Svalová vlákna kosterního svalu vzniklasplynutím mnoha buněk v průběhu vývo-je, a proto jsou mnohojaderná, což jimposkytuje velkou plasticitu při reakci naintenzitu a charakter zátěže. Během tétoreakce se „zapínají“ (exprimují) specific-ké geny, které zajišťují produkci dostateč-ného množství metabolických a struktu-rálních proteinů v souladu s charakteremzátěže, to umožňuje zvýšení výkonu svalu
a optimalizaci energetického metabolis-mu. „Zapínání“ může být vyvoláno nejenaktivací svalové kontrakce, tedy nervovoustimulací, ale i působením steroidníchhormonů, což určuje fenotyp svalu (vizdále). Rozdílné fenotypy svalů lze protopozorovat u mužů a žen, a mění se také přivýkonnostním sportu. K ovlivnění feno -typu dochází též za různých patologickýchsituací, jako jsou např. hormonální poru-
chy, mezi něž patří v současné době po -měrně časté změny v hladině thyroidníchhormonů (tyroxin, trijodthyronin) pro-dukovaných štítnou žlázou. Z hlediskafenotypu existují dva hraniční typy sva -lových vláken. Pomalá vlákna jsou málounavitelná, s velkým počtem mitochond-rií a vysoce oxidativním metabolismem.Tato vlákna jsou výhradně zastoupenav tzv. posturálních svalech, které udržujípostavení našeho těla po celý den, anižbychom nad tím přemýšleli. Pomalé svalyse diferencují až po narození a vyvíjejí sepod tonickou aktivitou neuronů (stálouaktivitou nízké intenzity dráždění). Rych-lost kontrakce (stažení) i relaxace (uvol-nění) tohoto typu svalu je pomalejší nežu svalu rychlého, který se naopak vyvíjípod rychlou fázickou nervovou stimulací(střídání vysoké intenzity dráždění s kli-dovou fází) a převažuje v embryonálnímvývoji a časném postnatálním období.Rychlá svalová vlákna se vyznačují niž-ším počtem mitochondrií a mají tedy vyš-ší podíl energie získané bez využití kys -líku anaerobní glykolýzou probíhajícív cytoplazmě, mimo mitochondrie. Svalys převahou těchto glykolytických vlákenjsou schopny krátkého a velkého výkonu,ale snadno se unaví. Produkují ve vyso-kém výkonu nadměrné množství kyselinymléčné, která se štěpí na laktát a vodíkovéprotony, což okyseluje okolní prostředí,a tímto se podílí na svalové únavě. Kyse-lina mléčná je transportována do krvea metabolizována sousedními oxidativní-mi vlákny kosterních svalů a srdcem, kdeslouží jako výhodný zdroj energie využí-vaný mitochondriemi při dostatečnémzásobení kyslíkem. Její zbytek putuje dojater, kde se stane substrátem pro synté-zu glukózy, jež může opět vstupovat dokrve. Tento proces se nazývá Coriho cyk -lus. K produkci laktátu dochází v malémmnožství i v pomalých oxidativních sva-lových vláknech a v srdci, kde je nezbytnýpro další průběh glykolýzy. Experimentál-ní inhibice syntézy laktátu vede k zasta-vení glykolýzy, což vyřadí metabolismusglukózy jako zdroj energie a zastaví svalo -vou kontrakci.
ziva.avcr.cz 4 živa 1/2014
Jitka Žurmanová
Kreatinkinázový systémv kosterním svalu
Možná si ještě pamatujeme, že se kosterní sval skládá ze snopců, ty ze snopeč-ků a snopečky jsou tvořeny svalovými vlákny. Svalová vlákna obaluje buněčnámembrána – sarkolema. Jejich nitro zcela vyplňují kontraktilní proteiny –myofibrily, které umožňují zkrácení svalu. Mezi nimi nacházíme mitochond-rie – semiautonomní organely obklopené dvěma membránami, které za pří-tomnosti kyslíku produkují chemickou energii ve formě adenosintrifosfátu(ATP). Energie ATP je využívaná pro veškeré buněčné procesy. Substrátem protvorbu ATP je většinou glukóza a mastné kyseliny, v období hladovění mohoubýt za tímto účelem degradovány i proteiny z kosterních svalů. Myofibrilyjsou obklopeny membránovým systémem, který je zásobárnou vápenatých ion-tů a nazývá se sarkoplazmatické retikulum. Následující řádky přinášejí infor-maci o významném mechanismu, jenž pomáhá udržovat energetickou rovnová-hu buněk v okamžiku náhlého zvýšení požadavku na dodání energie a zároveňje také schopen usnadnit přenos energeticky bohatých substrátů z místa jejichprodukce k místům spotřeby v buněčném prostoru při vysokém výkonu. Ten-to mechanismus je zvláště dokonale vyvinutý v buňkách schopných vysokéhovýkonu, jejichž požadavky na dodávku energie značně kolísají v čase. Tutoschopnost mají především svalová vlákna, srdeční myocyty, neurony, ale i sper-mie, které se po probuzení k aktivitě pokoušejí vyhrát závod s ostatními, a taksplnit svou biologickou úlohu.
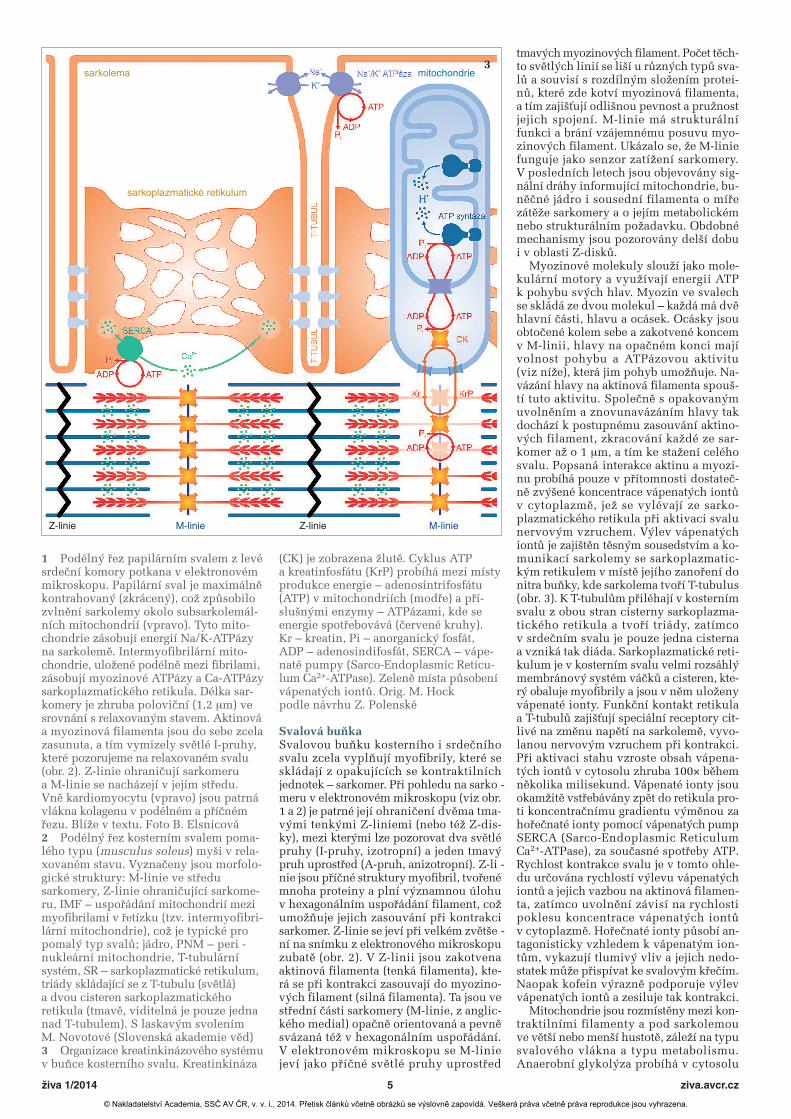
Svalová buňkaSvalovou buňku kosterního i srdečníhosvalu zcela vyplňují myofibrily, které seskládají z opakujících se kontraktilníchjednotek – sarkomer. Při pohledu na sarko -meru v elektronovém mikroskopu (viz obr.1 a 2) je patrné její ohraničení dvěma tma-vými tenkými Z-liniemi (nebo též Z-dis-ky), mezi kterými lze pozorovat dva světlépruhy (I-pruhy, izotropní) a jeden tmavýpruh uprostřed (A-pruh, anizotropní). Z-li -nie jsou příčné struktury myofibril, tvořenémnoha proteiny a plní významnou úlohuv hexagonálním uspořádání filament, cožumožňuje jejich zasouvání při kontrakcisarkomer. Z-linie se jeví při velkém zvětše -ní na snímku z elektronového mikroskopuzubatě (obr. 2). V Z-linii jsou zakotvenaaktinová filamenta (tenká filamenta), kte-rá se při kontrakci zasouvají do myozino-vých filament (silná filamenta). Ta jsou vestřední části sarkomery (M-linie, z anglic-kého medial) opačně orientovaná a pevněsvázaná též v hexagonálním uspořádání.V elektronovém mikroskopu se M-liniejeví jako příčné světlé pruhy uprostřed
tmavých myozinových filament. Počet těch-to světlých linií se liší u různých typů sva-lů a souvisí s rozdílným složením protei-nů, které zde kotví myozinová filamenta,a tím zajišťují odlišnou pevnost a pružnostjejich spojení. M-linie má strukturálnífunkci a brání vzájemnému posuvu myo-zinových filament. Ukázalo se, že M-liniefunguje jako senzor zatížení sarkomery.V posledních letech jsou objevovány sig-nální dráhy informující mitochondrie, bu -něčné jádro i sousední filamenta o mířezátěže sarkomery a o jejím metabolickémnebo strukturálním požadavku. Obdobnémechanismy jsou pozorovány delší dobui v oblasti Z-disků.
Myozinové molekuly slouží jako mole-kulární motory a využívají energii ATPk pohybu svých hlav. Myozin ve svalechse skládá ze dvou molekul – každá má dvěhlavní části, hlavu a ocásek. Ocásky jsouobtočené kolem sebe a zakotvené koncemv M-linii, hlavy na opačném konci majívolnost pohybu a ATPázovou aktivitu(viz níže), která jim pohyb umožňuje. Na -vázání hlavy na aktinová filamenta spouš -tí tuto aktivitu. Společně s opakovanýmuvolněním a znovunavázáním hlavy takdochází k postupnému zasouvání aktino-vých filament, zkracování každé ze sar-komer až o 1 μm, a tím ke stažení celéhosvalu. Popsaná interakce aktinu a myozi-nu probíhá pouze v přítomnosti dostateč-ně zvýšené koncentrace vápenatých iontův cytoplazmě, jež se vylévají ze sarko -plazmatického retikula při aktivaci svalunervovým vzruchem. Výlev vápenatýchiontů je zajištěn těsným sousedstvím a ko -munikací sarkolemy se sarkoplazmatic-kým retikulem v místě jejího zanoření donitra buňky, kde sarkolema tvoří T-tubulus(obr. 3). K T-tubulům přiléhají v kosternímsvalu z obou stran cisterny sarkoplazma-tického retikula a tvoří triády, zatímcov srdečním svalu je pouze jedna cisternaa vzniká tak diáda. Sarkoplazmatické reti-kulum je v kosterním svalu velmi rozsáhlýmembránový systém váčků a cisteren, kte-rý obaluje myofibrily a jsou v něm uloženyvápenaté ionty. Funkční kontakt retikulaa T-tubulů zajišťují speciální receptory cit-livé na změnu napětí na sarkolemě, vyvo -lanou nervovým vzruchem při kontrakci.Při aktivaci stahu vzroste obsah vápena-tých iontů v cytosolu zhruba 100× běhemněkolika milisekund. Vápenaté ionty jsouokamžitě vstřebávány zpět do retikula pro-ti koncentračnímu gradientu výměnou zahořečnaté ionty pomocí vápenatých pumpSERCA (Sarco-Endoplasmic ReticulumCa2+-ATPase), za současné spotřeby ATP.Rychlost kontrakce svalu je v tomto ohle-du určována rychlostí výlevu vápenatýchiontů a jejich vazbou na aktinová filamen-ta, zatímco uvolnění závisí na rychlostipoklesu koncentrace vápenatých iontův cytoplazmě. Hořečnaté ionty působí an -tagonisticky vzhledem k vápenatým ion-tům, vykazují tlumivý vliv a jejich nedo-statek může přispívat ke svalovým křečím.Naopak kofein výrazně podporuje výlevvápenatých iontů a zesiluje tak kontrakci.
Mitochondrie jsou rozmístěny mezi kon -traktilními filamenty a pod sarkolemouve větší nebo menší hustotě, záleží na typusvalového vlákna a typu metabolismu.Anaerobní glykolýza probíhá v cytosolu
živa 1/2014 5 ziva.avcr.cz
3
1 Podélný řez papilárním svalem z levésrdeční komory potkana v elektronovémmikroskopu. Papilární sval je maximálněkontrahovaný (zkrácený), což způsobilozvlnění sarkolemy okolo subsarkolemál-ních mitochondrií (vpravo). Tyto mito-chondrie zásobují energií Na/K-ATPázyna sarkolemě. Intermyofibrilární mito-chondrie, uložené podélně mezi fibrilami,zásobují myozinové ATPázy a Ca-ATPázysarkoplazmatického retikula. Délka sar-komery je zhruba poloviční (1,2 μm) vesrovnání s relaxovaným stavem. Aktinováa myozinová filamenta jsou do sebe zcelazasunuta, a tím vymizely světlé I-pruhy,které pozorujeme na relaxovaném svalu(obr. 2). Z-linie ohraničují sarkomerua M-linie se nacházejí v jejím středu. Vně kardiomyocytu (vpravo) jsou patrnávlákna kolagenu v podélném a příčnémřezu. Blíže v textu. Foto B. Elsnicová2 Podélný řez kosterním svalem poma-lého typu (musculus soleus) myši v rela-xovaném stavu. Vyznačeny jsou morfolo-gické struktury: M-linie ve středusarkomery, Z-linie ohraničující sarkome-ru, IMF – uspořádání mitochondrií mezimyofibrilami v řetízku (tzv. intermyofibri-lární mitochondrie), což je typické propomalý typ svalů; jádro, PNM – peri -nukleární mitochondrie, T-tubulárnísystém, SR – sarkoplazmatické retikulum,triády skládající se z T-tubulu (světlá)a dvou cisteren sarkoplazmatického retikula (tmavě, viditelná je pouze jedna nad T-tubulem). S laskavým svolenímM. Novotové (Slovenská akademie věd)3 Organizace kreatinkinázového systémuv buňce kosterního svalu. Kreatinkináza
(CK) je zobrazena žlutě. Cyklus ATPa kreatinfosfátu (KrP) probíhá mezi místyprodukce energie – adenosintrifosfátu(ATP) v mitochondriích (modře) a pří-slušnými enzymy – ATPázami, kde seenergie spotřebovává (červené kruhy).Kr – kreatin, Pi – anorganický fosfát,ADP – adenosindifosfát, SERCA – vápe-naté pumpy (Sarco-Endoplasmic Reticu-lum Ca2+-ATPase). Zeleně místa působenívápenatých iontů. Orig. M. Hock podle návrhu Z. Polenské
a poskytuje pouze dvě molekuly ATP z jed-né molekuly glukózy, zatímco oxidativnífosforylace (lokalizovaná na vnitřní mito-chondriální membráně) produkuje až 36molekul ATP z oxidace jedné molekulyglukózy nebo dvou molekul laktátu (vizŽiva 2013, 5: 202–205). V mitochondriíchje na konci dráhy oxidativní fosforylaceumístěna ATPsyntáza, využívající energiielektrochemického gradientu vodíkovýchprotonů (H+) na vnitřní mitochondriálnímembráně k syntéze ATP z adenosindifos -fátu (ADP) a anorganického fosfátu (Pi).[Pozn.: Za objasnění mechanismu syntézyATP enzymovým komplexem H+-ATPsyn-tázou získali Paul D. Boyer z Kalifornskéuniverzity v Los Angeles a John E. Wal-ker z Laboratoře pro molekulární biologiiv Cambridge, Velká Británie, v r. 1997 No -belovu cenu za chemii, společně s JensemCh. Skouem z univerzity v Kodani, Dán-sko, za vysvětlení mechanismu činnostiNa/K-ATPázy – viz dále v textu.] Lze tedyříci, že zvýšený vstup ADP do mitochond -rií podporuje tvorbu ATP a je nezbytný profunkci ATPsyntázy. Na druhé straně en -zymy, které mohou ATP štěpit a využítenergii skrytou ve vysokoenergetickýchchemických vazbách této molekuly, se na -zývají ATPázy. Dvě z nich už byly zmí něny,tvoří součást myozinové hlavy – myozi-nová ATPáza a SERCA pumpa na sarko -plazmatickém retikulu. K nim můžemepřidat ještě Na/K-ATPázu na sarkolemě,kde udržuje elektrochemický potenciál.ATPázy zajišťují hydrolýzu ATP a produk-tem této reakce, vedle uvolněné energie,je také ADP a anorganický fosfát. Tyto látkymohou stimulovat ATPsyntázu a současněve vyšší koncentraci inhibovat enzymatic-kou aktivitu ATPáz. ATPázy též produkujíH+ ionty, které lokálně okyselují prostře-dí buňky, a tím omezují její aktivitu. Sní-žené pH negativně ovlivňuje i jiné meta-bolické enzymy, které jsou na změny pHcitlivé. Okyselení buňky se podílí na sva-lové únavě, pokud nejsou H+ ionty efektiv-ně odstraňovány. Obecně platí, že izofor-my ATPáz přítomné v rychlých vláknechjsou méně citlivé na pokles pH ve srovná-ní s vlákny oxidativními, kde docházík podstatně menším výkyvům pH. V rych-lých vláknech může pH klesnout až nahodnotu 6,5 z původní 7,2. Z uvedeného
vyplývá, že akumulace produktů hydrolý-zy ATP (ADP, Pi, H+) silně ovlivňuje výkonsvalového stahu, stejně jako dostupnostATP z mitochondrií nebo z glykolýzy.
Enzym kreatinkináza – CK systémZásoba volného ATP v cytosolu, dostupnápříslušným ATPázám, je poměrně nízká,zhruba 5–8 mM. Představíme-li si rychlostnástupu svalové kontrakce u sprinteraa zvážíme-li koncentraci volného ATP, zdáse, že nutně musí dojít k poklesu volnéhoATP již na počátku výkonu a k následné-mu vychýlení energetické rovnováhy. Zvý-šení produkce ATP glykolýzou a oxidativ-ní fosforylací je stimulováno zvýšenoukoncentrací vápenatých iontů a nastupu-je s určitým zpožděním za počátkem sva-lové práce. Zhruba před 30 lety bylo zjiš-těno, že se hladina ATP během svalovékontrakce nemění. Tento nález vedl k hle-dání mechanismu, který udržuje hladinuATP. Centrem pozornosti se stal kreatin-fosfát, známý již od 30. let minulého stol.,avšak jeho funkce nebyla zcela jasná. Brzypoté byl objeven enzym kreatinkináza(CK), katalyzující přenos fosfátu z krea-tinfosfátu na ADP za vzniku ATP a kreati -nu. Tato reakce je vratná, může tedy pro-bíhat oběma směry s podobnou rychlostív závislosti na koncentraci substrátů a pHv buněčném kompartmentu (např. prostorpod sarkolemou nebo mezi myofibrila-mi). Kreatinkinázovou reakci známe podnázvem Lohmannova reakce.
Nedávné studie ukázaly, že kreatinki-názu v kosterním svalu zastupují dvě izo-formy, mitochondriální a cytosolická. Izo-formy se v buňce nacházejí jak v místechspotřeby energie, tak její produkce. Pokudjde o spotřebu energie, je enzym lokalizo-ván v blízkosti ATPáz a strukturně se vážezhruba na třech strategických místech –v centru sarkomery v M-linii poblíž myo-zinové ATPázy, dále na membráně sarko -plazmatického retikula poblíž vápníkovépumpy SERCA a na sarkolemě poblížNa/K-ATPázy, která zde udržuje iontovourovnováhu (obr. 3). V těchto místech vy -užívá kreatinkináza kreatinfosfát a pro-dukty hydrolýzy ATP (ADP, H+) k syntézeATP. Zajišťuje tak okamžité odstraňovánípotenciálních inhibitorů enzymatické akti-vity ATPáz (ADP, H+) a současně 10× rych-
leji recykluje ATP ve srovnání s jeho syn-tézou. Tím vzniká lokálně vysoký poměrATP/ADP, nezbytný pro dobrý výkon sva-lové kontrakce. Reakce katalyzovaná krea -tinkinázou tedy udržuje stálý vysoký poměrATP/ADP tím, že poskytuje ATP a naopaksnižuje obsah ADP ve svém okolí. Je protopovažována za lokální energetický pufr.
V oblasti produkce ATP v cytosolu, kdeje lokalizovaná glykolýza a v místě oxi-dativní fosforylace, v intermembránovémprostoru mitochondrií, má působení krea-tinkinázy opačný charakter. Reakce zdeprobíhá v obráceném směru, tedy ve pro-spěch syntézy kreatinfosfátu. Jeho moleku -la je ve srovnání s ATP podstatně menší,a proto její difuze z míst produkce k místůmspotřeby probíhá mnohem rychleji. Z to -hoto důvodu se nazývá kreatinkinázovýsystém též prostorovým energetickýmpufrem. Navíc kreatinkináza využívá krea -tin z cytosolu a ATP z mitochondrií v in -termembránovém prostoru mitochondriík produkci ADP, které zde přímo spo -třebovává ATPsyntáza pro tvorbu ATP.ATPsyntáza je tak udržovaná ve vysokémvýkonu. Bylo prokázáno, že tato přímá sti-mulace ATPsyntázy snižuje tvorbu reak-tivních forem kyslíku (tj. superoxidovéhoradikálu a jeho dalších produktů) v mito-chondriích, jež jsou při své nadměrné kon-centraci pro buňku toxické. Zásoba krea-tinfosfátu ve svalové buňce se pohybujemezi 15–30 mM, což pokryje zhruba 8–10sekund vysokého výkonu sprintera. Tatodoba postačuje k aktivaci glykolýzy sti-mulované zvýšenou koncentrací anorga-nického fosfátu uvolněného při hydrolý-ze ATP a vápenatými ionty uvolněnýmiz retikula. Z tohoto pohledu se kreatinki -názový systém též označuje jako časovýenergetický pufr. Je třeba zdůraznit, žev mitochondriích kreatinkinázová reakcetaké produkuje vodíkové protony, což zvy-šuje jejich elektrochemický potenciál navnitřní membráně pro syntézu ATP.
Závěrem lze říci, že kreatinkinázovýsystém patří mezi fosfagenové systémy,kterých se v živočišné říši vyvinulo cel-kem 8. Mají společnou funkci – doplňovathladinu ATP, liší se však chemickýmicharakteristikami. Vedle CK systému se nej-častěji setkáme s argininkinázovým sys -témem, který zahrnuje argininfosfát a argi-ninkinázu a jeho kinetické vlastnosti jsouodlišné. Existují organismy, jež mají obazmíněné systémy, buď odděleně v různýchtkáních, nebo společně v jednom typu tká-ně (např. ježovky ve svalech). Kreatinkiná -zový systém není pro buňky životně důle-žitý, neboť vypnutí genu pro kreatinkinázuu myší bylo úspěšně kompenzováno bě -hem jejich vývoje morfologicky – zmno-žením mitochondrií, a tedy zkrácenímdifuzní vzdálenosti mezi místy produkcea spotřeby energie. Zvířata poměrně dob-ře prospívala, vykazovala ale vyšší svalo-vou únavu. Funkce tohoto systému velmicitlivě reaguje na zvýšenou koncentracivolných radikálů a je v buňce potlačenav případě energetické nouze, kdy se patr-ně jeví jako nadbytečný konzument ATP.Kreatinkinázový systém můžeme tedy po -važovat za modulátor a synchronizátorenergetického metabolismu za fyziolo -gických podmínek a současně dokladdokonalosti přírody.