Hemicanales formados por conexinas en la regulación del volumen del cardiomiocito expuesto a estrés hiposmótico Directores de Memoria: Dr. Juan Carlos Sáez. Facultad Ciencias Biológicas, P. Universidad Católica de Chile Dr. Sergio Lavandero. Facultad Ciencias Químicas y Farmacéuticas, Universidad de Chile. Profesor Patrocinante Dr. Sergio Lavandero Facultad de Ciencias Químicas y Farmacéuticas Universidad de Chile Memoria para optar al título de Bioquímico Daniela Paz Salas Castro Santiago, 2009 Departamento de Bioquímica y Biología Celular Facultad de Ciencias Químicas y Farmacéuticas Universidad de Chile
Transcript
Hemicanales formados por conexinas en la regulación del volumen del cardiomiocito
expuesto a estrés hiposmótico
Directores de Memoria: Dr. Juan Carlos Sáez. Facultad Ciencias Biológicas, P. Universidad Católica de Chile
Dr. Sergio Lavandero. Facultad Ciencias Químicas y Farmacéuticas, Universidad de
Chile.
Profesor Patrocinante Dr. Sergio Lavandero
Facultad de Ciencias Químicas y Farmacéuticas Universidad de Chile
Memoria para optar al título de Bioquímico
Daniela Paz Salas Castro Santiago, 2009
Departamento de Bioquímica y Biología Celular Facultad de Ciencias Químicas y Farmacéuticas Universidad de Chile
2
DEDICATORIA Dedico esta memoria y todos mis logros académicos a mis padres, Lorena y
Claudio, por inculcarme el valor de los estudios y el trabajo constante como herramientas
para desarrollarme profesionalmente. Sin su constante apoyo y cariño para superar los
momentos difíciles, nada de esto sería posible.
3
AGRADECIMIENTOS
A mi familia por su apoyo y cariño incondicional. Gracias tías, primos, hermano,
sobrinos y padres por alegrarse y disfrutar con mis triunfos y darme palabras de aliento y
confianza cuando las cosas se volvían difíciles. Son responsables en gran medida de
todos mis logros.
A mis directores de tesis, Dres Sergio Lavandero y Juan Carlos Sáez. Por
aceptarme en su laboratorio cuando recién comenzaba en mi vida científica y haber
confiado en mis capacidades y siempre alentarme y enseñarme que la honestidad y la
perseverancia son la clave del éxito. Por su paciencia y por darme la oportunidad de
desarrollar mi tesis con libertad. Gracias por hacer del Laboratorio un lugar tan agradable
para trabajar.
A mis compañeros de Laboratorio por su amistad, alegría y consejos. Muchas
gracias por compartir conmigo tan generosamente sus conocimientos y experiencias. Son
responsables del ambiente increíble que se vive en el Laboratorio. En especial, muchas
gracias Kurt y Jessi por su ayuda en los experimentos y por transmitirme sus
conocimientos.
A Fidel, Ruth y Gladys por facilitarme las cosas y contribuir a que siempre tuviera
todo lo necesario para trabajar.
A mis amigas Jessi, Andrea y Vale por su amistad y cariño. Por su apoyo y
ayuda. Son responsables en gran medida de la culminación de esta Tesis. Agradezco la
oportunidad de haberlas conocido.
Finalmente, agradezco a mis padres por todas las enseñanzas y valores que me
inculcaron. Por celebrar todos mis logros y siempre apoyarme en mis decisiones.
4
FINANCIAMIENTO
Esta memoria se realizó en el laboratorio de Transducción de Señales del centro
Fondap Estudios Moleculares de la Célula, Facultades de Ciencias Químicas y
Farmacéuticas y Medicina, Universidad de Chile y en el Departamento de Fisiología,
Facultad de Ciencias Biológicas, Pontificia Universidad Católica de Chile. Esta memoria
se financió parcialmente por los proyectos Fondap 15010006 (SL) y Fondecyt 1070591
(JCS)
5
INDICE GENERAL Página
INDICE GENERAL ............................................................................................. 5
miso : pendiente de captación obtenida con medio isosmótico
mhipo : pendiente de captación obtenida con medio hiposmótico
mLa+3 : pendiente de captación obtenida al agregar LaCl3
MgCl2 : cloruro de magnesio
MgSO4 : sulfato de magnesio
min : minuto
mL : mililitro
mm : milímetro
mOsm : miliosmolar
mRNA : ácido ribonucleico mensajero
Na+ : ion sodio
Na2HPO4 : fosfato ácido disódico
NaCl : cloruro de sodio
NaF : fluoruro de sodio
NAD+ : Nicotinamida adenina dinucleótido
nm : nanómetros
NP-40 : nonidet p40
p38-MAPK : proteina quinasa de 38 kDa activada por mitógenos
PKC : proteina quinasa C
PMSF : p-fenilmetilsulfonilfluoruro
PVDF : polivinilidenfluoruro
10
rpm : revoluciones por minuto
ROI : region de interés
RVI : aumento regulado de volumen
RVD : reducción regulada de volume
SDS : dodecilsulfato de sodio
seg : segundos
SEM : error estándar de la media
TBS : amortiguador tris salino
TonEBP : proteína de unión al elemento de respuesta a tonicidad
V : voltios
Wt : tipo silvestre
11
RESUMEN
El correcto funcionamiento celular requiere condiciones físicas específicas, como
pH, temperatura y fuerza iónica. Cualquier alteración de estos parámetros puede producir
consecuencias graves para las células. La mantención del volumen celular es otro de los
parámetros importantes para la célula ya que regula la fuerza iónica y las concentraciones
de osmolitos y segundos mensajeros intracelulares. En patologías como la diabetes o
isquémicas como el infarto agudo al miocardio, ocurren alteraciones del volumen celular.
Durante la isquemia disminuye el aporte de nutrientes y oxígeno a las células por
lo que aumenta el catabolismo de nutrientes para obtener energía. Como consecuencia,
aumenta la osmolaridad intracelular y con ello, la entrada de agua a la célula
produciéndose un aumento de su volumen. Pero las células han desarrollado mecanismos
para regular su volumen y volver a la normalidad frente a cambios en la osmolaridad del
medio. En el caso del estrés hiposmótico, la estrategia consiste en sacar iones de la
célula, lo que disminuye su contenido de agua. Se ha descrito que los cardiomiocitos no
regulan espontáneamente su volumen en condiciones de estrés hiposmótico, lo que se ha
asociado a muerte. Este hecho es importante si consideramos que el corazón es uno de
los órganos más afectados por enfermedades que incluyen episodios isquémicos.
Las conexinas son proteínas de transmembrana que forman hexámeros
(hemicanal) y se insertan en la membrana plasmática de las células. Si dos hemicanales
de células adyacentes se unen forman un canal de una unión en hendidura, y permiten la
comunicación de los citoplasmas de las células vecinas. Se ha propuesto que los
hemicanales formados por la conexina 43 (Hcs-Cx43) podrían participar en la regulación
de volumen de las células, ya que forman verdaderos poros en la superficie celular que
permite el paso de agua e iones por difusión simple.
El objetivo de esta tesis consistió en determinar si los hemicanales formados por
conexinas participan en la regulación de volumen del cardiomiocito expuesto a estrés
hiposmótico. Para este fin cultivos primarios de cardiomiocitos de ratas neonatas se
expusieron a estrés hiposmótico y se estudió:
12
• Si el estrés hiposmótico modifica el estado funcional de los Hcs-Cx43 a través de
la técnica de captación de etidio
• Si los cambios en el estado funcional de los Hcs-Cx obedecen a modificaciones en
la cantidad de los Hcs-Cx43 expuestos en la membrana o cambios en su estado
de fosforilación, mediante la técnica de biotinilación de proteínas de superficie
• Si los Hcs-Cx43 participan en la regulación de volumen del cardiomiocito expuesto
a estrés hiposmótico, mediante el uso de calceina-AM y microscopía confocal
como indicador de los cambios de volumen de la célula e interviniendo el sistema
con el inhibidor específico de Hcs-Cx43, Gap26.
Los resultados muestran que los Hcs-Cx aumentan su estado funcional al exponer
las células a estrés hiposmótico, lo que impide la regulación de volumen del cardiomiocito,
ya que al inhibirlos con Gap26 recuperan su volumen. Además se sugiere que el aumento
funcional de los Hcs-Cx no se podría explicar por cambios en el estado de fosforilación o
alteraciones de la cantidad de Hcs-Cx expuestos en la membrana celular. De estos
resultados se concluye que los hemicanales formados por conexinas participan en el
control del volumen del cardiomiocito
13
SUMMARY
Role of connexin hemichannels in the regulation of cardiac myocyte volume by hyposmotic stress
The cell homeostasis requires specific physical conditions such as pH, temperature
and ionic strength. Any alteration in these parameters may produce serious consequences
to the cell. The maintenance of cell volume is key parameter because is involved in the
regulation of ionic strength, and concentration of osmolyte and intracellular second
messengers. Alterations in cell volume have been described in pathologies such as
diabetes, stroke and acute myocardial infarction.
During ischemia the nutrients and oxygen availability to the cells diminishes,
resulting in an increased catabolism in order to obtain energy. As a consequence,
intracellular osmolarity increases leading to water influx into the cell and an increase in cell
volume. Cells have developed different compensatory mechanisms to restore their volume
when they are exposed to changes in external osmolarity. In the case of hyposmotic
stress, ions are pumped out the cell to diminish water content. It has been shown that
cardiac myocytes are unable to spontaneously regulate their volume when exposed to
osmotic stress, and this event has been associated with increased cell death susceptibility.
This is important if we consider that cardiac tissue is one of the most affected organs by
ischemic diseases.
Connexins are transmembrane proteins forming hexamers (hemichannels) at the
cell membrane. When two hemichannels from adjacent cells reach each other, they form a
gap junction channel, which allow communication of both cytoplasms. It has been
proposed that hemichannels formed by connexin 43 (Hcs-Cx43) may participate in cell
volume regulation because they form pores in the cell surface allowing the passage of
water and ions by simple diffusion.
The aim of this work was to evaluate whether Hcs-Cx43 participates in the volume
regulation of cardiac myocytes exposed to hyposmotic stress. To this end, cultured
neonatal rat cardiac myocytes were exposed to hyposmotic stress and we study whether:
14
• Hyposmotic stress modifies the functional state of Hcs-Cx43 assessing the
ethidium uptake by the cells
• Changes in Hcs-Cx functional state are explained by the number of Hcs-Cx43
present in the cell membrane or by changes in their phosphorylation status.
• Hcs-Cx43 participates in the volume regulation of cardiac myocyte exposed to
hyposmotic stress. This was evaluated using calcein-AM and confocal microscopy
to measure changes in cell volume and Gap26 to inhibit Hcs-Cx43.
The results showed that the functional state of Hcs-Cx is enhanced in cells
exposed to hyposmotic stress. Such increase in the functional state of Hcs-Cx could not
be explained by changes in the phosphorylation state or alterations in the amount of Hcs-
Cx exposed in the cell surface. The increase in cardiac myocyte volume induced by
hyposmostic stress was inhibited by Gap26. These results collectively show that connexin
hemichannels participates in the regulation of cardiac myocyte volume.
15
1. INTRODUCCIÓN
1.1. Mantención del volumen celular
El mantenimiento del volumen celular es fundamental para el correcto
funcionamiento de las células. Pequeñas perturbaciones de la osmolaridad extracelular
producen cambios en el contenido de agua de la célula alterando la concentración de
mensajeros celulares, el equilibrio iónico y la estructura celular, lo que puede afectar
negativamente el funcionamiento de la célula (Ho, 2006). Bajo condiciones fisiológicas el
medio extracelular se mantiene relativamente constante. Sin embargo el medio
intracelular cambia continuamente debido a la generación de gradientes osmóticos
transitorios asociados a la incorporación de nutrientes, exocitosis de moléculas,
metabolismo y remodelamiento del citoesqueleto, por lo que la célula constantemente
debe poner en marcha mecanismos que permitan regular su volumen (Lang et al., 1998).
1.1.1. Vías de ingreso del agua a las células
Las membranas celulares son altamente permeables al agua, pudiendo entrar en
forma pasiva a través de la membrana plasmática, por canales de proteínas llamadas
aquaporinas o solvatando iones que ingresan a la célula a través de canales o
transportadores selectivos (Wright y Rees, 1998).
A pesar de la naturaleza lipídica de la membrana plasmática, las moléculas de agua
pueden difundir a través de ella e incluso hasta el descubrimiento de las aquaporinas, se
pensaba que era el principal mecanismo de entrada de agua a las células (Wright y Rees,
1998). Estudios en bicapas lipídicas artificiales mostraron una permeabilidad al agua
superior a 50x10-6 ms-1 en ausencia de colesterol y de 10-20x10-6 ms-1 con la incorporación
de concentraciones fisiológicas de colesterol (Fettiplace y Haydon, 1980). Pero estos
valores no permiten dar cuenta de cambios rápidos de volumen que ocurren en la célula
cuando está expuesta por ejemplo a estrés osmótico, para lo cual existirían las
aquaporinas.
16
Las aquaporinas (AQP) son proteínas integrales de membrana que forman poros
en la superficie de las células, permitiendo el paso de agua y algunas sales a favor de su
gradiente de concentración. Estas proteínas son especialmente importantes en las células
renales, donde cumplen un papel fundamental en la reabsorción de agua (Verkman y
Mitra, 2000). Su papel en células cardiacas es controversial por lo que no se puede saber
con certeza cuál es su función (Egan et al., 2006), aunque hay trabajos que sugieren que
podrían participar en la regulación de volumen, ya que se ha observado aumentos del
mRNA y expresión de AQP1 en condiciones de estrés hiposmótico (Jonker et al., 2003).
El tercer mecanismo que posibilita movimientos de agua entre los medio intra y
extracelulares son los iones. La célula tiene un cierto balance iónico que si se rompe
activa transportadores que movilizan los iones y, con ello, agua para solvatarlos (Wright y
Rees, 1998). Debido a que los iones contribuyen significativamente a la osmolaridad tanto
intra como extracelular, se estima que son responsables de aproximadamente 2/3 de la
regulación de volumen de la célula (Fisher y Spring, 1984).
1.2. Mecanismos de regulación del volumen celular
Cuando una célula se ve expuesta a cambios en la osmolaridad, se activan
mecanismos que contrarrestan las modificaciones en el volumen celular, alterando
principalmente la cantidad de osmolitos al interior de la célula y con ello, agua (Lang et al.,
1998).
1.2.1. Aumento regulado de volumen o RVI
Si el medio extracelular tiene una mayor concentración de osmolitos que el
intracelular, la célula se enfrenta a un estímulo hiperosmótico y pierde agua, por lo que se
encoge y se activa una respuesta que se conoce como “Aumento Regulado del Volumen”
o RVI, por su sigla en inglés (Regulated Volume Increase) que tiene un componente a
temprano o corto plazo y otro tardía o de largo plazo (Wright y Rees, 1998). En el primero
se activan mecanismos que posibilitan la entrada de iones como K+, Cl- y Na+ a través de
intercambiadores preexistentes de Na+-K+-2Cl- (Dunham et al., 1990), Na+/H+ (Grinstein et
17
al., 1983) y Cl-/HCO3- (Hebert, 1986) para que aumente la concentración de osmolitos en
la célula e ingrese agua para recuperar el volumen. Como consecuencia se producen
cambios en el pH intracelular y aumenta la fuerza iónica, afectando a largo plazo
principalmente a las proteínas (Lang et al., 1998). Si se trata de un estímulo sostenido en
el tiempo, se pone en marcha la respuesta adaptativa, que consiste en cambiar los iones
por osmolitos orgánicos como sorbitol, taurina y betaína, que son moléculas
osmóticamente activas pero que carecen de carga, con lo que se consigue mantener el
agua al interior de la célula sin cambiar la fuerza iónica (Ho, 2006). Con este fin se activan
genes para aumentar la producción o el transporte de estos osmolitos orgánicos, en su
mayoría bajo el control del factor de transcripción de respuesta a estrés osmótico TonEBP
(Handler y Kwon, 2001).
1.2.2. Disminución regulada de volumen o RVD
En el caso del estrés hiposmótico, la célula aumenta de volumen debido a que
existe una mayor concentración de osmolitos al interior de la célula respecto al medio
extracelular, por lo que ingresa agua a favor de su gradiente de concentración. En
condiciones fisiológicas esto ocurre durante el metabolismo, en que aumenta el número
de moléculas en el medio intracelular y se produce un pequeño gradiente osmótico
(Pasantes-Morales et al., 2006). Para contrarrestar este evento se activa la respuesta de
“Disminución Regulada del Volumen” o RVD por su sigla en inglés (Regulated Volume
Decrease), que consiste en eliminar K+ y Cl- de la célula para expulsar agua y recuperar el
volumen. Para ello se activan en forma paralela transportadores o co-transportadores de
K+ y Cl-, dependiendo del tipo celular, y de osmolitos como sorbitol e inositol, para
eliminarlos de la célula, además de activarse la degradación de osmolitos en algunos
tipos celulares (Lang et al., 1998). El objetivo de la activación de estos mecanismos es
conseguir la reducción de la concentración de osmolitos intracelulares para que se
produzca reducción del volumen de la célula.
18
1.3. Transducción de señales durante la regulación de volumen
Aún no está claro si en los cambios de volumen de la célula y la activación de las
vías de señalización involucradas participa un tipo de molécula o una variedad de ellas
que interactúan para detectar dichos cambios. Se ha propuesto la participación de
diversas moléculas como integrinas (vom Dahl et al., 2003), receptores de factores de
crecimiento (Franco et al., 2004), proteínas G (Davis et al., 1992), canales sensibles a
estiramiento (Sackin, 1995) y cambios en el citoesqueleto (Pedersen et al., 2001) como
sensores de los cambios de volumen, ya que se ha demostrado su activación durante
distintos estímulos osmóticos (Pasantes-Morales et al., 2006; Wright y Rees, 1998).
Se ha demostrado que el aumento en la concentración de Ca+2 intracelular es
fundamental para el RVD de múltiples tipos celulares, mientras que cambios en la
concentración de cAMP, pareciera ser fundamental para el RVI, relacionados ambos con
el funcionamiento de canales iónicos (McCarty y O’Neil, 1992). El Ca+2 también se ha
vinculado al funcionamiento de proteínas quinasas, como PKC, las que podrían activar
canales iónicos a través de su fosforilación durante la RVI (Grinstein et al., 1986). Por otro
lado, se ha sugerido que durante la RVD la desfosforilación sería el evento predominante
ya que se correlaciona con la salida de K+ y Cl- de la célula (Jennings y Schulz, 1991).
Son múltiples las proteínas que se activan río debajo de las respuestas de RVI y
RVD, encontrándose diferencias según el tipo celular de que se trate. A pesar de que
existe consenso en algunos mecanismos que operan en las células, hasta la fecha no se
ha podido encontrar un mecanismo universal que de cuenta de la regulación de volumen
(Lang et al., 1998).
Los cambios de volumen producto tanto del estrés hiposmótico e hiperosmótico
producen graves consecuencias en la célula debido a que se altera la concentración de
moléculas intracelulares como iones y segundos mensajeros que pueden derivar en una
alteración de la homeostasis celular e incluso la muerte. Dentro de las condiciones
patológicas que producen cambios de volumen en la célula, la isquemia es una de las
más importantes debido al daño que produce y por la alta frecuencia con que se presenta
en la población.
19
1.4. Isquemia cardiaca y regulación de volumen
Las enfermedades cardiovasculares constituyen una de las principales causas de
muerte en el mundo (Kannel, 2000), por lo que el estudio de los mecanismos moleculares
implicados en estas patologías, resulta de vital importancia. Dentro de ellas, las de tipo
isquémico, como infarto agudo al miocardio (IAM), son las más prevalentes.
La isquemia se caracteriza por la interrupción del flujo sanguíneo, disminuyendo el
aporte de oxígeno (hipoxia) y de nutrientes a las células (Cokkinos, 2006) y el lavado de
productos metabólicos o de desecho celular. Una de las consecuencias de la isquemia es
el aumento de la osmolaridad intracelular, producto del catabolismo de los nutrientes
almacenados (ej. ATP se convierte en ADP y fósforo inorgánico) y del aumento en la
producción de lactato debido a que la falta de suministro de oxígeno produce un cambio
de metabolismo de oxidativo a glicolítico (Reimer et al., 1981). Debido al aumento en la
cantidad de osmolitos intracelulares, la célula experimenta estrés hiposmótico, por lo que
el agua ingresa a la célula siendo impulsada por el gradiente de concentración de los
solutos y se produce un aumento en el volumen de la célula, produciendo consecuencias
deletéreas como acortamiento del potencial de acción en el caso de cardiomiocitos
(Vandenberg et al., 1996).
En un experimento realizado por Tranum-Jensen et al (1981), se determinó los
cambios en la osmolaridad en corazones de cerdo aislados sometidos a isquemia regional
por oclusión (ver Figura 1). Se observó que luego de 50 min de isquemia, la osmolaridad
promedio de la célula aumenta en aproximadamente 40 mOsM, lo que se correlaciona
con un discreto aumento del volumen celular de aproximadamente 16%. El hecho de que
la célula prácticamente no aumente de volumen durante la isquemia a pesar del aumento
en la osmolaridad intracelular, es bien documentado en la literatura (Vandenberg et al.,
1996) y se puede atribuir principalmente a la rápida salida de moléculas permeables a la
membrana celular y al rápido transporte del lactato fuera de la célula a través de
transportadores específicos (Poole y Halestrap, 1993), por lo que la osmolaridad
intracelular se equilibraría rápidamente con la extracelular, aliviando el estrés osmótico.
20
Al reestablecerse el flujo sanguíneo ingresa sangre normotónica al sitio isquémico,
desplazando el medio extracelular circundante ligeramente hipertónico. Debido a que
estas células presentan alta osmolaridad, el gradiente osmolar generado en estas
condiciones es mayor al que ocurre durante la isquemia, lo que induce un mayor aumento
del volumen celular (Jennings et al., 1985), ya que la célula no alcanza a equilibrar su
osmolaridad con la del medio extracelular. A este segundo aumento de volumen se ha
asociado la mayor parte del daño celular, correlacionándose con arritmias y en general,
un mayor grado de disfunción cardiaca (Tranum-Jensen et al., 1981). Esto se sustenta en
la evidencia de que al reperfundir con medio ligeramente hipertónico (con una
osmolaridad similar a la que se alcanza durante la isquemia), los parámetros cardiacos
mejoran notablemente (García-Dorado y Oliveras, 1993), revelando la importancia de la
osmolaridad en la regulación de volumen en la patología isquémica.
Figura 1. Aumento de volumen en cardiomiocitos expuestos a isquemia y reperfusión. (a) Se esquematiza cómo la isquemia afecta la osmolaridad intracelular y el rápido equilibrio que existiría con el medio extracelular al no existir flujo de sangre, por lo que no se producen cambios importantes de volumen. (b) Cuando ocurre la reperfusión con sangre normotónica, las células, cuya osmolaridad interior está aumentada, experimentan un brusco estrés hiposmótico al enfrentarse a la osmolaridad de la sangre. Como no pueden equilibrar su osmolaridad con el medio extracelular debido a que éste se recambia continuamente, ocurre un brusco aumento de volumen. Adaptado de Vandenberg et al., 1996.
↑ PCr
↑ Mg-ATP
↑ trigliceridos
↑glicógeno
← ↓ Cr + Pi
← ↓ (ADP,AMP) + ↓ Mg2++ ↓ Pi
← ↓ acetil-CoA
← ↓ ↓ lactato + ↓ H+
360 mOsm Kg-1
CapilarSangre Normotónica
290 mOsm Kg-1
(b) Reperfusión
↓ PCr
↓Mg-ATP
↓triglicéridos
↓glicógeno
→ ↑ Cr + Pi
→ ↑ (ADP,AMP) + ↑ Mg2++ ↑ Pi
→ ↑ acetil-CoA
→ ↑ ↑ lactato + ↑H+
360 mOsm Kg-1
Capilar
↑ ↑ lactato + ↑H+↑K+
360 mOsm Kg-1
(a) Isquemia
H2O
↑ PCr
↑ Mg-ATP
↑ trigliceridos
↑glicógeno
← ↓ Cr + Pi
← ↓ (ADP,AMP) + ↓ Mg2++ ↓ Pi
← ↓ acetil-CoA
← ↓ ↓ lactato + ↓ H+
360 mOsm Kg-1
CapilarSangre Normotónica
290 mOsm Kg-1
(b) Reperfusión
↓ PCr
↓Mg-ATP
↓triglicéridos
↓glicógeno
→ ↑ Cr + Pi
→ ↑ (ADP,AMP) + ↑ Mg2++ ↑ Pi
→ ↑ acetil-CoA
→ ↑ ↑ lactato + ↑H+
360 mOsm Kg-1
Capilar
↑ ↑ lactato + ↑H+↑K+
360 mOsm Kg-1
(a) Isquemia
H2O
21
1.5. Cardiomiocitos y regulación de volumen
Existe controversia respecto de si las células cardiacas son capaces o no de
regular su volumen en forma espontánea cuando se enfrentan a un medio anisosmótico.
Distintos investigadores señalan que los cardiomiocitos no ponen en marcha los
mecanismos de RVI o RVD (Drewnowska y Baumgarten, 1991; Wang et al., 1997), pero
en algunos tipos celulares como cardiomiocitos de pollo presentan RVD (Zhang et al.,
1993). En nuestro Laboratorio hemos demostrado que en cardiomiocitos de rata neonata
no existe RVD espontáneo, lo que sería inhibido por las especies reactivas del oxígeno
(EROs), que se producen durante el estrés hiposmótico (Diaz-Elizondo et al., 2006).
Aún no se conocen en detalle los mecanismos que gobiernan el RVD. Se ha
descrito un sinnúmero de moléculas y proteínas que podrían participar en este proceso,
siendo los hemicanales formados por la conexina 43 (Hcs-Cx43) uno de los últimos
candidatos atractivos para participar en regulación de volumen debido a su capacidad
para formar verdaderos poros en las células, dejando pasar agua e iones (Schulz y
Heusch, 2004).
1.6. Conexinas
La familia de las conexinas está formada por 20 integrantes en humano, siendo la
conexina-43 (Cx43) la más ubicua (Laird, 2006). El corazón es especialmente rico en
conexinas, y expresa las conexinas 40, 45 y 43 (John et al., 2003). Las conexinas (Cxs)
son proteínas que participan en la formación de uniones en hendidura (o gap junctions),
que comunican el citoplasma de dos células adyacentes.
Luego de la síntesis, las conexinas oligomerizan en el retículo endoplásmico para
formar hexámeros (hemicanal o conexón), los cuales son posteriormente exportados
hacia la membrana plasmática para insertarse en ella. Una vez que un hemicanal se
inserta en la membrana, difunde hasta encontrar el conexón de una célula vecina, dando
origen a la unión en hendidura y con ello la comunicación entre las células (ver Figura 2).
Los canales intercelulares se agregan formando diferenciaciones de la membrana
denominadas placas de unión (Laird, 2006). En el corazón, tales canales cumplen un rol
22
fundamental en la transmisión de señales eléctricas, permitiendo la propagación de ondas
de calcio que coordinan la contracción (Jalife et al., 1999).
Durante mucho tiempo se pensó que los hemicanales sólo eran el paso previo a la
formación de uniones en hendidura, pero en los últimos años se ha demostrado que los
hemicanales pueden permanecer en la membrana citoplasmática sin participar en la unión
intercelular (Goodenough y Paul, 2003) y formar poros poco selectivos que permiten el
paso de iones y moléculas de hasta ~1 kDa, en función de su gradiente de concentración,
con el consiguiente movimiento de agua (John et al., 2003). Se ha demostrado que a
través de estos hemicanales pueden pasar moléculas como ATP, glutamato,
prostaglandina E2 y NAD+, que actúan como mensajeros paracrinos (Sáez et al., 2003;
Jiang y Cherian, 2003). Debido a su baja selectividad y gran diámetro de poro, en células
en reposo los hemicanales presentan baja probabilidad de apertura pero ésta aumenta
con la desfosforilación (Hcs-Cx43), baja concentración de Ca2+ externo, estímulos
mecánicos, potencial de membrana positivos y disminución del potencial redox de la
célula (Hcs-Cx43) (Sáez et al., 2005). También, estímulos patológicos, como la inhibición
metabólica e isquemia/reperfusión aumentan la permeabilidad de la membrana celular a
través de los hemicanales (John et al., 2003).
23
Figura 2. Estructura de los hemicanales y las uniones en hendidura formadas por conexinas. La conexina presenta 4 dominios transmembrana y extremos NH2 y COOH-terminal orientados hacia el interior de la célula. Luego de oligomerizar en el retículo endoplasmático, se inserta en la membrana plasmática de la célula formando un hemicanal o conexón. Cuando un hemicanal se une a otro de una célula adyacente, se forma un canal de unión en hendidura que permite la comunicación física de los citoplasmas de dichas células. NT= NH2 terminal, CL= Loop citoplasmático, EL= Loop extracelular, CT= COOH-terminal, TM= Transmembrana. Adaptado de Decrock et al., 2009.
Recientemente se descubrieron las panexinas, que a diferencia de las conexinas
que se encuentran sólo en vertebrados, también están presentes en invertebrados
(Panchin, 2005). Aunque aún no está muy claro su rol, se sabe que pueden formar
uniones en hendidura, formar hemicanales y responden a estímulos similares a los de
conexinas (Barbe et al., 2006). Además, varios de los inhibidores que se usaban para
inhibir canales formados por conexinas, se ha descrito que también inhiben canales
formados por panexinas, lo que hace poner en duda el verdadero rol de las conexinas
(Spray et al., 2006).
Hemicanal / Conexón
Unión en Hendidura
Conexina
Intracelular (célula 2)
Extracelular
Intracelular (célula 1)
NT CL CT
EL
TM
24
1.7. Papel de las conexinas en el precondicionamiento isquémico e isquemia/reperfusión
Durante la isquemia aumenta la desfosforilación de la Cx43 (produce apertura), lo
que se asocia a daño cardiaco (Beardslee et al., 2000). El mecanismo propuesto para
esto es que al estar abiertos los canales formados por conexinas habría traspaso de
señales de muerte entre las células dañadas y sus vecinas, lo que sería responsable del
daño. Al someter los corazones a ciclos de hipoxia/normoxia subletales (proceso conocido
como precondicionamiento), aumenta la colocalización de Cx43 con p38-MAPK y la PKC,
manteniendo fosforilada la Cx43 en el miocardio preacondicionado (Schulz et al., 2003),
reduciendo el daño. El rol fundamental de la Cx43 queda en evidencia al observar que
ratones deficientes para esta proteína no experimentan precondicionamiento (Schwanke
et al., 2003). Recientemente se descubrió que Cx43 está presente en mitocondria
(Rodriguez-Sinovas et al., 2006) y podrían cumplir un rol importante en el
precondicionamiento (Ruiz-Meana et al., 2008).
1.8. Conexinas y regulación del volumen
Cuando se descubrió que las conexinas podían existir como parte de un hemicanal
sin necesidad de formar una unión en hendidura, se postuló que podían participar en la
regulación de volumen celular (Schulz et al., 2007). Sin embargo, son escasos los
reportes que apoyan esta hipótesis.
Una de las observaciones que apoya la tesis de la participación de las conexinas
en la regulación de volumen es que células carentes de conexinas no regulan su volumen
en respuesta a cambios en la concentración extracelular de Ca2+ (estímulo clásico que
produce su apertura), a menos que sean transfectadas con Cx43 (Quist et al., 2000),
sugiriendo que los hemicanales formados por Cx43 participan en la regulación de
volumen. Por otro lado, en células expuestas a estrés hiperosmótico aumentan los niveles
de la Cx43 fosforilada y su translocación a la membrana plasmática (Leykauf et al., 2003).
Además, el estrés hiperosmótico induce la apertura de hemicanales formados por
conexina43, lo que se asocia a desfosforilación de la Cx43 mediada por la activación de
25
proteínas fosfatasas (John et al., 2003). Otro antecedente que vincula a las conexinas con
la regulación de volumen es la observación de que el estrés hiposmótico disminuye la
conductancia de las uniones en hendidura (Ngezahayo y Kolb, 1990).
En base a los antecedentes expuestos se plantean las siguientes interrogantes:
¿Existe alguna relación entre la presencia y estado funcional de hemicanales
formados por conexinas y la ausencia de regulación espontánea de volumen del
cardiomiocito?
¿Modifica el estrés hiposmótico el estado funcional de los hemicanales formados por
conexinas? ¿Cuáles serían las consecuencias biológicas de dicho cambio?
2. HIPOTESIS
“El estrés hiposmótico aumenta la permeabilidad celular mediada por hemicanales
formados por conexinas en el cardiomiocito, impidiendo la regulación de su volumen”
3. OBJETIVO GENERAL
Demostrar que el estrés hiposmótico aumenta la permeabilidad de la membrana
celular mediada por hemicanales de conexina al exponer las células a medio hiposmótico,
impidiendo la regulación de su volumen.
26
4. OBJETIVOS ESPECÍFICOS
4.1. Determinar si el estrés hiposmótico estimula la apertura de los hemicanales.
4.2. Determinación del cambio en el número y/o estado de fosforilación de hemicanales
presentes en la superficie celular frente a estrés hiposmótico.
4.3. Estudiar la participación de hemicanales en la regulación del volumen en
cardiomiocitos expuestos a estrés hiposmótico.
27
5. MATERIALES Y MÉTODOS
5.1. Reactivos
Los siguientes reactivos se adquirieron en Sigma Chemical Co (St Louis, MO,
EEUU): medio DME, medio M-199, medio DMEM-F12 (sin rojo fenol ni bicarbonato de
sodio) Hanks, gelatina, pancreatina, Tritón X-100, 5-bromo-2’-deoxiuridina y LaCl3. En
Molecular Probes (Eugenia, Oregon EEUU) se adquirieron calceína-acetoximetil ester y
ácido plurónico. En Gibco BRL (Carlsbad, California EEUU) se adquirieron colagenasa II,
suero fetal de bovino, suero de caballo y bromuro de etidio. Los compuestos inorgánicos y
orgánicos, sales, ácidos y solventes se adquirieron en MERCK (Darmstadt, Alemania).
Los reactivos: Ez-link NHS-SS-Biotin y Immobilized Neutravidin Protein, se compraron en
Pierce (Rockford, IL). Los péptidos inhibidores Gap26 (VCYDKSFPISHVR) y Gap26
scrambled (PSFDSRHCIVKYV) se adquirieron a BIO-SYNTHESIS, INC (Lewisville,
Texas). El marcador preteñido de proteínas se adquirió en Winkler LTDA. El reactivo
quimioluminiscente para Western blot (Western Lightning) se obtuvo de PerKinElmer Life
Sciences, Inc. El material de plástico estéril para la obtención y cultivo de cardiomiocitos
neonatos se obtuvo en Falcon. Los anticuerpos secundarios anti-IgG de conejo y ratón
conjugados a peroxidasa se obtuvieron en Calbiochem (La jolla, CA, EEUU). El anticuerpo
para conexina 43 total (no comercial)l fue facilitado por el Dr. Juan Carlos Sáez, Pontificia
Universidad Católica de Chile.
5.2. Modelo animal
Se utilizó como modelo de estudio ratas neonatas Sprague-Dawley de 2 a 3 días
de edad, provenientes del bioterio de la Facultad de Ciencias Químicas y Farmacéuticas,
Universidad de Chile, cumplimiento de todas las normas éticas referidas a la utilización de
animales y autorización del Comité de Bioética.
28
5.3. Aislamiento y cultivo de cardiomiocitos ventriculares de ratas neonatas
Para el aislamiento y cultivo de los cardiomiocitos, los corazones se removieron y
lavaron a 37°C en medio Hanks estéril. Posteriormente se retiraron las aurículas y el tejido
ventricular resultante se homogenizó finamente para luego ser digerido enzimáticamente
en presencia de colagenasa tipo II y pancreatina. Para obtener una fracción enriquecida
en cardiomiocitos se aprovechó la capacidad diferencial de los distintos tipos celulares de
adherirse a superficies plásticas y el producto de la digestión se preplaqueó en botellas de
cultivo de 250 mL durante 2 h en medio DME:M199 (4:1) 10% FBS. Luego, las células no
adheridas presentes en el sobrenadante se recolectaron, centrifugaron a 1.000 RPM
durante 5 min y se resuspendieron en 21 mL de medio DME:M199 (4:1) 5% FBS y 10%
FCS. La concentración celular se determinó por microscopía utilizando como colorante
azul de tripán para discriminar las células vivas de aquellas muertas. Conocida su
concentración, las células se sembraron en medio DME:M199 (4:1) 5% FBS y 10% FCS a
distintas densidades, según las necesidades experimentales, sobre placas de cultivo
precubiertas con una monocapa de gelatina (2% p/v) en presencia de 5-bromo-2´-
deoxiuridina para inhibir la proliferación de fibroblastos. Las células se mantuvieron a
37°C en una atmósfera húmeda de CO2 al 5% y 95% de aire durante 24 h. Transcurrido
ese tiempo, las células se privaron de suero entre 18 y 24 h antes de su utilización.
5.4. Modelo experimental de inducción de estrés hiposmótico
Para inducir estrés hiposmótico, los cardiomiocitos se trataron con medio DMEM F-
12 diluido 30% con agua nanopura suplementada con CaCl2 y MgCl2, alcanzándose una
concentración hiposmolar de aproximadamente 202 mOsm, determinada mediante un
microsmómetro AdvancedTM 3MO plus.
29
5.5. Determinación del estado funcional de hemicanales formados por conexinas
La apertura de los hemicanales se determinó mediante los cambios en la cinética
de captación de etidio (Etd) por las células. Para el caso de cardiomiocitos 1,0 x 106
células se sembraron en cubreobjetos de vidrio de 12 mm de diámetro en placas de 35
mm (3 cubreobjetos por placa). Las células se preincubaron 5 min con 2 mL de medio
isosmótico (DMEM F-12) con 5 μM Etd y posteriormente se registraron los cambios en la
fluorescencia por 7 min, al cabo de los cuales se diluyó el medio con agua nanopura
suplementada que contenía 5 μM Etd hasta lograr 30% de hiposmolaridad. Finalmente, al
cabo de 7 min de registro con medio hiposmótico, se agregó el inhibidor LaCl3 (200 μM)
disuelto en 100 μL de medio 30% hiposmótico y se registró otros 7 minutos. La
preincubación y todos los registros se realizaron a temperatura ambiente. Los cambios en
la intensidad de fluorescencia se determinaron fotografiando las células cada 20 seg a
longitudes de onda de excitación y emisión de 528 y 598 nm, respectivamente, en un
microscopio de fluorescencia Olympus (Tokio, Japón) BX51W1I acoplado a un sistema de
registro de imágenes “Time Lapse”. En el caso de las células HeLa el experimento se
realizó de la misma manera, pero utilizando medio Locke (154 mM NaCl, 5,4 mM KCl, 2,3
mM CaCl2, 1,5 mM MgCl2, 5 mM glucosa, 5 mM Hepes) como medio isosmótico, el cual
fue diluido 30% con agua nanopura suplementada al igual que se hizo con cardiomiocitos
para obtener medio hiposmótico.
5.6. Determinación de niveles de hemicanales en la superficie celular.
Las cantidades relativas de hemicanales presentes en la membrana plasmática de
las células en cada condición se determinaron mediante la “Biotinilación de la Superficie
Celular” seguido de análisis de Western blot. Para estos fines, 7 x 106 células se
sembraron en placas de 100 mm y se expusieron a distintos tiempos con medio 30%
hiposmótico. Finalizado este período, las células se lavaron 3 veces con medio Hanks de
biotinilación (137 mM NaCl, 5 mM KCl, 1 mM CaCl2, 0,5 mM MgCl2, 0,4 mM KH2PO4, 0.4
mM MgSO4, 0,3 mM Na2HPO4, 5 mM glucosa) y se agregaron 3 mL por placa de una
30
solución 5 mg/mL de Sulfo-NHS-SS-Biotin. Después de 30 min, se retiró la biotina
agregada y se lavaron las células 3 veces con medio Hanks con glicina 15 mM pH 8,
para bloquear la biotina que no reaccionó con las proteínas de superficie.
Posteriormente las células se cosecharon en un tampón con inhibidores de
proteasas en ausencia de detergentes (446 mg PPi, 42 mg NaF, punta espátula de
75,2 mg EDTA y 1 mM PMSF para 10 mL de solución isosmótica) y se centrifugó 2 min a
14.000 RPM para obtener la pella. Luego de retirar todo el sobrenadante, se agregaron 50
μL del mismo tampón con que se cosecharon las células para sonicar en hielo hasta
obtener una mezcla homogénea.
Luego, la concentración de proteínas se determinó con el Método de Bradford
(Bio-Rad protein assay) y se calculó la cantidad de proteínas totales presentes en cada
muestra y se ajustó para que todos los tubos tuvieran la misma cantidad de proteínas.
Sabiendo que aproximadamente 40% de la biotina se une a las proteínas y
aproximadamente 1/3 de la neutravidina se une a la biotina, la cantidad de proteínas se
multiplicó por 0,4 y se dividió por 3 para calcular la cantidad de neutrevidina a agregar. Se
agregó la cantidad necesaria de neutravidina a la pella, se agitó en vórtex y se incubó por
1 h.
Luego, la pella se resuspendió en 1 mL de Hanks 0,1% SDS y 1% NP-40, se
centrifugó 2 min a 14.000 rpm y se descartó el sobrenadante, lo que se repitió 3 veces. A
continuación, se agregó Hanks con 0,1 M glicina pH 2,8 y se agitó con vórtex para separar
el complejo neutravidina-biotina de las proteínas de superficie y se centrifugó 2 min a
14.000 rpm para obtener el sobrenadante.
Finalmente se agregaron 10 μL de Tris-HCl 1M; pH 7,4 para ajustar el pH de las
muestras y buffer de carga 4x. Las muestras se hirvieron y almacenaron a -80°C hasta su
utilización.
Todos los pasos, desde el lavado de las células luego de los estímulos hasta la
obtención de las proteínas finales, se realizaron a 4°C. La cuantificación relativa de los
hemicanales formados por conexina se realizó por Western blot.
31
5.7. Electroforesis en geles de poliacrilamida
El fraccionamiento de las proteínas de acuerdo a su masa molecular se realizó
mediante electroforesis en geles denaturantes de poliacrilamida. Para la detección de los
niveles relativos de hemicanales formados por Cx43, se tomaron 20-30 μg de proteína
total (sin biotinilar) y todo el volumen obtenido en la biotinilación. Las concentraciones de
los geles concentrador y separador fueron 5 y 10%, respectivamente. La electroforesis se
realizó a voltaje constante a 90 V por 3 h en tampón de corrida (3,025 g/L Tris base; 14,4
g/L glicina, 1 g/L dodecil sulfato de sodio)
5.8. Western blot
Una vez realizada la electroforesis, las proteínas se electrotransfirieron a una
membrana de PVDF (BioRad) previamente activada con metanol por 5 min, a amperaje
constante de 400 mA durante 90 min en tampón de transferencia (3,025 g/L Tris base;
14,4 g/L glicina y 20 % v/v metanol).
Para la inmunodetección de hemicanales de Cx43 las membranas se incubaron en
solución de bloqueo (TBS 1X; 0,1% Tween-20 en 5% leche descremada) durante 1 h a
temperatura ambiente y posteriormente se incubaron con el anticuerpo anti Cx43 a un
título de 1:5.000 toda la noche a 4°C con agitación. Para retirar el anticuerpo primario no
unido, las membranas se lavaron 3 veces durante 30 min en TBS 1x/0,1%Tween-20 y se
incubaron durante 1 h a temperatura ambiente con el anticuerpo secundario anti-IgG de
conejo conjugado con peroxidasa, a un título de 1:5.000. Para retirar el exceso de
anticuerpo secundario no unido, las membranas se lavaron nuevamente 3 veces durante
20 min.
Posteriormente, las membranas se incubaron por 1 min en la reacción “Western
Lightning” y se expusieron a la película de fotografía Kodak-Biomax. Las películas se
32
digitalizaron y las imágenes se procesaron con los programas computacionales
Photoshop 6.0 y USI.
5.9. Determinación del volumen celular
Las determinaciones de volumen se realizaron para evaluar la participación de
hemicanales de Cx43 en la RVD. Con esta finalidad 1,0 x 106 células se sembraron sobre
cubreobjetos de vidrio de 25 mm de diámetro en placas de 35 mm. Los cardiomiocitos se
preincubaron a 37°C en presencia de CO2 con 5 μM calceína-acetoximetil ester (0,5%
ácido plurónico) por 10 min y lavaron 2 veces con medio isosmótico (DMEM F-12) para
retirar la calceína que no ingresó a las células. Posteriormente los cubreobjetos se
colocaron en una cámara de perfusión con 700 μL de medio isosmótico (DMEM F-12) y se
registró la fluorescencia basal de los cardiomiocitos (10 min aproximadamente). Para
estimular los cardiomiocitos con medio 30% hiposmótico, se agregó a la placa 300 μL de
agua nanopura suplementada y se homogenizó dos veces con pipeta. Para agregar el
inhibidor Gap26 o su péptido control Gap26 scrambled, se disolvieron minutos antes de su
utilización en 150 μL de medio 30% hiposmótico y se agregaron a las células a los
tiempos señalados, homogenizando 3 veces con pipeta. Todos los registros se realizaron
a temperatura ambiente. Los cambios en la intensidad de la fluorescencia se
determinaron fotografiando las células cada 20 seg a las longitudes de onda de excitación
y emisión a 488 y 510-545 nm, respectivamente, en un microscopio confocal Zeiss LSM
410.
Los valores se estandarizaron asignando a la fluorescencia basal (F0) el valor de 1.
Para la conversión de fluorescencia a volumen relativo se utilizó los datos de la curva de
calibración anteriormente obtenida por Jessica Díaz (Diaz-Elizondo et al., 2006). El factor
de corrección utilizado se obtuvo del intercepto de la ecuación de la recta (F0/FT vs π0/πT)
y a partir de la ecuación VT/V0= {(F0/FT)-n}/ (1-n) se realizó la conversión de los valores.
33
5.10. Expresión de resultados y análisis estadístico
Los resultados mostrados corresponden al promedio ± SEM de, al menos, tres
experimentos independientes. Los datos se analizaro para determinar la significancia
estadística utilizando la prueba t-Student (isosmótico vs hiposmótico en cardiomiocitos) y
la prueba Wilcoxon para el resto de los resultados.
34
6. RESULTADOS
6.1. Determinación del estado funcional de los hemicanales formados por conexina durante el estrés hiposmótico
Para determinar el estado funcional de hemicanales formados por conexina (Hcs-
Cxs) se determinó la cinética de incorporación del trazador de permeabilidad etidio (Etd).
El Etd es un fluorósforo catiónico relativamente grande con una masa molecular de 314
g/mol, que fluoresce al intercalarse en el DNA. Debido a su carácter apolar, el Etd no
difunde a través de la membrana plasmática por lo que ingresa a las células casi
exclusivamente a través de los hemicanales. A diferencia de otras moléculas que se
utilizan para medir la funcionalidad de Hcs-Cxs, el Etd sólo fluoresce una vez dentro de la
célula, por lo que no es necesario retirar la sonda que no ingresa y permite realizar
mediciones de los hemicanales en tiempo real (Schalper et al., 2008). Una mayor
velocidad de incorporación de Etd a la célula es indicativa de Hcs-Cxs más funcionales,
ya sea a causa de una mayor cantidad en la membrana plasmática o porque se abrieron.
Para medir la captación de Etd se registraron los cambios en la fluorescencia de
las células expuestas a las distintas condiciones mediante un microscopio de
fluorescencia acoplado a un sistema de registro de imágenes, como se indicó en
Materiales y Métodos. Para el análisis de las imágenes, se seleccionaron no menos de 30
células por experimento, analizándose dos zonas de cada célula para tener una
fluorescencia más representativa, lo que se muestra en la Figura 3.
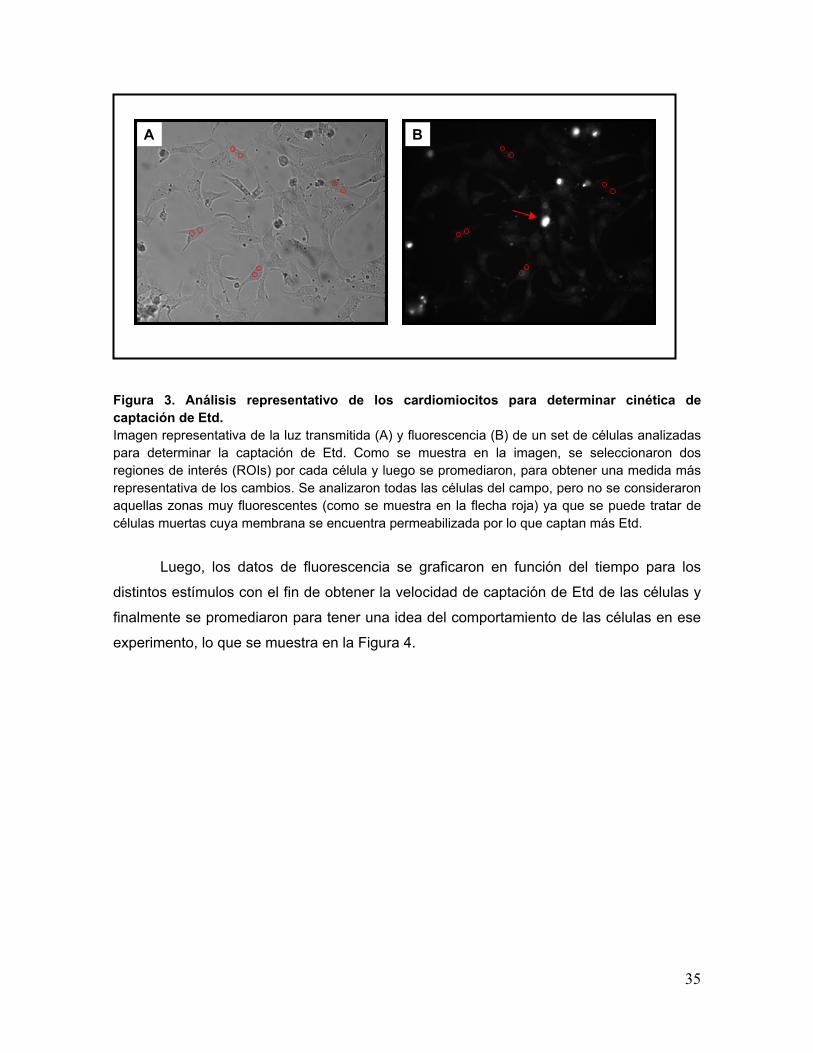
35
Figura 3. Análisis representativo de los cardiomiocitos para determinar cinética de captación de Etd. Imagen representativa de la luz transmitida (A) y fluorescencia (B) de un set de células analizadas para determinar la captación de Etd. Como se muestra en la imagen, se seleccionaron dos regiones de interés (ROIs) por cada célula y luego se promediaron, para obtener una medida más representativa de los cambios. Se analizaron todas las células del campo, pero no se consideraron aquellas zonas muy fluorescentes (como se muestra en la flecha roja) ya que se puede tratar de células muertas cuya membrana se encuentra permeabilizada por lo que captan más Etd.
Luego, los datos de fluorescencia se graficaron en función del tiempo para los
distintos estímulos con el fin de obtener la velocidad de captación de Etd de las células y
finalmente se promediaron para tener una idea del comportamiento de las células en ese
experimento, lo que se muestra en la Figura 4.
A B
36
5
10
15
20
25
30
0 5 10 15 20 25
fluor
esce
ncia
tiempo (min)
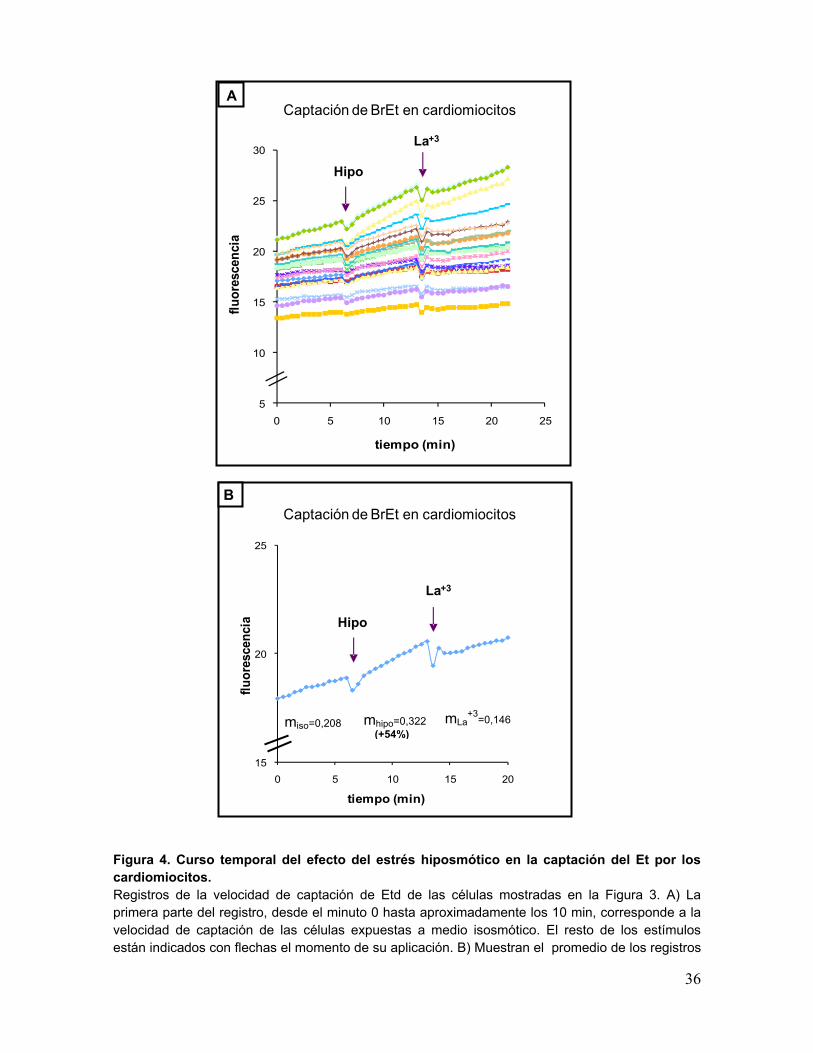
Captación de BrEt en cardiomiocitos
Figura 4. Curso temporal del efecto del estrés hiposmótico en la captación del Et por los cardiomiocitos. Registros de la velocidad de captación de Etd de las células mostradas en la Figura 3. A) La primera parte del registro, desde el minuto 0 hasta aproximadamente los 10 min, corresponde a la velocidad de captación de las células expuestas a medio isosmótico. El resto de los estímulos están indicados con flechas el momento de su aplicación. B) Muestran el promedio de los registros
A
La+3
Hipo
15
20
25
0 5 10 15 20
tiempo (min)
fluor
esce
ncia Hipo
La+3
Captación de BrEt en cardiomiocitosB
miso=0,208 mhipo=0,322 (+54%)
mLa+3=0,146
37
mostrados en A). Los valores de “m” corresponden a las pendientes de captación del estímulo señalado en el subíndice. El valor que aparece entre paréntesis corresponde al porcentaje de cambio en la pendiente con medio hiposmótico respecto al medio isosmótico. El signo positivo indica que la pendiente aumentó.
En las Figuras 4A y 4B se observa que la velocidad de captación de Etd aumenta
cuando se estimula las células con medio hiposmótico, implicando que los Hcs-Cxs se
encuentran más activos o que hay un mayor número de hemicanales en la superficie.
El uso del lantano luego de agregar el medio hiposmótico obedece a 2 razones
principales: 1) para comprobar que el Etd no ingresa a la célula debido a la perdida de la
integridad de su membrana, como ocurre cuando una célula muere por ejemplo, y 2) para
descartar que los aumentos en la captación observados durante el estrés hiposmótico se
deban a la apertura de hemicanales de panexina (Hcs-Pxs), ya que a través de éstos
también puede ingresar el Etd pero a diferencia de los Hcs-Cxs, no son inhibidos por La+3.
En las Figuras 4A y 4B se observa que el La+3 produce una disminución de la
captación de Etd, por lo que se descarta que el ingreso de Etd ocurra a través de la
membrana plasmática permeabilizada o que la incorporación ocurra principalmente a
través de Hcs-Pxs.
El análisis explicado en las Figuras 3 y 4 corresponde a un experimento
independiente. En la Figura 5 se muestra el análisis estadístico de los datos obtenidos en
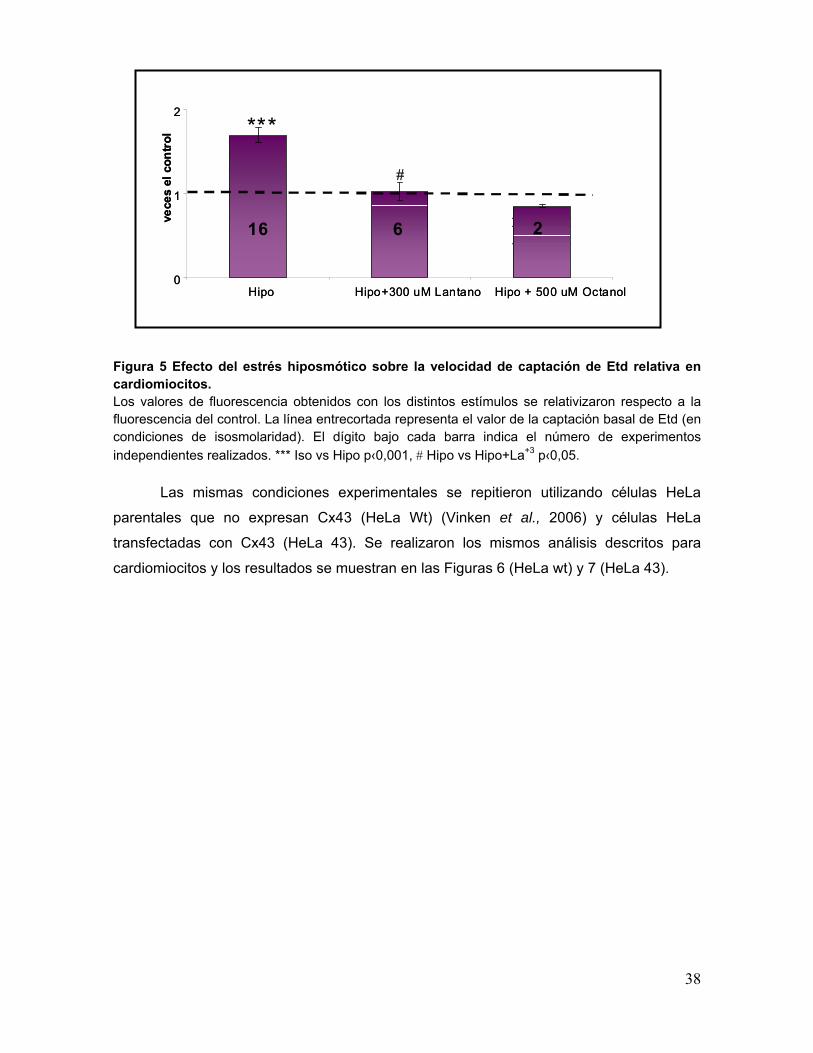
los distintos experimentos. Se observa que el estrés hiposmótico produjo un aumento
significativo de aproximadamente un 70% en la captación de Etd respecto a la condición
basal con medio isosmótico, lo que se revirtió al agregar La+3; alcanzándose valores
cercanos al basal. Además, se probó octanol, otro inhibidor de hemicanales formados por
conexinas, y se observó una tendencia similar a la observada con La+3.
38
Figura 5 Efecto del estrés hiposmótico sobre la velocidad de captación de Etd relativa en cardiomiocitos. Los valores de fluorescencia obtenidos con los distintos estímulos se relativizaron respecto a la fluorescencia del control. La línea entrecortada representa el valor de la captación basal de Etd (en condiciones de isosmolaridad). El dígito bajo cada barra indica el número de experimentos independientes realizados. *** Iso vs Hipo p‹0,001, # Hipo vs Hipo+La+3 p‹0,05.
Las mismas condiciones experimentales se repitieron utilizando células HeLa
parentales que no expresan Cx43 (HeLa Wt) (Vinken et al., 2006) y células HeLa
transfectadas con Cx43 (HeLa 43). Se realizaron los mismos análisis descritos para
cardiomiocitos y los resultados se muestran en las Figuras 6 (HeLa wt) y 7 (HeLa 43).
0
1
2
Hipo Hipo+300 uM Lantano Hipo + 500 uM Octanol
vece
s el
con
trol
***
#
16 6 2
0
1
2
Hipo Hipo+300 uM Lantano Hipo + 500 uM Octanol
vece
s el
con
trol
***
#
16 6 2
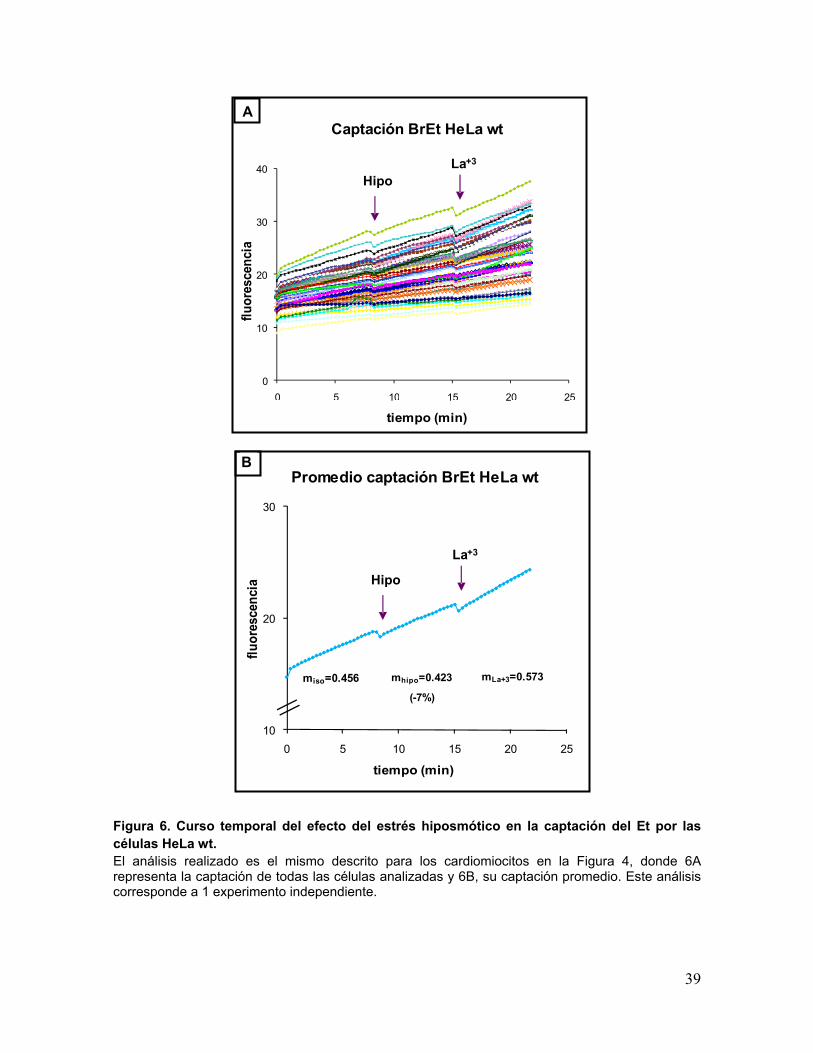
39
Figura 6. Curso temporal del efecto del estrés hiposmótico en la captación del Et por las células HeLa wt. El análisis realizado es el mismo descrito para los cardiomiocitos en la Figura 4, donde 6A representa la captación de todas las células analizadas y 6B, su captación promedio. Este análisis corresponde a 1 experimento independiente.
10 15 2
tiempo (min)
10
20
30
0 5 10 15 20 25
fluor
esce
ncia
Promedio captación BrEt HeLa wt
La+3
Hipo
B
miso=0.456 mLa+3=0.573mhipo=0.423
(-7%)
A
tiempo (min)
fluor
esce
ncia
Captación BrEt HeLa wt
0
10
20
30
40
0 5 10 15 20 25
La+3
Hipo
40
fluor
esce
ncia
0
20
40
60
80
0 5 10 15 20 25
tiempo (min)
Captación BrEt HeLa 43
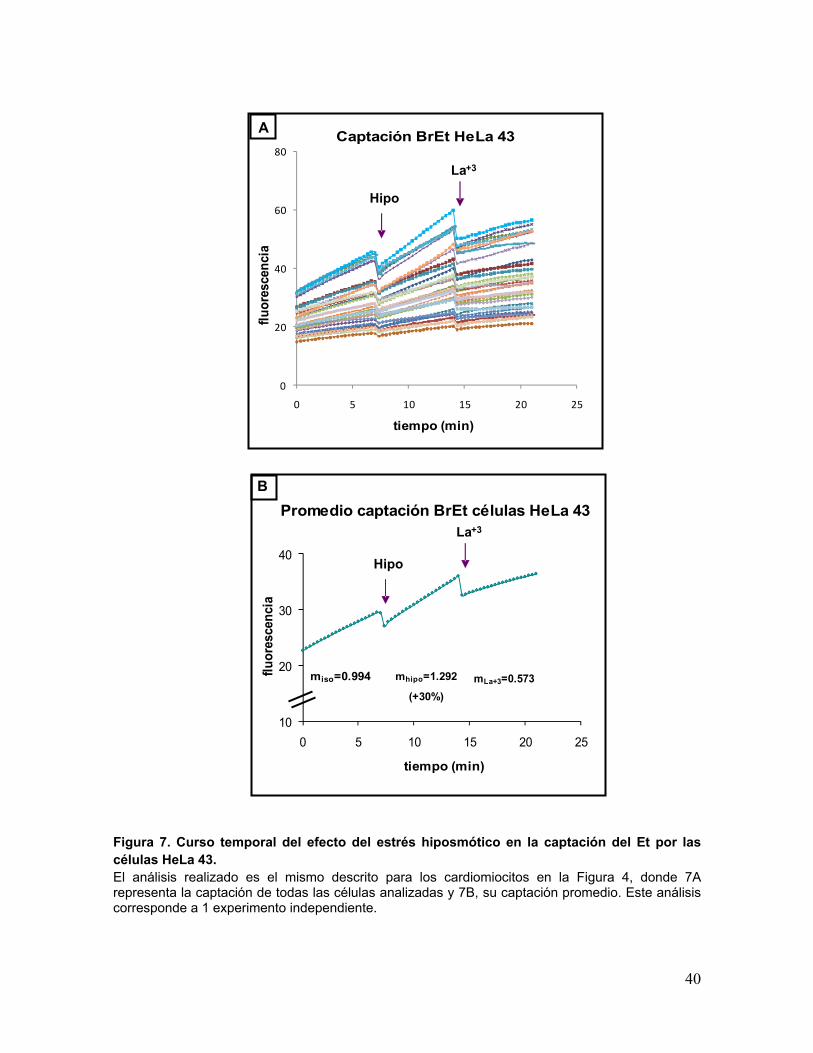
Figura 7. Curso temporal del efecto del estrés hiposmótico en la captación del Et por las células HeLa 43. El análisis realizado es el mismo descrito para los cardiomiocitos en la Figura 4, donde 7A representa la captación de todas las células analizadas y 7B, su captación promedio. Este análisis corresponde a 1 experimento independiente.
A
La+3
Hipo
B
tiempo (min)
fluor
esce
ncia
10
20
30
40
0 5 10 15 20 25
Promedio captación BrEt células HeLa 43
miso=0.994 mhipo=1.292
(+30%)mLa+3=0.573
La+3
Hipo
41
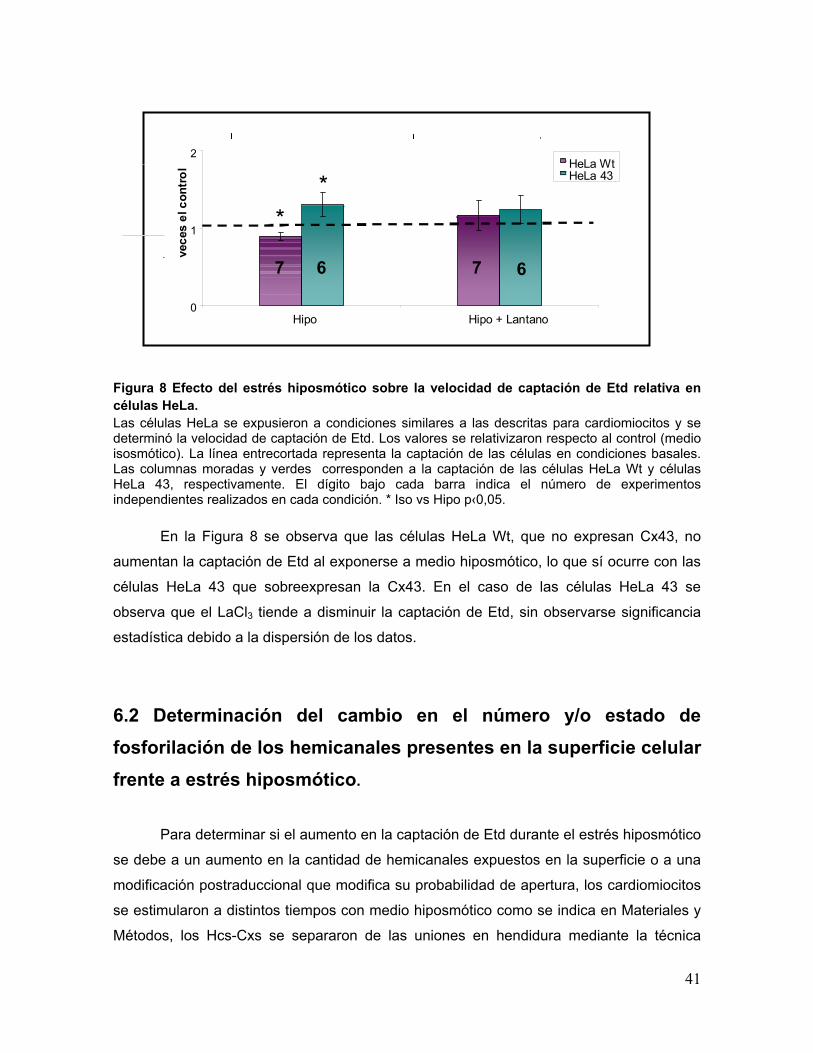
Figura 8 Efecto del estrés hiposmótico sobre la velocidad de captación de Etd relativa en células HeLa. Las células HeLa se expusieron a condiciones similares a las descritas para cardiomiocitos y se determinó la velocidad de captación de Etd. Los valores se relativizaron respecto al control (medio isosmótico). La línea entrecortada representa la captación de las células en condiciones basales. Las columnas moradas y verdes corresponden a la captación de las células HeLa Wt y células HeLa 43, respectivamente. El dígito bajo cada barra indica el número de experimentos independientes realizados en cada condición. * Iso vs Hipo p‹0,05.
En la Figura 8 se observa que las células HeLa Wt, que no expresan Cx43, no
aumentan la captación de Etd al exponerse a medio hiposmótico, lo que sí ocurre con las
células HeLa 43 que sobreexpresan la Cx43. En el caso de las células HeLa 43 se
observa que el LaCl3 tiende a disminuir la captación de Etd, sin observarse significancia
estadística debido a la dispersión de los datos.
6.2 Determinación del cambio en el número y/o estado de fosforilación de los hemicanales presentes en la superficie celular frente a estrés hiposmótico.
Para determinar si el aumento en la captación de Etd durante el estrés hiposmótico
se debe a un aumento en la cantidad de hemicanales expuestos en la superficie o a una
modificación postraduccional que modifica su probabilidad de apertura, los cardiomiocitos
se estimularon a distintos tiempos con medio hiposmótico como se indica en Materiales y
Métodos, los Hcs-Cxs se separaron de las uniones en hendidura mediante la técnica
**
3 3 3 3
Hipo Hipo + Lantano
HeLa parenta lHeLa-43
0
1
2
Hipo Hipo + Lantano
vece
s el
con
trol
HeLa WtHeLa 43
7 6 7 6
**
**
3 3 3 3
Hipo Hipo + Lantano
HeLa parenta lHeLa-43
0
1
2
Hipo Hipo + Lantano
vece
s el
con
trol
HeLa WtHeLa 43
**
3 3 3 3
Hipo Hipo + Lantano
HeLa parenta lHeLa-43
0
1
2
Hipo Hipo + Lantano
vece
s el
con
trol
HeLa WtHeLa 43
7 6 7 6
**
42
“biotinilación de proteínas de superficie” (Musil y Goodenough, 1991). Esta técnica se
basa en la utilización de biotina, una molécula que por su naturaleza semi-polar y gran
tamaño, es impermeable a la célula y reacciona con aminas, principalmente lisinas, que
se encuentran expuestas hacia el medio extracelular (Datasheet del Producto, Pierce
Biotechnology, IL). Esta técnica no permite separar conexinas del resto de las proteínas
de membrana, sino que permite discriminar entre las conexinas que se encuentran
formando parte de uniones en hendidura de aquellas que forman hemicanales, ya que
sólo estas últimas exponen lisinas libres hacia el medio extracelular. Luego de hacer
reaccionar la biotina con las proteínas de la superficie celular, se retira el exceso de
biotina, se lisan las células y se agregan esferas cubiertas de neutravidina que reacciona
con la biotina, para poder separar las proteínas biotiniladas del resto. A continuación se
separan las proteínas del complejo biotina-neutravidina utilizando glicina a pH ácido para
desplazar la unión de las lisinas a la biotina (Schalper et al., 2008). Finalmente, el extracto
proteico enriquecido en Hcs-Cxs se separa mediante electroforesis en geles de
poliacrilamida y posteriormente se cuantifica su número y estado de fosforilación mediante
Western Blot utilizando anticuerpos específicos. La normalización de las intensidades de
las bandas correspondientes a Cx43 biotinilada se realizó considerando el carril marcado
con rojo Ponceau, ya que no se disponen de marcadores proteicos de superficie
susceptibles a ser biotinilados cuya concentración se mantenga constante en las distintas
condiciones como ocurre con la actina en muchos experimentos.
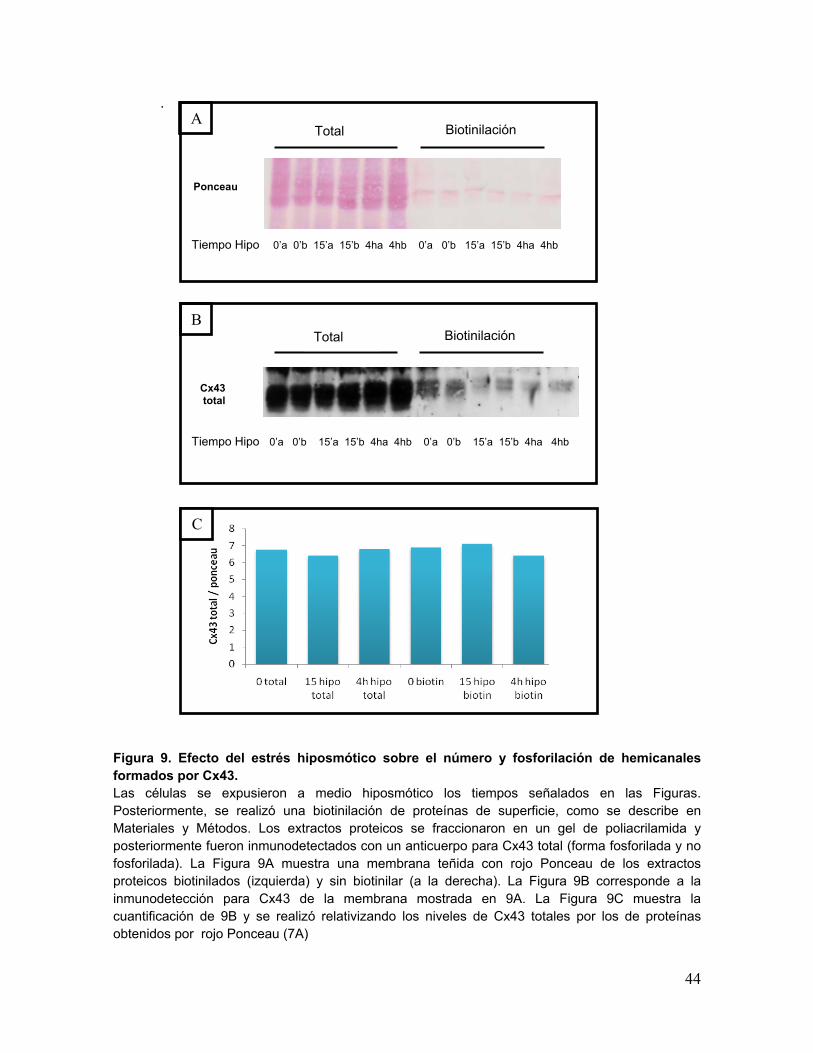
La Figura 9 muestra los resultados de la biotinilación de proteínas de superficie en
cardiomiocitos intactos. Luego de agregar biotina y lisar las células, se cuantificó la
concentración de proteínas y se separaron dos alícuotas: una de aproximadamente 30 µg
proteínas y que no fue tratada con neutravidina y otra que experimentó todo el proceso de
biotinilación (200 µg proteínas). Finalmente ambos extractos se resolvieron en geles de
poliacrilamida y los niveles totales de Cx43 se determinaron por Western blot. La Figura 9
(panel A) muestra el resultado de la tinción de las proteínas transferidas a la membrana
de PVDF con rojo Ponceau. El panel izquierdo de esta Figura muestra los niveles totales
de proteínas presentes en los extractos analizados. Adicionalmente esta misma
membrana permite determinar si el nivel de carga de proteínas entre los distintos carriles
fue homogéneo. Por otra parte, el panel derecho muestra los extractos biotinilados teñidos
con rojo Ponceau, apreciándose una reducción importante en el número de bandas
detectadas.
43
La Figura 9B muestra los niveles de Cx43 presentes en la membrana de la Figura
9A. Se observa la presencia de Cx43 en los extractos biotinilados, al igual que en los
extractos totales, como se esperaba. Las distintas bandas observadas para Cx43
corresponden a las distintas migraciones electroforéticas de la proteína debido a sus
distintos de grados de fosforilación. No se observaron cambios importantes en el patrón
de distribución de las distintas bandas para Cx43, por lo que no habría cambios en el
estado de fosforilación o éstos serían muy pequeños.
Debido a que se partió inicialmente tratando la misma cantidad de proteínas en las
distintas condiciones experimentales, cualquier cambio en los niveles de Cx43 se pueden
correlacionar con los cambios de hemicanales expuestos en la superficie. Para cuantificar
los niveles de hemicanales se sumaron todas las bandas de Cx43 y se relativizó por las
bandas detectadas por rojo Ponceau para cada estímulo. El análisis densitométrico
muestra que no hubo grandes diferencias entre los distintos estímulos (Figura 9C),
sugiriendo la ausencia de cambios en la cantidad de hemicanales presentes en la
membrana en las células expuestas a medio hiposmótico. Sin embargo, dado que se
requiere aumentar el número de experimentos, estos resultados aún no son concluyentes.
44
.
Figura 9. Efecto del estrés hiposmótico sobre el número y fosforilación de hemicanales formados por Cx43. Las células se expusieron a medio hiposmótico los tiempos señalados en las Figuras. Posteriormente, se realizó una biotinilación de proteínas de superficie, como se describe en Materiales y Métodos. Los extractos proteicos se fraccionaron en un gel de poliacrilamida y posteriormente fueron inmunodetectados con un anticuerpo para Cx43 total (forma fosforilada y no fosforilada). La Figura 9A muestra una membrana teñida con rojo Ponceau de los extractos proteicos biotinilados (izquierda) y sin biotinilar (a la derecha). La Figura 9B corresponde a la inmunodetección para Cx43 de la membrana mostrada en 9A. La Figura 9C muestra la cuantificación de 9B y se realizó relativizando los niveles de Cx43 totales por los de proteínas obtenidos por rojo Ponceau (7A)
6.3. Participación de los hemicanales formados por conexinas en la regulación del volumen en cardiomiocitos expuestos a estrés hiposmótico
Trabajos previos de nuestro laboratorio demostraron que los cardiomiocitos no
regulan espontáneamente su volumen cuando son expuestos a estrés hiposmótico (Díaz-
Elizondo et al., 2006), como ocurre con la mayoría de los tipos celulares. Para determinar
si los Hcs-Cxs tenían alguna participación en la regulación de volumen, se evaluó la
capacidad de regular volumen de los cardiomiocitos al exponerlos a medio hiposmótico
utilizando el inhibidor específico de los Hcs-Cx43, el péptido mimético Gap26 (Tabla 1).
Tabla 2 Secuencia aminoacídica del péptido Gap26 y Gap26scr Se muestra la secuencia aminoacídica del péptido mimético de Cx43 Gap26 y su control scrambled Gap26 scr, que posee la misma composición amino acídica, pero en distinta posición. La secuencia está escrita desde el extremo amino al carboxilo terminal
Para evaluar los cambios de volumen en los cardiomiocitos, se utilizó la sonda
fluorescente calceína-AM. Dado el carácter apolar que le confiere el grupo aceto-metil
éster (AM), la calceína puede difundir hacia el interior de la célula a través de la
membrana plasmática. Una vez dentro, las esterasas intracelulares procesan el grupo
AM, por lo que la calceína queda atrapada en el interior. Si una célula aumenta de
volumen, aumenta el contenido de agua al interior de la célula por lo que la calceína se
diluye, disminuyendo la fluorescencia. De la misma forma si una célula pierde volumen,
disminuye la cantidad de agua al interior de la célula, por lo que la calceína se concentra,
aumentando su fluorescencia. Por este motivo, la calceína se puede utilizar para
determinar cambios de volumen en la célula.
Péptido Secuencia aminoacídica
Gap26 V C Y D K S F P I S H V R
GAP26 scr P S F D S R H C I V K Y V
46
El experimento se realizó como se describe en Materiales y Métodos y para
transformar los datos de fluorescencia en volumen relativo, se utilizó los datos de la curva
de calibración anteriormente obtenida por Jessica Díaz (Diaz-Elizondo et al., 2006). En
cada experimento se analizó los cambios de volumen de entre 15-30 células que luego se
promediaron para obtener una curva representativa de cada experimento independiente.
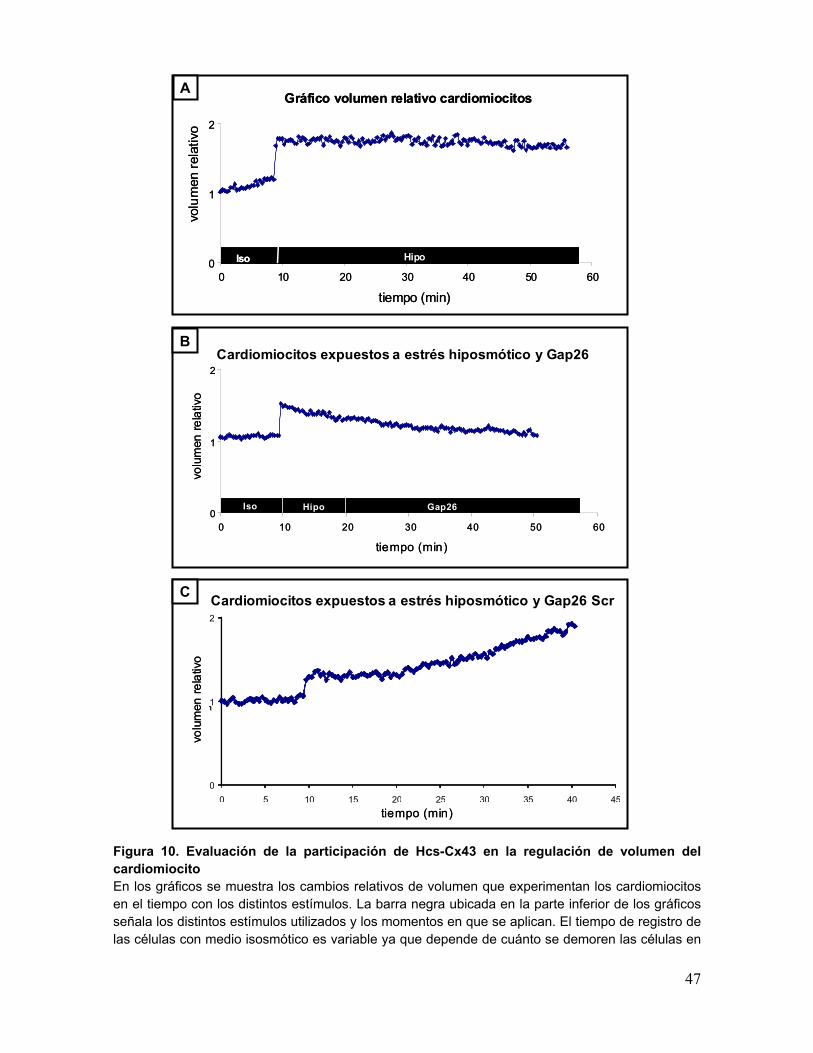
A continuación en la Figura 10 se muestra una imagen representativa de un experimento
independiente.
47
Figura 10. Evaluación de la participación de Hcs-Cx43 en la regulación de volumen del cardiomiocito En los gráficos se muestra los cambios relativos de volumen que experimentan los cardiomiocitos en el tiempo con los distintos estímulos. La barra negra ubicada en la parte inferior de los gráficos señala los distintos estímulos utilizados y los momentos en que se aplican. El tiempo de registro de las células con medio isosmótico es variable ya que depende de cuánto se demoren las células en
Gráfico volumen relativo cardiomiocitos
0
1
2
0 10 20 30 40 50 60
tiempo (min)
volu
men
rela
tivo
Iso HipoIso
Gráfico volumen relativo cardiomiocitos
0
1
2
0 10 20 30 40 50 60
tiempo (min)
volu
men
rela
tivo
Iso HipoIso
A
B
0
1
2
0 10 20 30 40 50 60
tiempo (min)
volu
men
rela
tivo
Iso Hipo Gap26Iso Hipo Gap26Iso Hipo
Iso
Gap260
1
2
0 10 20 30 40 50 60
tiempo (min)
volu
men
rela
tivo
Iso Hipo Gap26Iso Hipo Gap26Iso Hipo
Iso
Gap26
C
0
1
2
0 5 10 15 20 25 30 35 40 45
omn
o
volu
men
rela
tivo
volu
men
rela
tivo
tiempo (min)tiempo (min)
Cardiomiocitos expuestos a estrés hiposmótico y Gap26
Cardiomiocitos expuestos a estrés hiposmótico y Gap26 Scr
48
estabilizar la lectura. En la Figura 10A se muestra un control para demostrar la incapacidad de los cardiomiocitos de regular volumen de forma espontánea. En la Figura 10B se realiza el mismo experimento, pero se agrega el inhibidor de Cx43 Gap26 luego de exponer las células a medio hiposmótico y en Figura 10C se agrega el péptido Gap26 scrambled (Gap26 scr) como control.
En la Figura 10A los cardiomiocitos se expusieron aproximadamente 10 min al
medio isosmótico y luego este medio se diluyó con agua nanopura suplementada hasta
lograr 30% de dilución del medio como se describe en Materiales y Métodos. Se observa
un rápido aumento del volumen celular a casi el doble de su tamaño cuando se estimula
con medio hiposmótico, lo que no regresa a la normalidad durante todo el tiempo de
registro. Estos datos confirman que los cardiomiocitos no regulan espontáneamente su
volumen.
En la Figura 10B se repiten las condiciones del experimento que se muestra en la
Figura 10A, pero se deja aproximadamente 10 min con medio hiposmótico y luego se
agrega el inhibidor de Cx43 Gap26. Este péptido es análogo a una zona de la región C-
terminal de la Cx43 y a tiempos cortos (menores a 1h) produce la inhibición específica de
Hcs-Cx43 (Schalper et al., 2008) por un mecanismo que no está del todo claro. Se
observa que al adicionar Gap26 las células recuperan su volumen a valores cercanos al
que se registra con medio isosmótico, por lo que la Cx43 estaría involucrada en la
regulación de volumen del cardiomiocito.
Para demostrar que el efecto del Gap26 es específico y no se debe a un bloqueo
estérico de algún otro canal involucrado en la regulación de volumen, se utilizó como
control el péptido Gap26 scrambled que tiene la misma composición aminoacídica que el
Gap26, pero sus aminoácidos se encuentran en distinto orden (Tabla 1). En la Figura 10C
se observa que al agregar el péptido Gap26 scr a los cardiomiocitos expuestos a medio
hiposmótico, no regulan su volumen, al igual que ocurre en la Figura 10A, implicando que
es la inhibición específica producto del Gap26, muy probablemente actuando sobre Hcs-
Cx43, lo que posibilita que las células puedan regular su volumen en forma espontánea.
.
49
7. DISCUSIÓN
El corazón está formado principalmente por fibroblastos y cardiomiocitos, formando
estos últimos las unidades contráctiles funcionales del corazón. Debido a la incapacidad
de proliferar de los cardiomiocitos, el corazón es un órgano especialmente vulnerable a
situaciones de estrés como infarto agudo al miocardio, una de las enfermedades más
prevalentes en nuestro país y en países desarrollados (Kannel, 2000; Prieto et al., 1999).
Una de las consecuencias del infarto es el aumento de la osmolaridad intracelular,
produciéndose estrés hiposmótico en las células y con ello un aumento de su volumen
como una forma de contrarrestar las diferencias osmolares con el medio extracelular.
Producto del aumento de volumen, se producen cambios electrofisiológicos en el
cardiomiocito con severas consecuencias para el corazón. El aumento de volumen es
mucho más evidente durante la reperfusión, situación en la cual se reestablece el flujo
sanguíneo para rescatar al cardiomiocito (Wright y Rees, 1998).
Poco se conoce acerca del mecanismo que participa en la regulación de volumen
de los cardiomiocitos, pero se ha propuesto que los hemicanales formados por conexinas
podrían estar formando parte de este proceso al permitir la formación de verdaderos
poros en la membrana plasmática que permiten el movimiento de agua e iones a favor de
su gradiente de concentración (John et al., 2003). Por otro lado, diversas evidencias
sugieren que las conexinas sufren cambios durante los procesos de isquemia-reperfusión,
con importantes consecuencias para las células.
Considerando la evidencia obtenida previamente por nuestro laboratorio de que los
cardiomiocitos aumentan de volumen cuando son expuestos a estrés hiposmótico, pero
son incapaces de regular espontáneamente su volumen (Diaz-Elizondo et al., 2006) y la
creciente evidencia que involucra a las conexinas como actores importantes en la
regulación de volumen, en esta tesis investigó si los Hcs-Cxs están involucrados en la
incapacidad de regular volumen que presentan los cardiomiocitos cuando son expuestos
a estrés hiposmótico.
50
Estrés hiposmótico y estado funcional de los Hcs-Cxs
Para determinar el estado funcional de Hcs-Cxs se utilizó el trazador fluorescente
Etd, ya que dada sus características fisicoquímicas no puede traspasar la membrana
plasmática, siendo prácticamente su única vía de acceso los hemicanales de conexina
abiertos expuestos en la superficie celular. El Etd sólo fluoresce al intercalarse con el
DNA, característica que le confiere una ventaja importante respecto a otros trazadores
utilizados para medir la funcionalidad de Hcs-Cxs, ya que permite hacer mediciones en
tiempo real. Los otros trazadores que se utilizan como “Lucifer yellow”, fluorescen tanto
dentro como fuera de la célula, siendo necesario preincubar y luego lavar las células para
retirar el trazador que no ingresó, por lo que no es posible determinar cómo se modifica la
funcionalidad de los Hcs-Cxs en una misma célula al agregar distintos estímulos, lo que sí
es posible al utilizar Etd.
Al registrar la captación basal de las células con medio isosmótico, se observa que
hay ingreso de Etd a las células, lo que se puede explicar por dos razones: a) existe un
pool de Hcs-Cxs dispuestos en la membrana plasmática que se encuentran basalmente
abiertos, permitiendo el ingreso de Etd observado o b) basalmente los Hcs-Cxs se
encuentran cerrados y el Etd está ingresando por otra vía a las células. Debido a que
existe evidencia que sugiere que en condiciones basales existe apertura de Hcs-Cxs
(Contreras et al., 2003), nos inclinamos a pensar que la captación basal de Etd se debe a
Hcs-Cxs basalmente abiertos, aunque no se puede descartar la otra opción.
Al analizar la cinética de incorporación de Etd en cardiomiocitos estimulados con
medio hiposmótico se observó un aumento en la velocidad de captación de Etd de
aproximadamente un 70%, lo que indicaría Hcs-CXs más funcionales ya sea debido a una
mayor cantidad de Hcs-Cxs expuestos en la superficie o bien a un aumento en su
probabilidad de apertura. Además se observó que la pendiente de incorporación de Etd es
lineal, indicando que el proceso responsable del aumento de la captación ocurriría de
forma simultánea no cooperativa, en algún momento entre que se agrega el estímulo
hiposmótico y los 20 segundos que transcurren hasta que se registra la primera foto
después del estímulo. Dada la rapidez con que ocurre el fenómeno, se descarta que un
aumento en la expresión de la conexina sea responsable de los aumentos en la captación
51
de Etd observados. Sin embargo no se descarta que este proceso pueda operar en casos
de estrés hiposmótico más prolongados.
Recientemente con el descubrimiento de las panexinas, proteínas estructural y
funcionalmente muy similares a las conexinas, ha sido necesario ser más estrictos en el
análisis de los resultados y tratar de disecar su participación utilizando inhibidores
específicos. El La+3, un inhibidor no-selectivo de canales de cationes divalentes, permite
discriminar entre Hcs-Cxs y Hcs-Pxs, ya que está reportado que sólo inhibe Hcs-Cxs
(Schalper et al., 2008). Por esta razón y para descartar que el aumento de la captación
durante el estrés hiposmótico se debiera a la pérdida de integridad de la membrana
plasmática producto de la muerte de las células, se utilizó LaCl3 luego de estimular con
medio hiposmótico.
Se observó que al agregar LaCl3 las células disminuían la incorporación de Etd a
niveles similares a los basales en la mayoría de los casos. En ocasiones el LaCl3 inhibió y
otras veces aumentó mucho la captación de Etd. Estas diferencias se pueden explicar por
la aplicación del inhibidor, ya que al estar disuelto en un volumen pequeño y por el hecho
de que el objetivo del microscopio está sumergido en la solución de registro, es muy difícil
que el inhibidor quede homogéneamente distribuido, por lo que puede alcanzar
concentraciones tóxicas si se administra muy cerca de las células que se está registrando
o no inhibir, si se agrega muy lejos de las mismas. En el caso del octanol, otro conocido
inhibidor de Hcs-Cxs, también se observó una tendencia a la inhibición de la captación de
Etd, pero al igual que en el caso del LaCl3, fue común observar aumentos en la captación
lo que se atribuyó a los mismos problemas de administración comentados para el LaCl3,
sumándose además el hecho de que el octanol se administra disuelto en etanol absoluto,
lo que puede producir toxicidad en las células.
La interpretación de los datos obtenidos al utilizar LaCl3 como inhibidor debe ser
cuidadosa, ya que el LaCl3 es inespecífico y además de inhibibir Hcs-Cxs inhibe canales
de cationes divalentes en general, como por ejemplo canales de calcio. En el caso de la
determinación de captación de Etd, si bien no podemos descartar que el La+3 inhiba
canales divalentes, es poco probable que el Etd pase a través de éstos debido a su gran
tamaño, quedando casi como única alternativa para su ingreso los Hcs-Cxs, por lo que el
LaCl3 se puede utilizar como inhibidor en este caso.
52
Al determinar la captación de Etd en células HeLa parentales (no expresan
conexinas) estimuladas con medio hiposmótico, se observó que no aumenta la captación
de Etd a diferencia de lo observado en cardiomiocitos, incluso se observa una ligera
disminución en la captación, hecho que puede ser explicado considerando la existencia
de otra vía de ingreso para el Etd en estas células que durante el estrés hiposmótico se
inhibiría, observándose la disminución en la captación de Etd. En el caso de las células
HeLa 43 (que expresan la Cx43), al ser estimuladas con medio hiposmótico se observó un
aumento en la incorporación de Etd, de forma similar a lo que se observa en
cardiomiocitos pero de menos magnitud, lo que se puede deber a una menor expresión de
la Cx43 en células HeLa 43 respecto a cardiomiocitos o a la presencia de otras conexinas
en cardiomicitos que pudieran estar contribuyendo al efecto observado. Estos datos
sugieren que la presencia de Hcs-Cx43 es importante para el aumento en la captación de
Etd que se observa en células expuestas a estrés hiposmótico.
En términos generales los datos obtenidos de la captación de Etd sugieren que el
medio hiposmótico aumenta la funcionalidad de Hcs-CXs que podría ser responsable del
aumento de volumen observado en los cardiomiocitos al exponerse a estrés hiposmótico
(Díaz-Elizondo et al., 2006). Al estar abiertos los hemicanales, el agua y los iones se
moverían a través de su gradiente de concentración y considerando que la osmolaridad
intracelular de los cardiomiocitos en condiciones hiposmóticas es superior a la
extracelular, la tendencia del agua sería a ingresar a las células, produciéndose el
aumento de volumen observado.
Estrés hiposmótico y cambio en la actividad de los hemicanales: aumento de los Hcs-Cxs en la superficie v/s cambio en el estado de fosforilación
El aumento de la captación de Etd observado en los cardiomiocitos expuestos a
hiposmolaridad se puede explicar básicamente por dos mecanismos: a) aumento de HCs-
Cx en la superficie celulares o b) aumento de la probabilidad de apertura del HC-Cx
debido a una modificación postraduccional, como por ejemplo un cambio en el estado de
53
fosforilación. Para determinar ambas posibilidades, se realizó biotinilación de las proteínas
de superficie. Esta técnica permite separar Cxs que forman parte de uniones en hendidura
de aquellas que forman parte de hemicanales, ya que sólo estas últimas exponen lisinas
libres hacia el medio extracelular, requisito necesario para que la biotina reaccione y se
una. De esta forma, mediante Western blot de los extractos biotinilados, se puede
determinar los cambios en los niveles de Cx43 y en su estado de fosforilación.
Los resultados obtenidos sugieren que no hay diferencias importantes en los
niveles o estado de fosforilación de los hemicanales formados por Cx43 al exponer las
células a medio hiposmótico que permitan dar cuenta del aumento en la captación de Etd
observado. Sin embargo se debe tener en cuenta los siguientes aspectos en este análisis:
1) Para evaluar confiablemente la cantidad de hemicanales presentes en la
superficie por esta metodología se requiere partir de la misma cantidad inicial
de proteínas. Con este fin se partió de la misma cantidad de proteínas para
biotinilar en los distintos estímulos. Sin embargo, este proceso es complejo
experimentalmente y en varias de sus etapas (por ejemplo durante los lavados)
pueden perderse proteínas e inducir falsas interpretaciones. Por ello los
resultados de Cx43 se relativizaron con las bandas teñidas en la membrana
con rojo Ponceau.
2) No fue posible lograr una separación clara de las distintas bandas de Cx43. Se
observaron distintos patrones de migración electroforéticos para Cx43 producto
de sus cambios de fosforiilación pero que no pudieron ser analizadas
individualmente, pudiéndose realizar sólo un análisis cualitativo.
Si no se alterara el estado de fosforilación ni el número de hemicanales en la
superficie celular, una posible alternativa para explicar el aumento en la captación de Etd
sería que ocurriera otra modificación postraduccional de las conexinas que alterara su
probabilidad de apertura, como por ejemplo una modificación redox como nitrosilación. En
trabajos realizados en nuestro laboratorio se ha descrito que la formación de especies
reactivas del oxígeno (EROs) es importante en el proceso de regulación de volumen
(Rojas-Rivera et al., 2009), por lo que puede ser que estos EROs generados produzcan
modificaciones en las Cxs. Consecuentemente, se realizaron experimentos preliminares
54
de biotinilación para ver si se alteraba el patrón de nitrosilación de los hemicanales, sin
embargo hubo resultados contradictorios por lo que no se incluyeron en esta tesis.
Como se dijo anteriormente, la biotinilación de proteínas de superficie permite
separar proteínas que exponen lisinas libres hacia el medio extraceluar, no sólo
conexinas. El corazón expresa abundantemente Cx43, pero también expresa otras
conexinas como Cx45 y Cx40. En nuestro caso sólo se analizó la abundancia y estado de
fosforilación de la Cx43 y puede ser que otras conexinas se estén modificando durante el
estrés hiposmótico. Pero si consideramos que en los experimentos de volumen los
cardiomiocitos logran regular volumen luego de la administración de Gap26, un inhibidor
selectivo para Cx43, esta explicación parece menos probable.
Otra explicación a lo observado en la biotinilación es que los hemicanales estén
funcionando como mecanosensores que responden al estiramiento de la membrana
cuando la célula aumenta de volumen. Si este fuera el caso habría que plantearse cuáles
serían las posibles vías de ingreso del agua para permitir el hinchamiento de la célula.
Una posibilidad es que entre una pequeña cantidad por aquaporinas, produciendo un
pequeño aumento de volumen de la célula, lo que a su vez activaría a HCs-Cx de la
membrana plasmática que actuarían como mecanosensores. Otra posibilidad es que un
pequeño número de HCs-Cx estén abiertos basalmente y permitan el ingreso de agua
produciendo un pequeño aumento de volumen y posteriormente se abran más HCs-Cx
para permitir el hinchamiento de la célula.
Aumento de volumen durante el estrés hiposmótico y su relación con Hcs-Cxs
En nuestro laboratorio se había observado previamente que al estimular
cardiomiocitos con medio hiposmótico se producía un aumento de su volumen, el cual no
regresaba a valores basales espontáneamente como ocurre en células que son capaces
de regular su volumen, sino que lo recuperaban sólo cuando se restablecía el medio
isosmótico (Díaz-Elizondo et al., 2006). De esta observación surgió la hipótesis de que los
cardiomiocitos no son capaces de realizar el RVI y esta tesis se propuso determinar si los
hemicanales formados por conexina están relacionados con este proceso.
55
Para realizar estos experimentos de volumen se utilizó el marcador fluorescente
Calceína-AM, que debido al carácter apolar que le confiere su grupo aceto-metil ester
(AM) puede ingresar a la célula a través de la membrana lipídica. Una vez dentro no
puede volver a salir ya que las esterasas intracelulares cortan el grupo AM, perdiendo su
carácter apolar que le permite traspasar membranas. Debido a esta característica se
utiliza la calceína-AM para medir volumen, ya que su intensidad de fluorescencia refleja el
contenido de agua de la célula. De esta forma, al disminuir la cantidad de agua de la
célula, se concentra la calceína y aumenta la intensidad de fluorescencia y, por el
contrario, si aumenta la cantidad de agua de la célula la calceína se diluye y la
fluorescencia disminuye.
Debido a que se utilizó un medio distinto al utilizado previamente en nuestro
laboratorio (el nuevo medio no contenía rojo fenol) y a que las condiciones de la medición
de volumen variaron ya que esta vez se realizaron los experimentos sin reperfusición, en
primer lugar se comprobó que los hechos observados previamente y reportados en la
literatura (Díaz-Elizondo et al., 2006) eran reproducibles. Al igual que experimentos
anteriores, se observó que el medio hiposmótico produce un aumento de volumen de los
cardiomiocitos, el cual no retorna a valores basales por sí solo, tal como se había descrito
previamente (Díaz-Elizondo et al., 2006).
Al diluir el medio para lograr el medio hiposmótico, es necesario resuspender para
homogenizar la solución. Debido a que está documentado que las conexinas responden a
estímulos mecánicos (Cherian et al., 2005), se comprobó que la acción de resuspender
por sí sola no estuviera relacionada con los cambios de volumen observados durante la
aplicación del medio hiposmótico (datos no mostrados).
Para los experimentos de volumen se utilizó como inhibidor de Hcs-Cx43 el
péptido mimético Gap26, que a tiempos cortos inhibe específicamente hemicanales. Al
agregar Gap26 luego del estímulo hiposmótico, se observó que las células lograban
realizar el RVI, alcanzándose niveles similares al basal (obtenido con medio isosmótico),
lo que no ocurrió al agregar Gap26 scr por lo que se descarta una acción no específica
del péptido. También se utilizó La+3 como inhibidor obteniéndose resultados similares a
los obtenidos con Gap26 (datos no mostrados) pero en este caso no es posible
56
discriminar si el efecto del La+3 es debido al bloqueo de Hcs-Cxs o a la inhibición de por
ejemplo canales de calcio, que son importantes para el proceso de regulación de volumen
(Rojas-Rivera et al., 2009).
De los resultados obtenidos con los experimentos de volumen se puede deducir
que los Hcs-Cx43 son importantes para la regulación de volumen del cardiomiocito.
Debido a que no se utilizó un inhibidor más amplio de canales formados por Cxs, no
podemos descartar que otras Cxs pudieran cumplir también un rol importante en la
regulación de volumen, pero el hecho de que al inhibir selectivamente la Cx43 se logre
una recuperación de volumen a niveles similares al basal, nos hace pensar que
probablemente la Cx43 cumple un rol importante en este proceso. Si a estos resultados
sumamos los obtenidos de la captación de Etd, podemos inferir que el estrés hiposmótico
produce la apertura de Hcs-Cxs, impidiendo que el cardiomiocito regule espontáneamente
su volumen.
Las AQPs tradicionalmente se han asociado a la regulación de volumen de
diversos tipos celulares, cumpliendo un papel fundamental en las células renales (Brown
et al., 2009). En el caso de cardiomiocitos no está clara su función e incluso existe
controversia respecto a su expresión (Egan et al., 2006). Dado que no se utilizaron
inhibidores de AQPs para determinar su rol en la regulación de volumen durante el estrés
hiposmótico, no es posible descartar su contribución en el aumento de volumen que se
observa en el cardiomiocito.
Los resultados obtenidos en esta tesis permiten demostrar por primera vez que los
Hcs-Cx participan en la regulación de volumen del cardiomiocito, algo que se suponía
pero no había sido demostrado directamente. Además, se propone un mecanismo que
podría explicar por qué las células aumentan de volumen durante el estrés hiposmótico. Si
pensamos en lo que ocurre durante la reperfusión, donde ocurre el mayor daño, la
apertura de los Hcs-Cxs podría explicar el gran aumento de volumen que ocurre, ya que
durante la reperfusión se produce la mayor diferencia de osmolaridad entre el medio intra
y extracelular, por lo que el agua entraría rápidamente a la célula produciendo finalmente
su muerte
57
8. CONCLUSIONES
• El estrés hiposmótico aumenta la funcionalidad de hemicanales formados por
conexina en el cardiomcito
• El aumento en la funcionalidad de los hemicanales formados por Cx43 no se
debería a un aumento en el número de hemicanales ni a cambios en el nivel de
fosforilación de los mismos
• Los hemicanales formados por Cx43 cumplen un papel importante en la regulación
de volumen del cardiomiocito, ya que su cierre permite la regulación de volumen
espontánea del cardiomiocito en condiciones hiposmóticas.
58
9. BIBLIOGRAFÍA
Barbe, M. T., Monyer, H. and Bruzzone, R. (2006). Cell-cell communication beyond
connexins: the pannexin channels. Physiology (Bethesda). 21: 103-114
Beardslee, M. A., Lerner, D. L., Tadros, P. N., Laing, J. G., Beyer, E. C., Yamada, K. A.,
Kleber, A. G., Schuessler, R. B. and Saffitz, J. E. (2000). Dephosphorylation and
intracellular redistribution of ventricular connexin43 during electrical uncoupling induced by
ischemia. Circ Res. 87: 656-662
Brown, D., Breton, S., Ausiello, D.A. and Marshansky, V. (2009) Sensing, signaling and
sorting events in kidney epithelial cell physiology. Traffic. 10: 275-284
Cherian, P.P., Siller-Jackson, A.J., Gu, S., Wang, X., Bonewald, L.F., Sprague, E. and
Jiang, J.X. (2005) Mechanical strain opens connexin 43 hemichannels in osteocytes: a
novel mechanism for the release of prostaglandin. Mol Biol Cell. 16: 3100-3106.
Cokkinos, D.V., Pantos, C., Heusch, G. and Taegtmeyer, H. Myocardial Ischemia: from
mechanism to therapeutic potentials. USA, Springer, 204p, 2006.
Contreras, J. E., Sáez, J. C., Bukauskas, F. F. and Bennett, M. V. (2003). Gating and
regulation of connexin 43 (Cx43) hemichannels. Proc Natl Acad Sci U S A. 100: 11388-
11393
Davis, B. A., Hogan, E. M. and Boron, W. F. (1992). Role of G proteins in stimulation of
Na-H exchange by cell shrinkage. Am J Physiol. 262: C533-536
Diaz-Elizondo, J., Chiong, M., Rojas-Rivera, D., Olea-Azar, C., Kwon, H. M. and
Lavandero, S. (2006). Reactive oxygen species inhibit hyposmotic stress-dependent
volume regulation in cultured rat cardiomyocytes. Biochem Biophys Res Commun. 350:
1076-1081
59
Decrock, E., Vinken, M., De Vuyst, E., Krysko, D.V., D'Herde, K., Vanhaecke, T.,
Vandenabeele, P., Rogiers, V and Leybaert, L (2009). Connexin-related signaling in cell
death: to live or let die?. Cell Death Differ. 16:524-36
Drewnowska, K. and Baumgarten, C. M. (1991). Regulation of cellular volume in rabbit
ventricular myocytes: bumetanide, chlorothiazide, and ouabain. Am J Physiol. 260: C122-
131
Dunham, P. B., Jessen, F. and Hoffmann, E. K. (1990). Inhibition of Na-K-C1 cotransport
in Ehrlich ascites cells by antiserum against purified proteins of the cotransporter. Proc
Natl Acad Sci U S A. 87: 6828-6832
Egan, J. R., Butler, T. L., Au, C. G., Tan, Y. M., North, K. N. and Winlaw, D. S. (2006).
Myocardial water handling and the role of aquaporins. Biochim Biophys Acta. 1758: 1043-
1052
Fettiplace, R. and Haydon, D. A. (1980). Water permeability of lipid membranes. Physiol
Rev. 60: 510-550
Fettiplace, R. and Haydon, D. A. (1980). Water permeability of lipid membranes. Physiol
Rev. 60: 510-550
Fisher, R. S. and Spring, K. R. (1984). Intracellular activities during volume regulation by
Necturus gallbladder. J Membr Biol. 78: 187-199
Franco, R., Lezama, R., Ordaz, B. and Pasantes-Morales, H. (2004). Epidermal growth
factor receptor is activated by hyposmolarity and is an early signal modulating osmolyte
efflux pathways in Swiss 3T3 fibroblasts. Pflugers Arch. 447: 830-839
Garcia-Dorado, D. and Oliveras, J. (1993). Myocardial oedema: a preventable cause of
reperfusion injury? Cardiovasc Res. 27: 1555-1563
Goodenough, D. A. and Paul, D. L. (2003). Beyond the gap: functions of unpaired