VETERINÁRNÍ A FARMACEUTICKÁ UNIVERZITA BRNO FAKULTA VETERINÁRNÍ HYGIENY A EKOLOGIE Ústav hygieny a technologie mléka MIKROBIOLOGICKÉ LABORATORNÍ METODY MVDr. Šárka Bursová, Ph.D. Mgr. Marta Dušková, Ph.D. MVDr. Lenka Necidová, Ph.D. doc. MVDr. Renáta Karpíšková, Ph.D. Mgr. Petra Myšková BRNO 2014
Transcript
VETERINÁRNÍ A FARMACEUTICKÁ UNIVERZITA BRNO
FAKULTA VETERINÁRNÍ HYGIENY A EKOLOGIE
Ústav hygieny a technologie mléka
MIKROBIOLOGICKÉ LABORATORNÍ
METODY
MVDr. Šárka Bursová, Ph.D.
Mgr. Marta Dušková, Ph.D.
MVDr. Lenka Necidová, Ph.D.
doc. MVDr. Renáta Karpíšková, Ph.D.
Mgr. Petra Myšková
BRNO 2014
1
Tato skripta jsou spolufinancována z Operačního programu Vzdělávání pro konkurenceschopnost:
„Inovace bakalářského a navazujícího magisterského studijního programu v oboru Bezpečnost a
kvalita potravin“
(reg. č. CZ.1.07/2.2.00/28.0287)
Obsah
2
OBSAH
1. KULTIVAČNÍ METODY (MVDr. Šárka Bursová, Ph.D.) .............................................. 7
7.3.3.1. Provedení a vyhodnocení testu ............................................................................ 58
7.3.3.2. Výhody, nevýhody a využití Etestu v praxi ......................................................... 59
7.4. Genotypové metody stanovení rezistence ........................................................ 59 7.4.1. Odhalování mutací spojených s rezistencí k AML .............................................. 59
7.4.2. Výhody a nevýhody použití genotypových metod .............................................. 60
Velké využití má také při identifikaci klinicky významných multirezistentních bakterií.
2.2. Systém Biolog Pro identifikaci a charakterizaci mikrobiologických kultur vyvinula firma Biolog, Inc. systém
Biolog – rychlý a levný prostředek pro získání velkého množství informací o analyzovaných
mikroorganismech.
2.2.1. Základní principy Princip tohoto identifikačního systému spočívá ve schopnosti mikroorganismů metabolizovat
sacharidové substráty umístěné v 96-ti jamkové mikrotitrační destičce. Pokud naočkované
mikroorganismy mají enzymy schopné metabolizovat dané substráty, dochází k uvolnění
elektronů ze substrátu a probíhá oxidace. Tyto elektrony přijímá barvivo
trifenyltetrazoliumchlorid (TTC), čímž se redukuje a dochází ke vzniku trifenylformazanu
(TPF), reakce je doprovázena změnou barvy. Dostatečné množství buněk v jamce
mikrodestičky tak zajišťuje pozorovatelnou barevnou změnu. Tímto způsobem každý
mikroorganismus získá specifický kód, díky kterému je identifikován.
Postup analýzy je jednoduchý. Nejdříve se na agaru izoluje čistá kultura, poté se připraví
inokulum o předepsané koncentraci buněk a naočkuje se vhodná mikrotitrační destička.
V průběhu času dochází při inkubaci k vývoji barvy v jednotlivých jamkách podle toho, jak
mikroorganismus využívá jednotlivé substráty. Po inkubaci je mikrodestička umístěna do
mikrostanice pro analýzu, kde reader odečte absorbanci (k odečtům je použito dvou vlnových
délek, 590 nm a 750 nm). Výsledky jsou zaznamenány a srovnány s širokou databází
mikroorganismů, ve které je uloženo přes 2000 druhů. Systémy existují manuální (MicroLog),
poloautomatické (MicroStation) – zahrnující kromě softwaru i identifikační reader, a plně
automatické – systém OmniLog® (software, inkubátor/reader).
2.2.2. Výhody, nevýhody a využití systému Biolog v praxi Výhodou systému Biolog je rychlá a poměrně přesná identifikace mikroorganismů díky
široké škále biochemických reakcí obsažených v mikrodestičce a komplexní databázi
mikroorganismů (bakterie, kvasinky, plísně). Nevýhodou tohoto systému je možnost
identifikace pouze kultivovatelných mikroorganismů, pro vhodný výběr panelu testů se musí
předem nadefinovat, o jakou skupinu mikroorganismu se jedná. Dále je zde potenciální riziko
křížových reakcí, u manuální verze není možné měřit absorbanci (vyhodnocení je vizuální).
Původně měl systém Biolog sloužit pro snadnou identifikaci klinicky významných bakterií.
Nyní jsou mikrodestičky vyvinuty pro různé skupiny mikroorganismů – aerobní bakterie,
anaerobní bakterie, grampozitivní bakterie, gramnegativní bakterie, kvasinky a plísně.
V nejnovější verzi tohoto systému lze identifikovat na stejném panelu jak grampozitivní, tak
gramnegativní bakterie zároveň, a tím odpadá před vlastní analýzou barvení dle Grama a další
testování pro výběr vhodného panelu substrátů.
Systémy identifikace mikroorganismů
13
Kromě identifikace mikroorganismů má systém Biolog také možnost sledovat funkční
diverzitu mikrobiálních společenstev se speciálními testovacími panely pro ekologické a
ekotoxikologické studie.
2.3. Hmotnostní spektrometrie MALDI-TOF Rychlou a velmi přesnou identifikaci a typizaci mikroorganismů poskytuje hmotnostní
spektrometrie s laserovou desorpcí a ionizací za účasti matrice s průletovým analyzátorem (MALDI-TOF MS, Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass
Spectrometry). Hmotnostní spektrometrie MALDI-TOF patří mezi chemotaxonomické
metody. Proces identifikace je založen na analýze ribosomálních a dalších proteinů v buňce.
Ribosomální proteiny představují asi 20 % veškerých buněčných bílkovin a asi 3 % celkové
hmoty. Tyto makromolekuly jsou specifické pro jednotlivé bakteriální druhy a díky tomu
mohou být považovány za vhodné biomarkery. Hmotnostní spektrometrie je vysoce citlivá
analytická technika s detekčními limity řádově až desetiny femtomolů.
Obrázek 2: Schéma přístroje MALDI-TOF MS.
2.3.1. Základní principy Jedná se o šetrnou ionizační metodu, kdy biomolekuly (proteiny buněčných extraktů) nejsou
po ataku laseru štěpeny, ale pouze ionizovány pomocí matrice (obrázek 3). Nejčastěji se
využívají dusíkové UV lasery, v menší míře infračervené (IR) lasery. Ozáření o vlnové délce
337 nm trvá 4 ns. Matrice zajistí kontakt analyzované molekuly s laserem tak, aby
biomolekula nebyla atakována přímo a
štěpena nežádoucím způsobem. Matrice
dále zprostředkovává přenos energie, kdy
excitované molekuly matrice za vysokého
tlaku ionizují molekuly analytu
(biomolekuly) přenosem protonu.
Ionty, které přešly do plynné fáze, postupují
přes silné elektrické pole, ve kterém jsou
urychleny a zaostřeny. Vstoupí do vakuové
trubice hmotnostního analyzátoru doby
letu (TOF, Time of Flight), kde se pohybují
Obrázek 3: Schéma ionizace.
Systémy identifikace mikroorganismů
14
rychlostí úměrnou jejich hmotnosti a náboji (obrázek 2). Měří se doba letu částice, lehčí či
více nabité ionty dorazí k detektoru dříve než těžší nebo méně nabité. Detektor je propojen
s počítačem a pomocí softwaru jsou data zpracována.
Pro každý kmen je
vytvořeno hmotnostní
spektrum, neboli profil jeho
proteomu. Toto hmotnostní
spektrum je zobrazením
četnosti ionizovaných částic
buněčného proteomu.
Profily proteinů jsou pro
daný druh mikroorganismu
vysoce charakteristické.
Vlastní identifikace
mikroorganismů následně
spočívá ve srovnávání
proteinového spektra izolátu
se spektry referenčních
kmenů v databázi MALDI Biotyper. Z matice podobnosti spekter se provádí klastrová
(shluková) analýza. Shluky podobnosti se zobrazují jako dendrogram (obrázek 4). Na základě
získaného dendrogramu je založena klasifikace v rámci rodu, druhu, a v některých případech i
poddruhu či typizace kmene. Míra spolehlivosti identifikace (podobnost izolátu s referenčním
kmenem v databázi) se vyjadřuje jako log(score). Pokud je log(score) vyšší než hodnota 2,3,
jedná se o vysoce pravděpodobnou identifikaci na úrovni druhu. Log(score) v rozmezí 2,3 –
2,0 potvrzuje vysoce pravděpodobnou identifikaci na úrovni rodu a pravděpodobnou
identifikaci na úrovni druhu. Pravděpodobnou identifikaci na úrovni rodu mají kmeny
s log(score) 2,0 – 1,7. Výsledky s hodnotami pod 1,7 nejsou signifikantní. Samotná analýza je
velmi rychlá a výsledky jsou možné získat během několika minut.
2.3.1.1. Příprava izolátů
Příprava izolátů pro analýzu MALDI-TOF MS je velmi jednoduchá. Analyzovat se mohou
celé buňky nebo jejich extrakty. Nejrychlejším způsobem je nanesení mikroorganismů
vykultivovaných na agaru či v bujónu, promytých deionizovanou vodou a s přídavkem
matrice, přímo na MALDI destičku. Buněčné proteiny je doporučeno extrahovat zejména u
grampozitivních bakterií vzhledem ke struktuře jejich buněčné stěny (silná vrstva
peptidoglykanu) např. působením enzymů lysozymu či lysostaphinu, chemických
rozpouštědel (ethanolu, acetonitrilu, směsi chloroform-methanol), ultrazvukové sonikace či
tepelného záhřevu. Čisté 24 hodinové bakteriální
kultury jsou resuspendovány a inaktivovány v 75%
ethanolu. Po centrifugaci a důkladném odstranění
supernatantu je k buňkám přidána matrice v poměru
1:104. Matrice musí absorbovat vlnovou délku
použitého laseru, musí být fotostabilní a tvořit malé
krystaly. Nejčastěji se používají deriváty kyseliny
benzoové či skořicové. Výběr vhodné matrice je
zcela zásadní pro kvalitu analýzy.
Jako rozpouštědlo se používá acetonitril, aceton či
organická kyselina ve směsi s vodou a pro zvýšení
Obrázek 4: Dendrogram vytvořený na základě MALDI-TOF MS
profilů. (autor O. Šedo)
Obrázek 5: MALDI destička. (autor O. Šedo)
Systémy identifikace mikroorganismů
15
kvality signálu je přidávána kyselina trifluoroctová (TFA) či mravenčí (FA). Směs je
nanášena na speciální destičku z nerezové oceli či hliníku, která je inertní vůči matrici a
rozpouštědlům. Na této kovové destičce se může analyzovat až 384 izolátů. Po vysušení
vzorků při pokojové teplotě je destička umístěna do hlubokého vakua (10-4 Pa) v iontovém
zdroji hmotnostního spektrometru a proběhne analýza MALDI-TOF MS. Po analýze se
MALDI destička důkladně vyčistí a tím je připravena k dalšímu použití.
2.3.2. Výhody a nevýhody metody MALDI-TOF MS Tato metoda má velké přednosti díky levnému provozu, snadnosti a rychlosti provedení,
vysoké rozlišovací schopnosti, snadné reprodukovatelnosti a možnosti analyzovat velký počet
izolátů najednou. Pro analýzu je potřeba velmi malé množství vzorku. Je aplikovatelná pro
široké spektrum mikroorganismů a obejde se bez předchozí charakteristiky izolátu a jeho
zařazení do určité skupiny.
Limitujícím faktorem této metody je identifikace druhů mikroorganismů, které nejsou
obsaženy v databázi. Avšak hmotnostní spektra nových referenčních kmenů se neustále do
databáze doplňují. Mezi další nevýhody patří vysoké pořizovací náklady přístrojového
vybavení a komplikovaná identifikace některých fylogeneticky příbuzných druhů bakterií či
směsných kultur.
2.3.3. Využití metody MALDI-TOF MS v praxi Hmotnostní spektrometrie MALDI-TOF byla v nedávné době zavedena do rutinní
mikrobiologické diagnostiky. MALDI-TOF MS se začíná úspěšně využívat v různých
oblastech mikrobiologie, nejen v klinické praxi. Metodou lze identifikovat bakterie, kvasinky
a plísně izolované z různých biologických materiálů, potravin i vzorků prostředí. Stává se
klíčovým nástrojem v rámci kontroly bezpečnosti potravin. Mikroorganismy lze identifikovat
nejen do rodu, ale také na druhové úrovni a v některých případech i na úrovni poddruhů.
MALDI-TOF MS lze snadno a rychle identifikovat nejen patogenní bakterie izolované
z potravin, jako jsou E. coli O157:H7, Yersinia spp., Salmonella spp., Campylobacter spp.,
jednotlivé druhy listerií včetně L. monocytogenes, Clostridium spp., Bacillus cereus,
Staphylococcus aureus (včetně rozlišení meticilin rezistentních – MRSA, od meticilin
senzitivních kmenů S. aureus), Enterococcus spp., ale také technologicky významnou avšak
na identifikaci komplikovanou skupinu bakterií mléčného kvašení a další mikroorganismy.
Hmotnostní spektrometrií MALDI-TOF se mohou analyzovat také rekombinantní proteiny,
proteiny faktorů virulence, proteiny sporových stěn a S-vrstvy, enzymy, metabolity,
bakteriociny, atd.
2.4. Biosenzory Biosenzor je analytický přístroj s velmi širokým využitím. Uplatňuje se v humánní i
veterinární medicíně, farmaceutickém a potravinářském průmyslu, zemědělství, životním
prostředí, atd. Pomocí biosenzorů jsou zkoumány interakce protilátka-antigen, DNA-DNA,
DNA-protein, receptor-ligand, protein-ligand a mnoho dalších interagujících dvojic. Studium
biosenzorů spadá do několika vědních oborů – biochemie, biotechnologie, analytická a
fyzikální chemie, elektronika, informatika, nauky o materiálech a biologie.
2.4.1. Základní principy Biosenzor má dvě hlavní části (obrázek 6). Biorekogniční část – citlivý prvek biologického
původu, který má za úkol styk s testovaným vzorkem (analytem). Biorekogniční část je buď
součástí, nebo v těsném kontaktu s fyzikálně-chemickým převodníkem. Ten převádí
biologický nebo biochemický signál na měřitelný a kvantifikovatelný elektrický nebo optický
Systémy identifikace mikroorganismů
16
signál vhodný k dalšímu zpracování.
Signál je přímo úměrný koncentraci
stanovovaných látek ve vzorku.
Samotné měření může probíhat buď
v uzavřené nádobce, v průtočném
systému nebo přímo v kontaktu
s analytem. Měření v uzavřené nádobce je velmi jednoduché a nenáročné na vybavení, avšak
je potřeba manuální obsluha. Průtočné systémy měření mohou být automatizovány. Vzorek
může být v průtočné cele měřen neředěný či ředěný. Pokud se biosenzor nachází přímo ve
sledovaném prostředí (tkáň, krevní řečiště, fermentor), neměla by jeho přítomnost ovlivnit
koncentraci analytu a snížit jeho množství v důsledku měření.
2.4.1.1. Biorekogniční část
Biorekogniční část biosenzoru může být buď biokatalytická nebo bioafinitní. Pokud je analyt
přeměněn v průběhu chemické reakce, obvykle vystupuje jako substrát enzymové reakce,
jedná se o biokatalytický typ (biologickým prvkem je např. enzym, organela, buňka, tkáň,
orgán, mikroorganismus). V případě, že je analyt specificky vázán ve vznikajícím afinitním
komplexu, biorekogniční část je bioafinitní (např. lektin, protilátka, nukleová kyselina,
receptor).
Nejčastěji využívanou biorekogniční složkou je enzym. Ten katalyzuje přeměnu substrátu
v produkt, který se může stát zároveň markerem pro elektrochemický převodník.
Další rozšířenou biorekogniční složkou je protilátka, která je pevně imobilizovaná na povrch
převodníku. Tento typ biosenzoru velmi připomíná techniku ELISA s tím rozdílem, že
vzniklý imunokomplex je detekován pomocí převodníku, nikoliv pomocí kolorimetrické
změny. Velké využití se nabízí u kombinace protilátek s křemíkovým typem převodníků –
tzv. Guartz Crystal Microbalance.
Mikroorganismy patří mezi velmi výhodnou biologickou složku biosenzorů. Používají se
buňky bakterií (např. Bacillus subtilis, B. licheniformis), sinic, kvasinek či plísní. Mohou
nahradit antigeny, enzymy, proteiny (pokud jsou exprimovány na povrchu buňky), což
zjednodušuje a zvyšuje efektivitu výroby biosenzoru.
2.4.1.2. Fyzikálně-chemický převodník
Pro sestavení biosenzorů se používá celá škála fyzikálně-chemických převodníků. Mohou být:
- elektrochemické, tyto nejrozšířenější převodníky měří změnu napětí (amperometrie),
odporu (konduktometrie), proudu (voltametrie) či elektromotorického napětí
(potenciometrie); jsou velmi citlivé, levné a jednoduché ke konstrukci,
- optické převodníky jsou založeny na detekci změn ve světelné adsorpci nebo emisi
během reakce biorekogniční složky analytu, uplatňuje se fotometrie, fluorimetrie,
luminometrie a nelineární optika,
- kalorimetrické převodníky využívají změnu teploty v průběhu enzymatických reakcí,
- akustické,
- piezoelektrické převodníky, kde se využívá křemíkový krystal, který kmitá na určité
frekvenci v závislosti na převedeném napětí. Pokud je změněna hmotnost krystalu (např.
navázáním určitých látek), posouvá se i frekvence oscilace krystalu a z této změny lze
určit nárůst hmotnosti na krystalu. Tento typ převodníků se využívá např. při detekci
Salmonella Typhimurium.
Obrázek 6: Schéma složení biosenzoru.
Systémy identifikace mikroorganismů
17
2.4.2. Výhody a nevýhody biosenzorů Přednostmi biosenzorů jsou vysoká citlivost a selektivita, měření bez speciálních reagencií,
rychlost analýzy, nízké náklady a snadná obsluha. Analýzy probíhají v reálném čase. Častou
výhodou biosenzorů je jejich malá velikost a přenositelnost, tudíž stačí relativně malé
množství vzorku a potřebných chemikálií pro analýzu.
Velkým problémem je sterilizace biosenzorů. Některé typy pracují jen v úzkém rozsahu
koncentrace analytu, mají pomalou odezvu, jsou citlivé na elektrický šum a jejich
miniaturizace může mít vliv na velikost signálu. Při použití enzymů se obvykle liší optimální
provozní pH enzymu od pH prostředí.
2.4.3. Využití biosenzorů v praxi V dnešní době nacházejí biosenzory uplatnění zejména v medicíně. V klinické diagnostice se
používají pro detekci různých metabolických produktů, složek krve, ale i k identifikaci
patogenních mikroorganismů. Snad úplně nejrozšířenějším biosenzorem je glukosový
biosenzor pro kontrolu hladiny glukosy v krvi diabetiků.
Velký potenciál využití biosenzorů je i v potravinářství při kontrole kvality potravin. Pomocí
biosenzorů lze stanovit např. sacharidy, lipidy, vitaminy, ale také alergeny, rezidua
inhibičních látek, antibiotika, růstové stimulátory, pesticidy či methanol. Mezi důležité
aplikace patří detekce alimentárních patogenů jako Salmonella Enteritidis, Salmonella
Typhimurium, Listeria monocytogenes, Campylobacter jejuni, Escherichia coli O157:H7, S.
aureus, dále bakteriálních toxinů (např. stafylokokové enterotoxiny typ A a B), mykotoxinů
(např. aflatoxin) a biogenních aminů (např. histamin, putrescin).
Navzdory velkému rozvoji biosenzorů, jen málo systémů je komerčně vyráběných právě
pro oblast analýzy potravin. Na trhu jsou k dispozici následující biosenzory určené pro
Určené množství pomnoženého vzorku se otvorem v krytu testu napipetuje do tzv. okna pro
vzorek, kde je obvykle papírová vrstva umožňující absorpci vzorku. Pod touto vrstvou je
vrstva skleněných vláken obsahující vysušený konjugát – částice s navázanými specifickými
protilátkami proti cílovému antigenu (mohou to být např. částice zlata nebo barevné latexové
částice). Vrstva s konjugátem je připojena k nitrocelulózové membráně. Po aplikaci vzorku
proniká tekutina kapilární difúzí do vrstvy konjugátu a rehydratuje jej. Současně dochází ke
specifické reakci mezi konjugátem a cílovými antigeny.
Vytvořený komplex antigen-konjugát projde až na membránu a putuje směrem k linii
vazebných proteinů (tzv. vazebná nebo testovací linie). Vazebné proteiny jsou specifické
protilátky proti cílovému antigenu, které migrující komplex zachytí a imobilizují.
V testovacím okně dojde v místě vazebné linie k vytvoření zřetelného pruhu či linky, což
indikuje pozitivní výsledek. V závislosti na rychlosti průniku kapaliny porózní membránou,
trvá tento proces obvykle 15 – 20 minut.
Přebytečný (nenavázaný) konjugát vzlíná dál
membránou a je zachycen na dalším místě
membrány, tzv. kontrolní linii, kde jsou navázány
specifické protilátky proti konjugátu. Jejich
hromadění opět vyvolá vznik barevného proužku.
Tato linie slouží ke kontrole funkčnosti testu.
Pokud nedojde k jejímu zvýraznění, je test
neplatný i když vznikne proužek na testovací linii.
Vyhodnocení testu:
pozitivní výsledek – zřetelný barevný pruh
v místě vazebné i kontrolní linie (2 proužky)
negativní výsledek – pruh je vytvořen pouze
v místě kontrolní linie (1 proužek).
4.2.2. Výhody, nevýhody a využití imunochromatografie v praxi Jedná se o velmi rychlou, jednoduše proveditelnou metodu, která nevyžaduje zvláštní
přístrojové vybavení či školený personál a je proveditelná v provozních podmínkách. Určitým
omezením je požadavek na pomnožení vzorku před vlastním stanovením, což samozřejmě
prodlužuje dobu stanovení. Podobně jako u dalších imunologických technik, je nezbytné
potvrdit pozitivní výsledky konvenční metodou.
V klinické diagnostice jsou imunochromatografické metody rutinně používány. Využívají se
při detekci specifických složek moči, krve a ostatních tělních tekutin, dále hormonů, léčiv,
virů či dalších infekčních agens. V potravinářské mikrobiologii nachází své uplatnění při
detekci reziduí antibiotik, hormonů a alimentárních patogenů. Nečastěji se
imunochromatografická metoda používá k detekci Salmonella spp., E. coli O157 a
Shigatoxinů ST1 a ST2, L. monocytogenes či Campylobacter spp., a to v různých potravinách
či environmentálních vzorcích. Komerční soupravy dodává na trh řada firem, např. Merck
KGaA (řada Singlepath či Duopath), Oxoid Ltd, BioControl Inc. (řada VIP), Neogen Corp
(řada Reveal) či Celsis Ltd (řada Path-Stick). Některé testy byly validovány organizací
AOAC (Association of Official Chemists).
Obrázek 9: Výsledek testu.
Imunologické metody
28
4.3. Enzyme-Linked Immunosorbent Assay – ELISA ELISA (Enzyme-Linked Immunosorbent Assay), známá také jako EIA (Enzyme
Immunoassay), je široce používaná imunologická technika nejen pro detekci patogenních
mikroorganismů. Je alternativou ke kroku detekce či izolace mikroorganismů na živném
agaru. Je relativně jednoduchá, může být i semi-automatická, výsledky poskytuje dostatečně
rychle. Podle zaměření umožňuje detekovat konkrétní rod, druh či serotyp. Na druhou stranu
je to stále metoda screeningová, pozitivní výsledky musí být potvrzeny klasickým způsobem.
4.3.1. Základní principy Jako všechny imunologické metody, je i ELISA založena na principu specifické vazebné
reakce mezi antigenem a protilátkou. V ELISA testech se používají monoklonální nebo
polyklonální protilátky. Polyklonální protilátky jsou produkovány při imunitní odpovědi
hostitele na infekci organismu relativně velkou molekulou (protein, mikroorganismus), jsou
produktem mnoha aktivovaných klonů B lymfocytů, a proto jsou namířeny proti více
epitopům určitého antigenu. Monoklonální protilátky jsou produkované jedním klonem B-
lymfocytů po styku s jedním konkrétním antigenním epitopem, připravují se na tkáňových
kulturách.
Ke značení protilátek se v ELISA metodě používá enzym, obvykle schopný katalyzovat
přeměnu bezbarvého substrátu na barevný produkt (případně může dojít i ke změně barvy
substrátu). Díky tomu je hodnocení testu vizuální nebo spektrofotometrické. Nejčastěji se
využívá enzym alkalická fosfatasa reagující s para-nitrofenyl fosfátem nebo křenová
peroxidasa reagující s tetramethylbenzidinem. V obou případech vzniká barvený produkt.
Reakce probíhající v průběhu ELISA testu vždy vyžadují nějaký pevný nosič, na který jsou
kovalentně vázány či adsorbovány protilátky, příp. antigen. Nejčastěji se jedná o mikrotitrační
destičky či mikrozkumavky, lze použít i papírové membrány nebo polystyrenové tyčinky.
4.3.2. Konfigurace ELISA metod používaných v mikrobiologii potravin V mikrobiologii potravin je detekce obvykle zaměřena na průkaz antigenu (bakterie, toxin)
pomocí známých protilátek. Tomu odpovídají i typy ELISA metod, které se používají.
Prvním krokem je vždy pokrytí pevného nosiče specifickými protilátkami nebo antigeny.
V případě, že test používá statický nosič (např. mikrotitrační destičku),jsou vzorek a další
reagencie v několika fázích postupně přidávány do jamek. Při použití polystyrenových
tyčinek (tzv. dipstick) je pevný nosič navržen tak, aby byl přenosný. Antigeny navázané na
pevném nosiči (tyčince) mohou být přenášeny do různých živných médií (podpora růstu a
namnožení cílových buněk), po přenesení do roztoku substrátu dojde k barevné reakci.
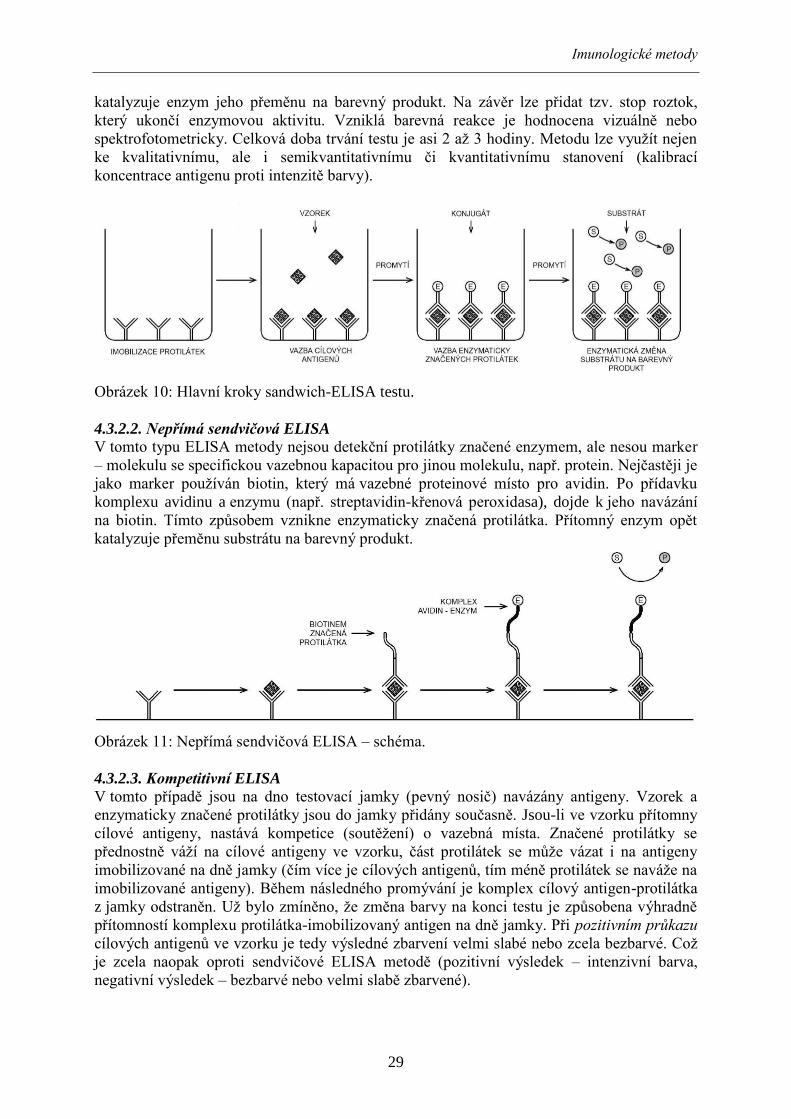
4.3.2.1. Sendvičová ELISA
Nejjednodušším a nejčastěji používaným formátem v komerčních testech používaných při
vyšetření potravin je sendvičová (angl. sandwich) ELISA. „Sendvič“ v tomto případě
znamená, že je cílový antigen obalen dvěma protilátkami. V průběhu testu je antigen nejprve
zachycen a poté detekován. Prvním krokem každého sendvičového ELISA testu je pomnožení
vyšetřované potraviny.
Vazebné (imobilizované) protilátky, specifické pro cílový antigen, jsou navázány na povrch
pevného nosiče, např. jamky mikrotitrační destičky. Po přídavku pomnoženého vzorku
potraviny dojde, v případě přítomnosti cílových antigenů, k jejich vazbě na protilátky. Po
promytí, kterým se odstraní zbytky vzorku, následuje přídavek detekčních, enzymaticky
značených protilátek. Detekční protilátky se váží na cílové antigeny a vytvoří se „sendvič“.
Několikanásobným promytím se odstraní nenavázané protilátky. Po přídavku substrátu
Imunologické metody
29
katalyzuje enzym jeho přeměnu na barevný produkt. Na závěr lze přidat tzv. stop roztok,
který ukončí enzymovou aktivitu. Vzniklá barevná reakce je hodnocena vizuálně nebo
spektrofotometricky. Celková doba trvání testu je asi 2 až 3 hodiny. Metodu lze využít nejen
ke kvalitativnímu, ale i semikvantitativnímu či kvantitativnímu stanovení (kalibrací
koncentrace antigenu proti intenzitě barvy).
Obrázek 10: Hlavní kroky sandwich-ELISA testu.
4.3.2.2. Nepřímá sendvičová ELISA
V tomto typu ELISA metody nejsou detekční protilátky značené enzymem, ale nesou marker
– molekulu se specifickou vazebnou kapacitou pro jinou molekulu, např. protein. Nejčastěji je
jako marker používán biotin, který má vazebné proteinové místo pro avidin. Po přídavku
komplexu avidinu a enzymu (např. streptavidin-křenová peroxidasa), dojde k jeho navázání
na biotin. Tímto způsobem vznikne enzymaticky značená protilátka. Přítomný enzym opět
katalyzuje přeměnu substrátu na barevný produkt.
Obrázek 11: Nepřímá sendvičová ELISA – schéma.
4.3.2.3. Kompetitivní ELISA
V tomto případě jsou na dno testovací jamky (pevný nosič) navázány antigeny. Vzorek a
enzymaticky značené protilátky jsou do jamky přidány současně. Jsou-li ve vzorku přítomny
cílové antigeny, nastává kompetice (soutěžení) o vazebná místa. Značené protilátky se
přednostně váží na cílové antigeny ve vzorku, část protilátek se může vázat i na antigeny
imobilizované na dně jamky (čím více je cílových antigenů, tím méně protilátek se naváže na
imobilizované antigeny). Během následného promývání je komplex cílový antigen-protilátka
z jamky odstraněn. Už bylo zmíněno, že změna barvy na konci testu je způsobena výhradně
přítomností komplexu protilátka-imobilizovaný antigen na dně jamky. Při pozitivním průkazu
cílových antigenů ve vzorku je tedy výsledné zbarvení velmi slabé nebo zcela bezbarvé. Což
je zcela naopak oproti sendvičové ELISA metodě (pozitivní výsledek – intenzivní barva,
negativní výsledek – bezbarvé nebo velmi slabě zbarvené).
separace, ultracentrifugace nebo filtrace elektricky nabitými filtry) a ošetření proteinasou K.
Po zakoncentrování virových částic je potřeba uvolnit nukleovou kyselinu z virových obalů,
odstranit nežádoucí zbytky hrubých lyzátů a nukleové kyseliny zakoncentrovat. Pro narušení
virových obalů lze použít specifické enzymy či detergenty (SDS, laurylsulfát sodný),
sonifikaci ultrazvukem nebo speciální přístroj „bead beater“, který protřepává biologický
materiál se skleněnými či zirkonovými kuličkami, které mechanicky narušují virový obal. Pro
alimentární viry, které nejsou obalené a mají pouze proteinovou kapsidu, stačí působení
fenolu (případně ve směsi s chloroformem). Nežádoucí proteiny z hrubého lyzátu se
odstraňují proteinasami, RNA (v případě izolace DNA) RNasami a DNA (v případě izolace
RNA) DNasami. Manipulaci s RNA je nutno provádět v ledové lázni, používat ochranné
rukavice a pracovat za aseptických podmínek.
Pro přímou extrakci nukleové kyseliny z potravinové matrice lze použít guanidin
isothiokyanát (GITC), který má destruktivní účinek na všechny komponenty vzorku kromě
nukleových kyselin. V potravinách živočišného původu se pro uvolnění nukleové kyseliny
z tkáně dá použít také přístroj „bead beater“. Poté následuje extrakce fenolem nebo
ultracentrifugace v chloridu cesném a precipitace nukleové kyseliny ethanolem či
izopropylalkoholem (u RNA v přítomnosti LiCl).
Existují komerční soupravy (silikátové kolonky, chromatografické systémy, atd.), které mají
výhodu snadného provedení, avšak jsou finančně nákladnější a hlavně nejsou vhodné pro
každou izolaci virové DNA či RNA.
5.2. Polymerázová řetězová reakce (PCR) Základem pro mnohé molekulárně biologické metody je polymerázová řetězová reakce
(Polymerase Chain Reaction). Pomocí této techniky může být za podmínek in vitro rychle a
vysoce selektivně namnožena konkrétní nukleotidová sekvence obsažená v DNA.
Problematika polymerázové řetězové reakce, včetně detekce PCR produktů, je podrobně studována v rámci
předmětu Mikrobiologie potravin a mikrobiologické laboratorní metody (viz skripta Mikrobiologie potravin –
praktická cvičení I. Obecná mikrobiologie).
5.2.1. Základní principy Principem PCR je opakované kopírování (amplifikace) templátové (vzorové) molekuly DNA
pomocí enzymu DNA-polymerasy. Syntéza DNA je řízena dvěma krátkými oligonukleotidy
(primery), které se komplementárně párují s templátovou DNA na počátku a na konci
amplifikovaného fragmentu, každý s jiným vláknem původní dvouřetězcové molekuly DNA.
V průběhu polymerázové řetězové reakce se opakovaně střídají 3 kroky. Prvním krokem je
denaturace, kdy obvykle při 95 °C dojde k uvolnění vodíkových můstků a z dvouřetězcové
DNA (dsDNA) vzniknou dvě jednořetězcové DNA (ssDNA). Následuje annealing – připojení (hybridizace) primerů k templátové ssDNA. Tím dojde k ohraničení cílové
Metody molekulární biologie
37
sekvence, která bude amplifikována. Tento krok probíhá při teplotě 50 až 65 °C. Ta závisí na
bodu tání primerů (Tm), jejich délce a nukleotidové sekvenci. U tohoto kroku je nutné provést
optimalizaci. Poté následuje poslední krok, syntéza nového vlákna DNA (vznik dsDNA), tzv.
elongace. Probíhá obvykle při teplotě 72 °C a kromě DNA-polymerasy jsou nutné všechny
čtyři 2´-deoxyribonukleosid-5´-trifosfáty (dNTPs – adenin, guanin, cytosin a thymin).
Přesné hodnoty teploty a dobu trvání jednotlivých kroků je třeba optimalizovat. Tyto kroky se
25 až 35 cyklicky opakují. Optimální počet cyklů je závislý na výchozí koncentraci
templátové DNA. Nově vytvořená DNA se v dalším cyklu stává matricí (templátovou DNA).
Celý proces amplifikace probíhá v přístroji termocykler, v němž se teplota mění automaticky
v naprogramovaných časových intervalech.
Aby byl zabezpečen správný průběh PCR, je potřeba připravit reakční směs (master mix)
obsahující templátovou DNA, primery, dNTPs, reakční pufr, hořečnaté kationty, PCR H2O a
DNA-polymerasu (nejčastěji se používá termostabilní Taq polymerasa izolovaná z termofilní
bakterie Thermus aquaticus). Výsledným produktem polymerázové řetězové reakce jsou
amplikony, úseky DNA s definovanou délkou, o velikosti obvykle desítky až tisíce párů bazí
(bp). Koncentrace a množství jednotlivých komponent reakční směsi hraje rozhodující roli při
tvorbě PCR produktu a stanovuje se empiricky.
Správný průběh reakce se sleduje systémem kontrol. Ty nám mohou indikovat např.
kontaminaci PCR komponent (negativní kontrola) nebo funkčnost všech složek reakce
(pozitivní kontrola). Aby nedocházelo ke křížovým kontaminacím, je potřeba mít od sebe
oddělené prostory pro izolaci DNA, přípravu reakční směsi (nejlépe laminární box) a
manipulaci s PCR produkty, včetně samostatných pomůcek pro každou z těchto činností. Tyto
místnosti by se měly pravidelně dekontaminovat UV světlem.
Pro rutinní provádění polymerázové řetězové reakce jsou dostupné komerčně předpřipravené
PCR master mixy (např. od firmy Top-Bio, s.r.o. či Qiagen, GmbH) obsahující DNA-
polymerasu, reakční pufr, MgCl2, barvivo a aditiva potřebná pro detekci amplikonů
agarózovou gelovou elektroforézou.
5.2.2. Detekce PCR produktů K analýze produktů polymerázové řetězové reakce mohou být použity různé metody. Např.
hybridizace, ELISA nebo sekvenční analýza. Avšak snadná, levná a asi nejužívanější je
separace amplikonů pomocí agarózové gelové elektroforézy (ELFO). PCR produkty migrují
agarózovým gelem na základě jejich záporného náboje od katody ke kladně nabité anodě,
rychlost pohybu molekul DNA je nepřímo úměrná logaritmu jejich velikosti. Amplikony jsou
obarveny např. ethidium bromidem (barvivo interkalující se mezi vlákna dsDNA).
V ultrafialovém světle obarvený fragment DNA oranžově fluoreskuje. Bezpečnější
alternativou ke karcinogenní, mutagenní a teratogenní látce ethidium bromid je barvení PCR
produktů pomocí látky Midori Green, která emituje zelenou fluorescenci.
Podle velikosti amplifikovaného fragmentu, určené porovnáním s markerem, tj. velikostním
standardem obsahujícím různě dlouhé fragmenty DNA, usuzujeme na přítomnost
specifického genu ve vzorku.
5.2.3. Modifikace PCR Polymerázová řetězová reakce je používána v široké škále variant, které jsou upraveny podle
účelu analýzy. U mnohonásobné multiplex PCR je možná detekce více genů v jedné reakci
současně. Podle počtu požadovaných amplikonů, obsahuje master mix počet párů primerů.
Primery musí být navrženy tak, aby se výsledné PCR produkty daly odlišit svou velikostí
nebo byly značeny různými fluorofory (u real-time PCR). Tato varianta PCR je výhodnější
Metody molekulární biologie
38
než provedení samostatných amplifikací, zejména z hlediska časového i ekonomického
(ušetření komponent reakční směsi). Avšak optimalizace metody je náročnější než s jednou
sadou primerů. Obecně platí, že jsou amplifikovány maximálně 4 různé cílové sekvence pro
zajištění adekvátní citlivosti, specifičnosti a snadné interpretace dat. Využívá se často pro
stanovení několika bakteriálních druhů současně (např. Campylobacter jejuni a C. coli) nebo
při detekci genů kódujících stafylokokové enterotoxiny či genů rezistence.
Odstupňovaná PCR (nested PCR) neboli PCR využívající vnějších a vnitřních primerů.
Tento typ PCR probíhá ve dvou krocích a vyznačuje se vysokou citlivostí. V prvním kroku
dochází k amplifikaci za účasti prvního, vnějšího páru primerů, poté se PCR produkt stává
matricí další polymerázové řetězové reakce a amplifikuje s druhým, vnitřním párem primerů.
Zpětná neboli reverzní PCR (RT-PCR) je metoda určená k amplifikaci molekul RNA. RNA
je nejprve přepsána na cDNA (komplementární DNA) enzymem zpětná transkriptasa. Tento
enzym je termolabilní (již nad 40 °C ztrácí svoji funkčnost), stringence (podmínky ovlivňující
sílu a specifitu hybridizace) reakce je poměrně nízká (dochází k nekomplementárnímu
párování bazí a transkripce tak není dokonalá). Proto se nyní využívá enzym Tth DNA-
polymerasa (RNA-dependentní DNA-polymerasa), která umožňuje nejen přepis RNA do
DNA při teplotě 72 °C, ale i vlastní amplifikaci a tvorbu PCR produktu.
Speciálním typem polymerázové řetězové reakce je in situ-PCR. Při této metodě probíhá
amplifikace specifické sekvence nukleové kyseliny přímo v buňce či tkáni. Postup zahrnuje
šetrnou fixaci buněk k podložnímu sklu a nastavení podmínek pro permeabilitu nezbytných
PCR reagencií do buňky. Detekce probíhá pomocí in situ hybridizace nebo imunochemicky.
Jak prokaryotické, tak eukaryotické genomy obsahují
repetitivní (opakující se) sekvence. Mezi těmito dlouhými
repetitivními elementy se v genomu prokaryot nacházejí
také relativně krátké nekódující sekvence. Při
interrepetitivní (repetitivní) PCR (REP-PCR) dochází
právě k amplifikaci těchto sekvencí oddělujících
v genomu různé typy repeticí. Tímto typem PCR se získá
soubor různě dlouhých amplikonů charakteristický pro
daný mikroorganismus (obrázek 18). Profil genomu
konkrétního mikroorganismu zobrazený spektrem PCR
produktů (případně restrikčních fragmentů) se nazývá
fingerprint. Pomocí počítačového softwaru se amplikony
jednotlivých izolátů porovnávají s typovými kmeny,
vytvoří se dendrogram a izoláty se identifikují. Touto
metodou za použití různých primerů (např. (GTG)5) lze
rozlišit některé druhy bakterií mléčného kvašení, např. při
studiu diversity startovacích a doplňkových kultur. Velké
využití má tato metoda jako typizační technika při
genotypizaci izolátů mikroorganismů.
Další fingerprintovou metodou založenou na PCR, pomocí níž lze zobrazit profil DNA
konkrétního organismu, je náhodná PCR (RAPD). Tato rychlá a jednoduchá metoda se
používá k identifikaci a typizaci některých druhů bakterií. U RAPD se krátký
oligonukleotidový primer náhodně váže k templátové DNA a vzniká soubor amplikonů
charakteristický pro daný mikroorganismus (podobně jako u rep-PCR). Pomocí agarózové
gelové elektroforézy jsou detekovány amplifikované fragmenty různé velikosti a tím se
vytvoří specifický fingerprint pro testovaný mikroorganismus.
Obrázek 18: Agarózová gelová
elektroforéza amplikonů vzniklých
pomocí rep-PCR. (U izolátů v běhu
č. 3, 5 a 6 se vytvořily stejné
amplikony, jedná se o stejný
bakteriální druh).
Metody molekulární biologie
39
5.2.3.1. Real-time PCR (qPCR)
Dnes velmi využívaným typem polymerázové řetězové reakce je real-time PCR (qPCR), která
umožňuje přímou detekci a kvantifikaci PCR produktu „v reálném čase“ (v průběhu celé PCR
reakce), nikoliv až po jeho skončení. Při stanovení PCR produktů se využívá sledování
fluorescenčního signálu. Jeho intenzita je permanentně snímána a analyzována speciálním
přístrojem, ve kterém zároveň probíhá PCR, tudíž se amplikony nemusí detekovat
elektroforeticky. Tím je tato technika zjednodušena a urychlena. Stanovení množství molekul
nukleových kyselin ve vzorku se využívá při studiu detekce patogenních mikroorganismů a
virů a také exprese genů (zejména studium transkripce – zda je a v jaké míře daný gen
přepisován z DNA do mRNA).
Předností metody qPCR je zejména rychlost bez nutnosti detekce PCR produktu (výsledek do
několika hodin), jednoduchost provedení, přesnost a citlivost.
K detekci vznikajícího PCR produktu lze využít fluorescenční barviva, fluorescenčně značené
hybridizační sondy nebo fluorescenčně značené primery. V prvním případě se používají
barviva (např. SYBR® Green I), která fluoreskují po vazbě na dsDNA. Fluorescenční signál
se zvyšuje se vzrůstajícím množstvím PCR produktu. Nevýhodou fluorescenčního
kyaninového barviva SYBR® Green I je to, že nelze
rozlišit PCR produkt od nespecificky
naamplifikovaných sekvencí a nedá se použít u
multiplex PCR. Druhý způsob detekce amplikonů je
prostřednictvím fluorescenčně značených
hybridizačních sond (krátké oligonukleotidy), které
se komplementárně vážou na vnitřní část
amplifikované sekvence. Existuje celá řada těchto
sond, které mohou obsahovat kromě různých typů
fluoroforů (např. FAM), také zhášeč (např.
TAMRA). U sondy TaqManTM je možné fluorescenci
snímat až po ukončení zhášení, kdy se zhášeč oddálí
od fluoroforu, a to rozložením sondy pomocí enzymu
Taq DNA-polymerasy při elongaci (obrázek 19).
Mezi další typy sond patří tzv. molekulární majáky.
Je možné také využít přenosu energie fluorescenční
rezonance – technologie FRET (Fluorescence
Resonance Energy Transfer). Třetí způsob detekce
amplionů při qPCR je pomocí fluorescenčně
značených primerů, např. technologie AmpliFluorTM
nebo LUX.
U klasické PCR je velmi obtížná kvantifikace. Korelace mezi počátečním množstvím
templátu a konečným množstvím PCR produktu (i přes známý počet cyklů) není spolehlivá.
Spolehlivější je vztah mezi množstvím templátu a počtem amplifikačních cyklů nutných pro
vytvoření detekovatelného PCR produktu (čím více templátu ve vzorku, tím dříve je možné
PCR produkt detekovat). Cyklus, ve kterém je již PCR produkt detekovatelný se nazývá jako
prahový cyklus CT (treshold cycle). A právě tato kvantifikace může být prováděna metodou
qPCR, která zaznamenává vznik amplikonů v průběhu celé reakce. Existují dva způsoby
kvantifikace. Relativní kvantifikace, při které dochází pouze ke srovnání dvou vzorků a
získání informace, ve kterém vzorku a kolikrát je více templátu. Při „absolutní“ kvantifikaci
je množství templátu vztaženo k externímu templátu a kvantifikováno přesné množství.
V tomto případě je nutné ze standardů sestrojit kalibrační křivku vyjadřující závislost CT na
známém množství DNA standardního vzorku, ze které se určuje počet kopií DNA v templátu.
Obrázek 19: Technologie TaqManTM.
Metody molekulární biologie
40
5.2.4. Inhibitory polymerázové řetězové reakce Teoreticky umožňuje technika PCR detekovat jednu jedinou buňku či molekulu DNA. Avšak
správný průběh reakce může být narušen přítomností celé řady inhibitorů. Zejména
potravinová matrice se vyznačuje jejich velkým množstvím.
Za snížení citlivosti či úplné selhání reakce mohou být zodpovědné bakteriální enzymy
(nukleasy, proteasy), polysacharidy, proteiny, lipidy a další složky potravin (např. ionty Ca2+
u mléčných výrobků, myoglobin u masných výrobků), pudr z laboratorních rukavic, složky
kultivačních médií, detergenty, antibiotika, aj.
Již při izolaci nukleové kyseliny může dojít k selhání buněčné lýzy nebo degradaci
templátové DNA či RNA. Poškozeny mohou být také primery (zejména ty s nižší teplotou
tání), případně může dojít k obsazení jejich cílových míst. Termostabilní DNA-polymerasu
mohou inhibitory inaktivovat. K inhibici dojde snáze, pokud je nízká koncentrace templátové
nukleové kyseliny a pokud se tvoří dlouhý amplikon nebo amplikon s nižším obsahem G+C
bazí. U real-time PCR může být inhibován také fluorescenční signál. Tyto nežádoucí vlivy se
mohou projevit jako falešně negativní výsledek.
5.2.5. Výhody, nevýhody a využití metody PCR v praxi Polymerázová řetězová reakce je nejpoužívanější amplifikační technika. Má však i svá
omezení. Její citlivost je tak vysoká (umožňující detekci jediné buňky), že u
kontaminovaných vzorků se mohou vyskytnout falešně pozitivní reakce. Naopak
v přítomnosti kontaminujících látek, které PCR inhibují, může dojít k falešně negativní
reakci.
V potravinářské mikrobiologii se polymerázová řetězová reakce využívá jednak k rychlé
detekci alimentárních patogenů přímo ze vzorku, jako jsou Salmonella spp., Listeria
monocytogenes, Shigella spp., Campylobacter jejuni/coli, patogenní serotypy E. coli, Vibrio
maso a masné výrobky, mořské plody, nápoje), testování vody, krmiv, environmentálních
vzorků či produktů kosmetického průmyslu.
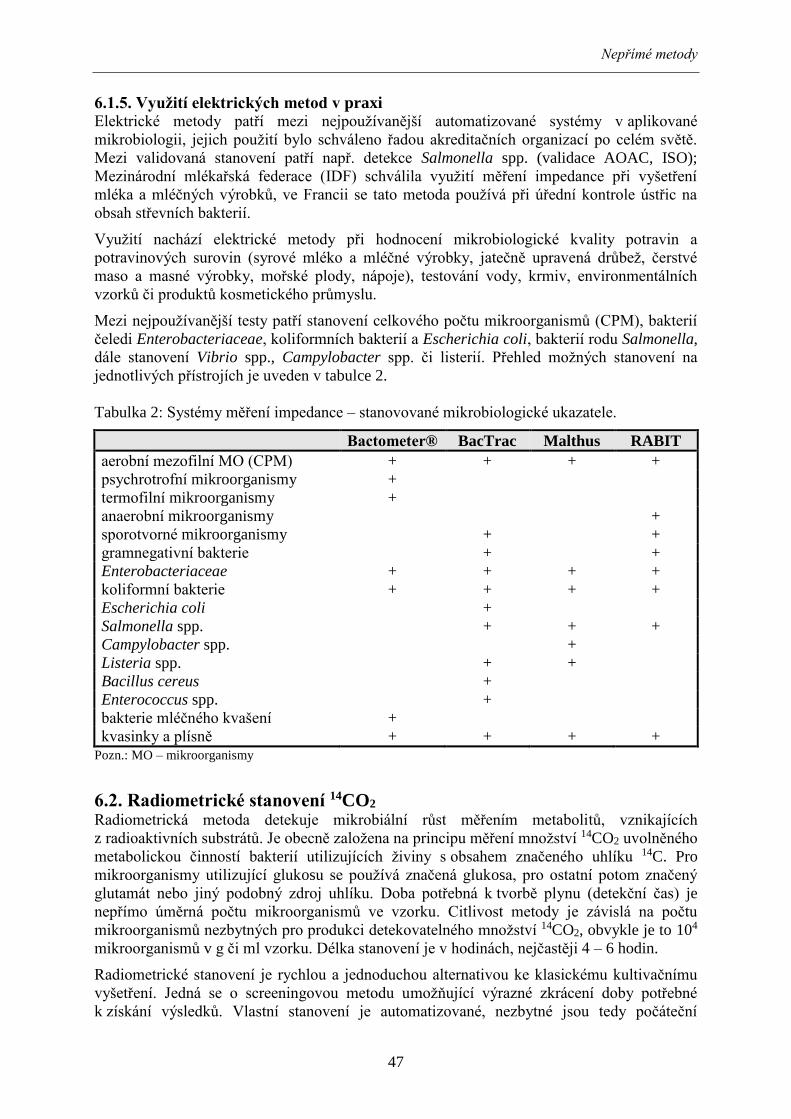
Mezi nejpoužívanější testy patří stanovení celkového počtu mikroorganismů (CPM), bakterií
čeledi Enterobacteriaceae, koliformních bakterií a Escherichia coli, bakterií rodu Salmonella,
dále stanovení Vibrio spp., Campylobacter spp. či listerií. Přehled možných stanovení na
jednotlivých přístrojích je uveden v tabulce 2.
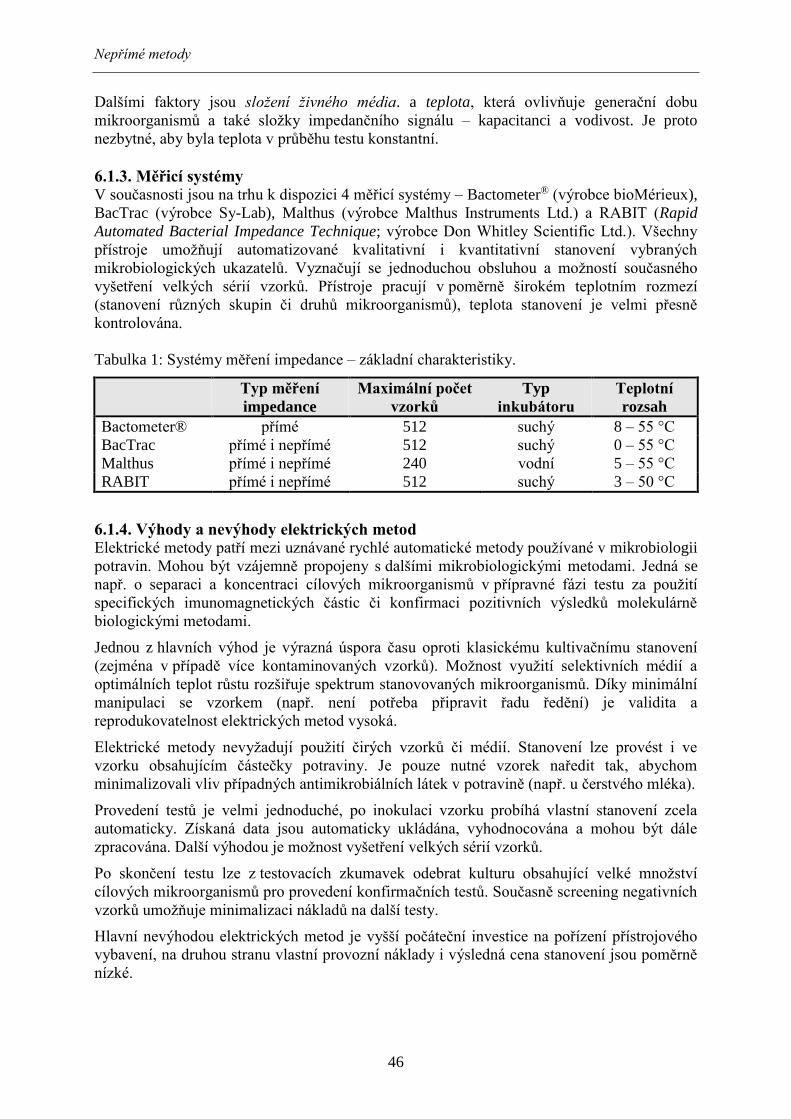
Tabulka 2: Systémy měření impedance – stanovované mikrobiologické ukazatele.
Bactometer® BacTrac Malthus RABIT
aerobní mezofilní MO (CPM) + + + +
psychrotrofní mikroorganismy +
termofilní mikroorganismy +
anaerobní mikroorganismy +
sporotvorné mikroorganismy + +
gramnegativní bakterie + +
Enterobacteriaceae + + + +
koliformní bakterie + + + +
Escherichia coli +
Salmonella spp. + + +
Campylobacter spp. +
Listeria spp. + +
Bacillus cereus +
Enterococcus spp. +
bakterie mléčného kvašení +
kvasinky a plísně + + + + Pozn.: MO – mikroorganismy
6.2. Radiometrické stanovení 14CO2 Radiometrická metoda detekuje mikrobiální růst měřením metabolitů, vznikajících
z radioaktivních substrátů. Je obecně založena na principu měření množství 14CO2 uvolněného
metabolickou činností bakterií utilizujících živiny s obsahem značeného uhlíku 14C. Pro
mikroorganismy utilizující glukosu se používá značená glukosa, pro ostatní potom značený
glutamát nebo jiný podobný zdroj uhlíku. Doba potřebná k tvorbě plynu (detekční čas) je
nepřímo úměrná počtu mikroorganismů ve vzorku. Citlivost metody je závislá na počtu
mikroorganismů nezbytných pro produkci detekovatelného množství 14CO2, obvykle je to 104
mikroorganismů v g či ml vzorku. Délka stanovení je v hodinách, nejčastěji 4 – 6 hodin.
Radiometrické stanovení je rychlou a jednoduchou alternativou ke klasickému kultivačnímu
vyšetření. Jedná se o screeningovou metodu umožňující výrazné zkrácení doby potřebné
k získání výsledků. Vlastní stanovení je automatizované, nezbytné jsou tedy počáteční
Nepřímé metody
48
náklady pro pořízení potřebného přístrojového vybavení. Na druhou stranu, používání
radioaktivních chemikálií je omezeno přísnými předpisy, což nedovoluje širší využití této
metody.
V potravinářství se tato metoda používá při mikrobiologickém hodnocení mražených
potravin, tepelně ošetřených potravin (např. masných výrobků) či ovocných šťáv. V klinické
mikrobiologii se radiometrické stanovení využívá např. k detekci Mycobacterium tuberculosis
v klinických vzorcích (BACTEC 460TB System, výrobce Becton, Dickinson and Company).
6.3. Limulus Amebocyte Lysate test – LAL test V 50. letech minulého století bylo zjištěno, že lyzát amebocytů z hemolymfy členovce
ostrorepa amerického (Limulus polyphenus) se sráží v přítomnosti endotoxinu
gramnegativních bakterií. Bakteriální endotoxin (lipopolysacharid) je součástí buněčné stěny
gramnegativních bakterií a je uvolňován do okolního prostředí po smrti a lýze bakteriální
buňky. Toto zjištění vedlo následně k vývoji in vitro testu pro detekci pyrogenů
kontaminujících přípravky určené k injekčnímu podávání. Od roku 1972 se v lékařském
prostředí začal LAL test masivně používat. Současné medicínské využití spočívá zejména v
detekci gramnegativních bakterií při meningitidách či infekcích močového traktu.
6.3.1. Detekce pozitivní reakce Klasickým způsobem detekce pozitivní reakce je vizuální posouzení tvorby gelu, jako je tomu
u zkumavkového testu.
Další možností je spektrofotometrické či turbidimetrické vyhodnocení, kdy se hodnotí změny
v optické denzitě. Příkladem tohoto typu testu je modifikace automatizovaného systému MS-2
(výrobce Abbott Laboratories), tzv. MS-2 LAL, který se využívá např. při vyšetření moči.
K vyhodnocení testu lze využít i chromogenní substrát. Vzniklá barevná reakce se odečítá
spektrofotometricky, intenzita barvy je přímo úměrná množství endotoxinu. Test probíhá
v mikrotitračních destičkách, pomocí speciálního readeru je vzniklá barevná změna
hodnocena v pravidelných časových intervalech po celou dobu trvání testu (celkem 241
měření za 60 minut). Koncentrace endotoxinu ve vzorcích jsou vypočteny automaticky na
základě standardní křivky odvozené z naměřených hodnot. Test je vysoce citlivý.
6.3.2. Zkumavkový LAL test K provedení zkumavkového testu se používá standardizovaný lyzát (obvykle 0,1 ml na jeden
test). Citlivost činidla se nejdříve testuje za použití standardizovaného endotoxinu. Poté se
stanoví obsah endotoxinu ve vzorcích. Ty se naředí a 0,1 ml každého ředění se přidá do
zkumavky s LAL činidlem. Zaočkované zkumavky jsou po promíchání inkubovány 1 hodinu
ve vodní lázni při teplotě 37 °C. Pozitivní reakcí je vznik pevného gelu, který zůstává ve
zkumavce i po jejím otočení o 180°. Jako titr se označí nejvyšší ředění s pozitivním
výsledkem. Ke stanovení množství endotoxinu v titru, se titr násobí předem stanovenou
citlivostí LAL činidla zjištěnou při reakci se standardizovaným endotoxinem. Počet
gramnegativních bakterií ve vzorku lze odhadnout na základě výpočtu počtu buněk, který
odpovídá zjištěnému množství endotoxinu.
6.3.3. Využití LAL testu v praxi Mimo široké využití ve farmakologii a klinické mikrobiologii, lze LAL test použít také při
mikrobiologickém hodnocení potravin. Metoda je dobře použitelná při screeningu mikrobiální
kontaminace. Například při vyšetření čerstvého masa indikuje nízký obsah endotoxinu maso
výborné mikrobiální kvality (vztaženo k obsahu gramnegativních bakterií – nejvýznamnější
Nepřímé metody
49
skupině mikroorganismů v čerstvém mase). Na druhou stranu vysoký obsah endotoxinu,
zjištěný metodou LAL, nemusí vždy korespondovat s vysokou hodnotou CPM zjištěnou
plotnovou metodou. A to třeba v případě, kdy došlo k destrukci gramnegativních bakterií
záhřevem či účinkem bakteriofágů, nebo když je přítomný endotoxin reprezentován
gramnegativními bakteriemi, které nerostou za podmínek stanovení CPM.
Protože v čerstvých potravinách zpracovaných za standardních podmínek existuje více méně
konstantní poměr gramnegativních a grampozitivních bakterií, lze výsledky LAL testu využít
nejen k odhadu celkového počtu mikroorganismů, ale i počtu bakterií grampozitivních.
Podobným způsobem lze odhadnout i počet plísní. Na druhou stranu LAL test nelze využít
k odhadnutí „celkového“ počtu mikroorganismů v tepelně ošetřených výrobcích, protože
gramnegativní bakterie jsou mnohem citlivější k záhřevu než bakterie grampozitivní.
LAL test lze využít při hodnocení mikrobiální kvality syrového a pasterovaného mléka,
mléčných výrobků, masa a masných výrobků, ryb, atd. Endotoxiny jsou termostabilní, proto
lze u tepelně opracovaných výrobků zpětně posoudit výchozí kontaminaci suroviny.
Výhodou LAL testu je především rychlost a jednoduchost provedení, dostupnost komerčně
připraveného lyzátu či kompletních testovacích souprav a vysoká citlivost (detekuje ng
endotoxinu, přítomného u gramnegativních bakterií). Na druhou stranu nelze rozlišit
přítomnost živých či mrtvých buněk.
6.4. ATP bioluminiscenční test
6.4.1. Základní principy Každá živá buňka obsahuje ATP (adenosintrifosfát), který je univerzálním donorem energie
pro její metabolické reakce. Množství ATP v buňkách je relativně konstantní, a proto může
intracelulární koncentrace ATP sloužit ke stanovení biomasy nebo počtu životaschopných
buněk. Po smrti buňky obsah ATP v důsledku autolytických procesů prudce klesá, veškeré
ATP je rozloženo během několika minut.
ATP bioluminiscenční metoda je založena na principu enzymové reakce mezi luciferinem
(substrát) a luciferasou (enzym). Enzym luciferasa, přirozeně se vyskytující u světlušek
(Photuris pyralis), katalyzuje za přítomnosti luciferinu, kyslíku a hořečnatých iontů přeměnu
chemické energie ATP ve světlo pomocí oxidačně-redukční reakce (obrázek 23).
Obrázek 23: Schéma ATP bioluminiscenční reakce. (McMeekin, 2003 – upraveno)
Luciferin je přeměňován na oxiluciferin, ATP na AMP (adenosinmonofosfát) za uvolnění
pyrofosfátu (PPi), CO2 a současné emise světla o vlnové délce v rozmezí 470 – 700 nm
(vrchol při 562 nm). Emitované světlo je měřeno pomocí luminometru, jeho množství se
stanovuje v relativních světelných jednotkách (RLU; Relative Light Units). Dostupné
luminometry jsou schopné detekovat méně než 0,1 pg ATP v kyvetě, což odpovídá počtu 102
bakteriálních buněk.
Nepřímé metody
50
Množství produkovaného světla je přímo úměrné koncentraci ATP vstupujícího do reakce a
souvisí s počtem metabolicky aktivních buněk ve vzorku. Metodu lze tedy využít i ke
stanovení celkového počtu mikroorganismů ve vzorku, protože mezi intracelulárním obsahem
ATP a počtem KTJ bakterií či kvasinek je lineární vztah. I když lze teoreticky při použití
vhodné techniky a činidel o vysoké čistotě stanovit množství ATP odpovídající přibližně stu
mikrobiálních buněk, v praxi je to obvykle množství odpovídající asi 103 – 104 KTJ.g-1.
Široké použití má při hodnocení čistoty povrchů přicházejících do kontaktu s potravinami.
6.4.2. Stanovení počtu mikroorganismů v potravinách ATP bioluminiscenční test byl původně vyvinut k odhadu celkového mikrobiálního zatížení
potravin. V potravinách je ATP přítomno v somatických buňkách, jako volné ATP nebo
v mikroorganismech. Při stanovení počtu mikroorganismů musí být mikrobiální ATP nejprve
separováno od ostatních zdrojů.
Původní techniky spoléhaly na selektivní lýzu somatických buněk neionogenním detergentem
(např. Triton X-100) následovanou enzymatickou či chemickou destrukcí uvolněného ATP.
Zbylý mikrobiální ATP byl poté extrahován pomocí kationaktivního detergentu a jeho obsah
stanoven za použití luciferin-luciferázového komplexu. Počet buněk byl vypočítán
z kalibračních přímek koncentrace ATP nebo emitovaného světla (RLU) k počtu buněk
(KTJ.ml-1). Tato metoda byla jednoduchá a rychlá, ale nedostatečně citlivá.
Další možností je využití koncentrace a separace mikroorganismů z potraviny ještě před
provedením ATP bioluminiscenčního testu. Využívá se filtrace nebo centrifugace vzorku
následovaná uvolněním mikrobiálního ATP a jeho kvantifikací. Nutnost dvou-krokové
procedury pro bioluminiscenční detekci mikroorganismů byla dána určitou nestabilitou
luciferasy v lyzačním činidle. Nově bylo vyvinuto činidlo pro jedno-krokovou analýzu. Jedná
se o mutantní formu luciferasy světlušek, která je více odolná k lytickým činidlům. Stabilní
luciferasa je použita v činidle kombinujícím lytické složky s luciferin-luciferázovým
komplexem a umožňuje tak nový zjednodušený formát testu.
Pro filtrovatelné roztoky byl zaveden systém MicroStar™ (výrobce Millipore Corp.)
umožňující stanovení počtu kvasinek (trvání testu několik minut) a bakterií (trvání testu
několik hodin). Nejprve dochází k filtraci vzorku přes speciální membránový filtr schopný
zadržet mikroorganismy. Systém umožňuje detekci jediné buňky kvasinek ihned po filtraci.
Detekce bakterií vyžaduje krátkou inkubaci membránového filtru na povrchu vhodného
živného média. Poté se na filtr nastříká činidlo uvolňující ATP a po krátkém sušení je přidáno
bioluminiscenční činidlo. To vyvolá vznik „světelných spotů“ korespondujících s každou
buňkou kvasinek či bakteriální mikrokolonií. CCD kamerou lze vytvořit obraz
membránového filtru s prostorovou distribucí světelných skvrn. Získané výsledky jsou
srovnatelné se standardní plotnovou metodou. Moderní verzí je automatizovaný systém
Milliflex® Rapid Microbiology Detection and Enumeration System (výrobce Merck
Millipore).
6.4.2.1. Využití v praxi
Při stanovení celkového počtu mikroorganismů v syrovém mléce se nejčastěji používá
filtrační technika, která je vhodná pro screening cisternových vzorků mléka ve
zpracovatelských závodech a monitoring kvality během prodlouženého skladování mléka.
Metodu lze využít také k hodnocení kvality pasterovaného mléka, kontrole sterility UHT
výrobků, k detekci antibiotik v mléce nebo při sledování aktivity bakterií mléčného kvašení
při hodnocení čistých mlékařských kultur. Testovací soupravy a potřebné přístrojové
vybavení nabízí v současnosti několik firem. Jedná se například o RapidScreen Dairy
Nepřímé metody
51
(výrobce Celsis Inc.) nebo 3M™ Microbial Luminescence System II (výrobce 3M) určený pro
testování sterility UHT mléka a mléčných výrobků.
Při hodnocení povrchové kontaminace jatečně opracované drůbeže se testuje oplachová voda
nebo se provede stěr sterilním tamponem, v případě prasat a skotu se stěr provádí sterilní
testovací houbičkou zvlhčenou fyziologickým roztokem s přídavkem Tweenu 80. Tekutina
z houbičky je dále zpracována pomocí filtračního zařízení Filtravette™ (výrobce New
Horizont Diag. Corp.). Jedná se o speciální luminometrickou kyvetu jejíž dno slouží jako
membránový filtr pro záchyt bakterií. Další možností je odebrání a zpracování vzorku
svaloviny z povrchu jatečně opracovaného těla. Systém Filtravette™ je využitelný také pro
stanovení mikrobiální kontaminace mletého masa. ATP bioluminiscenční metoda je
využitelná také při hodnocení vařených či vakuově balených masných výrobků.
Při detekci bakterií způsobujících kažení piva (Lactobacillus brevis) byl aplikován systém
MicroStar™ (výrobce Millipore Inc.), k vyšetření sycených nápojů lze použít např. filtrační
systém Haemocell Rapid Microbial Quality Assurance (RMQA) (výrobce GEM Biomedical,
Inc). Reagenční kit Bev-Trace™ (výrobce Biotrace Ltd.) je určený pro rychlou detekci
mikroorganismů kažení ve všech filtrovatelných nápojích, ať už pasterovaných či
sterilizovaných filtrací. Další možností je např. RapidScreen Food & Beverage (výrobce
Celsis Inc.).
6.4.2.2. Výhody a nevýhody
Vzhledem k tomu, že existuje vědeckými studiemi podložená korelace mezi množstvím ATP
a počtem mikroorganismů, je ATP bioluminiscenční test vhodnou alternativou k tradiční
plotnové metodě při rutinním mikrobiologickém hodnocení potravin a nápojů. Výhodou je
relativně jednoduché a rychlé provedení testu, v porovnání s klasickou kultivační metodou je
úspora času značná. Naměřené výsledky jsou archivovány a mohou být dále zpracovávány.
Naopak nevýhodou je nutnost počáteční úpravy vzorku tak, aby bylo mikrobiální ATP
separováno od ATP z jiných zdrojů (např. somatických buněk). Dále je to samozřejmě
počáteční investice do přístrojového vybavení. Test sám o sobě není specifický, v případě
detekce vybrané skupiny mikroorganismů je nezbytné zařazení vhodného specifického
postupu (např. imunomagnetická separace pro zachycení cílových bakterií či lýza buněk
druhově specifickými bakteriofágy).
6.4.3. Monitoring hygieny a sanitace v potravinářských provozech ATP bioluminiscenční techniky nachází široké uplatnění při hodnocení účinnosti sanitace
provozních povrchů a zařízení a monitoringu hygieny v rámci systémů HACCP, a to v celé
řadě potravinářských provozů (mlékárenství, výroba nápojů, masný průmysl, atd.) po celém
světě. Běžné stěrové či otiskové techniky kombinované se stanovením počtu mikroorganismů
plotnovou metodou mohou detekovat pouze povrchovou kontaminaci mikroorganismy, ale
neposkytují informace o tom, zda byl povrch dostatečně očištěn.
Metoda ATP bioluminiscence, provedená po rutinní sanitaci povrchu či zařízení, umožňuje
během několika minut posouzení celkové úrovně organického znečištění povrchu. V tomto
případě je detekován veškerý ATP přítomný na testovaném povrchu (mikrobiální ATP, ATP
ze somatických buněk, příp. volné ATP). Z vyšetřované plochy se provede stěr speciálním
sterilním tamponem, ten je vložen do zkumavky obsahující všechna potřebná činidla pro
vlastní bioluminiscenční reakci. Po vložení do přenosného luminometru je změřeno množství
emitovaného světla v RLU. Získané výsledky jsou následně hodnoceny jako vyhovující či
nevyhovující, a to podle předem nastavených limitních hodnot RLU pro každé kontrolované
místo. Provedení testu je velmi jednoduché, příprava vzorku a měření trvá pouze několik
Nepřímé metody
52
minut. Podobným způsobem lze testovat i oplachovou vodu. V tomto případě se k odběru
vzorku používají speciálně upravené odběrové tyčinky se žlábkem umožňující zachycení
konstantního množství vzorku.
Testovací soupravy a měřicí přístroje dodává na trh celá řada výrobců – např. 3M (3M™
Nespornou výhodou je jednoduché a rychlé provedení testu, při použití přenosného
luminometru se stanovení provádí přímo v místě kontroly. Naměřené výsledky lze ihned
porovnat se zadanými hraničními hodnotami, archivovat či dále zpracovávat.
Nezbytná je počáteční investice na pořízení měřicího přístroje, vlastní stanovení již není
finančně náročné. Pro záchyt ATP je důležité správné provedení stěru, vliv má také charakter
stíraného povrchu. Dalším omezením je případná přítomnost detergentů, sanitačních a dalších
čistících prostředků na testovaném povrchu. Tyto látky mohou interferovat s luminiscenční
reakcí a vyvolávat falešně pozitivní či negativní výsledky.
6.4.4. Využití ATP bioluminiscence při detekci vybraných patogenů Metodu ATP bioluminiscence lze využít také při detekci vybraných původců alimentárních
onemocnění. Obecný postup stanovení je následující: 1) rozeznání cílových bakterií, 2) lýza
cílových bakterií a uvolnění ATP a 3) vlastní ATP bioluminiscenční test. Rozeznání cílových
bakterií se provádí pomocí specifických protilátek nebo specifických bakteriofágů.
Výhodou fágů, oproti použití specifických protilátek, je vyšší specifita a rychlejší vazba na
cílové buňky. Masivní produkce bakteriofágů je také výrazně levnější než produkce
protilátek. Využití bakteriofágů umožňuje současnou koncentraci a detekci cílových buněk.
Bakteriální buňky jsou zachyceny na biosorbent obsahující fágy, následná lýza buněk je
detekována ATP bioluminiscenční metodou. Lze stanovit počet navázaných bakterií.
Jiným přístupem je využití rekombinantních fágů. Fágy specifické pro sledovanou bakterii
jsou geneticky upraveny vložením genů pro bakteriální luciferasu (tzv. lux geny). Samotné
fágy nejsou schopné gen exprimovat a zůstávají „tmavé“. Po infekci hostitelské buňky dojde
k syntéze luciferasy, bakteriální buňky se „rozsvítí“ a mohou být detekovány luminometrem.
Byly připraveny rekombinantní fágy pro detekci bakterií čeledi Enterobacteriaceae, Listeria
monocytogenes, Salmonella spp. či Mycobacterium tuberculosis.
6.4.5. Měřicí přístroje Na trhu jsou k dispozici 3 hlavní skupiny přístrojů schopných měřit nízké hladiny
bioluminiscence. První z nich jsou trubicové luminometry, které jsou uzpůsobeny k měření
intenzity luminiscence v jednotlivých zkumavkách, obvykle využívají fotonásobiče. Mohou
být víceúčelové pro široké spektrum vzorků či specifické pro určitý typ zkumavek a činidel
určených pro daný přístroj. Přenosné bateriové luminometry jsou vhodné pro monitoring
hygieny v potravinářském průmyslu.
Vzhledem k popularitě mikrotitračních destiček našli své uplatnění na trhu i luminometry
umožňující automatické odečítání destiček. Měření luminiscence je zde však doprovázeno
určitými konstrukčními problémy.
V některých případech jsou kromě intenzity luminiscence zapotřebí i informace o jejím
prostorovém uspořádání. Pro tento účel se používají zobrazovací zařízení pro hodnocení
objektů různého tvaru a velikosti jako jsou Petriho misky či blottovací membrány, mezi
nejpoužívanější patří CCD kamery různého typu.
Stanovení rezistence k AML
53
7. STANOVENÍ CITLIVOSTI MIKROORGANISMŮ K AML
Rezistence mikroorganismů k antimikrobiálním látkám (AML) je dnes celosvětově jedním
z největších problémů nejen klinické a veterinární praxe. Velký zájem o tuto problematiku je i
v oblasti mikrobiologie potravin, protože řada rezistentních bakterií je šířena nebo má svůj
rezervoár právě v potravinách a potravinových surovinách. Z tohoto důvodu se při vyšetření
potravinových izolátů stále častěji využívají i metody stanovení citlivosti k antimikrobiálním
látkám.
V současnosti jsou klinické laboratorní standardy pro testování citlivosti pravidelně
aktualizovány např. americkou organizací CLSI (Clinical Laboratory Standards Institute;
dříve NCCLS – National Committee for Clinical and Laboratory Standards). Metody
stanovení citlivosti, resp. rezistence mikroorganismů k antimikrobiálním látkám lze rozdělit
na semikvantitativní (disková difúzní metoda) a kvantitativní (tzv. diluční metody – např.
7.1.1. Antimikrobiální látky a jejich účinek na mikroorganismy Jako antimikrobiální látky se označují léčiva používaná k profylaxi či terapii infekčních
onemocnění. Tyto látky mohou být v podstatě dvojího druhu – antibiotika, která jsou
přírodního původu (dnes často chemicky modifikovaná či synteticky vyráběná) a
chemoterapeutika, která jsou připravená pouze chemicky.
Působení antimikrobiálních látek může být mikrobistatické (reverzibilní zastavení růstu a
množení mikroorganismů) nebo mikrobicidní (usmrcení mikroorganismů). Účinek
mikrobistatických látek nastupuje později, mikrobicidní látky působí ireverzibilně a rychle.
Mezi mikrobistatické látky patří např. tetracykliny, makrolidy, sulfonamidy; mikrobicidní
účinek mají např. β-laktamy, aminoglykosidy či polypeptidy.
Mimo antibiotika a chemoterapeutika mohou mít antimikrobiální účinek také další skupiny
látek. Jedná se o látky působící na mikrobiální enzymy (např. oxidační činidla, chelatační
látky, těžké kovy), látky reagující s DNA (např. chemické mutageny či cytostatika) či látky
působící jako imunosupresiva. Antibiotika a chemoterapeutika mohou zasahovat různými
způsoby na různých místech mikrobiální buňky.
Mezi hlavní mechanismy patří:
- inhibice syntézy buněčné stěny (např. β-laktamy, glykopeptidy),
- poškození syntézy cytoplasmatické membrány (např. polypeptidy – polymyxin B),
- inhibice proteosyntézy (např. aminoglykosidy, tetracykliny, chloramfenikol),
- porucha syntézy nukleových kyselin (např. chinolony, nitroimidazoly),
- antagonismus a kompetitivní inhibice (např. sulfonamidy).
7.1.2. Rezistence mikroorganismů k antimikrobiálním látkám Jako rezistenci obecně označujeme schopnost mikrobiální populace přežít účinek inhibiční
koncentrace dané antimikrobiální látky. Některé mikroorganismy jsou přirozeně (primárně)
rezistentní k některým antimikrobiálním látkám. Přirozená rezistence je geneticky podmíněná
necitlivost k dané látce, a to bez ohledu na to, zda došlo k předchozímu kontaktu
mikroorganismu s touto látkou. Příkladem může být rezistence gramnegativních střevních
tyčinek vůči penicilinu a makrolidům, rezistence streptokoků k aminoglykosidům či
enterokoků a listerií k cefalosporinům.
Stanovení rezistence k AML
54
Příčinou primární rezistence může být nepropustnost zevní membrány gramnegativních
bakterií pro danou antimikrobiální látku, produkce enzymů schopných antimikrobiální látku
inaktivovat, absence cílové struktury nebo její necitlivost k působení antimikrobiální látky.
U mikroorganismů se stále více setkáváme se získanou (sekundární) rezistencí. Tento typ
rezistence vzniká při působení antimikrobiální látky na populaci mikroorganismů, kdy se pod
selekčním tlakem antimikrobiální látky selektují rezistentní kmeny. Může se jednat o
rezistenci negenetického i genetického původu. V prvním případě může rezistence vzniknout
např. v důsledku ztráty specifických receptorových struktur. Sekundární rezistence
genetického původu může být chromosomální (např. spontánní mutace) či
extrachromosomální (reprezentovaná např. plasmidy či transpozony, viz též kapitola 9.2.3.).
Velkým problémem je multirezistence (mnohočetná rezistence), kdy je mikroorganismus
současně rezistentní k většímu počtu antimikrobiálních látek. Dále potom zkřížená rezistence
– současná necitlivost mikroorganismů na antibiotika s podobnou chemickou strukturou a
stejným mechanismem účinku.
7.1.2.1. Mechanismy rezistence mikroorganismů k AML
Jednou z možností je změna cílové struktury, tedy změna v místě působení antimikrobiální
látky. Mikroorganismy jsou schopné změnit strukturu cílové molekuly zásahu antimikrobiální
látky (obvykle esenciálního enzymu) nebo exprimovat alternativní cílovou molekulu, která
není antimikrobiální látkou inhibována.
Dalším mechanismem je změna propustnosti (permeability). Jde buď o úplnou nepropustnost
– impermeabilitu pro danou antimikrobiální látku, nebo dochází k jejímu aktivnímu vyloučení
z buňky. Ke změně propustnosti dochází např. změnou molekuly přenašeče či alterací
aktivního transportního proteinu, může se jednat i o omezení tvorby ATP v případech, kdy je
antimikrobiální látka přenášena aktivním transportem. Příkladem aktivního vyloučení již
nakumulované antimikrobiální látky je efflux systém, kdy prostřednictvím transmembránové
pumpy dochází k transportu antimikrobiální látky ven z buňky. Effluxní systém je schopen
zachytit molekuly AML nejen v periplasmatickém prostoru, ale i v cytoplasmě.
Mezi mechanismy rezistence patří i velmi častá změna antimikrobiální látky, kdy
mikroorganismus produkuje enzymy, které antimikrobiální látku pozmění nebo štěpí a tím ji
inaktivují. V některých případech mikroorganismus produkuje cílový enzym v takovém
množství, že jej nelze danou koncentrací antimikrobiální látky vyblokovat – hovoří se o tzv.
hyperprodukci cílové struktury.
7.1.2.2. Vznik a šíření rezistence mikroorganismů k AML
Rezistence genetického původu může vznikat v podstatě dvojím způsobem – modifikací genu
na chromosomu nebo převzetím genetického materiálu. K modifikaci chromosomálního genu
dochází obvykle spontánní mutací bez ohledu na předchozí kontakt s antimikrobiální látkou.
Vzniklá rezistence má trvalý charakter a přenáší se do další generace v populaci. Jinou
možností vzniku rezistence je převzetí genetického materiálu neseného na mobilních
elementech, jako jsou R plasmidy, transpozony, integrony či genové kazety, příp. i
bakteriofágy. Geny rezistence získávají mikroorganismy vertikálně (dědí je) nebo
horizontálně (od jiných buněk v ekosystému), mimo to tyto geny vznikají i mutacemi.
Mezi základní způsoby přenosu genetického materiálu řadíme konjugaci, transdukci a
transformaci. Při konjugaci dochází k přenosu plasmidu z donorové buňky (F+) do buňky
recipientní (F-) pomocí tzv. sex-pilů, konjugace může probíhat v rámci jednoho druhu
mikroorganismu nebo i mezidruhově. Transdukce představuje přenos pomocí bakteriofága, do
jehož genetické informace je včleněna chromosomální či plasmidová DNA, která je následně
Stanovení rezistence k AML
55
s bakteriofágem přenesena do další bakteriální buňky. Transformace je přenos uvolněné DNA
(např. z lyzované buňky) vedoucí ke vzniku nových forem rezistence.
7.2. Semikvantitativní metody Semikvantitativní metody nachází uplatnění zejména při rutinním screeningu citlivosti
mikroorganismů k antimikrobiálním látkám. Do této skupiny náleží disková difúzní metoda,
která se používá zejména pro vyšetření citlivosti u rychle rostoucích, nenáročných bakterií a
některých náročnějších bakterií. Hodnocení se provádí na základě měření velikosti
inhibičních zón vzniklých okolo disků s antimikrobiální látkou. Podle velikosti inhibiční
zóny jsou vyšetřované kmeny mikroorganismů zařazeny do kategorie citlivý (S),
intermediálně rezistentní (I) (kmeny na rozmezí mezi citlivými a rezistentními) nebo
rezistentní (R) k dané antimikrobiální látce. Při správném provedení prokazují výsledky
diskové difúzní metody a výsledky kvantitativních dilučních metod vysokou shodu.
7.2.1. Disková difúzní metoda První metodou hodnocení účinku antibiotik na bakterie byla disková difúzní metoda. Poprvé
byla standardizována jako paper disk method v roce 1954. Navzdory své jednoduchosti, je
disková difúzní metoda založena na sofistikovaných fyzikálně-chemických principech, které
upravují dynamiku šíření (difúze) antibiotika ve vztahu k růstu bakterií v agarovém systému.
Po kontaktu disku napuštěného antibiotikem s povrchem inokulovaného agaru, začnou
molekuly antibiotika okamžitě difundovat do agaru a vytváří dynamicky se měnící gradient
koncentrace antimikrobiální látky. Mikroorganismy se začínají dělit, jejich počet vzrůstá až
do kritického množství. Okraj inhibiční zóny vzniká v okamžiku, kdy je koncentrace
antibiotika ještě schopná inhibovat mikroorganismus narůstající do velké buněčné masy.
Současně je hustota buněk dostatečně vyšší než absorbovatelné množství antibiotika
v bezprostřední blízkosti, subinhibiční množství antibiotika dovoluje mikroorganismu růst.
Difúzní koeficient antimikrobiální látky je ovlivněn nejen její molekulovou hmotností,
velikostí, iontovým nábojem či rozpustností ve vodě, ale také viskozitou a výškou agaru,
teplotou a dalšími inkubačními podmínkami. Růst mikroorganismu ovlivňuje zejména
dostupnost živin, hustota populace, růstová fáze inokula a inkubační teplota.
Výsledky in vitro testů citlivosti mohou být použity jako kvalifikovaný odhad k predikci
léčebného výsledku běžných dávek antibiotik u zdravých pacientů. Na druhou stranu
potenciální klinická účinnost antibiotika může být ovlivněna řadou dalších faktorů, proto
výsledky kvalitativních testů musí být v korelaci s kvantitativními hodnotami MIC
(minimální inhibiční koncentrace).
Některé disky mohou být využity i pro screening některých speciálních mechanismů
rezistence. Jedná se např. o detekci rezistence k vysokým hladinám aminoglykosidů u
enterokoků nebo detekci přítomnosti širokospektré β-laktamasy (ESBL) u izolátů E. coli,
Klebsiella pneumoniae či K. oxytoca.
Mezi hlavní výhody diskové difúzní metody patří jednoduché provedení bez nutnosti
náročného vybavení, flexibilní změna spektra vyšetřovaných antimikrobiálních látek a nízká
cena. Hlavní nevýhodou, mimo toho, že se nejedná o kvantitativní metodu, je do jisté míry
časová náročnost, zahrnující zejména přípravu agarových ploten a dobu potřebnou
k manuálnímu odečítání inhibičních zón a interpretaci výsledků. Přesto má v klinické praxi
disková difúzní metoda svou nezastupitelnou roli.
Provedení a využití diskové difúzní metody je detailně studováno v rámci předmětu Mikrobiologie
potravin a mikrobiologické laboratorní metody (viz skripta Mikrobiologie potravin – praktická cvičení
I. Obecná mikrobiologie).
Stanovení rezistence k AML
56
7.3. Kvantitativní – diluční metody Diluční metody jsou kvantitativní metody určené ke stanovení minimální inhibiční
koncentrace (MIC), tedy nejnižší testované koncentrace daného antibiotika, která inhibuje
viditelný růst mikroorganismu. MIC se obvykle vyjadřuje v mg.l-1, příp. v µg.ml-1. Odečtené
hodnoty MIC se porovnávají s interpretačními kriterii a testovaný mikroorganismus se označí
jako citlivý (S), intermediárně rezistentní (I) či rezistentní (R) k danému antibiotiku.
Vyšetření se provádí na agarových nebo bujonových půdách, které obsahují zvolené
koncentrace antibiotika, obvykle v dvojnásobné geometrické řadě. Do půd se očkuje
standardní inokulum testovaného mikroorganismu. Kvalita získaných výsledků závisí na
přesném dodržení metody a ověřuje se systémem kontrol.
Stanovení hodnoty MIC je důležité pro určení terapeutické dávky antimikrobiální látky. Mezi
nejpoužívanější metody patří agarová diluční metoda, diluční mikrometoda a Etest.
7.3.1. Agarová diluční metoda Agarová diluční metoda je referenční metodou stanovení citlivosti mikroorganismů
k antimikrobiálním látkám, používá se mimo jiné pro ověřování spolehlivosti ostatních
metod.
7.3.1.1. Provedení a vyhodnocení testu
Jako živné médium se používá Mueller-Hinton agar, který je obohacený o testovanou
antimikrobiální látku. Antimikrobiální látka se naředí a jednotlivá ředění se přidávají
k rozpuštěnému a vytemperovanému agaru (45 – 50 °C), který je po promíchání rozléván do
Petriho misek tak, aby výsledná vrstva agaru byla 3 – 4 mm. Po ztuhnutí lze připravené plotny
skladovat při chladničkové teplotě po dobu několika dnů. Obvykle se připravuje 12 – 15
koncentrací jednoho antibiotika ředěných dvojnásobně geometrickou řadou.
Inokulum vyšetřovaných bakterií se aplikuje na povrch předsušeného agaru formou spotů
speciálním vzorkovačem. Na jedné Petriho misce (tj. jedné koncentraci antibiotika) můžeme
testovat až 20 různých kmenů bakterií. Soubor vyšetřovaných kmenů takto naočkujeme na
všechny připravené koncentrace testovaného antibiotika. Po příslušné době inkubace
odečteme pro každý testovaný bakteriální kmen hodnotu MIC. Inkubační podmínky se volí
tak, aby odpovídali testovanému mikroorganismu. Příklad provedení a vyhodnocení agarové
diluční metody je uveden na obrázku 24.
1.
2.
Obrázek 24: Agarová diluční metoda: 1. inokulované plotny, 2. plotny po inkubaci s nárůstem
mikroorganismů. (Engelkirk and Duben-Engelkirk, 2008 – upraveno)
V uvedeném příkladu se jedná se o testování rezistence k penicilinu, kdy bylo připraveno 5
ředění antibiotika. Na agarových plotnách bylo současně testováno šest různých kmenů,
každý z nich byl vždy aplikován na stejnou pozici. Po ukončení inkubace se hodnotí nárůst
Stanovení rezistence k AML
57
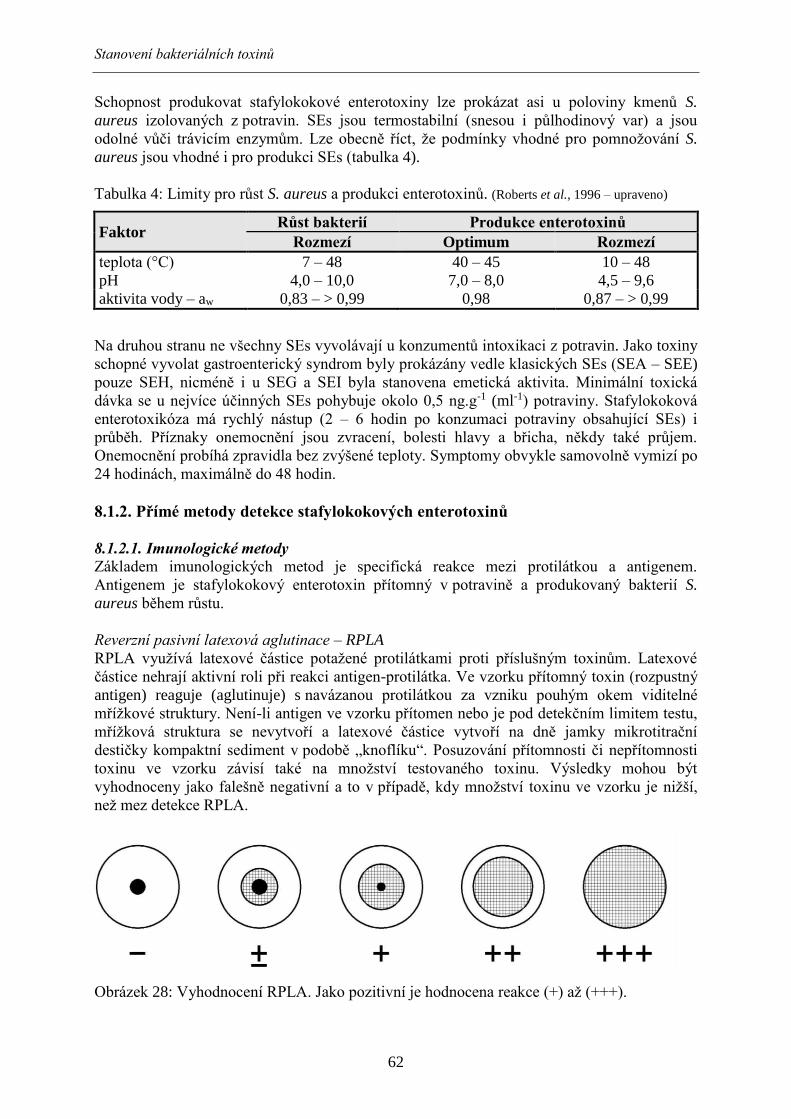
mikroorganismů na jednotlivých plotnách. Jako MIC se odečte první koncentrace bez
9.2.4. Makrorestrikční analýza a pulsní gelová elektroforéza (PFGE) Makrorestrikční analýza a následná pulsní gelová elektroforéza jsou považovány za zlatý
standard při konfirmaci epidemických kmenů různých patogenních agens. Jedná se o izolaci
DNA ukotvenou v agarózových bločcích a její štěpení v místech specifických pro daný
restrikční enzym, který štěpí pouze na limitovaném počtu míst v genomu. Používány jsou
restrikční enzymy izolované z různých mikroorganismů a pro každé agens je vhodný jiný
restrikční enzym, například XbaI pro salmonely nebo E. coli, SmaI pro kampylobaktery, ApaI
pro listerie. Rozdělení fragmentů probíhá v agarózovém gelu ve speciálně upravené
elektroforetické vaně hexagonálního tvaru. Gel je poté obarven v roztoku ethidium bromidu a
fragmenty DNA jsou vizualizovány v UV světle. Gely jsou analyzovány vhodným software
například programem BioNumerics (Applied Maths, Belgie), který vypočítá podobnost
srovnávaných kmenů.
Typizační metody
75
Metoda makrorestrikční analýzy je poměrně náročná na čas (postup podle standardního
protokolu trvá 2-3 dny). I přesto je tato metoda považována za standard v typizačních
metodách a umožňuje mezilaboratorní (mezinárodní) porovnávání kmenů.
9.2.5. Sekvenační metody Nejmodernější typizační metody jsou založeny na principu sekvenace, ať už se jedná o
sekvenování určitého úseku DNA (MLVA, MLST) nebo celého bakteriálního genomu.
Při metodě MLVA (Multi-Locus Variable-Number Tandem Repeat Analysis, multilokusová
analýza tandemových repetic) jsou zjišťovány sekvence určitých oblastí obsahující různé
počty tandemových repetic. Podle počtu těchto repetic je pak definována příbuznost kmenů.
Tato metoda umožňuje detailní rozlišení kmenů, protože ke změnám v repeticích dochází
velmi snadno a často. Tato metoda je vhodná k porovnávání blízce příbuzných kmenů
jednoho sérotypu, například v rámci jedné epidemie v jedné lokalitě.
Při metodě MLST (Multi-Locus Sequence Typing, multilokusová sekvenační typizace) jde
naopak o sekvenaci oblastí tzv. housekeeping – provozních genů, kdy jsou oblasti těchto genů
obvykle konzervativní. Metoda je vhodná například po porovnání kmenů jednoho sérotypu
z různých zdrojů nebo různých států pro vytvoření fylogenetických stromů.
Celogenomová sekvenace poskytuje maximální diskriminační schopnost. Z výsledné
sekvence lze zjistit například celkové údaje o genech rezistence (rezistom) anebo genech
virulence (virulom). Tyto metody umožňují získání detailních informací o genetické stavbě
bakterií, odpadá při tom tedy vybírání nejvhodnějších cílů analýz. Získaná data poskytují
velkou variabilitu výběru porovnávaných vlastností. Nicméně je tato metoda a především pak
nové rychlé variace sekvenování (např. pyrosekvenace) velmi drahá a tudíž nevhodná
k rutinní typizaci bakterií. Do budoucna však metody celogenomové sekvenace budou
představovat jedinou možnost, jak v krátkém čase zjistit o daném patogenu co nejvíce
informací a tím zrychlit diagnostiku a typizaci při šetření hromadných výskytů.
9.3. Případové studie
9.3.1. Případová studie 1 Na luxusní lodi při plavbě po středomoří došlo k hromadnému výskytu alimentárního
onemocnění. Na lodi cestovalo 850 cestujících, délka výletu byla naplánována na 5 dní.
Šéfkuchař den před vyplutím trpěl střevními potížemi, ale v den plavby nastoupil do práce.
Šéfkuchař se svým týmem připravoval stravu pro pasažéry i posádku, prvním společným
pokrmem byla večeře. Téže noci se objevily trávicí obtíže u 150 cestujících, problémy se
vyznačovaly rychlým nástupem nevolnosti a prudkým průběhem onemocnění, z příznaků
bylo zaznamenáno zvracení, bolesti břicha, průjem, schvácenost, bolest celého těla. Už druhý
den většina příznaků u postižených osob ustoupila a třetí den po onemocnění došlo k úpravě
zdravotního stavu. Postupně se však onemocnění šířilo na další cestující, především na
příbuzné a nejbližší spolucestující osob, kteří již onemocnění prodělali.
Na základě explozivního charakteru epidemie, klinických příznaků postižených osob a
anamnézy kuchaře lze usuzovat na onemocnění virového původu. Přenos se obvykle děje
fekálně orální cestou. U pacientů by mělo být provedeno bakteriální a virologické vyšetření
stolice či zvratků. Pokud jsou k dispozici zbytky potravin, pak je indikováno jejich virologické
vyšetření.
Typizační metody
76
9.3.2. Případová studie 2 Na svatební hostině se podávala polévka s játrovými knedlíčky, svíčková na smetaně
s karlovarským knedlíkem a jako moučník tiramisu z domácích vajec. Druhý den večer 10 lidí
z 50 přítomných začalo trpět bolestmi břicha, průjmem bez příměsi krve, nevolností a
teplotou. Všech 50 účastníků konzumovalo polévku a hlavní jídlo. Do týdne došlo k úpravě
zdravotního stavu postižených osob.
Klinické příznaky onemocnění (včetně teploty) naznačují, že se jedná o bakteriální infekci.
Přenos se děje obvykle kontaminovaným vehikulem (potravinou). U pacientů by mělo být
provedeno bakteriální vyšetření stolice. Pokud jsou k dispozici zbytky potravin, pak je
indikováno jejich bakteriologické vyšetření. Podezřelými potravinami jsou zejména játrová
zavářka, houskový knedlík a tiramisu. Možným etiologickým agens je salmonela.
9.3.3. Případová studie 3 V restauraci nižší cenové skupiny s nízkým hygienickým standardem bylo podáváno jednotné
menu – těstoviny se sýrovou omáčkou. Pokrm byl připravován v době od 10 do 11 hodin a
poté byl uchováván při pokojové teplotě až do doby vydání (od 12. do 14. hodiny). Porce byly
vydávány postupně a ohřívány v mikrovlnné troubě. U části strávníků (porce vydané mezi 13.
a 14. hodinou) došlo přibližně hodinu po konzumaci k náhlé nevolnosti a zvracení, u části
postižených se přidal i průjem. Během několika hodin došlo k samovolné úpravě zdravotního
stavu postižených.
Podle klinických příznaků a rychlého nástupu onemocnění lze usuzovat na alimentární
intoxikaci. Pravděpodobně došlo k pomnožení toxinogenních bakterií v průběhu nesprávného
uchovávání pokrmu (těstoviny se sýrovou omáčkou). U pacientů by mohlo být provedeno
vyšetření zvratků či stolice na přítomnost toxinů (např. metodou ELFA). Pokud jsou
k dispozici zbytky potravin, pak je indikováno jejich vyšetření na přítomnost toxinů, případně
i bakteriologické vyšetření na počty S. aureus a B. cereus.
9.3.4. Případová studie 4 V průběhu letní dovolené někteří rekreanti kempu konzumovali grilované kuře prodávané
z pojízdného stánku. Do 48 hodin se u některých konzumentů projevily horečka, křeče
v břiše, průjem, u některých postižených s příměsí krve a celková slabost. Přidružila se
horečka, a zatímco u části pacientů došlo k samovolné úzdravě, u několika pacientů (děti
předškolního věku a jeden senior) klinické příznaky neustupují a museli navštívit zdravotní
středisko, jedna osoba byla pro závažné potíže hospitalizována v místním zdravotnickém
zařízení. V nemocnici byla provedena rehydratace a lékaři podali postiženému antibiotika ze
skupiny makrolidů.
Klinické příznaky onemocnění (včetně teploty) naznačují, že se jedná o bakteriální infekci.
Přenos se děje obvykle kontaminovaným vehikulem (potravinou). U pacientů by mělo být
provedeno bakteriální vyšetření stolice. Pokud jsou k dispozici zbytky potravin, pak je
indikováno jejich bakteriologické vyšetření. Podezřelým pokrmem je grilované kuře. U
pacientů by mělo být provedeno bakteriální vyšetření stolice. Pokud jsou k dispozici zbytky
potravin, pak je indikováno jejich bakteriologické vyšetření. Klinické příznaky (křeče v břiše)
a suspektní vehikulum naznačují, že by se mohlo jednat o kampylobakterovou infekci.
Použitá literatura
77
POUŽITÁ LITERATURA
ALBERTS, B., BRAY, D.; JOHNSON, A., LEWIS, J., RAFF, M., ROBERTS, K., WALTER, P. (eds.)
Základy buněčné biologie (Úvod do molekulární biologie). 1. vyd. Ústí nad Labem, CZ: Espero Publishing,
s.r.o. 2005. 630 s.
ANDERSON, E.S., WARD, L.R., DE SAXE, M.J., DE SA, J.D. Bacteriophage typing designations of
Salmonella typhimurium. The Journal of Hygiene, 1977, vol. 78, no. 2, p. 297-300.
Ground Beef by the Limulus Amoebocyte Lysate Test as Rapid Indicator of Microbial Quality. Applied and Environmental Microbiology, 1979, vol. 38, no. 5. p. 885-890.
JEßBERGER, N., DIETRICH, R., BOCK, S., DIDIER, A. Bacillus cereus Enterotoxin Act as Major
Virulence Factors and Exhibit Distinct Cytotoxicity to Different Human Cell Lines. Toxicon, 2014, vol. 77,
p. 49-57.
LAY, J.O. MALDI-TOF mass spectrometry and bacterial taxonomy. Trac-Trends in Analytical Chemistry,
2000, vol. 19, no. 8, p. 507-516.
LAY, J.O. MALDI-TOF mass spectrometry of bacteria. Mass Spectrometry Reviews, 2001, vol. 20, no. 4,
p. 172– 194.
LEWIS, K., SALYERS, A.A., TABER, H.W., WAX, R.G. (eds.) Bacterial Resistance to Antimicrobials.
1st ed. New York, USA: Marcel Dekker, Inc. 2002. 495 p.
LI, X.H., MA, H.M., DONG, S.Y., DUAN, X.J., LIANG, S.C. Selective labeling of histidine by a designed
fluorescein-based probe. Talanta, 2004, vol. 62, no. 2, p. 367-371.
LÓPEZ-CAMPOS, G., MARTÍNEZ-SUÁREZ, J.V., AQUADO-URDA, M., LÓPEZ-ALONSO, V.
Microarray Detection and Characterization of Bacterial Foodborne Pathogens. 1st ed. New York, USA:
Springer. 2012. 135 p.
LORENCOVÁ, A., VAŠÍČKOVÁ, P. Viry jako původci alimentárních onemocnění. Maso, 2013, vol. 24,
no. 2, s. 34-40.
MAZZEO, M.F., SORRENTINO, A., GAITA, M., CACACE, G., DI STASIO, M., FACCHIANO, A., COMI, G, MALORNI, A., SICILIANO, R.A. Matrix-assisted laser desorption ionization-time of flight
Použitá literatura
79
mass spectrometry for the discrimination of food-borne microorganisms. Applied and Environmental Microbiology, 2006, vol. 72, no. 2, p. 1180-1189.