29
Nukleové kyseliny Martin Hubálek Ústav organické chemie a biochemie, AV ČR, Praha

Nukleové kyseliny
Martin Hubálek
Ústav organické chemie a biochemie, AV ČR, Praha

Obsah
• Nukleové kyseliny – složení • MS analýza oligonukleotidů
– Historie MS analýzy nukleotidů – EI, FAB
– MALDI – ESI
• MSMS oligonukleotidů – Fragmentace – CID, IRMPD, UVPD, ECD, ETD, EDD
• MS nukleových bází, nukleosidů a nukleotidů
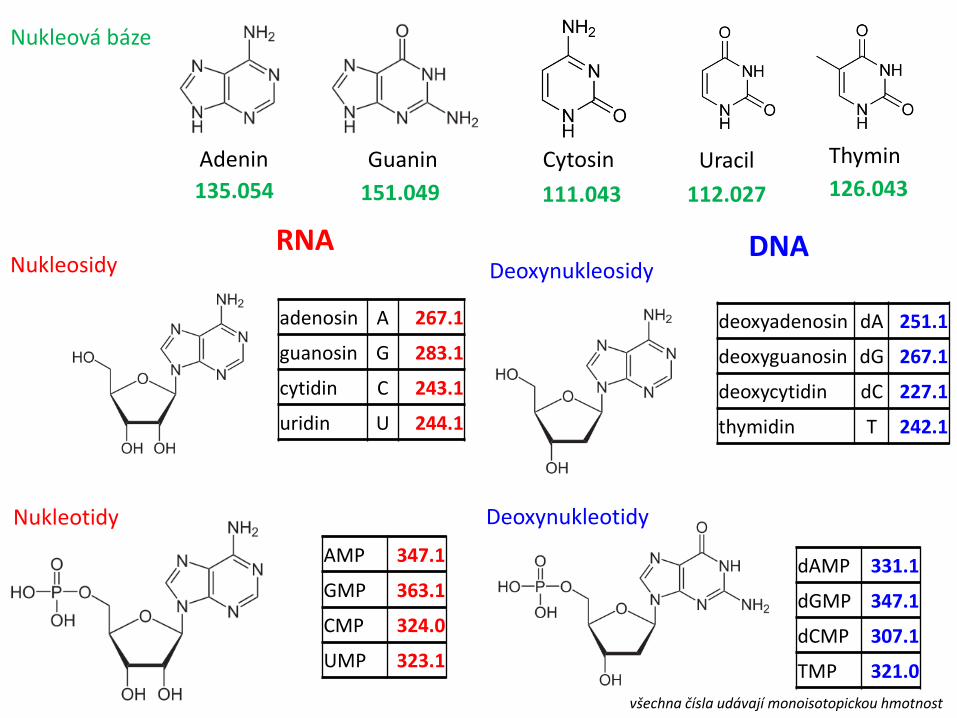
Adenin Guanin Thymin Uracil Cytosin
Nukleosidy Deoxynukleosidy
Nukleová báze
112.027 111.043 126.043 151.049 135.054
Nukleotidy Deoxynukleotidy
AMP 347.1
GMP 363.1
CMP 324.0
UMP 323.1
dAMP 331.1
dGMP 347.1
dCMP 307.1
TMP 321.0
adenosin A 267.1
guanosin G 283.1
cytidin C 243.1
uridin U 244.1
deoxyadenosin dA 251.1
deoxyguanosin dG 267.1
deoxycytidin dC 227.1
thymidin T 242.1
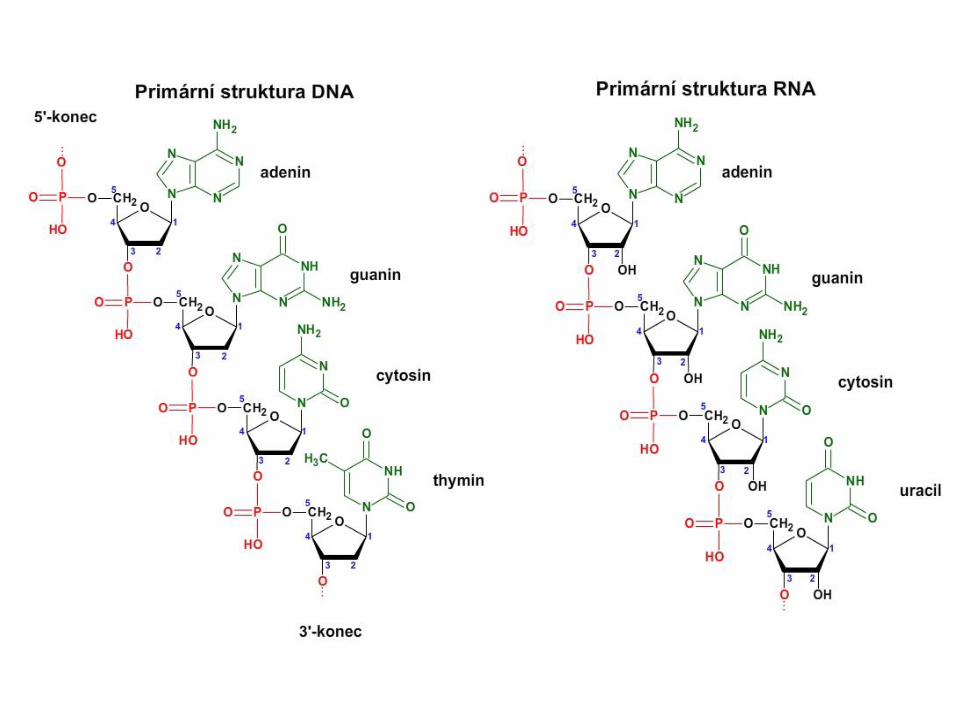
RNA DNA
všechna čísla udávají monoisotopickou hmotnost
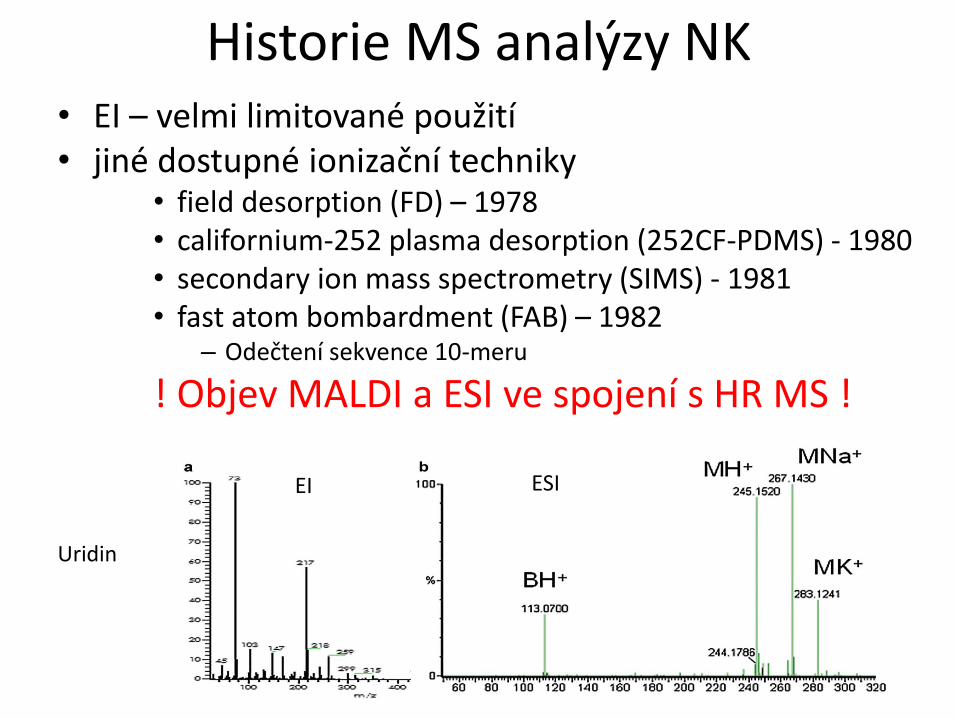
Historie MS analýzy NK • EI – velmi limitované použití • jiné dostupné ionizační techniky
• field desorption (FD) – 1978
• californium-252 plasma desorption (252CF-PDMS) - 1980
• secondary ion mass spectrometry (SIMS) - 1981
• fast atom bombardment (FAB) – 1982 – Odečtení sekvence 10-meru
! Objev MALDI a ESI ve spojení s HR MS !
Uridin
EI ESI

• Budzikiewicz – charakterizace dinukleotidů pomocí FD - 1978 “Now the question arises: is all this research on the sequencing of nucleic acids by mass spectrometry of purely academic interest? Mass spectrometric sequencing of peptides has its well established place in the armamentarium for structure elucidation since units comprising up to ten amino acids and
beyond can be handled. For oligonucleotides the limit for a direct analysis will probably be much lower. It has also to be taken into account that sequencing of both ribo and deoxyribonucleotides can readily be achieved by biochemical methods which require only minute amounts of material. Yet, there are three areas, where mass spectrometry can compete successfully, viz., a) if unusual bases are present (originally or after transformation by chemical reagents or by radiation) which are not readily recognised by the standard methods; b) for control purposes in synthetic or other chemical work, and c) for the structure elucidation of small units carrying characteristic
genetic information isolated from RNA or DNA which cannot be handled by the standard methods.”
• Základní použití • Určení molekulové hmotnosti nukleových kyselin
• Detailní strukturní charakterizace

MS NK obecně • v porovnání s proteiny/peptidy - komplikovaná MS analýza –
důvody: 1. Silná vazba fosfodiesterové skupiny a alkalických kationtů (Na+, K+) –
adukty s několika atomy
2. Snadná fragmentace – ztráta nukleobáze (1,2 eliminace) 3. Nízká iontová výtěžnost
• Nutná purifikace NK před konečným měřením – Gelová filtrace (spin kolonky) – SPE - C18
– Iontově výměnná
– Precipitace etanolem

MALDI • Běžně používaná technika pro analýzu NK
• Matrice
– 3-hydroxypikolinová kyselina – DNA, RNA – směs 2,3,4- a 2,4,6-trihydroxyacetofenon – RNA, DNA
• Nezbytná zpožděná extrakce (DE) • Snadná (až příliš) fragmentace
– ztráta nukleobáze
• protonizace báze - destabilizace glykosidické vazby – ztráta
• Platí více pro AGC, polyT – stabilnější ionty
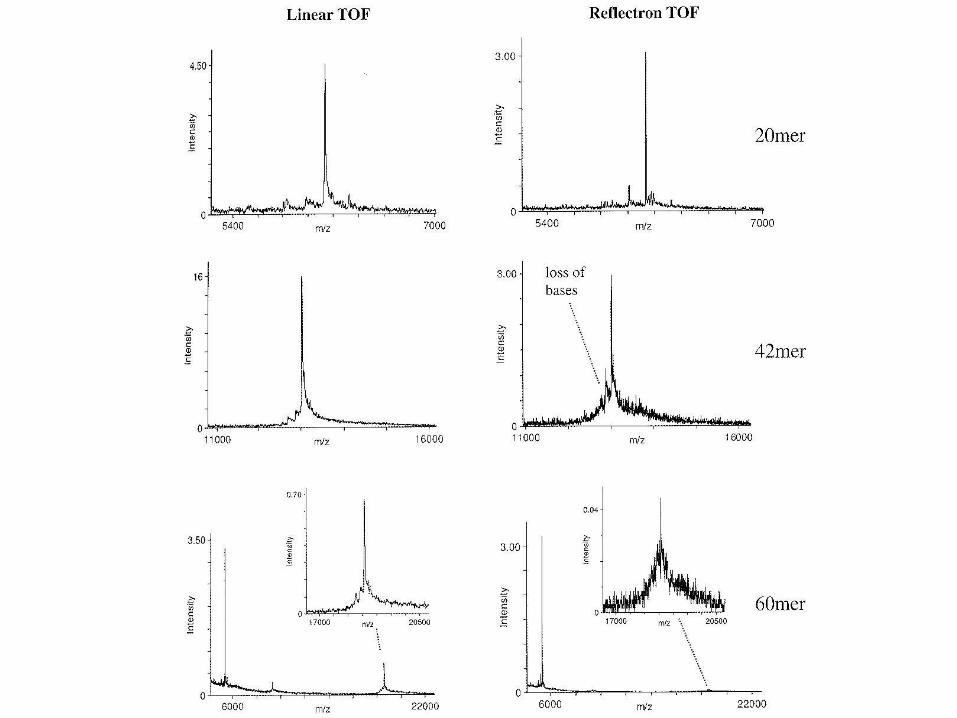
• Komplikace pro analýzu velkých NK
• DNA limit - 25 kDa (~80mer) • RNA stabilnější – 2’-hydroxyl stabilizuje glykosidickou vazbu
• méně závislé na vysoké čistotě vzorku než ESI
3-HPA
THAP
gallacetophenone
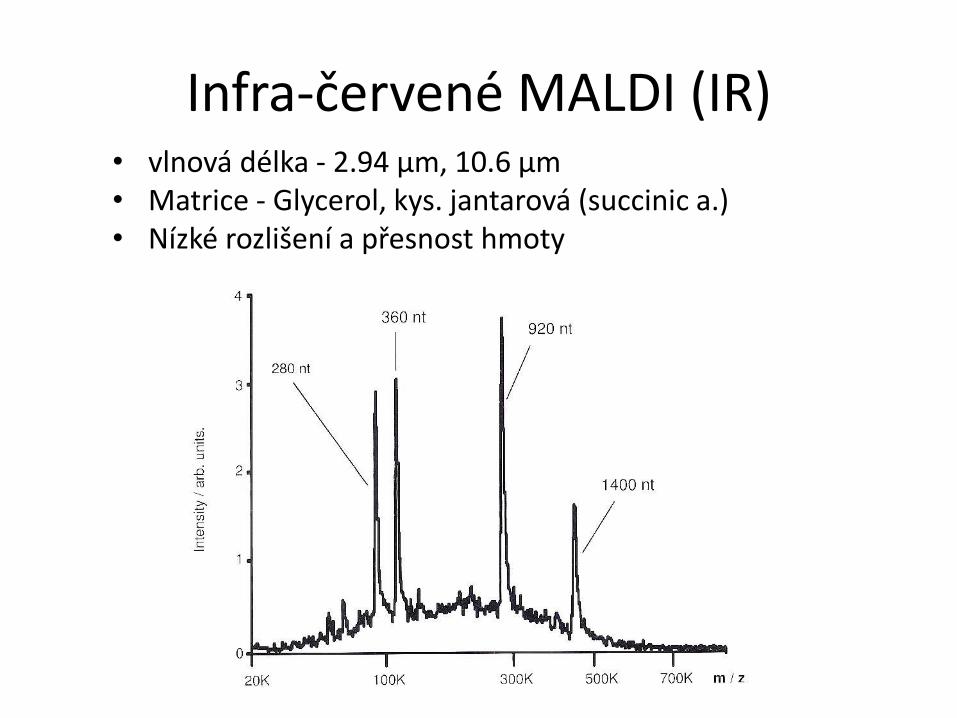
Infra-červené MALDI (IR) • vlnová délka - 2.94 µm, 10.6 µm
• Matrice - Glycerol, kys. jantarová (succinic a.) • Nízké rozlišení a přesnost hmoty
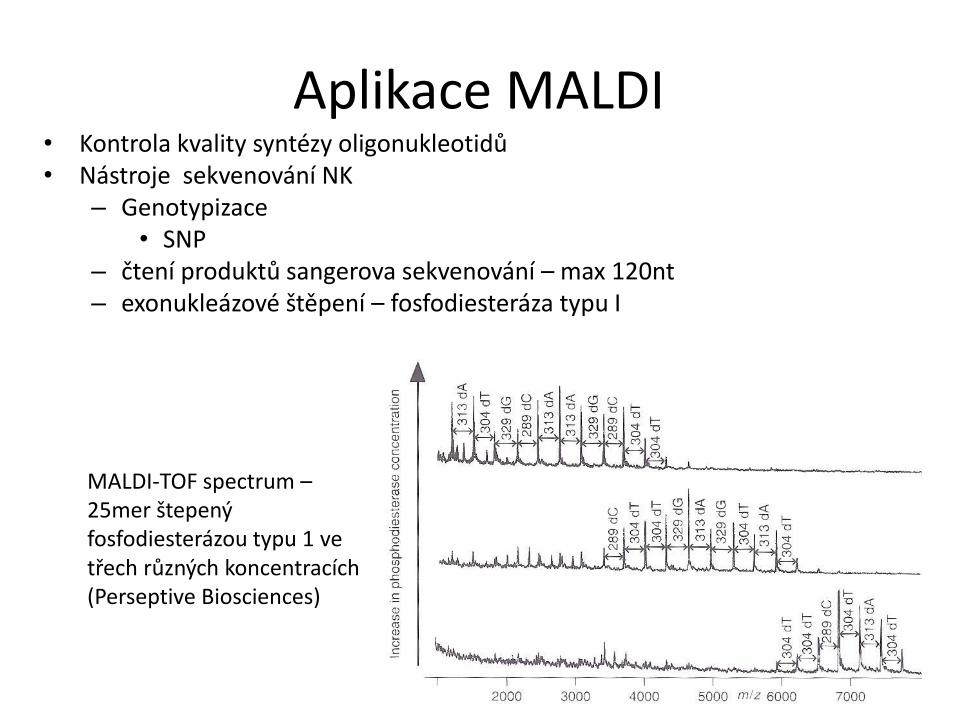
Aplikace MALDI • Kontrola kvality syntézy oligonukleotidů
• Nástroje sekvenování NK
– Genotypizace
• SNP
– čtení produktů sangerova sekvenování – max 120nt – exonukleázové štěpení – fosfodiesteráza typu I
MALDI-TOF spectrum – 25mer štepený fosfodiesterázou typu 1 ve třech různých koncentracích (Perseptive Biosciences)

ESI • Negativní mód – citlivější
• S výhodou udržovat pH ≥7
• Přídavek organické báze (triethylamine, piperidine,
imidazole)
• LC - octan amonný (10mM, pH 7-9)
Aplikace ESI • Odhalení modifikovaného místa
• Sekvenování antisense oligonukleotidů
• Identifikace patogenních agens
• HT diagnostika
• Objevy léčiv
• Farmakogenetika
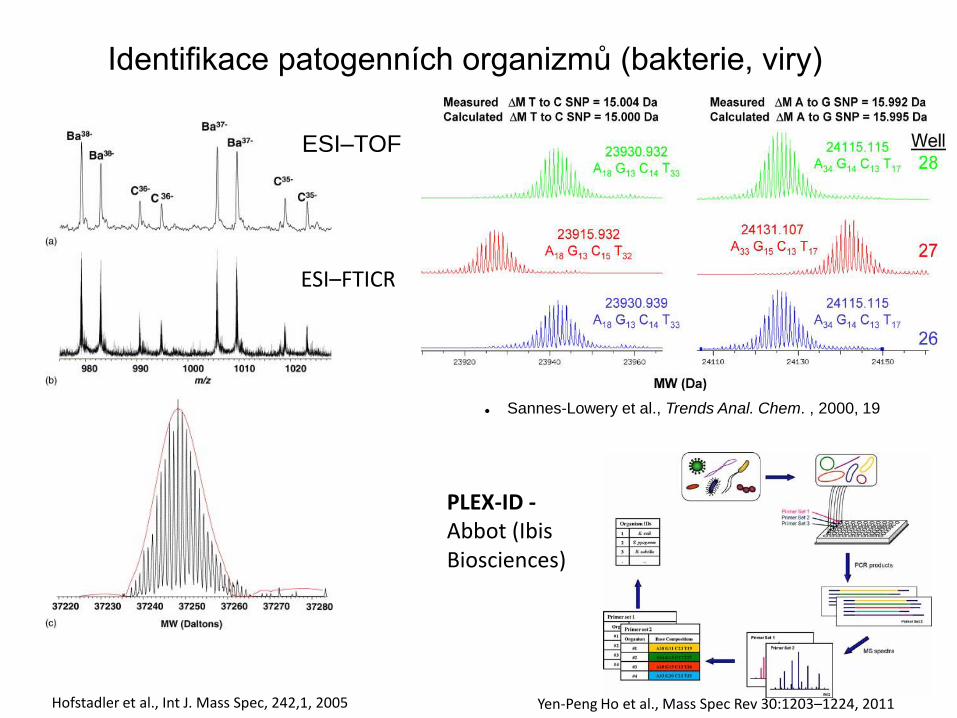
Identifikace patogenních organizmů (bakterie, viry)
Yen-Peng Ho et al., Mass Spec Rev 30:1203–1224, 2011
PLEX-ID - Abbot (Ibis Biosciences)
ESI–TOF
ESI–FTICR
Hofstadler et al., Int J. Mass Spec, 242,1, 2005
Sannes-Lowery et al., Trends Anal. Chem. , 2000, 19
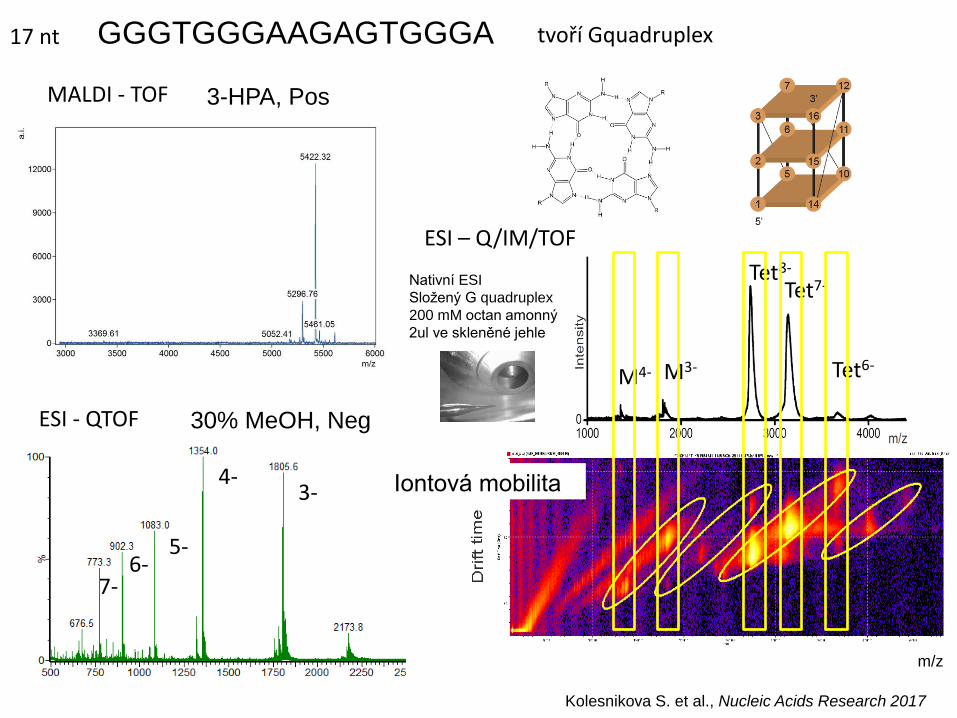
GGGTGGGAAGAGTGGGA
Nativní ESI
Složený G quadruplex
200 mM octan amonný
2ul ve skleněné jehle
tvoří Gquadruplex
4- 3-
5- 6-
7-
MALDI - TOF
ESI - QTOF
M4- M3-
Tet8- Tet7-
Tet6-
Iontová mobilita
m/z
3-HPA, Pos
30% MeOH, Neg
ESI – Q/IM/TOF
Kolesnikova S. et al., Nucleic Acids Research 2017
17 nt
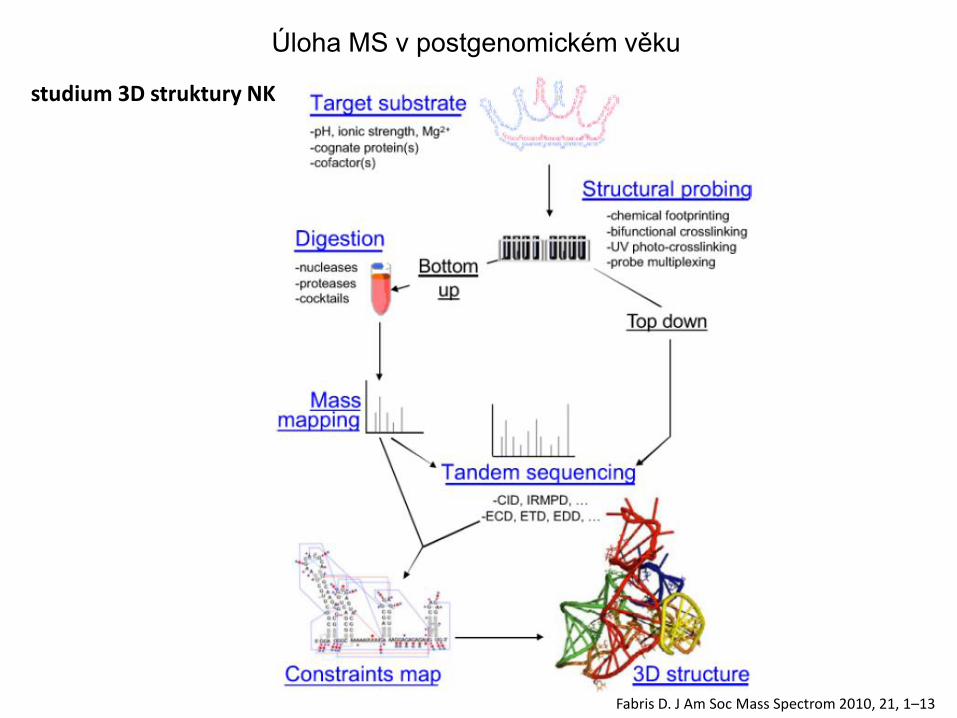
studium 3D struktury NK
Fabris D. J Am Soc Mass Spectrom 2010, 21, 1–13
Úloha MS v postgenomickém věku

McLuckey, Van Berkel and Glish (1992)
MSMS
2 hlavní pochody disociace v plynné fázi • Ztráta neutrální nebo nabité nukleobáze • Štepení vazeb mezi nukleosidy
• Klíčové štepení pro určení sekvence
Komplikace při interpretaci 1. pouze 4 stavební bloky spojené symetrickou
fosfodiesterovou vazbou – pravděpodobnost tvorby isobarických fragmentační iontů, které nelze v MS spektru rozlišit
2. různé disociační dráhy, některé poskytují neinformativní fragmenty – nukleobáze, sekundární štěpení hlavního řetězce

MSMS postřehy
• DNA x RNA rozdíl
• DNA fragmenty – typicky [a-B]- / w-
• RNA stabilnější, fragmenty - typicky c- / y-
• Závislé na E fragmentace
• Redukce nábojového stavu (0,2 náboje na nt) – větší šance na
kompletní pokrytí sekvence (př. 0,05% kys. octová, pH 2,5)
• De-novo x databázové prohledání
• Kompletní nebo částečné pokrytí sekvence
Důležité studie disociace v plynné fázi
• Určení protonových afinit nukleobází a jejich vlivu na ztrátu nukleobáze
(Greco et al., 1990)
• Výzkum stérických a konformačních faktorů (Gidden & Bowers, 2003)
• Studium mechanistických procesů, které řídí disociaci aktivovaných
oligonukleotidů (Wu & McLuckey, 2004)

Fragmentační metody NK
Iontové disociační metody
• CID (Collision Induced Dissociation)
• IRMPD (InfraRed Multiphoton Disociation)
• UVPD (UltraViolet PhotoDisociation)
Radikálové disociační metody
• Electron Capture Dissociation (ECD)
• Electron Transfer Dissociation (ETD)
• Negative Electron Transfer Dissociation (NETD)
• Electron Detachment Dissociation (EDD)
• Electron PhotoDetachment (EPD)
Hybridní
• ETcaD (Electron Transfer collision activated Dissociation
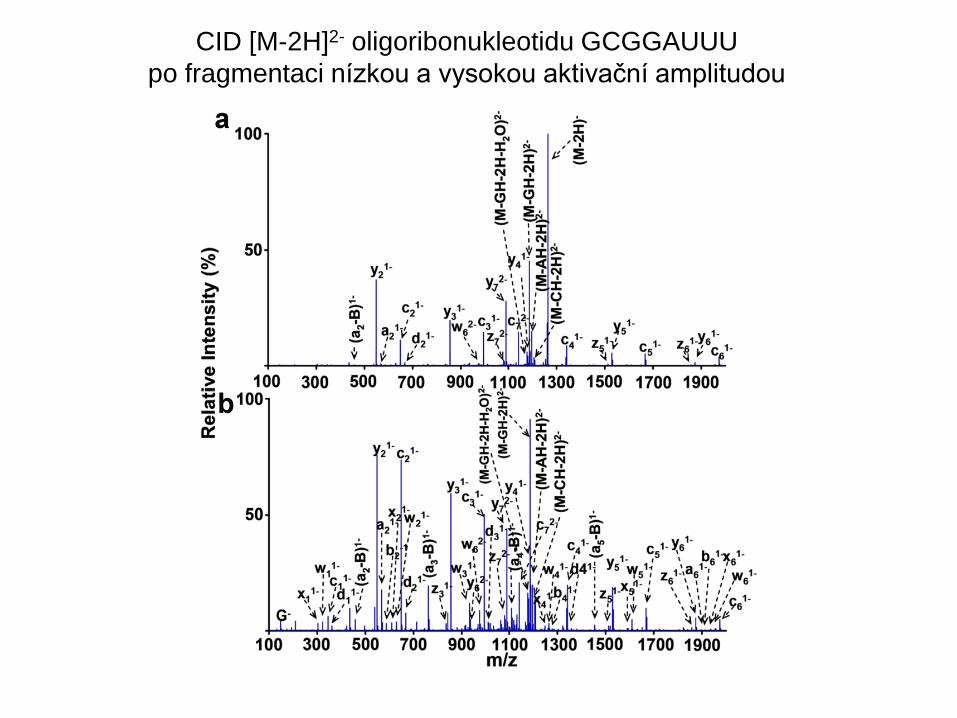
CID [M-2H]2- oligoribonukleotidu GCGGAUUU
po fragmentaci nízkou a vysokou aktivační amplitudou
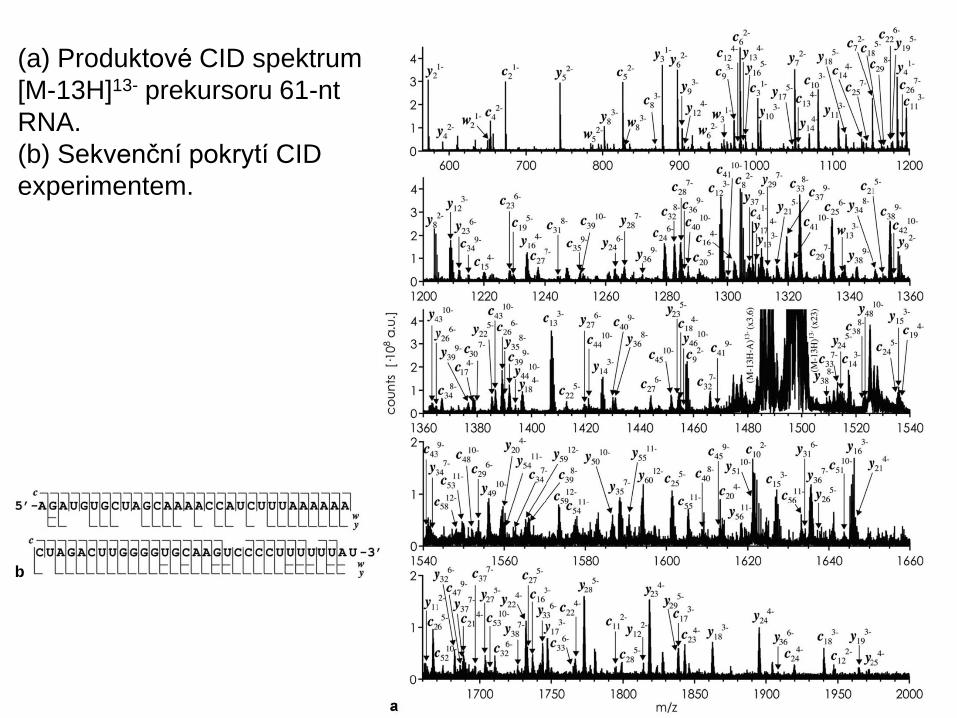
(a) Produktové CID spektrum
[M-13H]13- prekursoru 61-nt
RNA.
(b) Sekvenční pokrytí CID
experimentem.
b
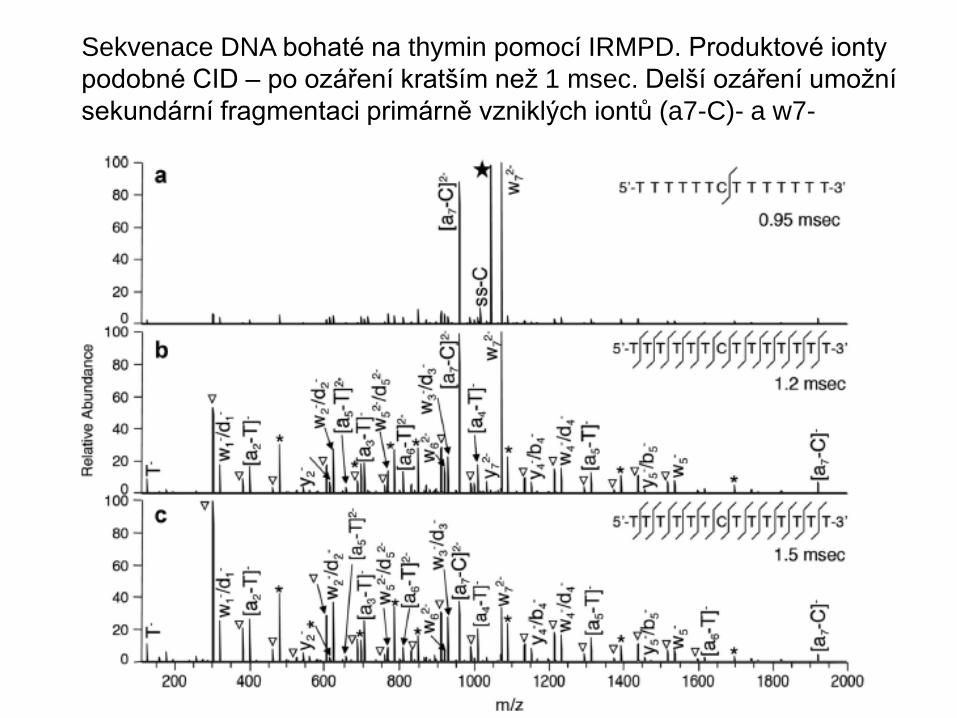
Sekvenace DNA bohaté na thymin pomocí IRMPD. Produktové ionty
podobné CID – po ozáření kratším než 1 msec. Delší ozáření umožní
sekundární fragmentaci primárně vzniklých iontů (a7-C)- a w7-
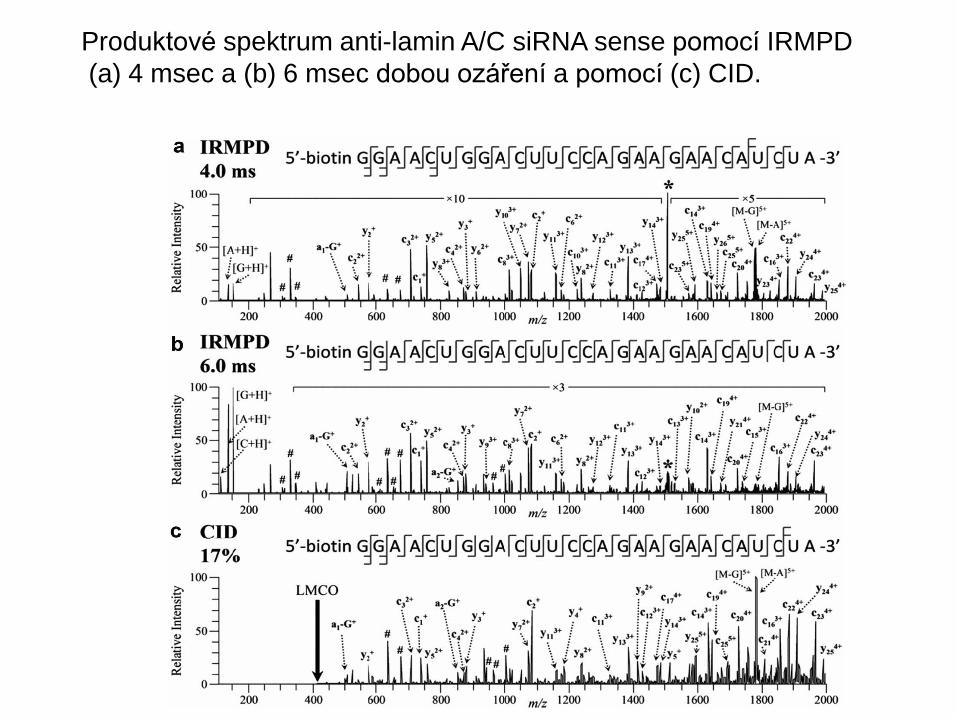
Produktové spektrum anti-lamin A/C siRNA sense pomocí IRMPD
(a) 4 msec a (b) 6 msec dobou ozáření a pomocí (c) CID.
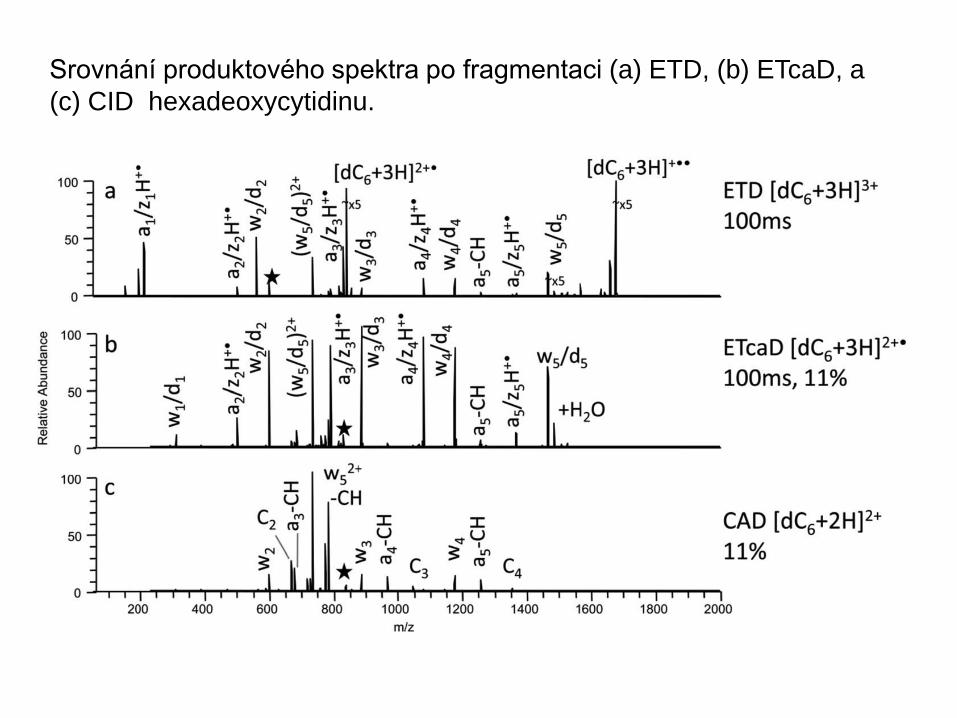
Srovnání produktového spektra po fragmentaci (a) ETD, (b) ETcaD, a
(c) CID hexadeoxycytidinu.
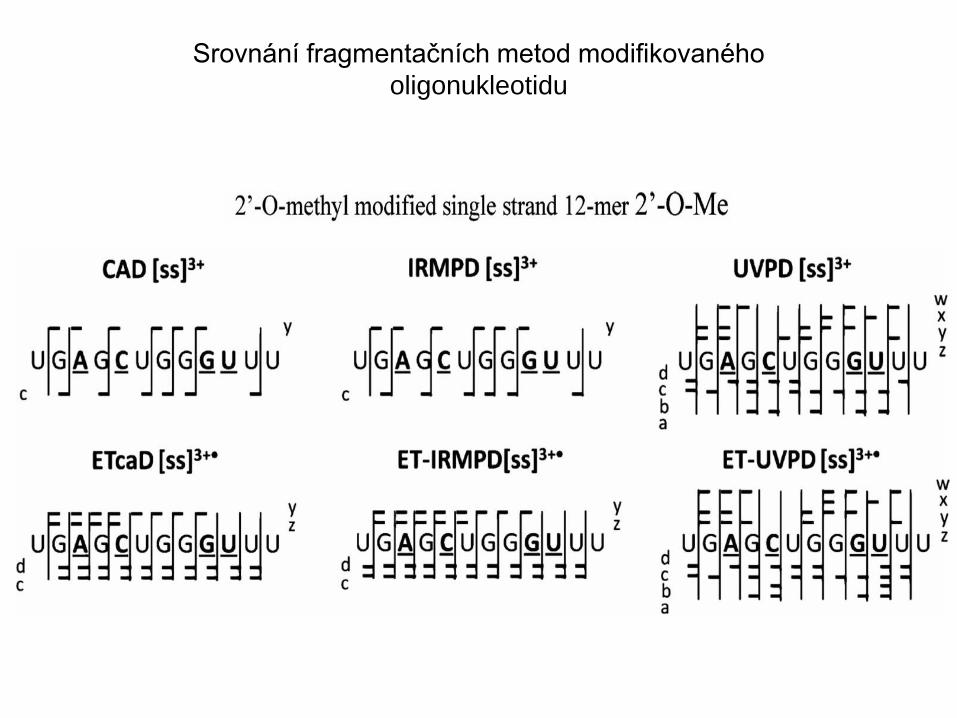
Srovnání fragmentačních metod modifikovaného
oligonukleotidu

MS nukleobází, nukleosidů a nukleotidů
• Nukleobáze a nukleosidy – chovají se jako jiné „malé molekuly“
– Báze jsou dostatečně bazické – LC na reverzní fázi v kyselém prostředí bez problémů
– nukleosidy lze také – retence na C18 malá
– měření v pozitivním módu
• Nukleotidy – větší problém – Fosfátová skupina snižuje náboj – Na+, K+ vícenásobně nabité stavy
– Vhodnější v NEGativním módu
– Reverzní fáze nevhodná – HILIC
• Typická metabolomická analýza
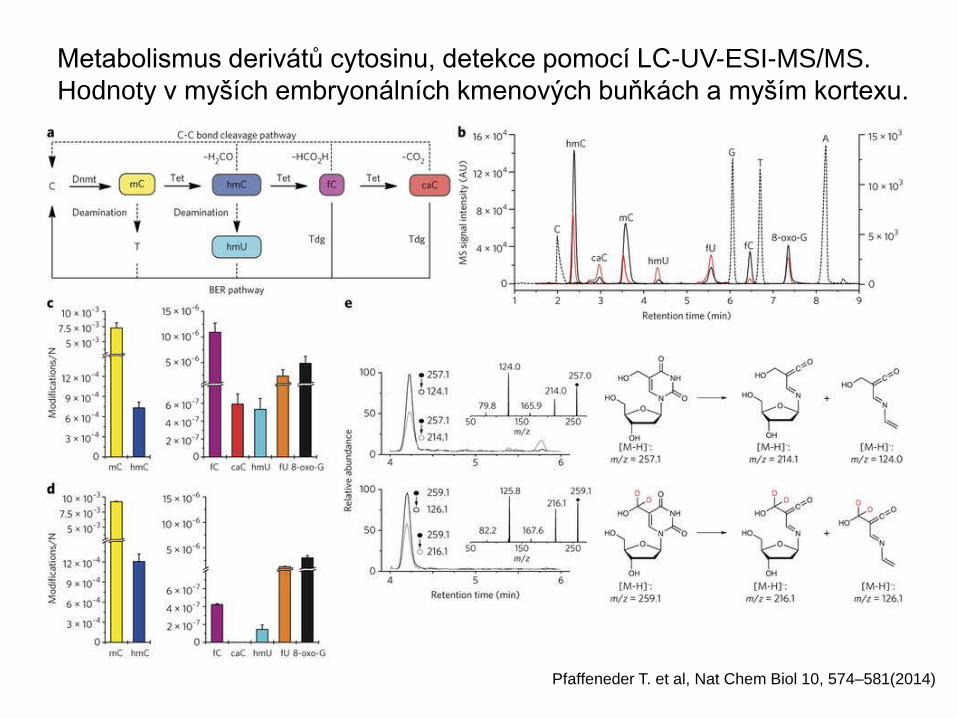
Metabolismus derivátů cytosinu, detekce pomocí LC-UV-ESI-MS/MS.
Hodnoty v myších embryonálních kmenových buňkách a myším kortexu.
Pfaffeneder T. et al, Nat Chem Biol 10, 574–581(2014)
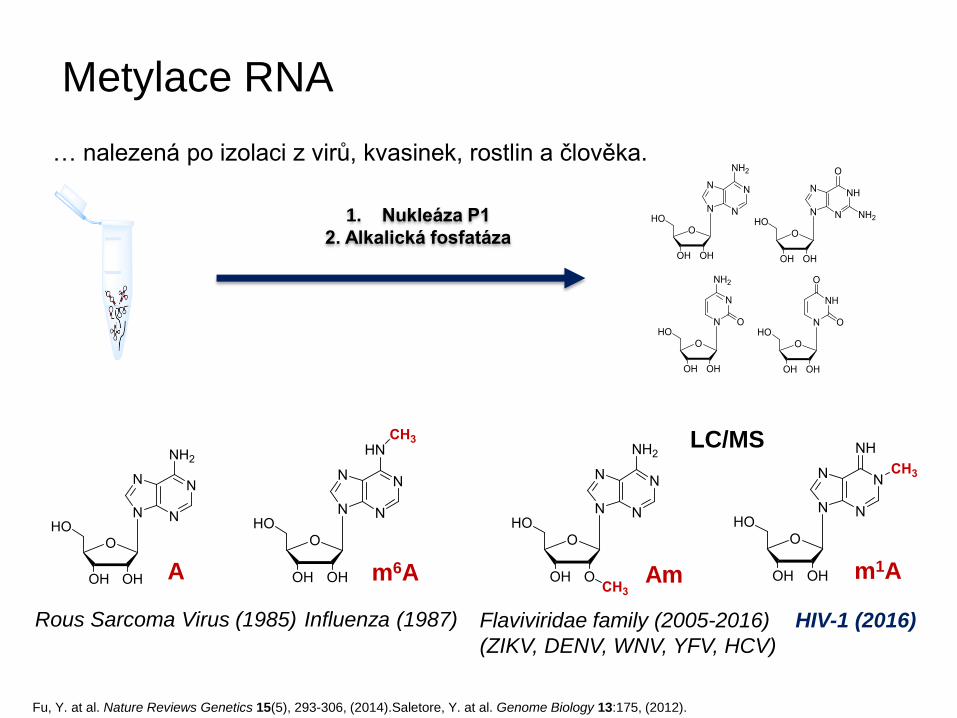
Metylace RNA
… nalezená po izolaci z virů, kvasinek, rostlin a člověka.
Am m6A m1A A
Fu, Y. at al. Nature Reviews Genetics 15(5), 293-306, (2014).Saletore, Y. at al. Genome Biology 13:175, (2012).
Rous Sarcoma Virus (1985) Influenza (1987) Flaviviridae family (2005-2016)
(ZIKV, DENV, WNV, YFV, HCV)
HIV-1 (2016)
1. Nukleáza P1
2. Alkalická fosfatáza
LC/MS
Time5.00 10.00 15.00 20.00 25.00
%
0
100
5.00 10.00 15.00 20.00 25.00
%
0
100
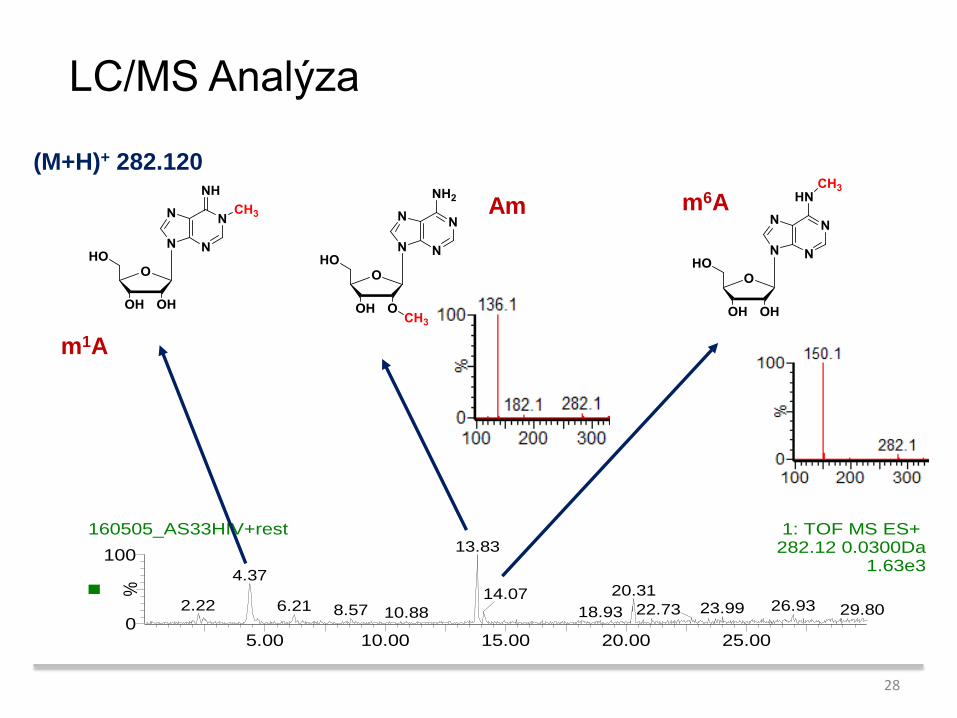
160505_AS33HIV+rest 1: TOF MS ES+ 282.12 0.0300Da
1.63e3
13.83
4.37
2.22 6.21 8.57 10.88
20.3114.07
18.93 26.9323.9922.73 29.80
160505_AS33HIV+rest 1: TOF MS ES+ TIC
5.63e6
1.882.14
28.8825.109.61
2.93 8.515.29 20.5118.0915.06
10.34 21.10 28.4729.73
28
LC/MS Analýza
Am m6A
m1A
(M+H)+ 282.120

Použitá literatura
• Mass Spectrometry, Principles and Application, Second edition – Hoffmann E.,
Stroobant V., 2001, Willey and Sons
• MALDI MS, A Practical Guide to Instrumentation, Methods and Aplication – Hillenkamp
F., Peter-Katalinic J., 2007, Willey-VCH
• CHARACTERIZATION OF NUCLEIC ACIDS BY TANDEM MASS SPECTROMETRY -
THE SECOND DECADE (2004–2013): FROM DNA TO RNA AND MODIFIED
SEQUENCES, Schurch S., Mass Spec. Rev., 2016, 35, 483–523
• MASS SPECTROMETRY ANALYSIS OF NUCLEOSIDES AND NUCLEOTIDES Mass
Spectrometry Reviews, 2014, 33, 302–331 , DOI 10.1002/mas.21388, Ed Dudley and
Liz Bond
• Multimerization rules for G-quadruplexes Kolesnikova S. et al., Nucleic Acids
Research 2017, gkx637
• A Role for the MS Analysis of Nucleic Acids in the Post-Genomics Age, Fabris D. J Am
Soc Mass Spectrom 2010, 21, 1–13
• Tet oxidizes thymine to 5-hydroxymethyluracil in mouse embryonic stem cell DNA,
Pfaffeneder T. et al, Nat Chem Biol 10, 574–581(2014)

















