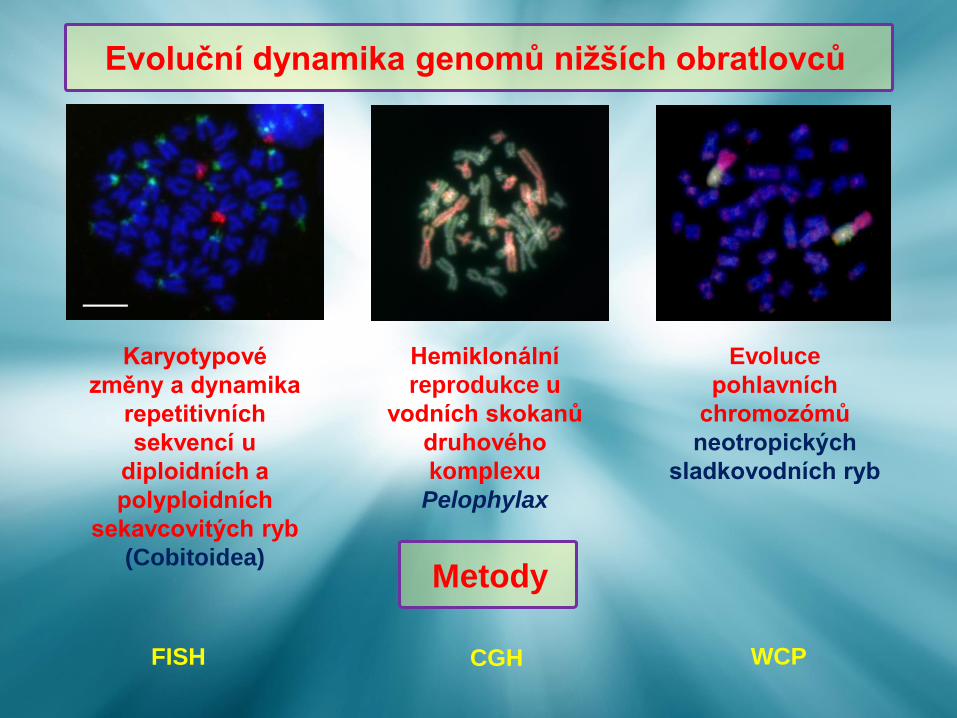
Evoluční dynamika genomů nižších obratlovců
Hemiklonální
reprodukce u
vodních skokanů
druhového
komplexu
Pelophylax
Evoluce
pohlavních
chromozómů
neotropických
sladkovodních ryb
Karyotypové
změny a dynamika
repetitivních
sekvencí u
diploidních a
polyploidních
sekavcovitých ryb
(Cobitoidea)Metody
FISH CGH WCP
Page 7
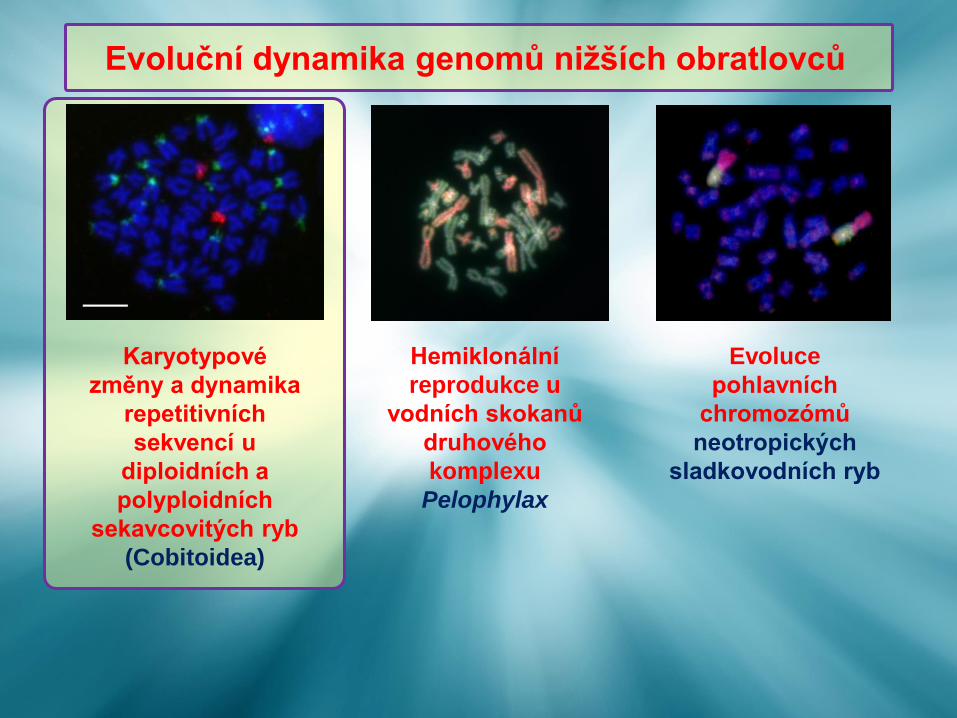
Evoluční dynamika genomů nižších obratlovců
Hemiklonální
reprodukce u
vodních skokanů
druhového
komplexu
Pelophylax
Evoluce
pohlavních
chromozómů
neotropických
sladkovodních ryb
Karyotypové
změny a dynamika
repetitivních
sekvencí u
diploidních a
polyploidních
sekavcovitých ryb
(Cobitoidea)
Page 8
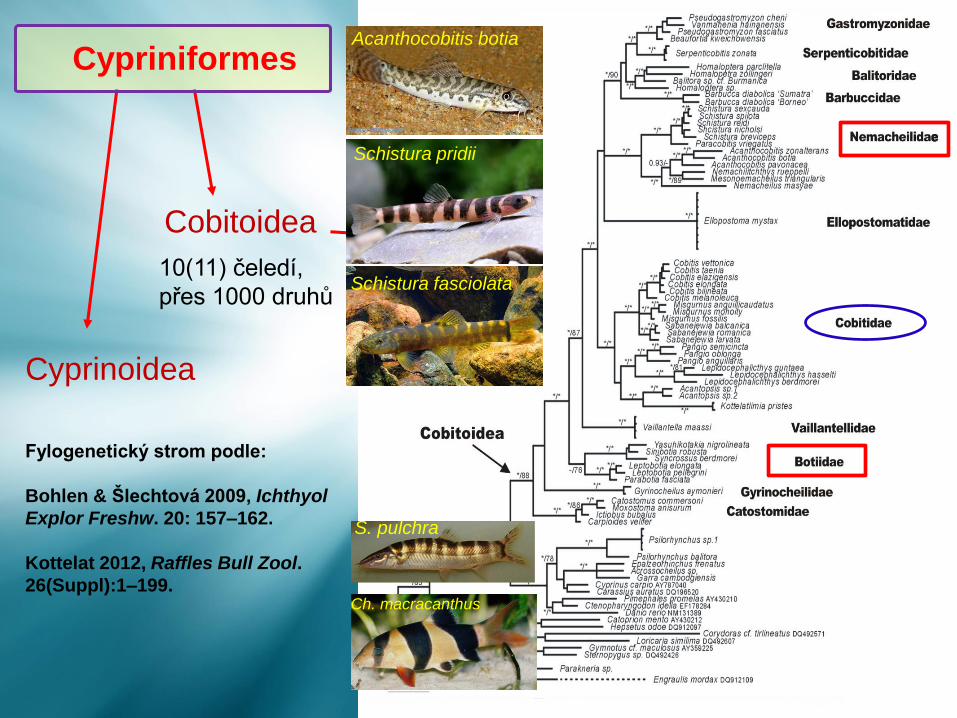
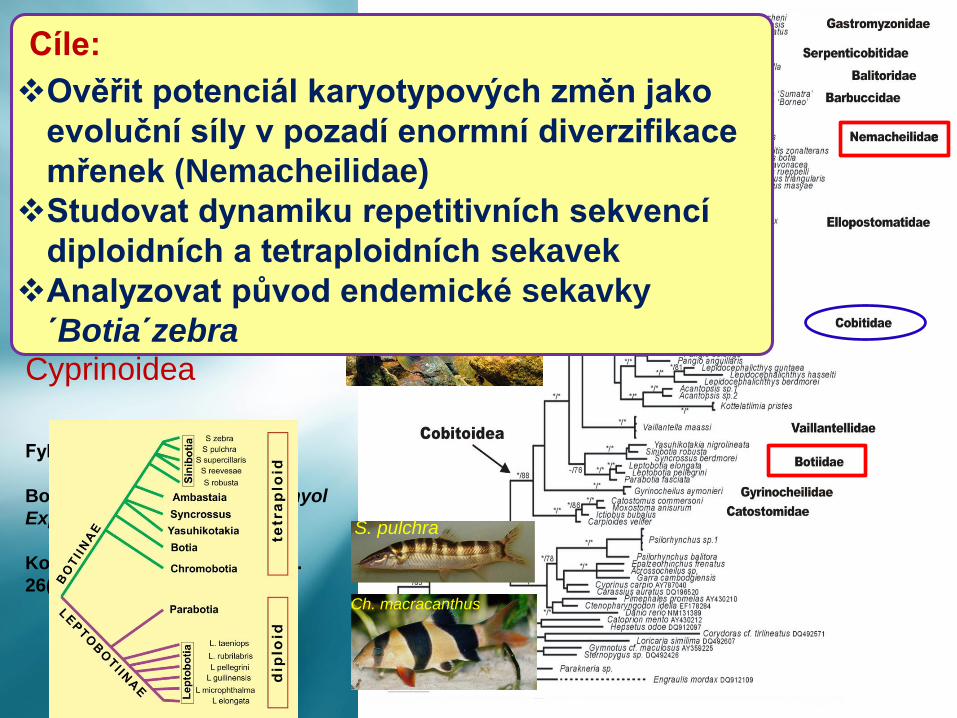
Cyprinoidea
Cobitoidea
10(11) čeledí,
přes 1000 druhů
Fylogenetický strom podle:
Bohlen & Šlechtová 2009, Ichthyol
Explor Freshw. 20: 157–162.
Kottelat 2012, Raffles Bull Zool.
26(Suppl):1–199.
CypriniformesAcanthocobitis botia
Schistura fasciolata
Schistura pridii
S. pulchra
Ch. macracanthus
Page 9
Cyprinoidea
Cobitoidea
10(11) čeledí,
přes 1000 druhů
Fylogenetický strom podle:
Bohlen & Šlechtová 2009, Ichthyol
Explor Freshw. 20: 157–162.
Kottelat 2012, Raffles Bull Zool.
26(Suppl):1–199.
CypriniformesAcanthocobitis botia
Schistura fasciolata
Schistura pridii
S. pulchra
Ch. macracanthus
Cíle:
❖Ověřit potenciál karyotypových změn jako
evoluční síly v pozadí enormní diverzifikace
mřenek (Nemacheilidae)
❖Studovat dynamiku repetitivních sekvencí
diploidních a tetraploidních sekavek
❖Analyzovat původ endemické sekavky
´Botia´zebra
Page 10
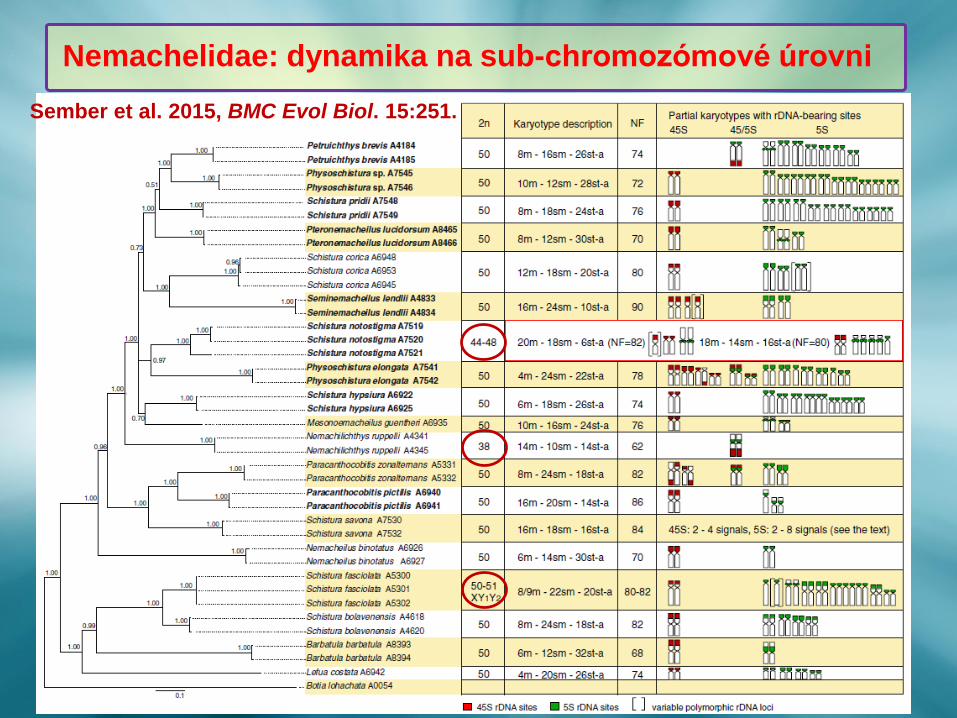
Nemachelidae: dynamika na sub-chromozómové úrovni
Sember et al. 2015, BMC Evol Biol. 15:251.
Page 11
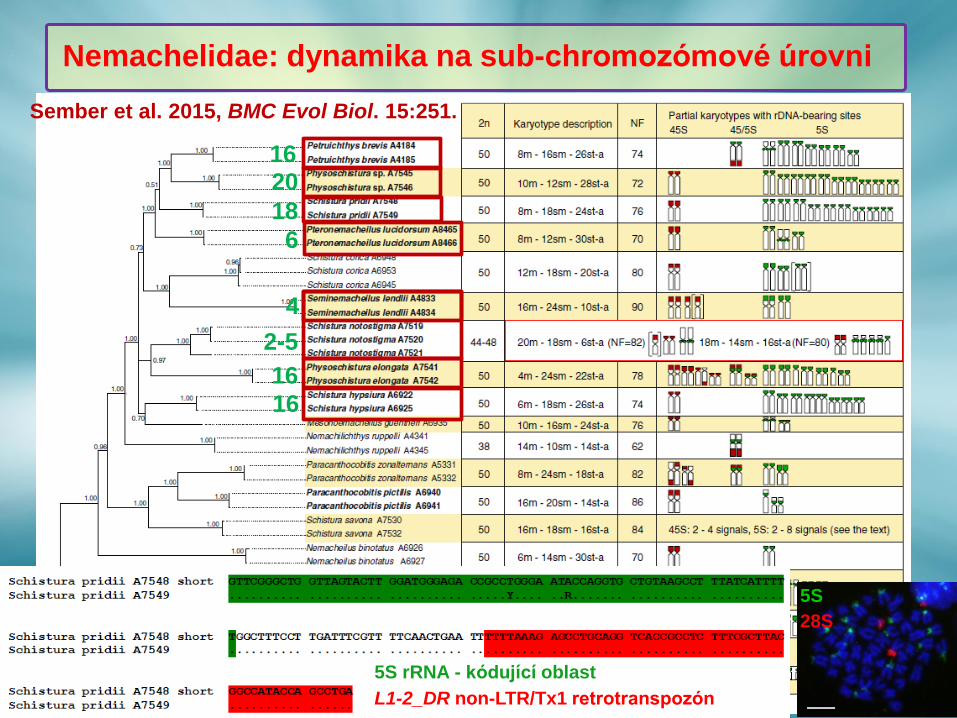
Nemachelidae: dynamika na sub-chromozómové úrovni
Sember et al. 2015, BMC Evol Biol. 15:251.
1620
18
6
4
2-5
16
16
5S
28S
5S rRNA - kódující oblast
L1-2_DR non-LTR/Tx1 retrotranspozón
Page 12
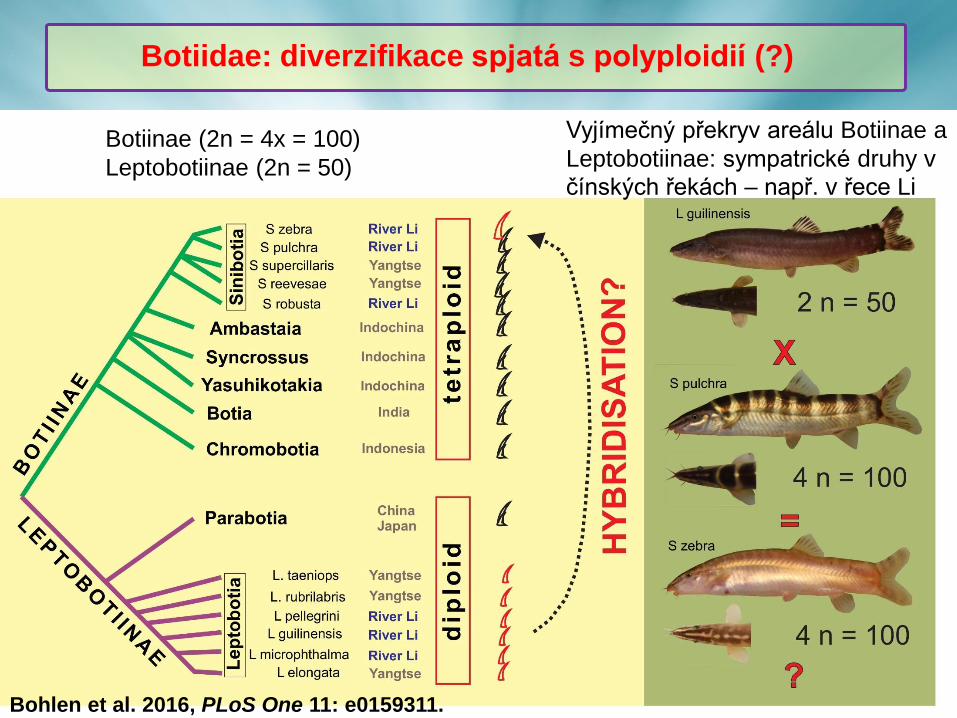
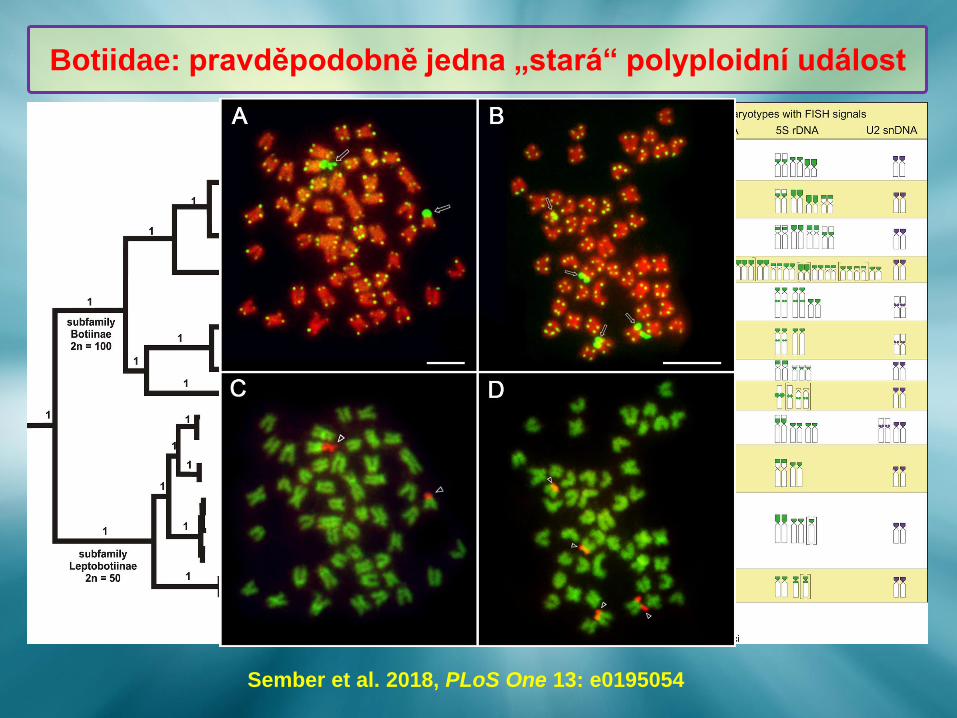
Botiinae (2n = 4x = 100)
Leptobotiinae (2n = 50)
Botiidae: diverzifikace spjatá s polyploidií (?)
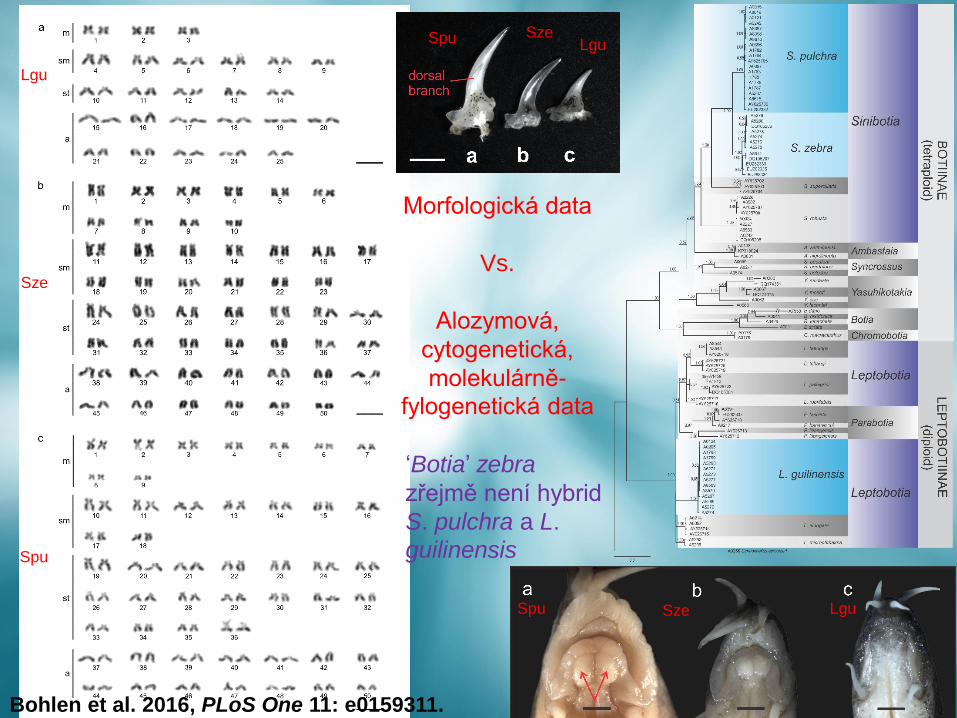
Bohlen et al. 2016, PLoS One 11: e0159311.
Vyjímečný překryv areálu Botiinae a
Leptobotiinae: sympatrické druhy v
čínských řekách – např. v řece Li
Page 13
Morfologická data
Vs.
Alozymová,
cytogenetická,
molekulárně-
fylogenetická data
‘Botia’ zebra
zřejmě není hybrid
S. pulchra a L.
guilinensis
Spu SzeLgu
Lgu
Spu Sze Lgu
Sze
Spu
Bohlen et al. 2016, PLoS One 11: e0159311.
Page 14
Botiidae: pravděpodobně jedna „stará“ polyploidní událost
Sember et al. 2018, PLoS One 13: e0195054
Page 15
Botiidae: pravděpodobně jedna „stará“ polyploidní událost
Sember et al. 2018, PLoS One 13: e0195054
Page 16
Evoluční dynamika genomů nižších obratlovců
Hemiklonální
reprodukce u
vodních skokanů
druhového
komplexu
Pelophylax
Evoluce
pohlavních
chromozómů
neotropických
sladkovodních ryb
Karyotypové
změny a dynamika
repetitivních
sekvencí u
diploidních a
polyploidních
sekavcovitých ryb
(Cobitoidea)
Page 17
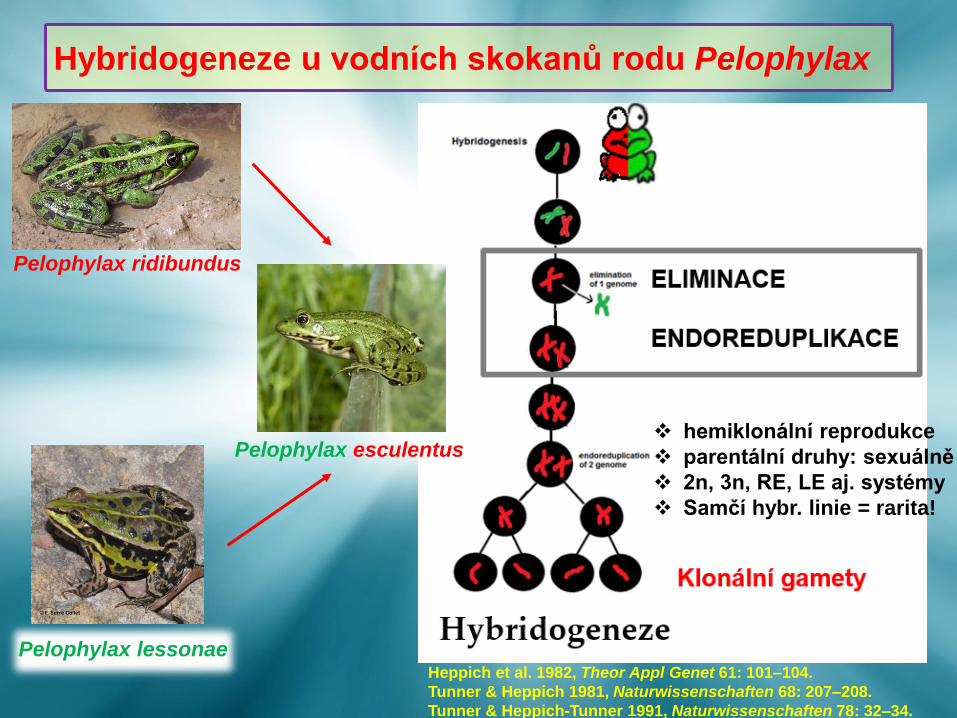
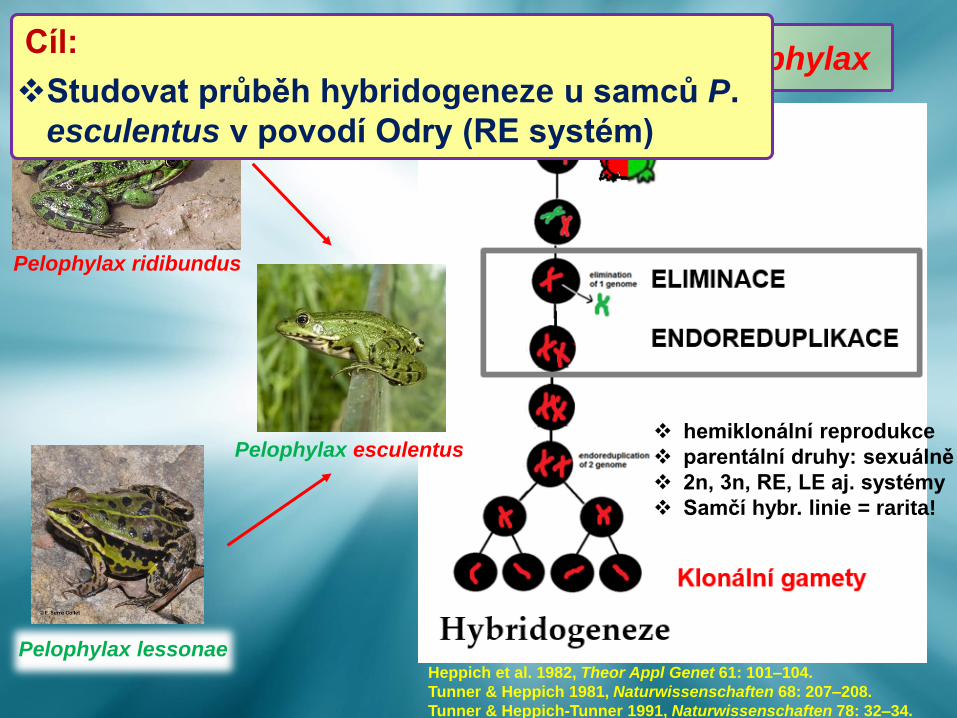
Pelophylax esculentus
Pelophylax ridibundus
Pelophylax lessonae
Hybridogeneze u vodních skokanů rodu Pelophylax
Heppich et al. 1982, Theor Appl Genet 61: 101–104.
Tunner & Heppich 1981, Naturwissenschaften 68: 207–208.
Tunner & Heppich-Tunner 1991, Naturwissenschaften 78: 32–34.
❖ hemiklonální reprodukce
❖ parentální druhy: sexuálně
❖ 2n, 3n, RE, LE aj. systémy
❖ Samčí hybr. linie = rarita!
Page 18
Pelophylax esculentus
Pelophylax ridibundus
Pelophylax lessonae
Hybridogeneze u vodních skokanů rodu PelophylaxCíl:
❖Studovat průběh hybridogeneze u samců P.
esculentus v povodí Odry (RE systém)
Heppich et al. 1982, Theor Appl Genet 61: 101–104.
Tunner & Heppich 1981, Naturwissenschaften 68: 207–208.
Tunner & Heppich-Tunner 1991, Naturwissenschaften 78: 32–34.
❖ hemiklonální reprodukce
❖ parentální druhy: sexuálně
❖ 2n, 3n, RE, LE aj. systémy
❖ Samčí hybr. linie = rarita!
Page 19
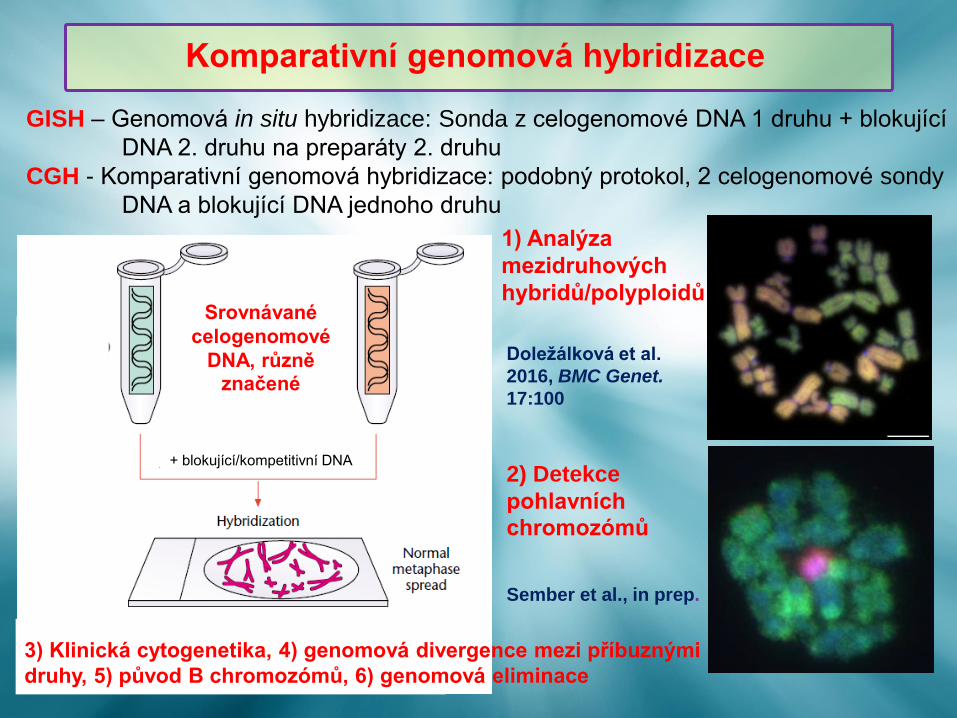
GISH – Genomová in situ hybridizace: Sonda z celogenomové DNA 1 druhu + blokující
DNA 2. druhu na preparáty 2. druhu
CGH - Komparativní genomová hybridizace: podobný protokol, 2 celogenomové sondy
DNA a blokující DNA jednoho druhu
+ blokující/kompetitivní DNA2) Detekce
pohlavních chromozómů
Sember et al., in prep.
Doležálková et al.
2016, BMC Genet.
17:100
1) Analýza
mezidruhových
hybridů/polyploidů
3) Klinická cytogenetika, 4) genomová divergence mezi příbuznými
druhy, 5) původ B chromozómů, 6) genomová eliminace
Komparativní genomová hybridizace
Srovnávané
celogenomové
DNA, různě značené
Page 20
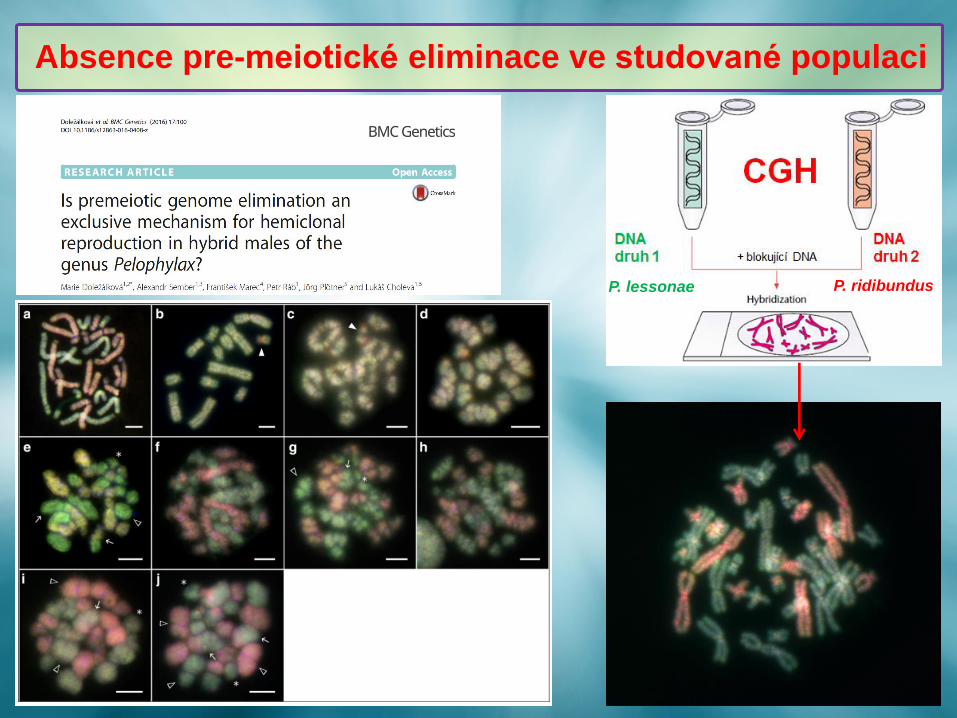
Absence pre-meiotické eliminace ve studované populaci
P. lessonae P. ridibundus
Page 21
Symonová R, Sember A, Majtánová Z, Ráb P.
(2015): Characterization of fish genomes by
GISH and CGH. In: Fish Cytogenetic
Techniques (Chondrichthyans and Teleosts),
Eds: Catherine Ozouf-Costaz, Eva Pisano,
Fausto Foresti, Lurdes Foresti de Almeida
Toledo CRC Press, Inc., Enfield, NH 03748,
USA
Popis možností využití metod GISH a CGH
v cytogenetické analýze ryb a optimalizace
protokolu pro analýzu vybraných skupin
paprskoploutvých ryb.
Page 22
Evoluční dynamika genomů nižších obratlovců
Hemiklonální
reprodukce u
vodních skokanů
druhového
komplexu
Pelophylax
Evoluce
pohlavních
chromozómů
neotropických
sladkovodních ryb
Karyotypové
změny a dynamika
repetitivních
sekvencí u
diploidních a
polyploidních
sekavcovitých ryb
(Cobitoidea)
Page 23
❖ nejbohatší sladkovodní ichtyofauna (cca 45% z celkového druhového
bohatství; cca 20-25% všech ryb)
❖ 17 řádů, přibližně 5 160 druhů
❖ převážně recentní diverzifikace (3-10 Mya; DNA barcoding; Pereira et al.,
2013, BMC Genet. 14:20)
❖ bohatá geologická historie, členitost vodních toků, překážky, izolované
populace
❖ Kryptické druhy, druhové komplexy❖ Apareiodon affinis
❖ Astyanax altiparanae
❖ Astyanax scabripinnis
❖ Bunocephalus corracoideus
❖ Erythrinus erythrinus
❖ Erythrinus erythrinus
❖ Geophagus brasiliensis
❖ Gymnotus carapo
❖ Iheringichthys labrosus
❖ Imparfinis spp.
❖ Piabina argentea
❖ Serrasalmus rhombeus
❖ Synbranchus marmoratus
Neotropická ichtyofauna, druhové komplexy
Page 24
Řád Čeleď Druh
Systém pohlavních
chromozómů Zdroj
Clupeiformes Clupeidae Brevorita aurea ♂X1X2Y/♀X1X1X2X2 Brum et al. 1992
Cypriniformes Parodontidae Apareiodon affinis ♂ZZ/ ♀zW1W2 Moreira-Filho et al. 1980, de Jesus et al. 1999
Gymnotiformes Hypopomidae Brachypopomus gauderio ♂X1X2Y/♀X1X1X2X2 Almeida-Toledo et al. 2000
Brachyhypopomus pinnicaudatus ♂X1X2Y/♀X1X1X2X2 Cardoso et al. 2015
Brachyhypopomus n. sp. FLAV ♂X1X2Y/♀X1X1X2X2 Cardoso et al. 2015
Gymnotidae Gymnotus bahianus ♂XY1Y2/♀XX Almeida et al. 2015
Gymnotus carapo ♂XY/♀XX Milhomem et al. 2008
Gymnotus coropinae ♂X1X2Y/♀X1X1X2X2 Silva et al. 2014
Gymnotus pantanal ♂X1X2Y/♀X1X1X2X2 Margarido et al. 2007
Gymnotus sp. ♂X1X2Y/♀X1X1X2X2 da Silva and Margarido 2005
Sternopygidae Eigenmannia aff. trilineata ♂ZZ/♀Z0 Araya-Jaime et al. 2017
Eigenmanni trilineata ♂X1X2Y/♀X1X1X2X2 Fernandes et al. 2010
Eigenmannia sp. B ♂XY/♀XX Silva et al. 2015
Eigenmannia sp.2 ♂X1X2Y/♀X1X1X2X2 Almeida-Toledo et al. 1984, 2001
Eigenmannia virescens ♂XY/♀XX Almeida-Toledo et al. 2001
Eigenmannia virescens ♂ZZ/♀ZW Almeida-Toledo et al. 2002, Silva et al. 2009
Characiformes Anostomidae Leporinus elongatus ♂Z1Z1Z2Z2/ ♀Z1W1Z2W2 Parise-Maltempi et al. 2007
Leporinus (9 druhů) ♂ZZ/♀ZW Oliveira et al. 2009
Crenuchidae Characidium (5 druhů) ♂ZZ/♀ZW Machado et al. 2011
Erythrinidae Hoplias malabaricus (D) ♂X1X2Y/♀X1X1X2X2 Bertollo et al. 1997
Hoplias malabaricus (G) ♂XY1Y2/♀XX Bertollo et al. 2000
Erythrinus erythrinus (B-D) ♂X1X2Y/♀X1X1X2X2 Bertollo et al. 2004
Triportheidae Triportheus (11 druhů) ♂ZZ/♀ZW Bertollo and Cavallaro 1992,Sanchez and
Jorge 1999, Artoni and Bertollo 2002, Yano
et al. 2017
Perciformes Coryphaenidae Coryphaena equielis * ♂X1X2Y/♀X1X1X2X2 Soares et al. 2014
Gobiidae Awaous strigatus ♂X1X2Y/♀X1X1X2X2 Oliveira and Almeida Toledo 2006
Pleuronectiformes Achiridae Achirus achirus ♂X1X2Y/♀X1X1X2X2 Bitencourt et al. 2016
Siluriformes Aspredinidae Bunocephalus coracoideus ♂X1Y1X2Y2/♀X1X1X2X2 Ferreira et al. 2016
Loricariidae Ancistrus dubius ♂XY1Y2/♀XX Oliveira et al. 2008
Ancistrus cf. dubius ♂ZZ/♀ZW nebo ♂XY/♀XX Mariotto et al. 2004, 2006
Ancistrus dolichopterus ♂Z1Z1Z2Z2/ ♀Z1W1Z2W2 Oliveira et al. 2008
Ancistrus n. sp. ♂X0/♀XX Alves et al. 2006
Ancistrus (3 species) ♂ZZ/♀ZW Oliveira et al. 2007, Takagui et al. 2017
Ancistrus (2 species) ♂XY/♀XX Oliveira et al. 2008, 2009, Favarato et al. 2015
Loricariidae Hartia carvalhoi ♂XY1Y2/♀XX Centofante et al. 2006
Hartia punctata ♂X1X2Y/♀X1X1X2X2 Blanco et al. 2014
Tetraodontiformes Monacanthidae Stephanolepis seiter * ♂X1X2Y/♀X1X1X2X2 Nirchio and Oliveira 2007
Apareiodon affinis
♂XY/♀XX
♂XY/♀XX
♂XY1Y2/♀XX
♂X1X2Y/♀X1X1X2X2
♂XY1X2Y2/♀X1X1X2X2
♂ZZ/♀ZW
♂ZZ/♀Z0
♂ZZ/ ♀ZW1W2
♂Z1Z1Z2Z2/♀Z1W1Z2W2
~ 65 %
Heteromorfní pohlavní
chromozómy u studovaných
druhů:
❖ 5% Teleostei
❖ 6% neotropických ryb
* = příklady mořských ryb
Pohlavní chromozómy neotropických ryb
Page 25
❖ tři rody:
❖ Erythrinus
❖ Hoplias
❖ Hoplerythrinus
❖ alespoň 18 popsaných
druhů, výhradně JA
Erythrinidae (Characiformes)
Page 26
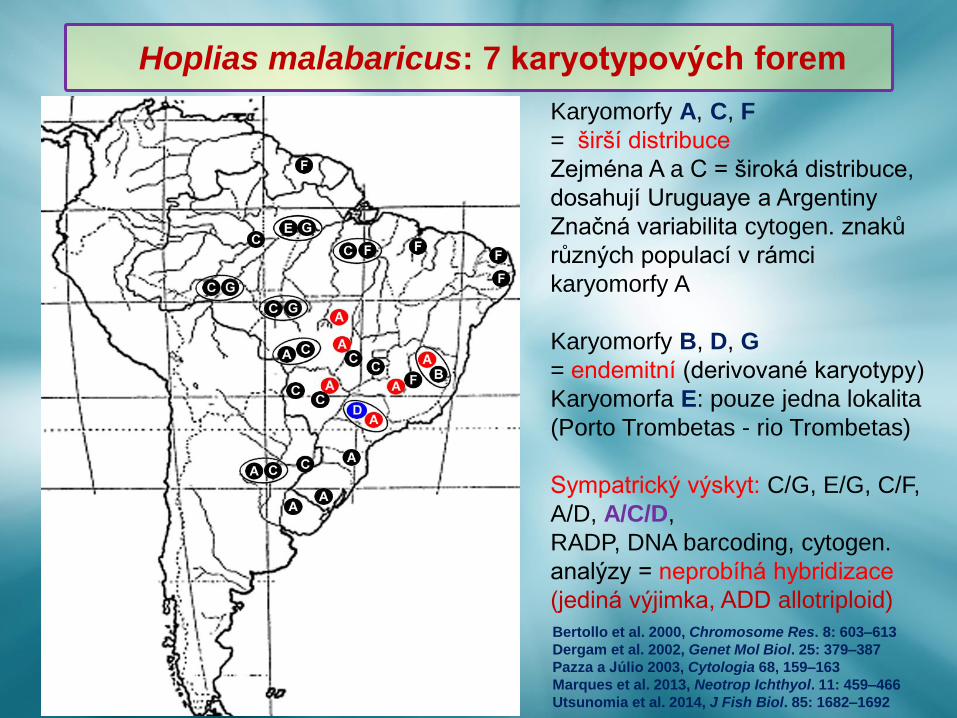
Karyomorfy A, C, F
= širší distribuce
Zejména A a C = široká distribuce,
dosahují Uruguaye a Argentiny
Značná variabilita cytogen. znaků
různých populací v rámci
karyomorfy A
Karyomorfy B, D, G
= endemitní (derivované karyotypy)
Karyomorfa E: pouze jedna lokalita
(Porto Trombetas - rio Trombetas)
Sympatrický výskyt: C/G, E/G, C/F,
A/D, A/C/D,
RADP, DNA barcoding, cytogen.
analýzy = neprobíhá hybridizace
(jediná výjimka, ADD allotriploid)
Bertollo et al. 2000, Chromosome Res. 8: 603–613
Dergam et al. 2002, Genet Mol Biol. 25: 379–387
Pazza a Júlio 2003, Cytologia 68, 159–163
Marques et al. 2013, Neotrop Ichthyol. 11: 459–466
Utsunomia et al. 2014, J Fish Biol. 85: 1682–1692
Hoplias malabaricus: 7 karyotypových forem
Page 27
27
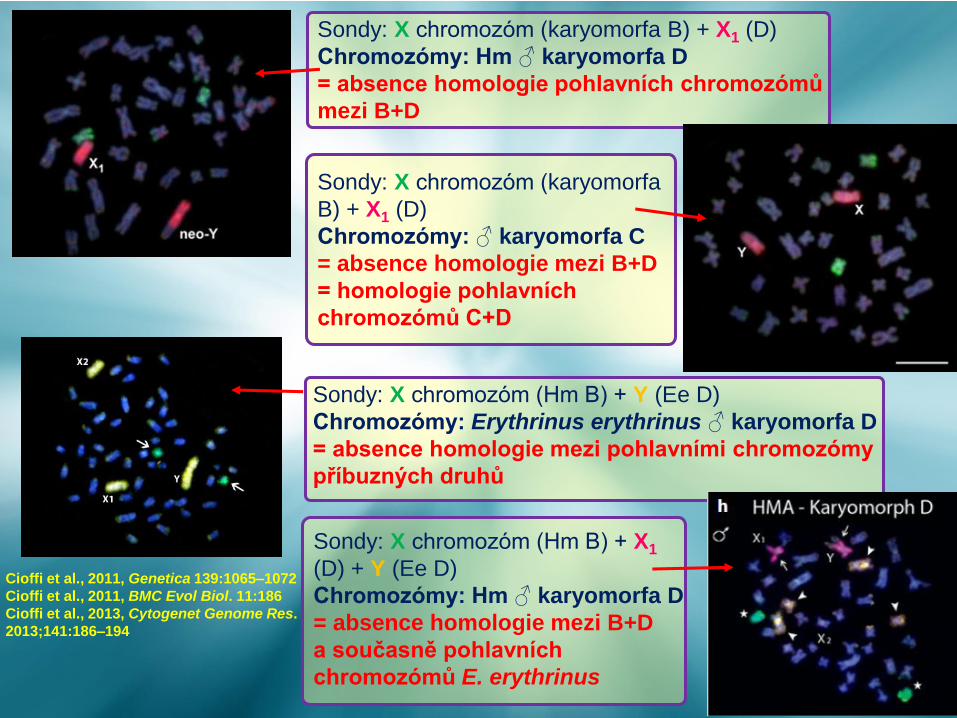
Cioffi et al., 2011, Genetica 139:1065–1072
Cioffi et al., 2011, BMC Evol Biol. 11:186
Cioffi et al., 2013, Cytogenet Genome Res.
2013;141:186–194
Sondy: X chromozóm (karyomorfa B) + X1 (D)
Chromozómy: Hm ♂ karyomorfa D
= absence homologie pohlavních chromozómů
mezi B+D
Sondy: X chromozóm (karyomorfa
B) + X1 (D)
Chromozómy: ♂ karyomorfa C
= absence homologie mezi B+D
= homologie pohlavních
chromozómů C+D
Sondy: X chromozóm (Hm B) + Y (Ee D)
Chromozómy: Erythrinus erythrinus ♂ karyomorfa D
= absence homologie mezi pohlavními chromozómy
příbuzných druhů
Sondy: X chromozóm (Hm B) + X1
(D) + Y (Ee D)
Chromozómy: Hm ♂ karyomorfa D
= absence homologie mezi B+D
a současně pohlavních
chromozómů E. erythrinus
Page 28
Federal University of São Carlos, Brazil
Prof. Dr. Marcelo de Bello Cioffi
Prof. Dr. Luiz Antônio Carlos Bertollo
Page 29
Federal University of São Carlos, Brazil
Prof. Dr. Marcelo de Bello Cioffi
Prof. Dr. Luiz Antônio Carlos Bertollo
Cíle:
❖Identifikovat přítomnost homomorfních
pohlavních chromozómů (karyomorfy A + F)
❖Detekovat pohlavně-specifické oblasti na
nascentních (C), diferencovaných (B) a neo-
pohlavních (D+G) pohlavních chromozómech
❖Objasnit mechanizmus vzniku XY1Y2 u
karyomorfy G
❖Objasnit míru homologie mezi pohlavními
systémy (WCP)
Page 30
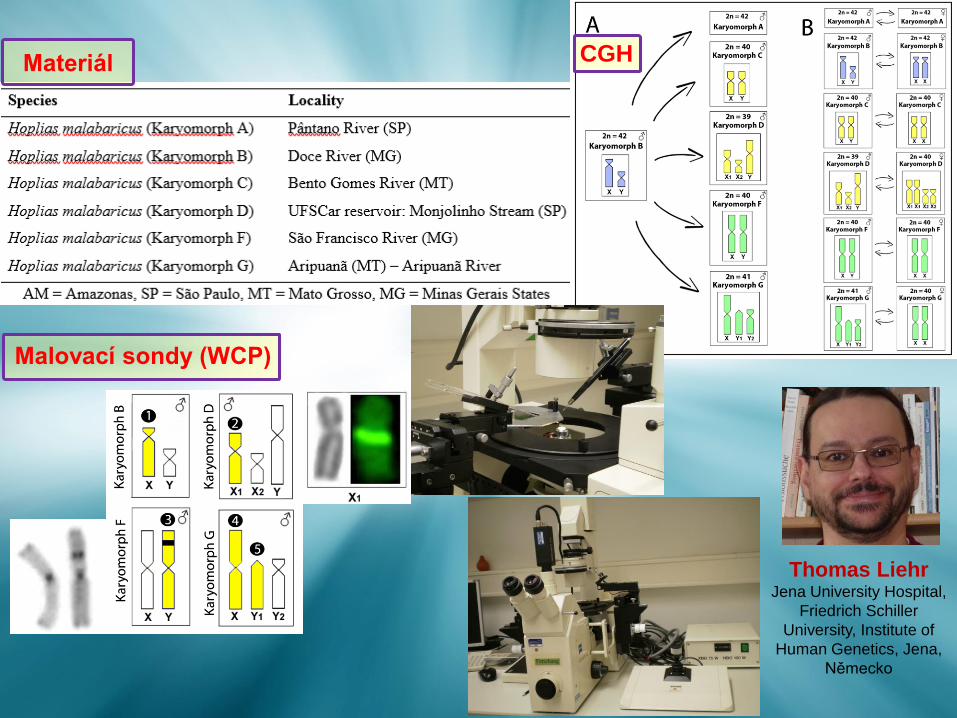
Thomas LiehrJena University Hospital,
Friedrich Schiller
University, Institute of
Human Genetics, Jena,
Německo
CGHMateriál
Malovací sondy (WCP)
Page 31
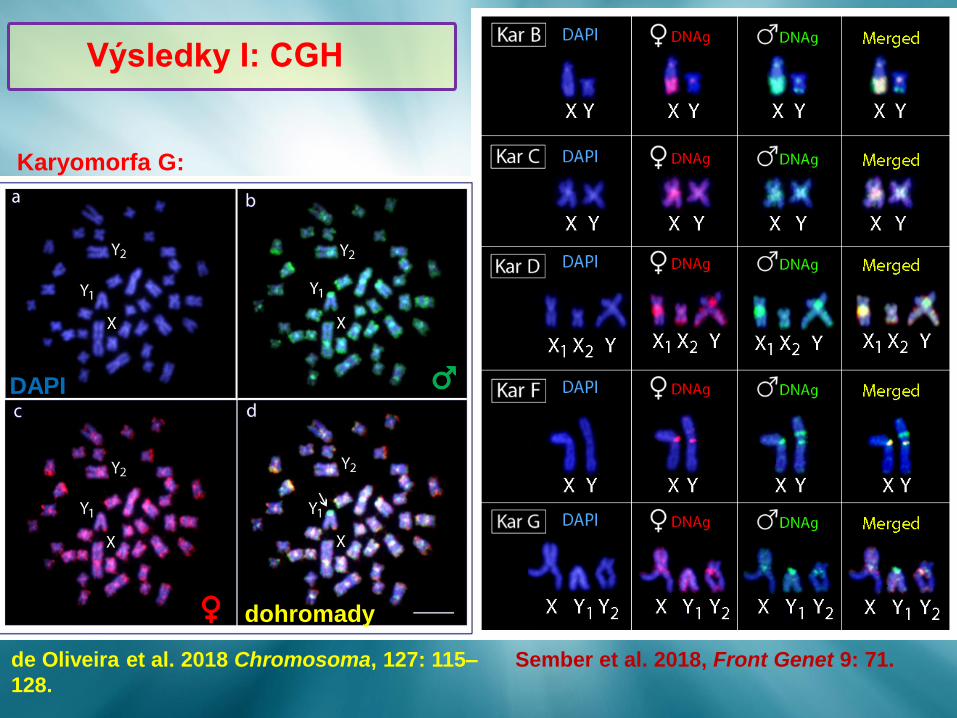
de Oliveira et al. 2018 Chromosoma, 127: 115–
128.
Sember et al. 2018, Front Genet 9: 71.
DAPI ♂
Karyomorfa G:
dohromady ♀
Výsledky I: CGH
Page 32
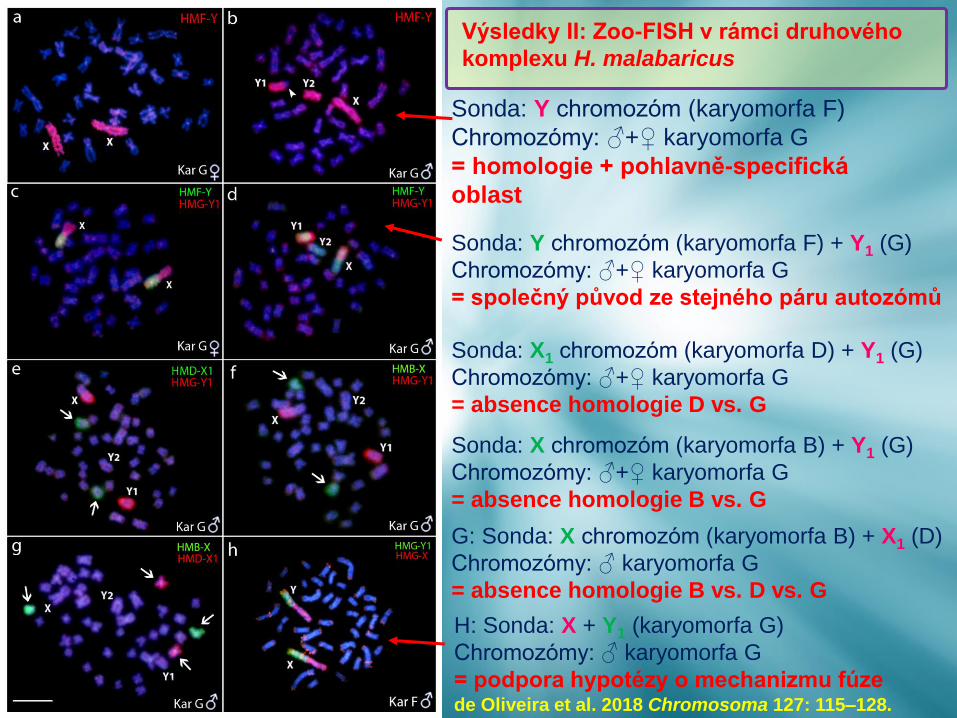
Sonda: Y chromozóm (karyomorfa F) + Y1 (G)
Chromozómy: ♂+♀ karyomorfa G
= společný původ ze stejného páru autozómů
Sonda: X chromozóm (karyomorfa B) + Y1 (G)
Chromozómy: ♂+♀ karyomorfa G
= absence homologie B vs. G
G: Sonda: X chromozóm (karyomorfa B) + X1 (D)
Chromozómy: ♂ karyomorfa G
= absence homologie B vs. D vs. G
H: Sonda: X + Y1 (karyomorfa G)
Chromozómy: ♂ karyomorfa G
= podpora hypotézy o mechanizmu fúze
Sonda: Y chromozóm (karyomorfa F)
Chromozómy: ♂+♀ karyomorfa G
= homologie + pohlavně-specifická
oblast
Sonda: X1 chromozóm (karyomorfa D) + Y1 (G)
Chromozómy: ♂+♀ karyomorfa G
= absence homologie D vs. G
Výsledky II: Zoo-FISH v rámci druhového
komplexu H. malabaricus
de Oliveira et al. 2018 Chromosoma 127: 115–128.
Page 33
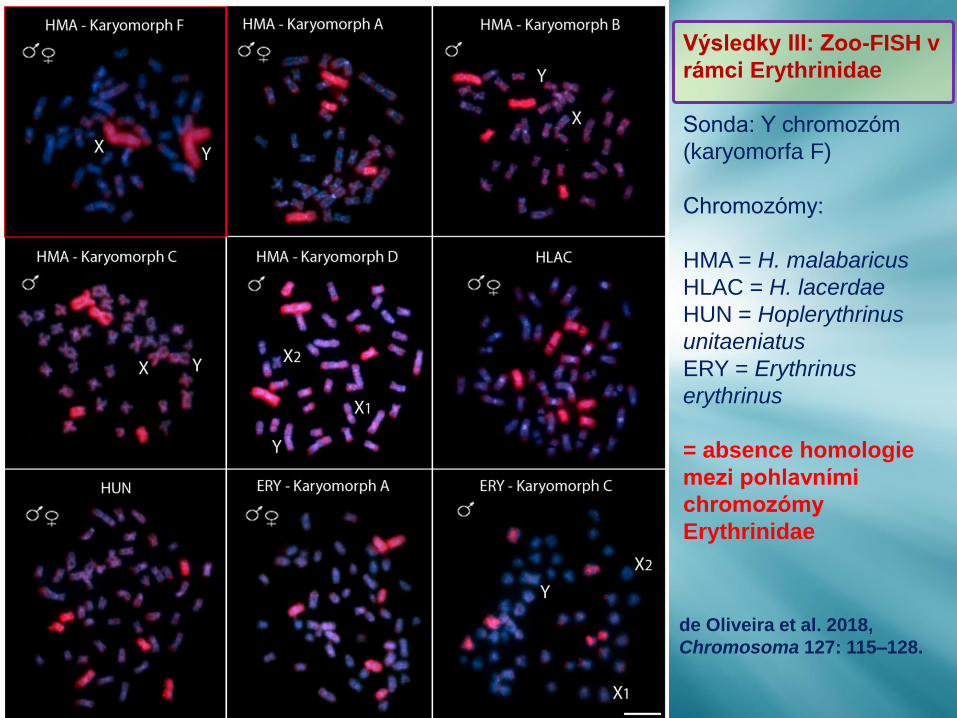
Výsledky III: Zoo-FISH v
rámci Erythrinidae
Sonda: Y chromozóm
(karyomorfa F)
Chromozómy:
HMA = H. malabaricus
HLAC = H. lacerdae
HUN = Hoplerythrinus
unitaeniatus
ERY = Erythrinus
erythrinus
= absence homologie
mezi pohlavními
chromozómy
Erythrinidae
de Oliveira et al. 2018,
Chromosoma 127: 115–128.
Page 34
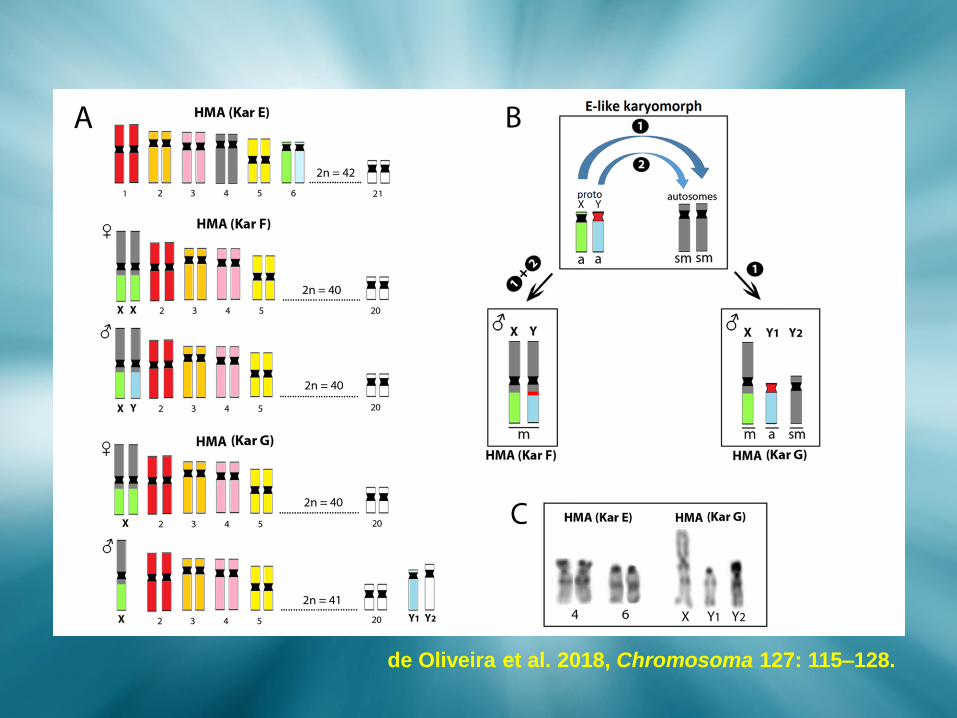
de Oliveira et al. 2018, Chromosoma 127: 115–128.
Page 35
Evoluční dynamika genomů nižších obratlovců
Hemiklonální
reprodukce u
vodních skokanů
druhového
komplexu
Pelophylax
Evoluce
pohlavních
chromozómů
neotropických
sladkovodních ryb
Karyotypové
změny a dynamika
repetitivních
sekvencí u
diploidních a
polyploidních
sekavcovitých ryb
(Cobitoidea)
Page 36
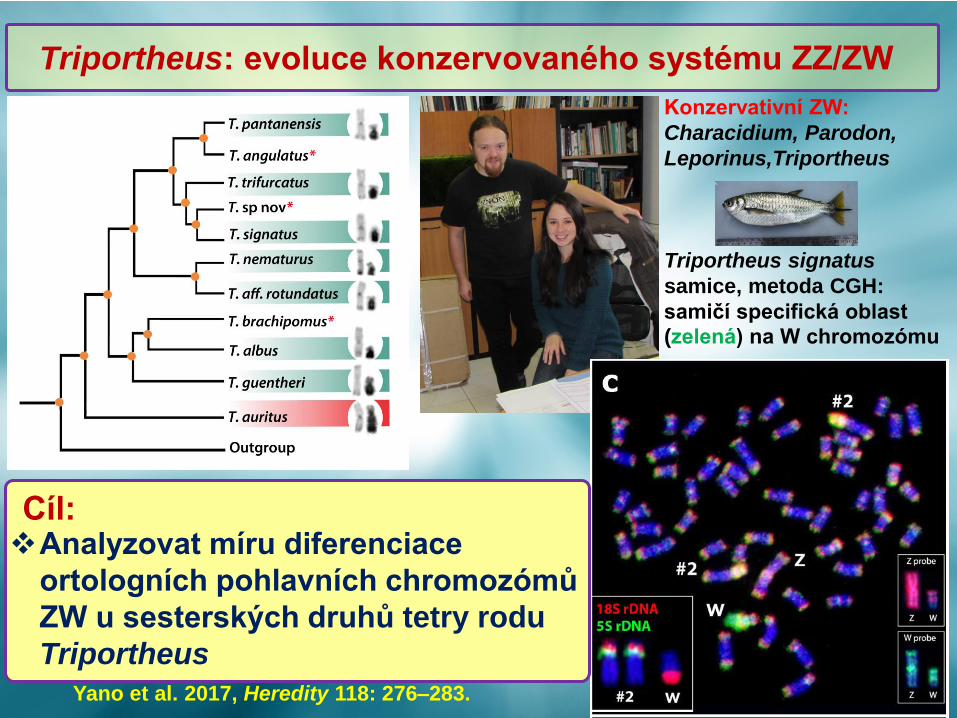
Konzervativní ZW:
Characidium, Parodon,
Leporinus,Triportheus
Triportheus signatus
samice, metoda CGH:
samičí specifická oblast
(zelená) na W chromozómu
Yano et al. 2017, Heredity 118: 276–283.
Triportheus: evoluce konzervovaného systému ZZ/ZW
Cíl:❖Analyzovat míru diferenciace
ortologních pohlavních chromozómů
ZW u sesterských druhů tetry rodu
Triportheus
Page 37
❖ Dynamika genomů v diverzifikované čeledi Nemacheilidae probíhá na sub-
chromozómové úrovni, nekoreluje s fylogenetickou hypotézou a je
výraznější u endemitů.
❖ Pokročilý stav re-diploidizace genomů sekavek čeledi Botiinae, odlišnoá
druhově-specifická dynamika repetitivních sekvencí a odlišný molekulární
drive multigenových rodin (U2 snDNA < 45S rDNA < 5S rDNA).
❖ Endemit Sinibotia zebra není výsledek mezidruhového křížení (morfologická
adaptace, mimikry).
❖ Mechanizmus hybridogeneze/genomové eliminace není uniformní u
různých populací skokana zeleného P. esculentus.
❖ Nezávislý původ a různá dynamika pohlavních chromozómů druhového
komplexu H. malabaricus (platí i ve vztahu k příbuzným druhům).
❖ Konvergence systémů XY (karyomorfa F) a XY1Y2 (kar. G) z karyotypu
společného předka (tandemová fúze)
❖ Podpora hypotézy pokročilé reproduktivní izolace a samostatných druhů
Závěry
❖ Výskyt asociovaných bloků rDNA/“teomere-like“ sekvencí zasluhuje větší
pozornost. (funkce?)
❖ Společný původ, avšak odlišná dynamika systému ZZ/ZW u tetry rodu
Triportheus; absence korelace s fylogenetickou hypotézou.
Page 38
❖ Dynamika genomů v diverzifikované čeledi Nemacheilidae probíhá na sub-
chromozómové úrovni, nekoreluje s fylogenetickou hypotézou a je
výraznější u endemitů.
❖ Pokročilý stav re-diploidizace genomů sekavek čeledi Botiinae, odlišnoá
druhově-specifická dynamika repetitivních sekvencí a odlišný molekulární
drive multigenových rodin (U2 snDNA < 45S rDNA < 5S rDNA).
❖ Endemit Sinibotia zebra není výsledek mezidruhového křížení (morfologická
adaptace, mimikry).
❖ Mechanizmus hybridogeneze/genomové eliminace není uniformní u
různých populací skokana zeleného P. esculentus.
❖ Nezávislý původ a různá dynamika pohlavních chromozómů druhového
komplexu H. malabaricus (platí i ve vztahu k příbuzným druhům).
❖ Konvergence systémů XY (karyomorfa F) a XY1Y2 (kar. G) z karyotypu
společného předka (tandemová fúze)
❖ Podpora hypotézy pokročilé reproduktivní izolace a samostatných druhů
Závěry
❖ Výskyt asociovaných bloků rDNA/“teomere-like“ sekvencí zasluhuje větší
pozornost. (funkce?)
❖ Společný původ, avšak odlišná dynamika systému ZZ/ZW u tetry rodu
Triportheus; absence korelace s fylogenetickou hypotézou.
Take home message:
❖Prezentované práce (8 publikací + 1
kapitola v knize) demonstrují značný
potenciál metod molekulární
cytogenetiky v současném výzkumu.
❖Metody jsou efektivní zejména pokud:❖ je zacíleno na vhodné
evoluční/ekologické/taxonomické
téma
❖ jsou data kombinována s
výsledky jiných biologických
disciplín
Page 39
Bohlen J Choleva L Janko K Rábová M
Šlechta V Šlechtová V Pelikánová ŠŠlechtová V
Pokorná M Symonová R Doležálková M Majtánová Z Šejnohová P
Altmanová M
Petr Ráb
Page 40
Granty:
GAČR, projekty č. 206/08/0637, 523/08/0824, P506/11/P596, 13-37277S,
14-02940S, GAUK 43-251468, RVO: 67985904, Conselho Nacional de
Desenvolvimento Científico e Tecnológico - CNPq (Proc. nos
304992/2015-1, 306896/2014-1 a 152105/2016-6), Fundação de Amparo
à Pesquisa do Estado de São Paulo- FAPESP (Proc. No 2016/21411-7)
EXCELLENCE CZ.02.1.01/0.0/0.0/15_003/0000460 OP RDE .
Thomas LiehrJena University Hospital, Friedrich
Schiller University, Institute of Human
Genetics, Jena, Germany
Vladimir TrifonovInstitute of Molecular and Cellular
Biology SB RAS, Novosibirsk,
Russia
Terumi Hatanaka, Geovanna Malimpensa,
Ezequiel Aguiar de Oliveira, Cassia Yano,
Felipe Barby, Marcelo Cioffi, Pedro
Carvalho, Renáta Moraes, Piau, Luiz
Antonio Carlos Bertollo, Natália Freitas
Orlando Moreira-Filho