Page 1
U N I V E R Z I T A K A R L O V A V P R A Z E
P ř í r o d o v ě d e c k á f a k u l t a
K a t e d ra f yz ik á l n í a ma k ro mo l e ku l á rn í ch emi e
OPTIMALIZACE HPLC METODY PRO SEPARACI
ENANTIOMERŮ AMINOKYSELIN OBSAŽENÝCH V BCAA
DOPLŇCÍCH PRO SPORTOVCE
D i p l o m o v á p r á c e
s t u d i j n í h o o b o r u C h e m i e ž i v o t n í h o p r o s t ř e d í
Praha 2008 Simona Srkalová
Page 2
1
Předmětová hesla:
Separace
Aminokyseliny
Chromatografie
Klíčová slova:
Větvené aminokyseliny
HPLC
Teikoplanin
Teikoplanin aglykon
Chirální separace
Page 3
2
Prohlášení
Prohlašuji, že jsem tuto diplomovou práci vypracovala samostatně, pod vedením
školitele Doc. RNDr. Evy Tesařové, CSc. a že jsem všechny použité prameny řádně
citovala.
Jsem si vědoma toho, že případné využití výsledků, získaných v této práci, mimo
Univerzitu Karlovu v Praze je možné pouze po písemném souhlasu této univerzity.
V Praze dne……………………
………………………………………….
podpis
Page 4
3
PODĚKOVÁNÍ
Mé poděkování patří zejména Doc. RNDr. Evě Tesařové, CSc. za odborné
vedení diplomové práce a podnětné rady a připomínky při jejím sepisování. Dále velice
děkuji Mgr. Květě Kalíkové za úvodní zaškolení v chromatografickém systému Breeze
a za veškerý čas, který mi vždy ochotně věnovala.
Na závěr děkuji svým rodičům a Bc. Filipu Baumgartnerovi za velkou trpělivost
a duševní podporu.
Page 5
4
OBSAH
Seznam zkratek a symbolů................................................................................................6
1 Úvod..............................................................................................................................8
2 Teoretický úvod...........................................................................................................9
2.1 Enantiomery - základní pojmy a popis vlastností...................................................9
2.2 Význam enantiomerů v potravinách.....................................................................10
2.3 Aminokyseliny......................................................................................................12
2.4 Potenciální toxicita aminokyselin.........................................................................14
2.5 Sportovní potravinové doplňky............................................................................16
2.5.1 Aminokyseliny s rozvětveným řetězcem.....................................................16
2.5.2 Vliv BCAA na snižování únavy při fyzické námaze...................................19
2.5.2.1 Hypotéza centrální únavy...............................................................19
2.5.2.2 Serotonin.........................................................................................21
2.5.3 Terapeutické využití BCAA........................................................................22
2.6 Principy enantioseparace v HPLC........................................................................24
2.6.1 Chirální stacionární fáze..............................................................................25
2.6.2 Makrocyklická antibiotika...........................................................................25
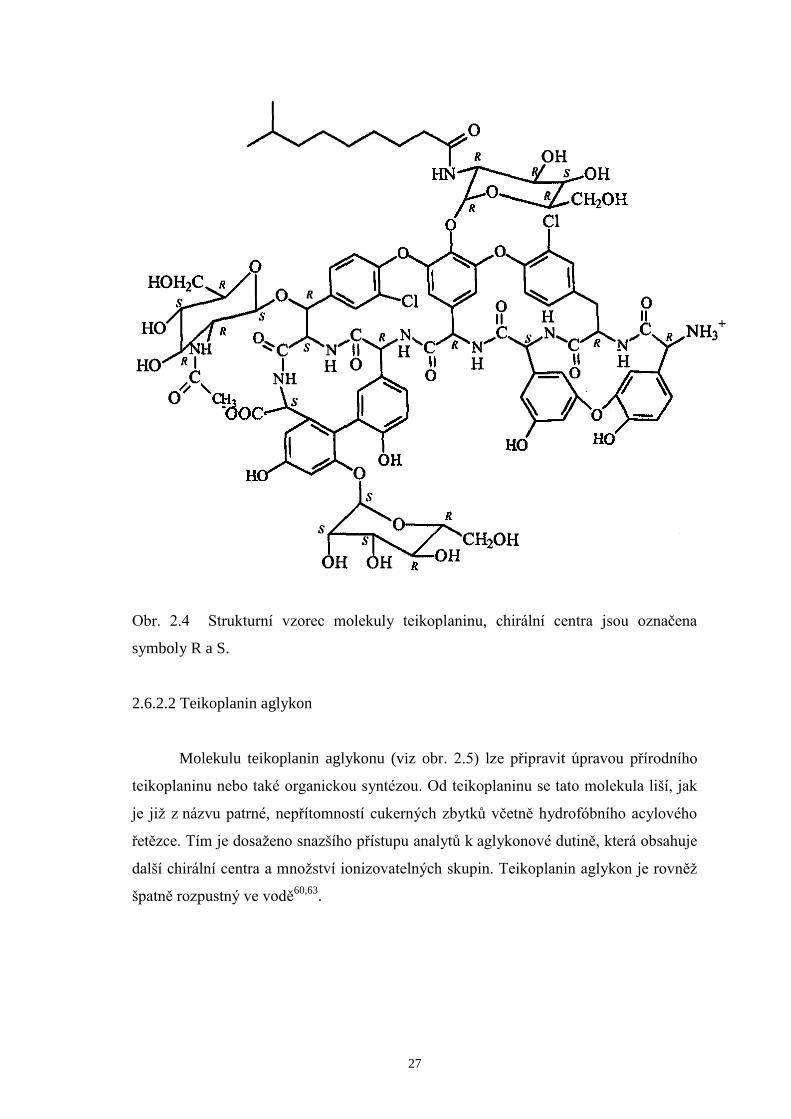
2.6.2.1 Teikoplanin.....................................................................................26
2.6.2.2 Teikoplanin aglykon.......................................................................27
3 Cíl práce......................................................................................................................29
4 Experimentální část...................................................................................................30
4.1 Seznam použitých chemikálií...............................................................................30
4.2 Charakterizace chirálních aminokyselin...............................................................31
4.3 Použité přístroje....................................................................................................31
4.4 Příprava vzorků.....................................................................................................32
4.5 Příprava mobilní fáze............................................................................................32
4.6 Podmínky měření..................................................................................................33
4.7 Zpracování naměřených dat..................................................................................33
4.8 Postup Waltersova testu........................................................................................35
4.9 Racemizační postupy............................................................................................35
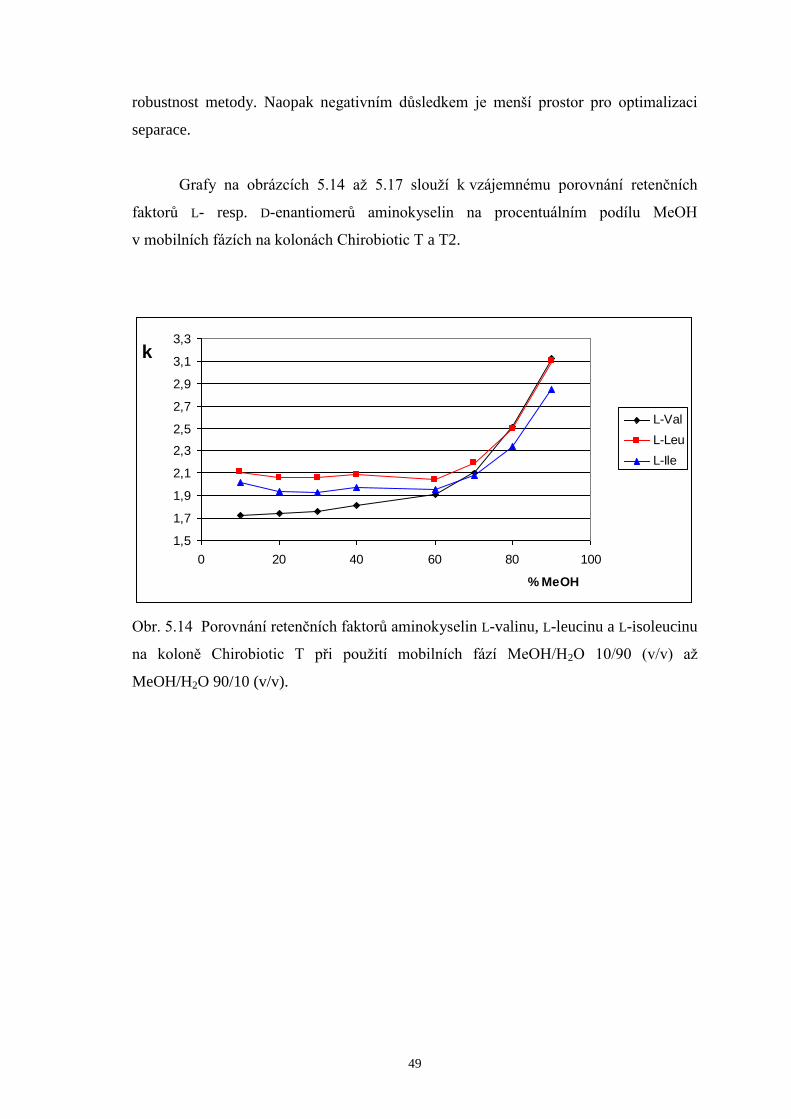
5 Výsledky a diskuze.....................................................................................................36
5.1 Optimalizace metody pro separaci enantiomerů valinu, leucinu a isoleucinu.....36
Page 6
5
5.2 Charakterizace a vzájemné porovnání chirálních stacionárních fází na bázi
teikoplaninu.........................................................................................................43
5.2.1 Waltersův test..............................................................................................43
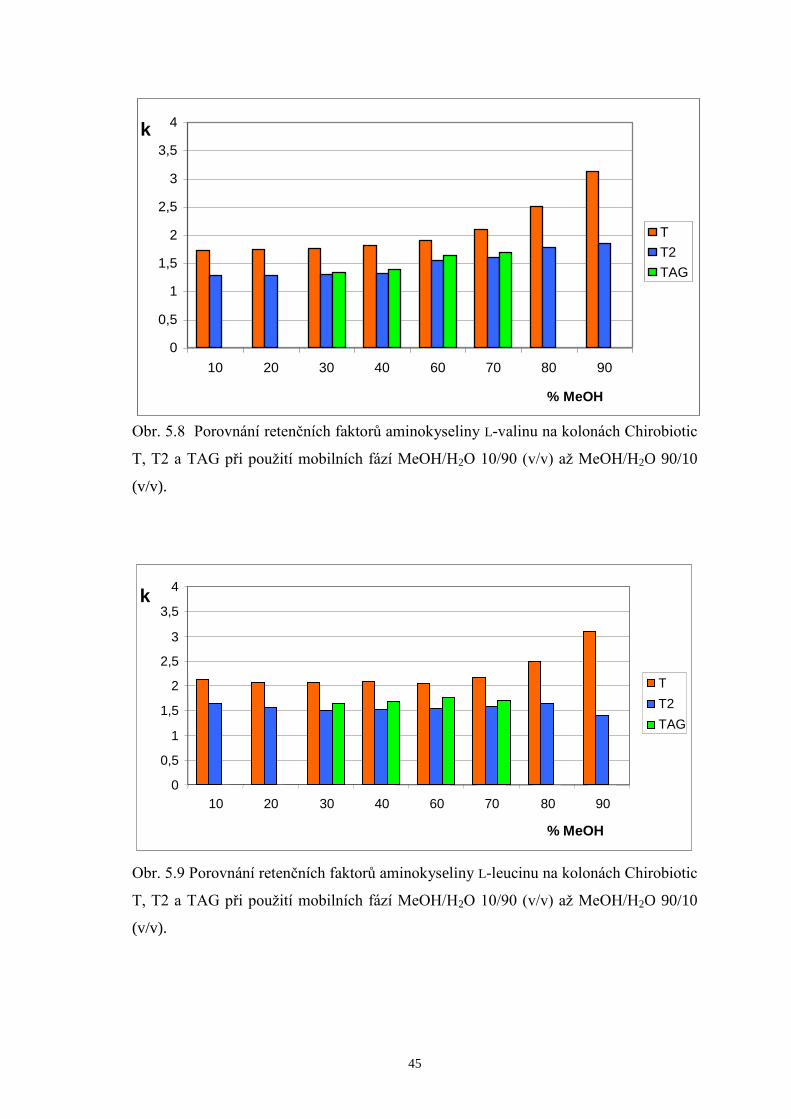
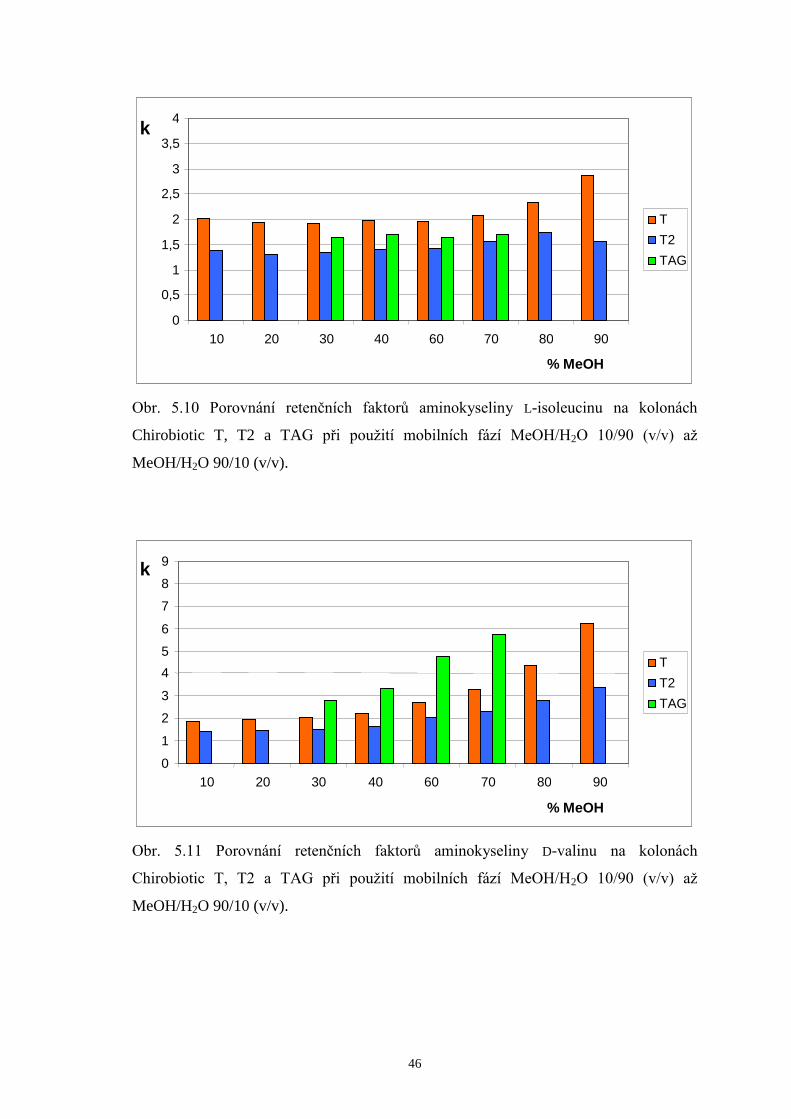
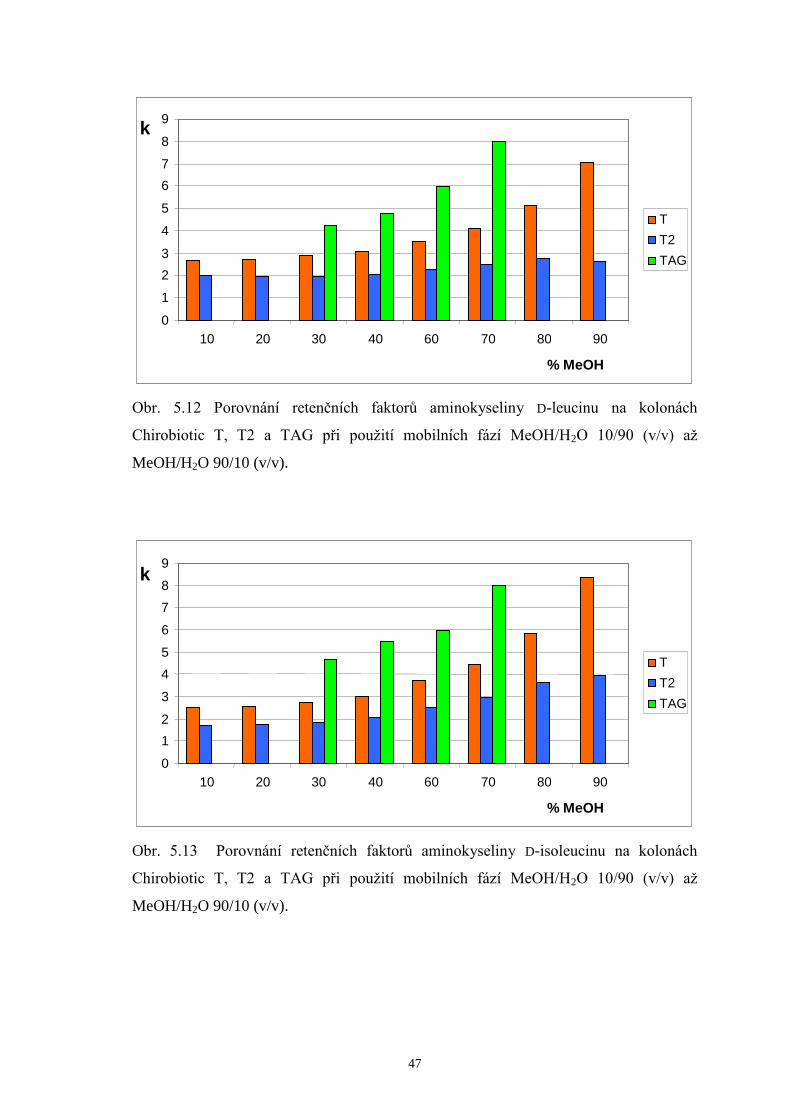
5.2.2 Porovnání retenčních faktorů enantiomerů valinu, leucinu a isoleucinu
na kolonách Chirobiotic T, T2 a TAG.........................................................44
5.2.3 Porovnání separačních faktorů enantiomerů valinu, leucinu a isoleucinu
na kolonách Chirobiotic T a T2...................................................................52
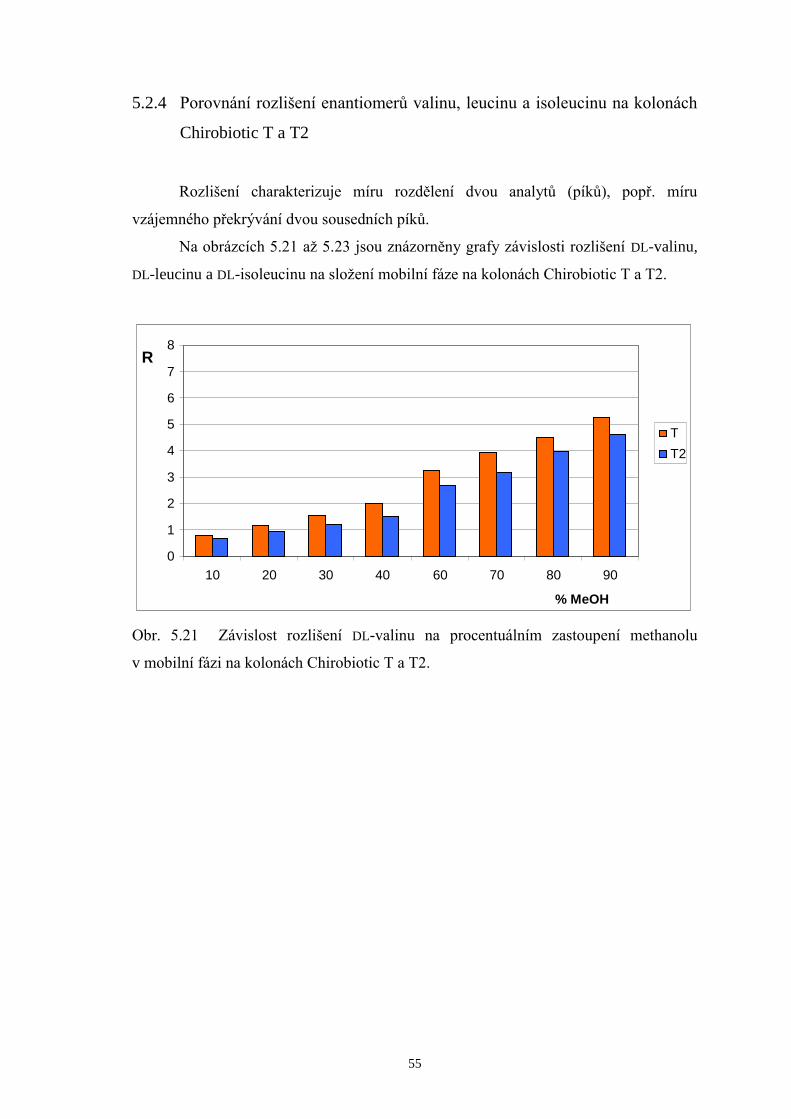
5.2.4 Porovnání rozlišení enantiomerů valinu, leucinu a isoleucinu na kolonách
Chirobiotic T a T2.......................................................................................55
5.2.5 Porovnání symetrie píků enantiomerů valinu, leucinu a isoleucinu
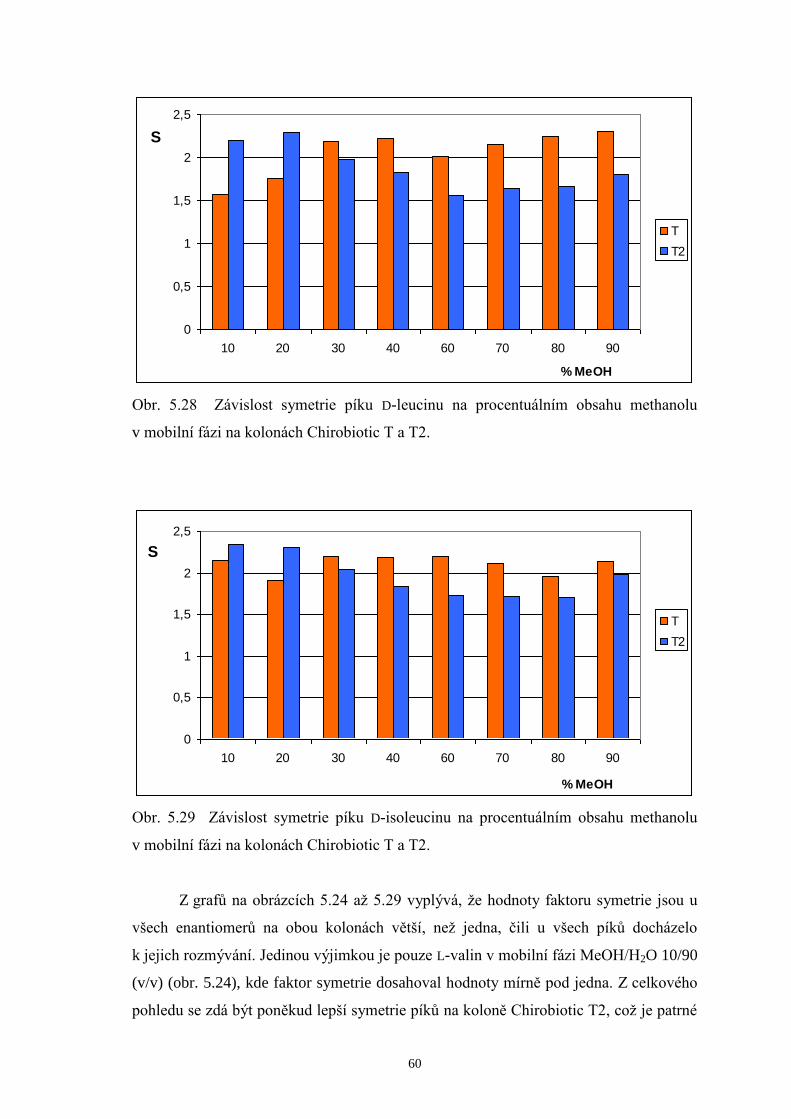
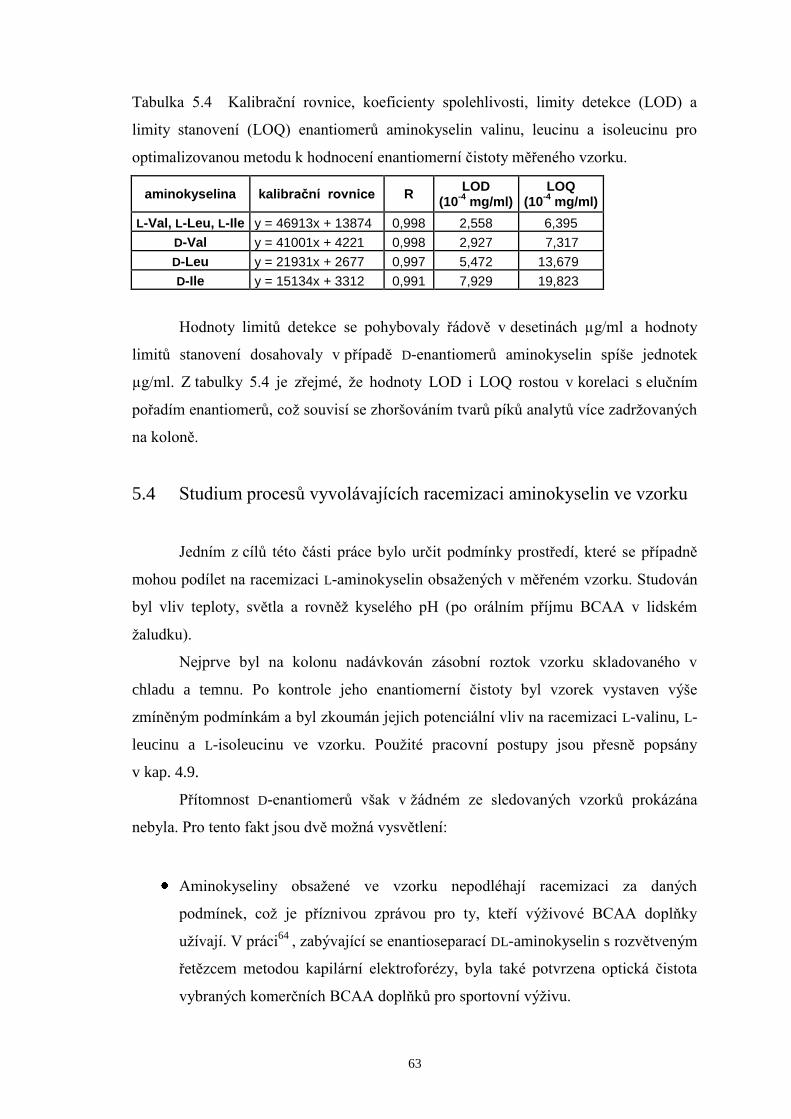
na kolonách Chirobiotic T a T2...................................................................57
5.3 Určení meze detekce a meze stanovení DL-valinu, DL-leucinu a DL-isoleucinu..62
5.4 Studium procesů vyvolávajících racemizaci aminokyselin ve vzorku.................63
6 Závěr...........................................................................................................................65
Seznam literatury.............................................................................................................67
Page 7
6
SEZNAM ZKRATEK A SYMBOLŮ
ACN - acetonitryl
BCAA - Aminokyseliny s rozvětveným řetězcem (z angličtiny Branched Chain Amino
Acids)
CIP - Cahn-Ingold-Prelog
CSP - chirální stacionární fáze
D-Ile - D-isoleucin
D-Leu - D-leucin
D-Val - D-valin
fTrp - volný tryptofan
HI - index hydrofobicity
HPLC - vysokoúčinná kapalinová chromatografie
k - retenční faktor
L-Ile - L-isoleucin
L-Leu - L-leucin
L-Val - L-valin
LOD - limit detekce
log P - rozdělovací poměr
LOQ - limit stanovení
M - mol·dm-3
(jednotka molární koncentrace)
MA - makrocyklická antibiotika
MeOH - methanol
MF - mobilní fáze
Mr - relativní molekulová hmotnost
n - počet teoretických pater kolony
N,N-DETA - N,N-diethyltoluamid
pI 25
- izoelektrický bod při teplotě 25°C
pKA25
- disociační konstanta kyseliny při teplotě 25°C
pKB25
- disociační konstanta zásady při teplotě 25°C
R - koeficient spolehlivosti
R1,2 - rozlišení složek 1 a 2
S - faktor symetrie
Page 8
7
SI - silanolový index
T - teikoplaninová chirální stacionární fáze
T2 - teikoplaninová chirální stacionární fáze s vyšším pokrytím chirálním selektorem
TAG - teikoplanin aglykonová chirální stacionární fáze
tM - mrtvý čas kolony
tR - retenční čas
v/v - objem/objem (poměr objemů složek MF)
w - šířka píku při základně
αij - separační faktor složek i a j
σ - výška šumu základní linie
%obj. - objemová procenta
Page 9
8
1 ÚVOD
Významná část přírodních i uměle připravených sloučenin je chirálních. Je
známo, že v důsledku rozdílného prostorového uspořádání mohou mít enantiomery
často odlišnou biologickou aktivitu a fyziologické vlastnosti. Zejména při aplikaci léčiv
je nutno věnovat enantiomernímu zastoupení patřičnou pozornost, neboť vlivem
rozdílných interakcí chirálních léčiv může mít jeden z enantiomerů požadovaný
terapeutický účinek, zatímco druhý izomer je vysoce toxický. V posledních desetiletích
proto vzrostl zájem o sledování stereoselektivního chování sloučenin a chirální
separační metody dosáhly v oboru analytické chemie významného postavení.
Aminokyseliny se v nedávné době staly populárními výživovými doplňky
užívanými zejména sportovci. Aminokyseliny s rozvětveným řetězcem (BCAA), které
jsou schopné svaly chránit před jejich degradací po náročném tréninku, a mohou se také
podílet na nárůstu svalové hmoty, se proto staly součástí běžně prodávaných
potravinových doplňků. Výše popsané vlastnosti mají pouze L-enantiomery větvených
aminokyselin, proto přítomnost D-forem vzniklých již během výrobního procesu nebo
následně vlivem nevhodného skladování může vést ke snížení výživových hodnot
výrobku. Pro kontrolu optické čistoty BCAA doplňků jsou nezbytným nástrojem právě
analytické separační metody.
Tato práce je zaměřena na optimalizaci metody vysokoúčinné kapalinové
chromatografie, která by umožnila rozdělit a kvantifikovat šest enantiomerů větvených
aminokyselin. Pro tyto účely byly využívány chirální stacionární fáze na bázi
teikoplaninu, proto součástí práce je i jejich charakterizace a vzájemné porovnání
z hlediska retence analytů, selektivity a účinnosti.
Page 10
9
2 TEORETICKÝ ÚVOD
2.1 Enantiomery – základní pojmy a popis vlastností
Enantiomery jsou prostorové izomery, které mají stejnou chemickou strukturu,
ale je mezi nimi vztah jako mezi předmětem a jeho zrcadlovým obrazem. Tyto
molekuly mají v izotropickém (achirálním) prostředí stejné fyzikální a chemické
vlastnosti (teplotu tání, teplotu varu, rozpustnost, spektrum, reaktivitu). Výjimkou je
jejich optická aktivita, nebo-li schopnost otáčet rovinou polarizovaného světla1.
Enantiomery otáčejí rovinu polarizovaného světla o stejný úhel, ale v opačném směru2.
Naměřený úhel závisí na podmínkách měření (koncentraci vzorku, délce kyvety, vlnové
délce použitého světla, použitém rozpouštědle, teplotě). Enantiomery se liší v chirálním
prostředí. Jsou jinak rozpustné v chirálním rozpouštědle, jinak se chovají vůči enzymům
a chirálním činidlům3.
Opticky aktivní jsou látky, které jsou asymetrické1,2,4
. Takové látky nemají
v žádné své konformaci prvek symetrie a nazývají se chirální. Slovo chiralita je
odvozeno ze starořeckého slova chiros – dlaň4.
U sacharidů, aminokyselin a peptidů se k rozlišení absolutní konfigurace na
asymetrickém uhlíku dosud používá označení D- a L- (Fisherova projekce)4. U ostatních
enantiomerů se absolutní konfigurace vyjadřuje pomocí konvence podle autorů Cahna,
Ingolda, a Preloga (CIP systém), která používá označení R- a S- 5.
Směs dvou enantiomerů o stejné koncentraci, bez ohledu na jejich skupenství, se
nazývá racemát2,4
. Protože enantiomery mají stejnou absolutní hodnotu optické
otáčivosti, ale liší se smyslem otáčení, příspěvky obou forem se v racemické směsi
vzájemně ruší, a proto má racemát nulovou hodnotu optické rotace3. Racemizace je
proces, při kterém nevratně vzniká z výchozí neracemické chirální sloučeniny racemát4.
Významná část přírodních látek je chirálních, vyskytují se ve formě
enantiomerů, případně diastereomerů6. Lze říci, že chiralita je základní vlastností živé
hmoty.
Obecně se předpokládá, že přírodní makromolekuly jsou „homochirální“
(molekuly, které obsahují stereogenní centra se stejnou konfigurací)7. Aminokyseliny,
ze kterých se skládají peptidy a bílkoviny, se v organizmech vyskytují převážně v L-
formě. Příčinou „homochirality“ může být fakt, že L-aminokyseliny jsou stabilnější než
Page 11
10
D-formy8. Pro sacharidy, které jsou např. složkou nukleových kyselin, jsou typické D-
izomery. Přírodní výskyt D-aminokyselin nebo L-sacharidů je vzácný nebo je
výsledkem neobvyklých transformací1.
Enantiomery se vyznačují rozdíly nejen v prostorovém uspořádání, ale
v důsledku toho také v chuti a vůni. Např. monoterpen S-(+)-karvon voní jako
kmín, ale R-(–)-karvon voní po mátě. S-limonen voní po citronech, zatímco jeho R-
analog má pomerančové aroma. Enantiomery se také dále mohou lišit ve své biologické
aktivitě a toxicitě. Příkladů lze najít celou řadu. Kyselina askorbová je účinná pouze ve
své S-formě. Léčivo S-propranolol je β-blokátor a R-izomer je neúčinný, R-sotalol je
antiarytmikum, zatímco S-sotalol je β-blokátor, R-penicilamin je antiarytmikum a S-
penicilamin je extrémně toxický1. S-ketoprofen má protizánětlivé účinky, R-enantiomer
se užívá při onemocnění ozubice a může být prodáván jako přísada do zubní pasty7.
Nechvalně známý důsledek aplikace racemátu thalidomidu v 50. a na počátku 60. let
minulého století měl také souvislost se stereochemií9; thalidomid byl nasazen ve 48
zemích jako lék na ranní nevolnosti pro těhotné ženy, vlivem jeho užívání se však
narodilo 12 000 postižených dětí. R-thalidomid má sice sedativní účinky, avšak jeho S-
enantiomer je vysoce teratogenní. Navíc při podání R-thalidomidu dochází k racemizaci
in vivo a vzniká druhý enantiomer s teratogenními účinky10
. Tyto rozdílné biologické a
fyziologické vlastnosti jsou způsobeny stereoselektivními interakcemi mezi
jednotlivými stereoizomery a bílkovinnými receptory1.
Z výše uvedeného je zřejmé, že je zapotřebí věnovat pozornost sledování
stereoselektivního chování či působení sloučenin a z toho vyplývajících důsledků při
aplikaci léčiv, agrochemikálií a při kontrole složek potravin.
2.2 Význam enantiomerů v potravinách
Potraviny obsahují přírodní sloučeniny - některé z nich jsou přítomné pouze jako
čisté enantiomery, jiné se zde vyskytují ve specifickém enantiomerním poměru nebo
jako diastereomery. Chiralita byla shodou okolností poprvé pozorována právě v nápoji
L. Pasteurem, který si všiml, že krystalky soli kyseliny vinné, vytvořené kolem hrdla
lahve od vína, se lišily od těch, které se objevily na dně lahve. Obě rozdílné krystalické
struktury byly později identifikovány jako enantiomery7.
Dlouhou dobu nebyla
významu stereochemie (enantiomernímu zastoupení) u složek potravin věnována
Page 12
11
odpovídající pozornost. Teprve v posledních desetiletích, s rozvojem stereoselektivních
analytických metod, dosáhlo sledování enantiomerů významnějšího postavení i
v potravinářské chemii11,12
.
Chuť a vůně jsou tvořeny komplexní směsí stovek sloučenin, z nichž některé se
vyskytují pouze ve stopových množstvích13
. V potravinách a nápojích bylo
identifikováno množství těkavých látek, jejichž značná část je známá svou chiralitou.
Řada chirálních složek je přirozeného původu, zatímco jiné vznikají během výrobního
procesu (fermentace, sušení, pražení, alkalického zpracování atd.)7. Chuťové a čichové
vnímání je způsobeno interakcí molekuly s receptorem, který je tvořen chirálními
látkami (např. proteiny). Receptor má tedy možnost enantioselektivního rozpoznávání
daného rozdílnou interakcí s enantiomery14
. Např. D-sacharidy jsou kalorické a
kariogenní. L-sacharidy by neměly být kalorické, ani způsobovat zubní kaz, což by
mohlo být využito při uplatnění L-sacharidů jako náhražky sacharózy. Obě enantiomerní
formy sacharidů mají sladkou chuť 15
. Chirální chuťové a vonné složky v přírodních
produktech jsou obecně charakterizovány specifickou distribucí enantiomerů;
sloučeniny nejsou vždy přítomné ve formě jednoho čistého enantiomeru, ale vyskytují
se ve specifickém enantiomerním poměru. Tento poměr se může lišit podle druhu a
stupně zrání, ale i podle zeměpisného původu. Vyhodnocením specifického
enantiomerního zastoupení sloučenin, které nejsou citlivé k racemizaci, mohou být
rozlišeny přírodní a umělé (obvykle racemické) složky přidané do potravin. Detekce
přítomnosti jiných enantiomerů může mít velký význam pro stanovení kvality daného
produktu. Např. proteiny v potravinách mohou racemizovat následkem nevhodného
technologického zpracování, jako je nadměrný ohřev, ozáření nebo úprava za
extrémních hodnot pH. Eventuelně mohou být do produktu přidány syntetické přísady,
čímž dochází k jeho znehodnocení. Během minulých let proto vzrostl zájem o
prověřování enantiomerní čistoty složek potravin. Napomohl tomu i fakt, že
enantiomery mohou mít rozdílné výživové hodnoty a zejména biologickou aktivitu7.
Enantioselektivní analýza může být využita při kontrole technologického
zpracování, identifikaci příměsí, kontrole fermentačních procesů, stáří, hodnocení
mikrobiální kontaminace a skladovacích podmínek, určování zeměpisného původu a při
studiu a kontrole příchutí a aditiv v potravinách a nápojích1,7
.
Page 13
12
2.3 Aminokyseliny
Enantiomery aminokyselin obecně, včetně těch přítomných v potravinách, jsou
jednoznačně nejlépe prozkoumanou skupinou chirálních sloučenin. Je to dáno nejen
četností výskytu, ale i relativně dobře zvládnutou možností stanovení enantiomerů
aminokyselin. Lze říci, že většina stávajících enantioselektivních separačních systémů
umožňuje dobré rozdělení a následnou kvantifikaci enantiomerů aminokyselin11,12
.
Enantiomery aminokyselin se často liší chutí. Například většina hydrofóbních
L-aminokyselin je hořkých, zatímco příslušné D-formy mají velmi sladkou chuť (např.
D-asparagin)1. Čajové aroma a chuť ovlivňují kromě řady dalších těkavých složek také
chirální aminokyseliny. Mezi aminokyselinami běžně se vyskytujícími v čaji má
unikátní postavení theanin (5-N-ethylglutamin)13
. L-theanin je majoritní volnou
aminokyselinou v čaji a tvoří téměř 2 % sušiny z čaje16
. Při analýze několika druhů čaje
bylo zjištěno, že celkové množství theaninu a podíl D-izomeru se mění podle druhu
analyzovaného čaje17
. Sledováním procesů racemizace mohou být získány informace o
výrobě, skladování a přepravě čajových produktů.
D-aminokyseliny mohou sloužit jako vhodné ukazatele podmínek zpracování.
Jsou obsaženy v potravinách a nápojích vystavených vysokoteplotní úpravě,
fermentačním procesům a extrémním hodnotám pH. Všechny aminokyseliny přítomné
v peptidech a proteinech racemizují, avšak rozdílnou rychlostí, což je dáno odlišnými
konstantami racemizace. Rychlost racemizace je dále ovlivňována vnějšími faktory,
jakými jsou právě teplota, pH, ale i koncentrace aj.13
. Bylo prokázáno, že L-
aminokyseliny vázané v proteinech podléhají racemizaci desetkrát rychleji než volné
aminokyseliny. Racemizace aminokyselin během zpracovávání za vysokých teplot nebo
v alkalickém prostředí může být zodpovědná za tvorbu DD-, DL- a LD-peptidových
vazeb. Taková variabilita vazeb následně snižuje schopnost proteolytických enzymů
tyto proteiny trávit, a tím dochází ke snižování nutriční hodnoty daných potravin18
.
Faktory, které ovlivňují racemizaci aminokyselin v proteinech během jejich zpracování,
byly již dříve zkoumány. Výsledky studie19
zabývající se vlivem pasterizace a dalších
vysokoteplotních procesů na racemizaci aminokyselin a proteinů v mléce ukázaly, že
krátkodobé vystavení relativně vysokým teplotám v průběhu zpracování racemizaci
nevyvolalo.
Page 14
13
Pražení kávových a kakaových bobů představuje další příklad vysokoteplotních
procesů, kde lze využít racemizace aminokyselin pro hodnocení kvality finálního
výrobku. Stupeň pražení bývá popisován hodnotou organického úbytku při pražení,
která je zjišťována jako hmotnostní úbytek. Vzájemný vztah mezi stupněm pražení a
racemizací aminokyselin byl popsán matematickou funkcí20
. Tak lze z poměru D/L
enantiomerů a celkového množství aminokyselin zjistit hodnotu organického úbytku při
pražení.
Čerstvě nadojené mléko je většinou kontaminováno mikroorganizmy
přítomnými v bachoru, zejména anaerobními bakteriemi rodů Bacteroides,
Ruminococcus a Butyrivibrio.
Nízké, avšak významné hladiny D-aminokyselin
nalezených v mléce jsou výsledkem trávení a autolýzy bachorových bakterií. D-
aminokyseliny jsou v bakteriích tvořeny z L-analogů za katalýzy racemáz a epimeráz 21
.
D-aminokyseliny by neměly být přítomné v kvalitních nekvašených, nijak
nezpracovaných potravinách. Racemizace aminokyselin je obecně ukazatelem buď
přítomnosti příměsí, a nebo jiného znehodnocení potravin 7.
Enantioselektivní analýza aminokyselin je novým prostředkem pro hodnocení
kvality dalších nekvašených potravinových výrobků, jakými jsou třeba ovocné džusy a
med.7 V ovocných nápojích lze přídavek levné, uměle připravené aminokyseliny
použité k zamaskování zřeďování produktu vodou snadno rozpoznat z přítomnosti
syntetické, tedy obvykle racemické směsi aminokyselin15
. Sedm aminokyselin,
konkrétně DL-arginin, DL-prolin, DL-asparagin, DL-serin, DL-alanin, DL-glutamová
kyselina a DL-asparágová kyselina, je zodpovědných za 90 % veškerého obsahu
aminokyselin nalezených v pomerančových džusech22
. D-arginin, D-alanin, D-
glutamová kyselina a D-asparágová kyselina byly však detekovány v pomerančových
džusech rovněž jako přirozeně se vyskytující D-enantiomery z rostlinných zdrojů, půdy
nebo mikroorganizmů23
. Významné hladiny D-aminokyselin byly nalezeny také v medu.
Určení enantiomerního poměru vybraných aminokyselin může být použito při
zjišťování doby skladování a podmínek, za kterých byl med vyráběn. Enantioselektivní
analýza aminokyselin však také může poskytnout informace o bylinném a zeměpisném
původu medu24
. Nejvýznamnější aminokyselina v medu je L-prolin, jehož obsah činí 50
až 85 % z celkového množství volných aminokyselin. Zpracováním medu za vysokých
teplot se jednak snižuje celkové množství aminokyselin, a zároveň se zvyšuje relativní
zastoupení D-aminokyselin13
.
Page 15
14
Výskyt D-aminokyselin v kvašených potravinách je přirozený. Důvodem je
jednak přítomnost D-aminokyselin v buněčné stěně bakterií, odkud se mohou uvolňovat
do okolí, a dále pak působení bakteriálních racemáz či epimeráz, které přeměňují L-
aminokyseliny na jejich D-analogy25
. V sýru, jogurtu, víně, octu atd. jsou ve větším
množství zastoupené D-alanin, D-glutamová kyselina a D-asparágová kyselina.
Významné je zastoupení těchto aminokyselin ve zrajících sýrech. Stejné D-
aminokyseliny převládají také v jogurtech. Množství těchto enantiomerů je v korelaci s
bakteriálním nárůstem Streptococcus thermophilus a Lactobacillus bulgaricus. Zvláště
obsah D-alaninu, který je produkován pouze bakterií L. bulgaricus, je používán jako
platný rychlý ukazatel pro měření koncentrace L. bulgaricus v jogurtu26
.
D-aminokyseliny mohou vznikat v potravinách nejen činností mikroorganizmů,
ale také v kyselém prostředí z L-analogů za přítomnosti glukózy, fruktózy nebo
sacharózy27
. Hlavní zdroje D-aminokyselin pro člověka jsou potraviny a nápoje. Dále
získává člověk D-aminokyseliny degradací buněčných stěn mikroorganizmů ve střevě a
de novo syntézou28
. Pouze 10-20 % přijatých D-aminokyselin je vyloučeno, zbylých 80-
90 % je zachyceno ve střevech a přeměněno na α-oxokyseliny za působení oxidázy D-
aminokyselin, která je přítomná zejména v játrech, ledvinách a mozku29
. Toxicita D-
aminokyselin pro lidský organizmus nebyla dosud jasně prokázána, také jejich
metabolická dráha není dodnes zcela známá19
. Nahromadění D-aminokyselin
v organizmu, způsobené nízkou aktivitou oxidázy, však může vyvolat řadu poškození,
např. inhibici enzymové syntézy nebo potlačení růstu30
. Metabolizmus těchto
enantiomerů může být ovlivněn i dalšími faktory, jako např. rychlostí transportu,
činností střevních enzymů a bakterií, absorpcí a ledvinovým oběhem7. Při
intravenózním podání čisté racemické směsi aminokyselin dospělým osobám i dětem se
neprojevily žádné toxické účinky, přestože podaná dávka obsahovala vyšší koncentraci
aminokyselin, než je množství přijímané v běžné potravě21
.
2.4 Potenciální toxicita aminokyselin
Toxicita aminokyselin samotných není známá (kromě zažívacích obtíží
vyvolaných příjmem jejich nadměrného množství), nicméně jejich sekundární
metabolity nebo produkty degradace v potravinách již s sebou mohou přinášet řadu
závažných zdravotních rizik.
Page 16
15
Rostliny a také některé jiné organismy jsou schopné rozkladem kyanogenních
sloučenin (tzv. kyanogeneze) produkovat kyanovodík. Předpokládá se, že tyto
sloučeniny plní v rostlinách ochrannou funkci před škůdci a predátory.
Nejrozšířenějšími kyanogeny jsou kyanogenní glykosidy, jejichž molekula je většinou
složena z monosacharidu β-D-glukosy (výjimečně disacharidu) a na něj navázané
necukerné složky. Kyanogenní glykosidy se obvykle dělí podle aminokyselin, ze
kterých vznikly biosyntézou. Nejjednodušším kyanogenním glykosidem je linamarin
odvozený od valinu. Je obsažen v manioku (Manihot esculenta), který je významnou
součástí jídelníčku obyvatel subsaharské Afriky a Indonésie31
.
Dekarboxylací příslušných aminokyselin vznikají biogenní aminy, které jsou
v nízkých koncentracích přirozenou složkou řady potravin. V živočišných tkáních a
rostlinných pletivech vykonávají mnoho důležitých funkcí. V živočišných produktech
jsou hlavními biogenními aminy histamin (vznikající z histidinu), kadaverin (z lysinu),
putrescin (z ornitinu) a tyramin (z tyrosinu). Při skladování masa, ryb a sýrů dochází ke
zvyšování jejich obsahu vlivem enzymové aktivity přítomných mikroorganismů. Obsah
některých biogenních aminů lze proto využít jako ukazatele čerstvosti masa. Růst
koncentrace biogenních aminů je především ovlivněn teplotou při skladování a druhem
kontaminující mikroflóry. Potraviny, které mají vysoký obsah biogenních aminů, jsou
zdravotně závadné; jejich konzumace vyvolává zvracení, dýchací obtíže, pocení, bušení
srdce, hypotenzi nebo hypertenzi a migrény. Působením monoaminooxidas a
diaminooxidas je část biogenních aminů v lidském organismu odbourána, avšak vysoké
koncentrace těchto látek není tělo schopné odstranit. Koncentrace histaminu přesahující
500 – 1000 mg . kg-1
jsou pro člověka považovány za nebezpečné. Zvýšené množství
tohoto aminu může vyvolat až anafylaktický šok31
.
Zatímco v rozvojových zemích se na vzniku nádorů nejvíce podílejí chronické
infekce virové, mikrobiální a parazitární (celosvětově vyvolávají 20 – 25 % všech
nádorů), v hospodářsky vyspělých státech je to zejména strava, složená především
z rafinovaných potravin s převahou tuku, cukru, výrobků z bílé mouky, velkého podílu
živočišných a malého podílu přirozených rostlinných potravin. Vnější faktory se mohou
u lidí spolupodílet na vzniku až 90 % nádorů32
.
Dusitany přidané do potravin nebo vzniklé bakteriální redukcí z dusičnanů (v
dutině ústní nebo v žaludku) reagují za určitých podmínek se sekundárními aminy nebo
amidy, které v potravě vznikají degradací bílkovin a jiných dusíkatých potravinových
složek a tvoří nitrosaminy nebo nitrosamidy, které jsou klasifikovány jako mutageny a
Page 17
16
potenciální karcinogeny. Nitrosaminy také vznikají ve kvašených potravinách a jsou
produkovány rovněž během tepelné úpravy potravin, zejména při smažení tučných
masných výrobků. N-Nitrososloučeniny se mohou tvořit z dusičnanů a dusitanů a
aminoskupin i přímo v lidském těle při pH 3 – 7. Snížení produkce nitrosaminů
v potravinách lze dosáhnout přídavkem vitaminu C (přibližně 500 mg . kg-1
) nebo
vitaminu E 33
.
Heterocyklické aminy patří do skupiny velmi škodlivých látek, které mají
genotoxické účinky a u hlodavců (krysy a myši) byla prokázána jejich mutagenní a
karcinogenní aktivita. Tyto sloučeniny vznikají rozkladem proteinů nebo jednotlivých
aminokyselin, především tryptofanu, fenylalaninu a kyseliny glutamové. Tvoří se
během tepelné úpravy potravin. Z každé aminokyseliny vzniká jeden nebo více
heterocyklických aminů33
.
2.5 Sportovní potravinové doplňky
2.5.1 Aminokyseliny s rozvětveným řetězcem
Aminokyseliny s rozvětveným řetězcem jsou označovány jako BCAA
(z angličtiny Branched Chain Amino Acids) – jedná se o valin, leucin a isoleucin.
Obr. 2.1 Strukturní vzorec aminokyseliny L-valinu
Obr. 2.2 Strukturní vzorec aminokyseliny L-leucinu
Page 18
17
Obr. 2.3 Strukturní vzorec aminokyseliny L-isoleucinu
Tyto tři aminokyseliny patří do skupiny tzv. esenciálních aminokyselin, tedy pro
člověka nepostradatelných a je proto nutné, je v nezbytných množstvích dodávat do
organismu potravou. V klinické medicíně se větvené aminokyseliny uplatňují všude
tam, kde je potřeba chránit vlastní bílkoviny organismu před devastací34
. Struktura
myofibrilárních bílkovin je přibližně z 18 % tvořena právě aminokyselinami
s rozvětveným řetězcem35
(některé zdroje34
uvádí až z 35 %). Tyto aminokyseliny
představují okamžitý zdroj energie, neboť jsou využitelné přímo z krevního oběhu,
odkud přecházejí do svalových buněk. Jsou proto schopné svaly chránit před úbytkem
svalové hmoty (antikatabolický efekt) a mohou také přispívat k nárůstu objemu svalů.
Jejich přirozené množství však nevystačí např. při náročném tréninku, následkem je
devastace svalové hmoty související s katabolickými procesy. Z těchto důvodů jsou
rozvětvené aminokyseliny vhodnými potravinovými doplňky pro sportovce (zejména
kulturisty), případně osoby s nadměrnou fyzickou zátěží34,36
. Doplňkový příjem L-
valinu, L-leucinu a L-isoleucinu může být užitečný i v určitých situacích – např. pro
prevenci ztráty svalů ve vysokých nadmořských výškách a zvyšování odolnosti při
výkonech prováděných v teplém prostředí. Mohou také zlepšit stav při úbytku
duševních schopností, vzniklých v důsledku záteže37
(viz kap. 2.5.2.1).
Aminokyseliny s rozvětveným řetězcem zaujímají mimořádné postavení
v metabolismu. V metabolismu těchto aminokyselin lze najít řadu analogií
s odbouráváním mastných kyselin. Katabolismus větvených aminokyselin v organismu
se skládá ze tří základních kroků: transaminace, oxidační dekarboxylace a třetího kroku,
který je podobný β-oxidaci mastných kyselin. Pro rozklad větvených aminokyselin je
typické, že se jen nepatrně metabolizují v játrech. Jsou totiž zpracovávány na periferii,
zejména ve svalu, pro který představují vynikající zdroj energie. Při užívání BCAA
dochází k mnohem nižší tvorbě kyseliny mléčné, což znamená, že tělo při výkonu
využívá menší množství svalového glykogenu k tvorbě energie. Dalším důležitým
faktem je, že doplňování větvených aminokyselin po sportovním výkonu udržuje
Page 19
18
hladinu glutaminu v krvi (snížená hladina glutaminu v krvi způsobuje oslabení
obranyschopnosti a organismus je tak více náchylný vůči infekcím). Jak tyto
aminokyseliny samotné, tak z nich odvozené produkty mají zvláštní terapeutický
význam – používají se při chorobách jater, svalů a při selhání ledvin (viz kap. 2.5.3). L-
valin, L-leucin a L-isoleucin jsou součástí umělých diet a mají kladný vliv na
metabolismus bílkovin – lze jej označit za antikatabolický, jelikož vedou ke zvýšení
syntézy bílkovin v srdci a játrech34
. Při studiu účinků BCAA bylo rovněž zjištěno, že
doplňkový příjem BCAA přispívá k redukci břišního tuku, naopak efekt na svalový
objem byl nulový, pokud byly tyto doplňky kombinovány s dalšími aminokyselinami38
.
K nejdůležitějším funkcím BCAA z hlediska doplňkového příjmu osobami
s nadměrnou fyzickou zátěží patří zejména tyto34,36
:
Uplatňují se všude tam, kde je zapotřebí chránit vlastní bílkoviny organismu
před devastací.
Jsou užitečné v období svalové regenerace, kdy urychlují novou tvorbu bílkovin.
Jsou rychle vstřebatelné, čímž představují okamžitý zdroj energie.
Pomáhají léčit svalová zranění.
Spolu se svými ketoanalogy jsou velice často používány k léčbě jaterních a
ledvinových onemocnění.
Z běžných potravin je největší množství větvených aminokyselin obsaženo
v kuřecím a krůtím mase, v mořských rybách, mléčných výrobcích a luštěninách. Strava
většiny lidí poskytuje dostatečné množství BCAA, což je okolo 25 – 65 mg / kg / den.
Denní spotřeba leucinu je ze všech tří aminokyselin nejvyšší a pro mladého
nesportujícího jedince se pohybuje v rozmezí 20 – 40 mg / kg / den. Pro sportovce
trénujícího denně tři a více hodin pak vychází dávka minimálně na 60 mg / kg/ den.
Výše spotřeby valinu byla stanovena mírně pod hranicí 40 mg / kg / den, pro intenzivně
trénující jedince je doporučována dávka minimálně 50 mg / kg / den. U isoleucinu se
spotřeba stanovuje velice těžko kvůli obtížnosti měření jeho štěpení, nicméně odborníci
doporučují denní dávku zhruba 20 mg / kg / den. Ideální poměr zastoupení jednotlivých
aminokyselin (v pořadí leucin, isoleucin a valin) je 2 : 1 : 0,5 34
. Právě leucin se výrazně
podílí na zrychlení nárůstu svalové hmoty38
- syntézu bílkoviny v organismu nastartuje
Page 20
19
mnohem dříve než ostatní aminokyseliny. Protože je celý tento proces závislý na
hladině inzulínu, nemůže pracovat sám, bez dalších aminokyselin a sacharidů.
Celková denní dávka BCAA by neměla překročit 20 g (vyšší množství se sice
nepovažuje za škodlivé, ale může způsobit zažívací obtíže). Denní dávka je součtem
množství větvených aminokyselin ze všech zdrojů přijímaných během dne, tzn. ze
stravy a z doplňků. Ideální je dané množství rozdělit do minimálně šesti denních porcí a
užívat je jak ve dnech tréninku, tak ve dnech regenerace. Je doporučováno užít 5 g
BCAA zhruba 60 minut před tréninkem a dalších 5 g ihned po tréninku spolu s potravou
bohatou na proteiny. V souvislosti s užíváním BCAA nebyly zaznamenány žádné
vedlejší účinky34
.
2.5.2 Vliv BCAA na snižování únavy při fyzické námaze
2.5.2.1 Hypotéza centrální únavy
Fyzická únava je definována jako neschopnost udržet silový výkon a může být
původem centrální nebo periferní. Faktory související s únavou jsou ovlivňovány
intenzitou a délkou cvičení, nutričním příjmem a fyzickou kondicí konkrétního
jedince39
. Příčinami periferní únavy mohou být např. vyčerpání svalového glykogenu
nebo fosfokreatinu, nahromadění H+ iontů (laktátu) nebo nedostatečný nervově svalový
přenos. Faktory, které by mohly být příčinou centrální únavy, jsou méně známé, avšak
několik mechanismů, souvisejících s centrální únavou, již bylo navrženo. Prvním z nich
může být nárůst koncentrace některých klíčových sloučenin ve svalech během tělesné
aktivity, např. H+ iontů, K
+ iontů, bradykininů, fosfátů, prostaglandinů, které se mohou
navázat na specifické receptory ve svalech. Touto vazbou pak mohou být přenášeny
informace ze svalu do mozku za pomoci senzorických nervů. Dalším z mechanismů
může být vyčerpání zásoby glykogenu v játrech, následný pokles hladiny glukózy
v krvi, snížení této hladiny v mozku a z toho vyplývající omezené využití glukózy
některými neurony v těch částech mozku, které jsou zodpovědné za kontrolu motorické
aktivity. Posledním z mechanismů podílejících se na vzniku centrální únavy je nárůst
koncentrace aminokyseliny tryptofanu v krvi, s čímž souvisí zvýšení koncentrace
neurotransmiteru 5-hydrohytryptaminu (serotoninu) v některých neuronech,
zodpovídajících za kontrolu motorické aktivity v mozku40
.
Page 21
20
Množství tohoto neurotransmiteru v presynaptickém neuronu závisí na rychlosti
jeho syntézy, která je regulována koncentrací tryptofanu v krvi (příjem tryptofanu
mozkem je významným faktorem při řízení syntézy 5-hydroxytryptaminu a tedy jeho
množství v presynaptickém nervu)40
a transportem tryptofanu přes hematoencefalickou
bariéru39
. Tyto procesy pak vedou ke vzniku únavy40
. První studie, která poukázala na
to, že koncentrace 5-hydroxytryptaminu je ovlivňována fyzickým cvičením, byla
publikována v roce 1963 Barchasem a Freedmanem41
. Ti zjistili zvýšené množství
serotoninu v mozku krys, které před tím plavaly až do naprostého vyčerpání.
Transport tryptofanu do mozku není řízen pouze koncentrací této aminokyseliny
v krevním řečišti, ale také množstvím dalších aminokyselin, zejména aminokyselin
s rozvětveným řetězcem, které při transportu přes hematoencefalickou bariéru
s tryptofanem soupeří 38,40
. Během delšího fyzického výkonu jsou BCAA oxidovány v
pracujícím svalu a jejich koncentrace v plasmě klesá. Pokud se během cvičení zvyšuje
v plasmě hladina volných mastných kyselin, které s tryptofanem soutěží o vazebné
místo na albuminu (tryptofan je jedinou aminokyselinou navázanou v krevní plasmě na
albumin; pouze 10% z celkové koncentrace v plasmě se vyskytuje ve volné formě39
),
dochází rovněž k růstu koncentrace volného tryptofanu v plasmě. Rostoucím poměrem
mezi volným tryptofanem a BCAA (fTrp/BCAA) v plasmě, který je pozorován během
námahy a zejména po ní, je upřednostňován transport tryptofanu do mozku. Cvičením
tedy může být zvýšena rychlost syntézy, koncentrace a uvolňování serotoninu
z některých neuronů zodpovědných za únavu během delší námahy40
.
Orálním podáváním BCAA roste jejich koncentrace v krevní plasmě, tím
dochází ke snížení poměru koncentrací fTrp/BCAA, a tedy nižšímu transportu
tryptofanu k presynaptickým neuronům v mozku. Tím klesá podle této hypotézy
syntéza serotoninu v mozku, a následně tak může být redukována velikost únavy38,40
.
Zmiňovaná hypotéza ovšem nemusí být zcela správná. Např. lze namítnout, že
pozitivní efekt orálně podávaných větvených aminokyselin během namáhavého cvičení
nesouvisí se snižováním koncentrace 5-hydroxytryptaminu v mozku, nýbrž s jinými
biochemickými procesy v centrální nervové soustavě. Eventuálně lze podotknout, že
BCAA (zejména pak leucin) mohou být přeměněny na metabolit, který může fungovat
jako nový neurotransmiter s účinky snižujícími únavu40
. Na druhou stranu infuze
větvených aminokyselin v pacientech s cirhózou jater zamezila abnormálnímu příjmu
tyrosinu mozkem42
.V další studii43
bylo rovněž naznačeno, že zvýšením koncentrace
valinu v krvi se snížil transport aromatických kyselin - tyrosinu a tryptofanu přes
Page 22
21
hematoencefalickou bariéru. Některé studie na lidských jedincích podporují teorii
zapojení 5-hydroxytryptaminu při vzniku únavy44,45
, jiné nikoliv46,47
. Tyto rozpory
mohou být vysvětleny rozdíly v lécích užívaných konkrétní osobou, dávkách BCAA,
době podání dávky, individuálních odchylkách v neuroendokrinní odezvě, stejně tak
jako v typu, intenzitě a délce cvičení39
.
Dalším zajímavým faktem je, že příjem sacharidů během fyzického výkonu
snižuje v krevní plasmě kompetici o vazebné místo na albuminu mezi volnými
mastnými kyselinami a volným tryptofanem38
. Tímto procesem je inhibován cvičením
indukovaný nárůst množství volného tryptofanu v plasmě, a tím také transport
tryptofanu do mozku39
. Tak může být opožděn možný pozitivní efekt BCAA při únavě.
Naopak při studii48
zkoumající vliv infuze větvených aminokyselin na tělesný výkon
jedince ve stavu vyčerpaného svalového glykogenu (dosaženého krátkodobým půstem)
nebyl zjištěn žádný pozitivní efekt podaných BCAA na svalovou výdrž. Doplňkový
příjem aminokyselin s rozvětveným řetězcem měl dokonce škodlivý účinek na tělesný
výkon u pacientů s nedostatkem enzymu glykogen fosforylasy v důsledku rostoucí
produkce amoniaku49
.
2.5.2.2 Serotonin
Biologicky aktivní látka serotonin byla objevena v krevním séru a jako první byl
prokázán její vliv na napětí hladkého svalstva - tonus. Název látky proto vznikl
složením slov sérum a tonus50
. Poprvé byla izolována Irvinem Pagem v roce 1948 a
později byla dána do souvislosti s centrální nervovou soustavou. V organismu
průměrného lidského jedince je obsaženo pouze 5 – 10 mg serotoninu, z toho 90 % se
nalézá ve střevě a zbytek v krevních destičkách a mozku51
.
Významnou funkcí 5-hydroxytryptaminu je přenos nervových vzruchů
(neurotransmiter), podílí se na procesech souvisejících se vznikem nálad50
, kontrolou
chuti, pamětí a učením, termoregulací, chováním, kardiovaskulárními funkcemi,
svalovými kontrakcemi a endokrinní regulací51
. Serotonin podporuje kontrakce
hladkého svalstva a krevní srážlivost. Proto hraje značnou roli při krvácivých
poraněních, kdy vasokonstrikční účinek snižuje únik krve z těla. Serotonin je také
spojován se vznikem únavy50
.
Nedostatek serotoninu způsobuje snížení přenosu nervových vzruchů –
způsobuje tak změny nálad, depresi, popřípadě poruchy spánku, podrážděnost až
Page 23
22
agresivitu. Změny v jeho metabolismu mohou být zodpovědné také za určité psychické
poruchy – např. schizofrenii, maniodepresivní psychózu, chorobnou úzkost a další.
Nedostatek této biomolekuly lze upravit příjmem potravy, která obsahuje bílkoviny
bohaté na L-tryptofan. Serotonin v těle vzniká z této aminokyseliny působením enzymů.
Na syntézu serotoninu v těle má také velký vliv hladina světelného záření. Proto se jeho
nedostatek projevuje zejména v zimním období, kdy lidé častěji trpí úzkostnými stavy,
depresemi, jsou podráždění a útoční. Zvýšená hladina této látky vyvolává projevy
předávkování, označované jako serotoninový syndrom, který se projevuje třesem,
vyšším krevním tlakem, zrychlením srdeční činnosti, zmateností a bezvědomím50
.
2.5.3 Terapeutické využití BCAA
Schopností BCAA je aktivovat glutamát dehydrogenázu, enzym, jehož deficit se
projevuje u choroby amyotrofická laterální skleróza, zvané též jako Lou Gehrigova
choroba. Provedená studie prokázala, že doplňkový příjem 26 g větvených
aminokyselin denně pomáhá osobám trpícím touto nemocí udržet svalovou sílu.
Malá skupina lidí s poruchami nervového systému, souhrnně označovanými jako
spinoceleberální degenerace, zaznamenala v rámci předběžné studie zlepšení
zdravotního stavu při podávání BCAA.
Lidé s onemocněním jater, tzv. hepatální encefalopatií, mají sníženou hladinu
větvených aminokyselin, a naopak zvýšenou hladinu jiných aminokyselin. Při několika
provedených experimentech byl zjištěn celkový pozitivní účinek BCAA na příznaky
tohoto jaterního onemocnění. Příznivé účinky větvených aminokyselin byly rovněž
prokázány u dětí se selháním jater a dospělých s jaterní cirhózou.
Při intravenózním podání aminokyselin s rozvětveným řetězcem pacientům
s chronickým ledvinovým selháním dochází ke zlepšení kvality spánku a dýchání 37
.
Fenylketonurie je dědičně podmíněné metabolické onemocnění spočívající v
poruše přeměny fenylalaninu na tyrosin způsobené nefunkčním enzymem
hydroxylázou. Vysoké hladiny fenylalaninu v krvi poškozují mozek52
. V rámci
kontrolované studie bylo prokázáno, že pravidelné užívání BCAA adolescenty a
mladými dospělými lidmi trpícími touto nemocí, zvyšuje výkonnost při některých
testech duševních schopností. BCAA totiž mohou vytěsňovat fenylalanin, a tím snižovat
jeho toxický účinek37
.
Page 24
23
Dlouhotrvající pooperační stav nebo pokročilé nádorové bujení vedou k
nadměrnému systémovému katabolismu, který je charakterizován rostoucí spotřebou
energie, negativní rovnováhou dusíku, oslabením imunitního systému, zvyšující se
spotřebou glutaminu a narušením metabolismu aminokyselin – dochází k úbytku
svalových proteinů a oxidaci větvených aminokyselin v kosterních svalech (během
normálních podmínek je oxidací BCAA v kosterních svalech poskytováno 6 – 7 %
energie, při stavech s vysokým stupněm katabolismu je to až 20 %.
Ztráta váhy a podvýživa se řadí mezi nejběžnější doprovodné rysy pozorované u
pacientů v pooperačních stavech a onkologických pacientů, kteří jsou vystaveni
přetrvávajícímu katabolickému stresu. Zachováním adekvátního příjmu živin lze zlepšit
šance na přežití pacientů ve stavech vysokého stupně katabolismu. Běžná výživa však
nebývá příliš účinná.
Oxidací aminokyselin s rozvětveným řetězcem je poskytována energie pro
činnost svalů a dalších orgánů, tyto aminokyseliny také hrají důležitou roli při regulaci
metabolismu. Exogenně podávané BCAA, zejména leucin, stimulují syntézu proteinů a
snižují jejich degradaci v izolovaných svalových preparátech a při orálním podání in
vivo. Mnoho příznivých účinků větvených aminokyselin během katabolických procesů
je spojeno se syntézou glukoneogenních aminokyselin, alaninu a zejména glutaminu,
klíčové sloučeniny pro udržení životních funkcí orgánů. Oxidací BCAA ve svalech je
poskytován dusík využívaný pro tvorbu glutaminu – zvyšují jeho syntézu a uvolňování
v kosterních svalech a udržují tak koncentraci glutaminu v krevní plasmě. Tyto
vlastnosti větvených aminokyselin podpořily jejich využití v rámci specielní výživy
zaměřené na zlepšení životních funkcí a klinických výsledků u vážně nemocných
pacientů53
.
Ovšem při studiu účinků potravinových doplňků bohatých na větvené
aminokyseliny zaměřené na středně až těžce nemocné pacienty byly pozorovány určité
rozpory; výsledky některých klinických pokusů ukázaly příznivé účinky BCAA na
zdravotní stav pacientů54,55
, zatímco jiné studie tyto účinky nepotvrdily56,57
. Možné
pozitivní vlivy potravinových doplňků bohatých na BCAA (zejména leucin) u pacientů
po chirurgických zákrocích a pacientů trpících rakovinou nejsou tedy zcela jasné.
Důkazy zatím naznačují, že potravinové doplňky se zvýšeným obsahem BCAA mohou
pozitivně působit pouze u těžce nemocných pacientů53
.
Unikátní vlastností nádorového onemocnění je schopnost karcinomu využít
aminokyseliny pro vlastní metabolické pochody a proliferativní a invazivní činnost.
Page 25
24
Proto ochrana organismu vystaveného katabolickému stresu během onkologického
onemocnění doplňováním větvených aminokyselin může být méně efektivní v důsledku
příjmu BCAA nádorem, čímž je podporováno jeho rozšiřování, růst a schopnost
metastázovat. Nádor, stejně jako zdravé buňky, využívá BCAA pro syntézu bílkovin.
Metoda pozitronové emisní tomografie využívající 11
C-leucin, 11
C-methionin,
11C-tyrosin,
11C-fenylalanin,
11C-tryptofan a
11C-valin, používaná pro zobrazování
mozkových nádorů, poukázala na vysoké výživové nároky tumoru na množství
dostupných aminokyselin. Myšlenka, že doplňková výživa obohacená o aminokyseliny
může podněcovat nádorové bujení zvyšující se syntézou bílkovin je však méně
znepokojující pro BCAA doplňky, neboť právě větvené aminokyseliny jsou méně
zapojené v procesu buněčného množení, na rozdíl např. od methioninu nebo glutaminu.
Na druhou stranu BCAA jsou nezbytnými prekurzory a substráty pro zachování
fyziologických funkcí organismu i pro nádorový růst. Ideální by proto bylo nalézt
takovou dávku doplňkově přijímaných větvených aminokyselin, která by maximálně
zohledňovala nutriční nároky samotného organismu nad nároky zhoubného nádoru35
.
Z výše uvedené podrobné rešerše o používání a účincích větvených
aminokyselin sportovci, jedinci s vysokou tělesnou zátěží a nemocnými není možno
učinit jednoznačný závěr o pozitivním či negativním působení valinu, leucinu a
isoleucinu v organismu. O případných rozdílných účincích jednotlivých enantiomerů
těchto aminokyselin je známo ještě méně. Objasnění této problematiky je proto velkou
výzvou, neboť případné poznatky o stereoselektivním působení těchto látek by našly
široké využití - nejen z hlediska výživy sportovců, ale zejména v medicíně.
2.6 Principy enantioseparace v HPLC
V izotropickém (achirálním) prostředí mají chirální látky shodné vlastnosti a za
těchto podmínek tedy nemohou být rozděleny. Techniky separace enantiomerů jsou
založeny na principech chirálního rozpoznávání pozorovaného v přírodě - enantiomery
je možné rozdělit pouze v chirálním prostředí1.
Při separaci chirálních sloučenin jsou využívány dva základní postupy. Nepřímé
metody dělení jsou založeny na derivatizační reakci racemického analytu s opticky
čistým činidlem. Při této reakci vzniká pár diastereomerních sloučenin, které mají
Page 26
25
rozdílné fyzikálně-chemické vlastnosti. K následné separaci je pak využíván achirální
chromatografický systém7.
Většího praktického využití nalezly přímé metody dělení, při kterých je do
separačního systému zaveden vhodný chirální selektor. Chirální selektor se nejběžněji
používá ve formě chirální stacionární fáze (CSP), tj. vázaný na pevný nosič, méně často
je přidáván do mobilní fáze1. Pro úspěšné rozdělení enantiomerů jsou zapotřebí
minimálně tři současně probíhající interakce mezi enantiomerem a chirálním selektorem
(tzv. model tříbodové interakce, odvozený v roce 1952 Dalglishem)58
, přičemž alespoň
jedna z interakcí má mít stereoselektivní charakter. K typům interakcí probíhajících
mezi chirálním analytem a chirálním selektorem se řadí: elektrostatické interakce,
vodíková vazba, hydrofóbní interakce, stérické stínění, π-π interakce a dipólové
interakce. Na míře rozdílných afinit enantiomerů k chirálnímu selektoru pak závisí
vlastní enantioseparace v chromatografickém systému59
.
2.6.1 Chirální stacionární fáze
V kapalinové chromatografii jsou pro chirální separace nejčastěji využívány
stacionární fáze tvořené silikagelovým nosičem s navázanými chirálními selektory na
bázi polysacharidů, cyklodextrinů, proteinů, makrocyklických antibiotik, ligandově
výměnné CSP a fáze tvořené syntetickými polymery59
. Právě makrocyklickým
antibiotikům bude, vzhledem k zaměření této práce, věnována větší pozornost.
2.6.2 Makrocyklická antibiotika
Makrocyklická antibiotika (MA) patří do skupiny nejmodernějších chirálních
selektorů. Nejčastěji používanými MA jsou glykopeptidy, mezi něž se řadí teikoplanin,
vankomycin, ristocetin A a avoparcin58
. Tato antibiotika patří do skupiny inhibitorů
syntézy buněčné stěny a jsou účinná proti aerobním a anaerobním grampozitivním
bakteriím60
.
Struktura makrocyklických antibiotik je značně složitá, obsahuje velké množství
chirálních center a vázaných funkčních skupin, které umožňují řadu interakcí
Page 27
26
s chirálními analyty. CSP na bázi glykopeptidů lze použít v normálním pracovním
modu (stacionární fáze je polárnější než mobilní, uplatňují se zejména π-π, dipólové
interakce a H-vazby), v reverzním pracovním modu (mobilní fáze je polárnější,
převažují hydrofóbní interakce, vodíkové vazby a také inkluze) i v polárně organickém
modu (mobilní fáze je složena z acetonitrilu, methanolu, případně pouze methanolu a
malého množství triethylaminu a kyseliny octové na ovlivnění selektivity), ve kterém se
uplatňuje většina výše zmíněných interakcí - omezené jsou hydrofóbní interakce a
inkluze58,60
.
2.6.2.1 Teikoplanin
Teikoplanin je přirozeným produktem plísně Actinoplanes teicomycetus.
Struktura této biomolekuly (viz obr. 2.4) je velmi zajímavá; tzv. aglykonový koš je
tvořen čtyřmi makrocykly. Molekulu tvoří hexapeptidový řetězec, na který je navázáno
sedm benzenových jader, pět z nich je vzájemně propojeno třemi etherovými
skupinami. Na dvou benzenech je navázán atom chloru a na šesti jsou vázány
hydroxylové skupiny. Aglykon také obsahuje samostatnou karboxylovou skupinu a
primární amin, čímž molekula získává amfoterní charakter. Na aglykonový koš jsou
dále navázány tři cukerné zbytky, tvořené dvěma D-glukosaminy a D-mannosou. Na
jeden D-glukosamin je dále vázán acylový řetězec; přítomnost tohoto hydrofóbního
řetězce významně přispívá k nízké rozpustnosti molekuly ve vodě. V přírodě se
teikoplaninové glykopeptidy liší právě tímto uhlovodíkovým řetězcem61
.
CSP na bázi teikoplaninu jsou tvořeny směsí pěti izomerů, které se liší větvením
acylového řetězce. Molekuly chirálního selektoru jsou vázány kovalentní vazbou na
silikagelový nosič60
. Komerčně dostupné chirální kolony Chirobiotic T a T2 obsahují
silikagelový nosič o velikosti zrn 5 μm, ovšem velikost pórů je u kolony Chirobiotic T
120 Å a u Chirobiotic T2 200 Å. Navíc délka řetězce, který zprostředkovává vazbu
teikoplaninu na silikagel je u Chirobiotic T2 téměř dvojnásobná oproti délce řetězce
v Chirobiotic T62
.
Page 28
27
Obr. 2.4 Strukturní vzorec molekuly teikoplaninu, chirální centra jsou označena
symboly R a S.
2.6.2.2 Teikoplanin aglykon
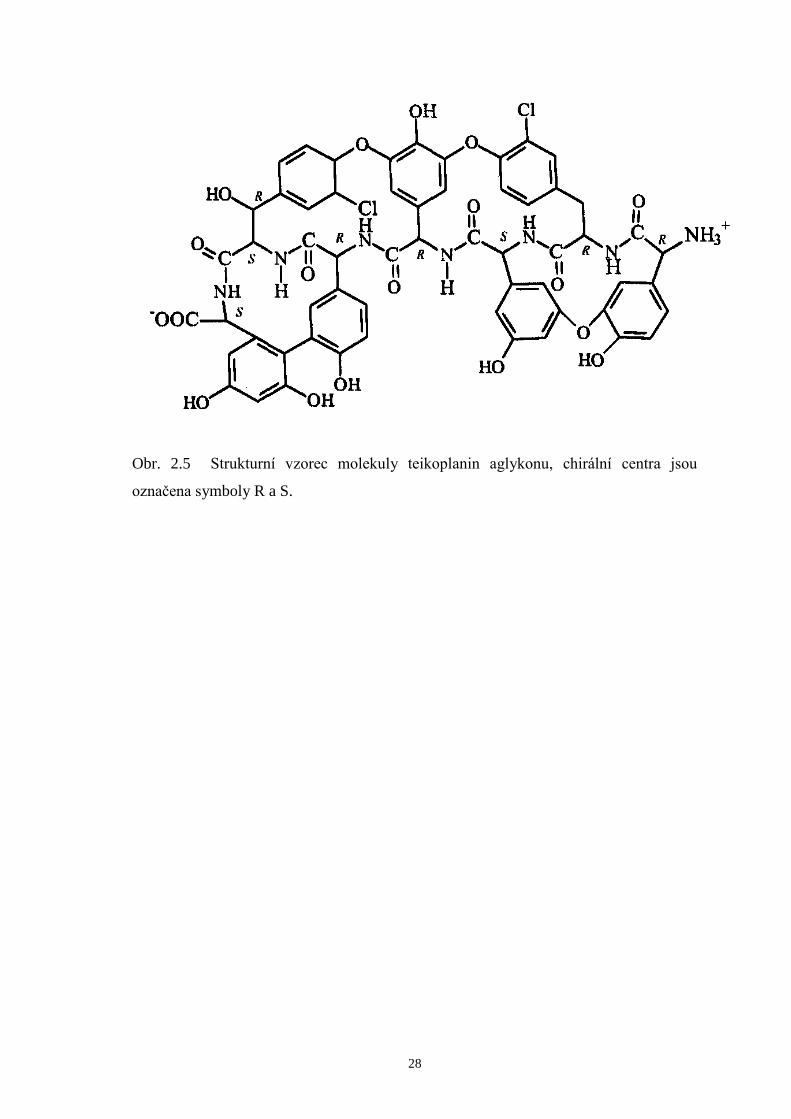
Molekulu teikoplanin aglykonu (viz obr. 2.5) lze připravit úpravou přírodního
teikoplaninu nebo také organickou syntézou. Od teikoplaninu se tato molekula liší, jak
je již z názvu patrné, nepřítomností cukerných zbytků včetně hydrofóbního acylového
řetězce. Tím je dosaženo snazšího přístupu analytů k aglykonové dutině, která obsahuje
další chirální centra a množství ionizovatelných skupin. Teikoplanin aglykon je rovněž
špatně rozpustný ve vodě60,63
.
Page 29
28
Obr. 2.5 Strukturní vzorec molekuly teikoplanin aglykonu, chirální centra jsou
označena symboly R a S.
Page 30
29
3 CÍL PRÁCE
Teikoplaninové CSP jsou významnou skupinou chirálních stacionárních fází,
které umožňují dělení široké škály strukturně odlišných analytů. Velmi dobré uplatnění
našly v oblasti separace aminokyselin. Novější komerční stacionární fáze, u které má
být silikagelový nosič více pokrytý teikoplaninem (Chirobiotic T2), je zatím málo
známý a málo vyzkoušený produkt.
Aminokyseliny se v nedávných letech staly velmi populárními výživovými
doplňky užívanými zejména sportovci. Významná skupina těchto doplňků pro sportovní
výživu obsahuje L-valin, L-leucin a L-isoleucin. Tyto výrobky jsou volně dostupné a
široce využívané zejména atlety a kulturisty. Během výrobního procesu ale mohou
v těchto produktech vznikat také příslušné D-formy daných aminokyselin. D-
aminokyseliny mají rozdílné biologické a fyziologické vlastnosti, proto je důležité
kontrolovat enantiomerní čistotu těchto výrobků, aby nedocházelo k výživovým ztrátám
z důvodu vzniku nemetabolizovatelných a biologicky nevyužitelných forem těchto
aminokyselin64
.
Cíle této práce lze shrnout do několika bodů:
Optimalizace metody pro separaci enantiomerů aminokyselin valinu, leucinu a
isoleucinu.
Charakterizace a vzájemné porovnání chirálních stacionárních fází na bázi
teikoplaninu.
Určení limitu detekce a limitu stanovení enantiomerů valinu, leucinu a
isoleucinu na chirální stacionární teikoplaninové fázi.
Sledování procesů vyvolávajících racemizaci aminokyselin obsažených
v měřeném vzorku BCAA 500.
Page 31
30
4 EXPERIMENTÁLNÍ ČÁST
4.1 Seznam použitých chemikálií
methanol (MeOH): pro chromatografii (Sigma-Aldrich, Praha, Česká republika)
acetonitryl (ACN): pro chromatografii (Sigma-Aldrich, Praha, Česká republika)
kyselina chlorovodíková 35 %: (Lachema a.s., Neratovice, Česká republika)
deionizovaná voda (H2O) filtrovaná Milli-Q filtrovacím systémem (Millipore, Milford,
MA, USA)
anthracen: p. a. (Sigma, St. Louis, MO, USA)
benzen: p. a. (Sigma, St. Louis, MO, USA)
N,N-diethyltoluamid: p. a. (Sigma, St. Louis, MO, USA)
standardy aminokyselin: L-valin (L-Val): čistota ≥ 99,5 % (Fluka, Praha, Česká
republika), DL-valin (DL-Val): čistota ≥ 99,0 % (Fluka, Praha, Česká republika), L-
leucin (L-Leu): pro biochemii (Merck, Německo), DL-leucin (DL-Leu): čistota ≥ 99,0 %
(Fluka, Praha, Česká republika), L-isoleucin (L-Ile): čistota ≥ 99,5 % (Fluka, Praha,
Česká republika), D-isoleucin (D-Ile): čistota ≥ 98,0 % (Sigma-Aldrich, Praha, Česká
republika)
měřený vzorek: BCAA 500 tobolky: obsah: L-leucin 218,75 mg, L-valin 156,25 mg, L-
isoleucin 125,00 mg (Petr Novák - Bolid Beroun, ve spolupráci s NUTRI-EXACT CZ
s.r.o., Česká republika)
Page 32
31
4.2 Charakterizace chirálních aminokyselin
V tabulce 4.1 jsou uvedeny základní charakterizace studovaných analytů.
Tabulka 4.1 Studované aminokyseliny a jejich vlastnosti: relativní molekulová
hmotnost (Mr), disociační konstanty (pKA25
, pKB25
), izoelektrický bod (pI25
) a
rozdělovací poměr (log P) tří měřených chirálních analytů, aminokyselin valinu, leucinu
a isoleucinu
Název VALIN LEUCIN ISOLEUCIN
Funkční vzorec CH3CH(CH3)CH(NH2)COOH CH3CH(CH3)CH2CH(NH2)COOH CH3CH2CH(CH3)CH(NH2)COOH
Mr 117,148 131,175 131,175
pKA25
9,719 9,744 9,758
pKB25
11,711 2,328 11,682
pI 25
6,002 6,036 6,038
log P *
-2,26
-1,52
-1,70
* http://www.syrres.com/esc/est_kowdemo.htm
4.3 Použité přístroje
Tato práce byla měřena na kapalinovém chromatografu Breeze System, který se
skládá z HPLC gradientové pumpy 1525, autosampleru 717Plus, termostatu Jetstream 2
Plus , UV/VIS dvoukanálového detektoru 2487 a osobního počítače s nainstalovaným
softwarem Breeze, verze 3.30 SPA (vše od firmy Waters, Milford, USA).
Pro separaci analytů byly využívány kolony s navázaným chirálním selektorem
teikoplaninem resp. jeho analogem teikoplanin aglykonem:
1) Chirobiotic T: kovová kolona, 25 cm x 4,6 mm, naplněná teikoplaninem vázaným
na silikagelový nosič, velikost zrn 5 µm, velikost póru 8 nm (Astec - Advanced
Separation Technologies, Inc., Whipanny, NJ, USA).
2) Chirobiotic T2: kovová kolona, 25 cm x 4,6 mm, naplněná teikoplaninem
navázaným na silikagelový nosič, velikost zrn 5 µm, velikost póru 8 nm (Supelco,
Bellefonte, USA).
3) Chirobiotic TAG: kovová kolona, 25 cm x 4,6 mm, naplněná teikoplanin aglykonem
vázaným na silikagelový nosič, velikost zrn 5 µm, velikost póru 8 nm (Astec -
Advanced Separation Technologies, Inc., Whipanny, NJ, USA).
Page 33
32
Dále byly při práci používány ultrazvuková lázeň Ultrasonic LC 30 (Elma,
Německo), pH metr PHM-210 standardní (Radiometer, Kodaň, Dánsko), termostat,
který je sestaven z ponorného ohřívacího cirkulátoru MB a vodní lázně F12 (Julabo,
Seelbach, Německo), analytické váhy Mettler AE 240 (Greifensee, Švýcarsko),
míchačka MS2-S8 (IKA Works, Inc., Wilmington, NC, USA) a centifuga MC2
(Qualitron, Inc., Ohio, USA).
4.4 Příprava vzorků
Zásobní roztoky standardů aminokyselin byly připraveny rozpuštěním 1 mg
látky v 1 ml solventu tvořeného 60-i objemovými procenty (%obj.) methanolu a 40-i
objemovými procenty deinizované vody. Poté se tyto roztoky nechaly 10 minut
odstředit na centrifuze a následně byly přefiltrovány. Při měření kalibrační závislosti a
při sledování vlivu teploty na racemizaci byly zásobní roztoky standardů L-Val, L-Leu a
L-Ile připraveny rozpuštěním 1 mg látky v 1 ml deinizované vody a poté centrifugovány
10 minut.
Zásobní roztoky měřeného vzorku byly připraveny rozpuštěním 1mg BCAA 500
v 1 ml deinizované vody a poté se tyto roztoky nechaly 10 minut odstředit na centrifuze.
Příprava vzorků pro Waltersův test byla provedena podle návodu převzatého
z literatury65
.
4.5 Příprava mobilní fáze
Při chirálních separacích aminokyselin byla používána mobilní fáze složená
z methanolu a deionizované vody podle tabulky 4.2.
Tab. 4.2 Složení studovaných mobilních fází - objemová procenta methanolu a vody.
Složka S l o ž e n í m o b i l n í f á z e
MeOH (%obj.) 10 20 30 40 60 70 75 80 90
H2O (%obj.) 90 80 70 60 40 30 25 20 10
Page 34
33
Při provádění Waltersova testu byla pro stanovení indexu hydrofobicity
používána mobilní fáze složená z 65 %obj. ACN a 35 %obj. H2O, pro stanovení
silanolového indexu byla používána mobilní fáze tvořená čistým ACN.
4.6 Podmínky měření
Používané mobilní fáze jsou uvedeny v kap. 4.5. Destilovaná voda pro přípravu
mobilní fáze byla před měřením vždy odvzdušňována v ultrazvukové lázni po dobu 10
minut. Průtoková rychlost mobilní fáze byla pro všechna měření 1 ml/min. Teplota
kolony v termostatu byla vždy nastavena na 25°C. Detekce aminokyselin byla
prováděna při vlnové délce 200 nm, pro Waltersův test při vlnové délce 254 nm.
Dávkované množství látky na kolonu bylo vždy 20 μl a jednotlivá měření byla
prováděná minimálně dvakrát, při měření kalibrační závislosti bylo každé měření
provedeno třikrát.
4.7 Zpracování naměřených dat
Naměřená data byla vyhodnocena programem Breeze, verze 3.30 SPA. Tímto
programem byly u jednotlivých analytů určeny retenční časy, retenční faktory, rozlišení,
a faktory symetrie, dále využité pro zpracování výsledků.
Hodnoty retenčních faktorů byly vypočítány podle vztahu
M
MR
t
ttk (1)
kde tR je retenční čas, tedy celkový čas, který analyt setrvá v koloně a tM je mrtvý objem
kolony, tedy retenční čas analytu, který se na koloně nezadržuje.
Page 35
34
Hodnoty separačních faktorů byly vypočítány podle vztahu
i
j
k
k
ji, (2)
)
kde kj resp. ki jsou retenční faktory později resp. dříve eluující složky.
Hodnoty rozlišení byly vypočítány podle vztahu
21
12 ).(2
2,1 ww
tt RRR (3)
kde tR2 resp. tR1 jsou retenční časy později resp. dříve eluující složky, w1 a w2 jsou šířky
píků při základně.
Hodnota rozlišení je závislá na počtu teoretických pater kolony, separačním a
retenčním faktoru podle vztahu
j
j
ij
ij
k
knjiR
1
1
4,,
,
(4)
kde n je počet teoretických pater dané kolony.
Hodnoty faktoru symetrie (S) byly vypočítány ze vztahu (5) podle obrázku 4.1.
f
WS
2
05,0
(5)
Obr. 4.1 Názorné schéma ilustrující výpočet faktoru symetrie66
.
Page 36
35
4.8 Postup Waltersova testu
Pro provedení Waltersova testu byl využit postup dle literatury65
.
4.9 Racemizační postupy
Při zkoumání vlivu nevhodných skladovacích podmínek na racemizaci byl
vzorek ponechán v letním období, tzn. červenec - září 2008, na světle a při laboratorní
teplotě v těchto formách: chráněný v tobolce, volně vysypaný na dno kádinky,
rozpuštěný v deionizované vodě a rozpuštěný v ovocném čaji. Okraje kádinek byly
překryty parafilmem. Vzorek chráněný tobolkou a vzorek volně vysypaný byly vždy
před měřením rozpuštěny, a to 1 mg látky v 1 ml deionizované vody, a poté odstředěny
po dobu 10 minut. Vzorky rozpuštěné v deionizované vodě a ovocném čaji byly
přefiltrovány a rovnou dávkovány na kolonu.
Na vznik racemizačních procesů ve vzorku byl také zkoumán vliv pH. K 1 mg L-
Val, L-Leu, L-Ile a vzorku byl přidán vždy 1 ml vodného roztoku HCl o molární
koncentraci 0,01 mol·dm-3
(pH 2,04; tj. přibližné pH kyseliny chlorovodíkové v lidském
žaludku) a o molární koncentraci 6,00 mol.dm-3
. Před nadávkováním na kolonu byly
vzorky rozpuštěné v 0,01 M HCl naředěny deionizovanou vodou v poměru 1:1. Vzorky
rozpuštěné v 6,00 M HCl byly deionizovanou vodou naředěny v poměru 1:5 a
dávkovány na kolonu jeden den a sedm dní po přípravě.
Dále byl zkoumán vliv teploty. Vzorky v tobolce, volně vysypaný a rozpuštěný
v dionizované vodě byly zahřívány v termostatu po dobu 15 minut při teplotách 25°C,
37°C, 50°C a 60°C. Dále byly jak zásobní roztoky standardů L-Val, L-Leu, L-Ile a
vzorku ve vodě, tak jejich roztoky v 6,00 molární HCl (před nadávkováním na kolonu
naředěné deionizovanou vodou v poměru 1:5) zahřívány v termostatu po dobu 30 minut
při teplotách 60°C a 90°C. Dalším z postupů bylo přelití obsahu jedné tobolky (500 mg)
500 ml destilované vody a ovocného čaje přivedených k varu. Tyto roztoky vzorku byly
po úplném vychladnutí přefiltrovány a dávkovány na kolonu.
Page 37
36
5. VÝSLEDKY A DISKUZE
5.1 Optimalizace metody pro separaci enantiomerů valinu, leucinu a
isoleucinu
V BCAA potravinových doplňcích jsou v určitém poměru obsaženy
aminokyseliny L-valin, L-leucin a L-isoleucin, které však v důsledku nevhodných
skladovacích podmínek, mikrobiální kontaminace nebo již během výrobního procesu
mohou racemizovat. Vzniklé D-analogy jsou pro člověka z hlediska výživy nevyužitelné
a jejich přítomnost ve výrobku tak snižuje jeho nutriční hodnotu. Jedním z cílů této
práce proto bylo nalézt vhodnou HPLC metodu, která by umožnila rozdělení všech šesti
enantiomerů aminokyselin s dostatečnou hodnotou rozlišení a s jejíž pomocí by pak
bylo možné kontrolovat enantiomerní čistotu těchto výrobků. Za tímto účelem byla
měření prováděna postupně na chirálních kolonách Chirobiotic TAG, Chirobiotic T a
Chirobiotic T2.
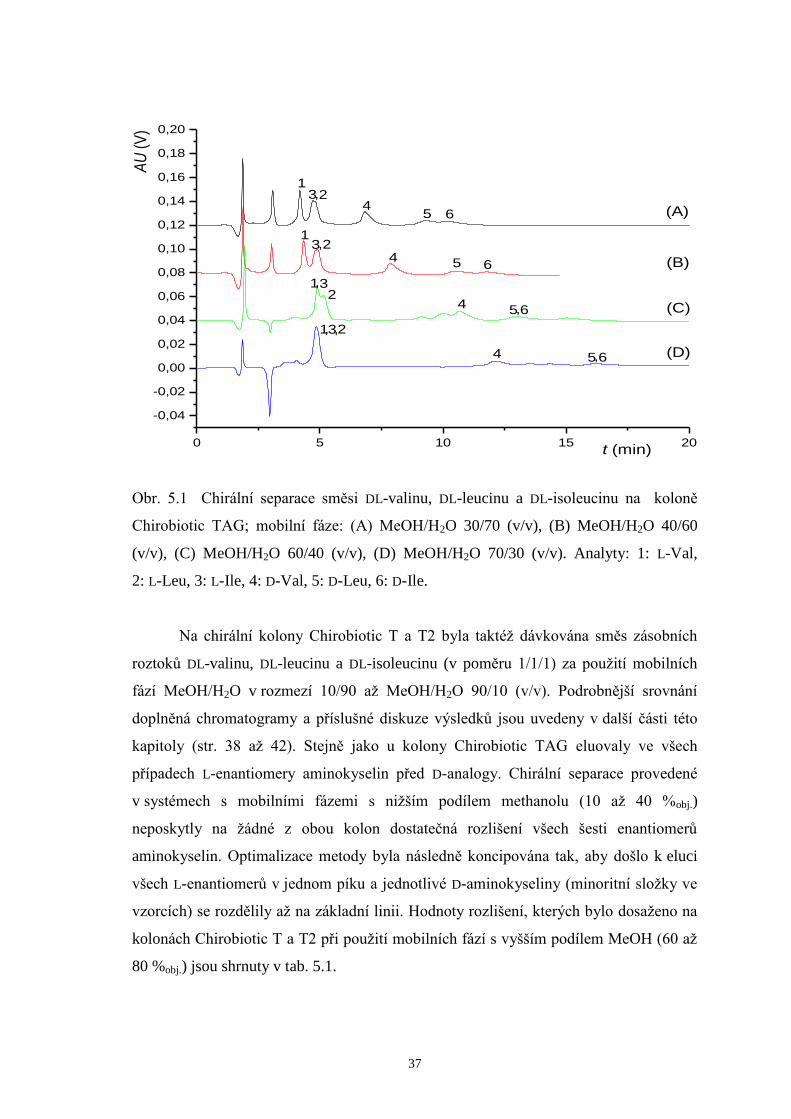
Na chirální kolonu Chirobiotic TAG byla dávkována směs zásobních roztoků
DL-valinu, DL-leucinu a DL-isoleucinu (v poměru 1/1/1) a měření byla prováděna za
použití mobilních fází MeOH/H2O 30/70 (v/v), MeOH/H2O 40/60 (v/v), MeOH/H2O
60/40 (v/v) a MeOH/H2O 70/30 (v/v). Chromatografy zobrazující enantioseparaci na
koloně Chirobiotic TAG při použití daných mobilních fází jsou znázorněny na obrázku
5.1. Ve všech případech eluovaly L-enantiomery aminokyselin před D-analogy. Při
žádné provedené chirální separaci však nebylo dosaženo požadovaných výsledků
z hlediska dostatečného rozlišení všech enantiomerů a symetrie píků. Z těchto důvodů
proto nebyla na této koloně prováděna další měření.
Page 38
37
0 5 10 15 20
-0,04
-0,02
0,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
0,16
0,18
0,20
,6
,, 23
,65
,
6
,
6
,3
t (min)
54
1
4
32
1
543 2
1
(D)
(C)
(B)
(A)54
21
AU
(V
)
Obr. 5.1 Chirální separace směsi DL-valinu, DL-leucinu a DL-isoleucinu na koloně
Chirobiotic TAG; mobilní fáze: (A) MeOH/H2O 30/70 (v/v), (B) MeOH/H2O 40/60
(v/v), (C) MeOH/H2O 60/40 (v/v), (D) MeOH/H2O 70/30 (v/v). Analyty: 1: L-Val,
2: L-Leu, 3: L-Ile, 4: D-Val, 5: D-Leu, 6: D-Ile.
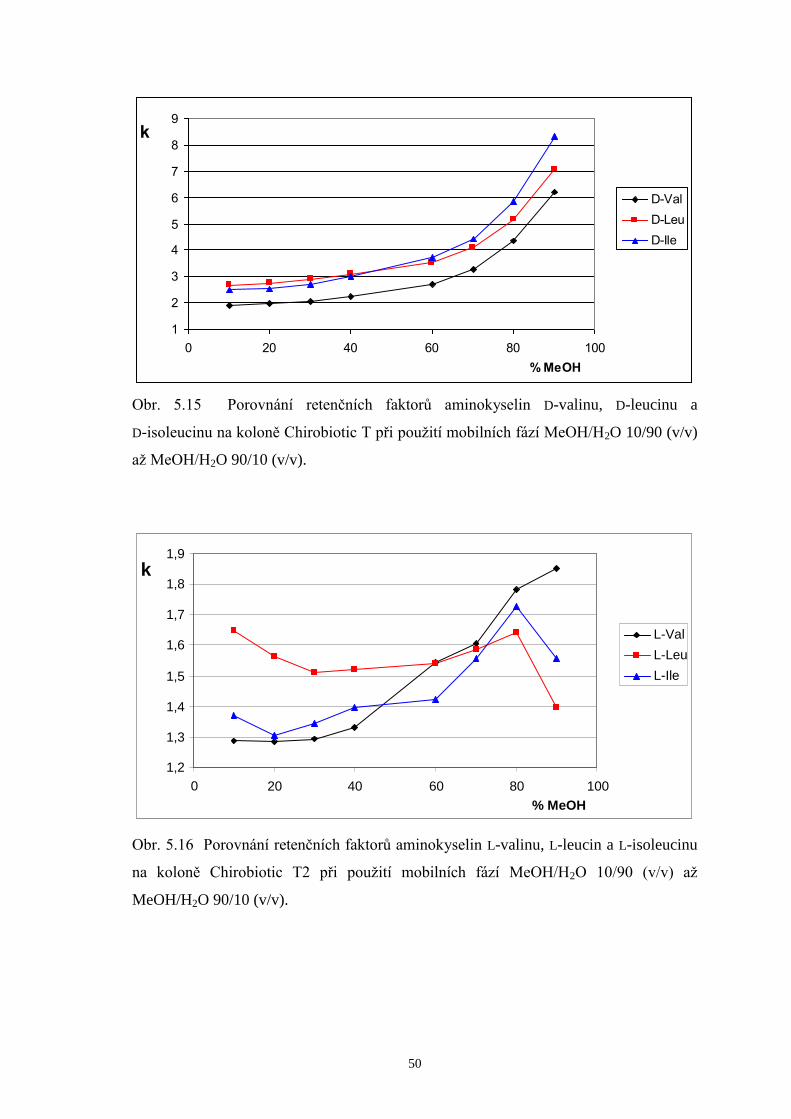
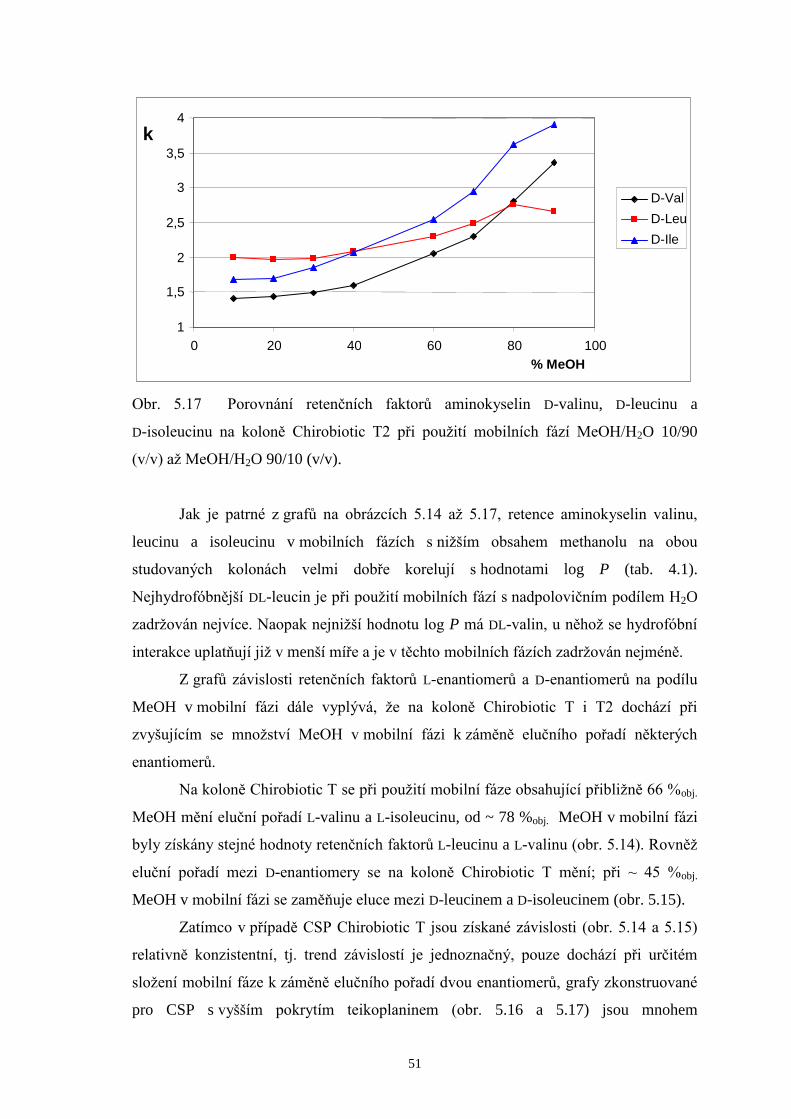
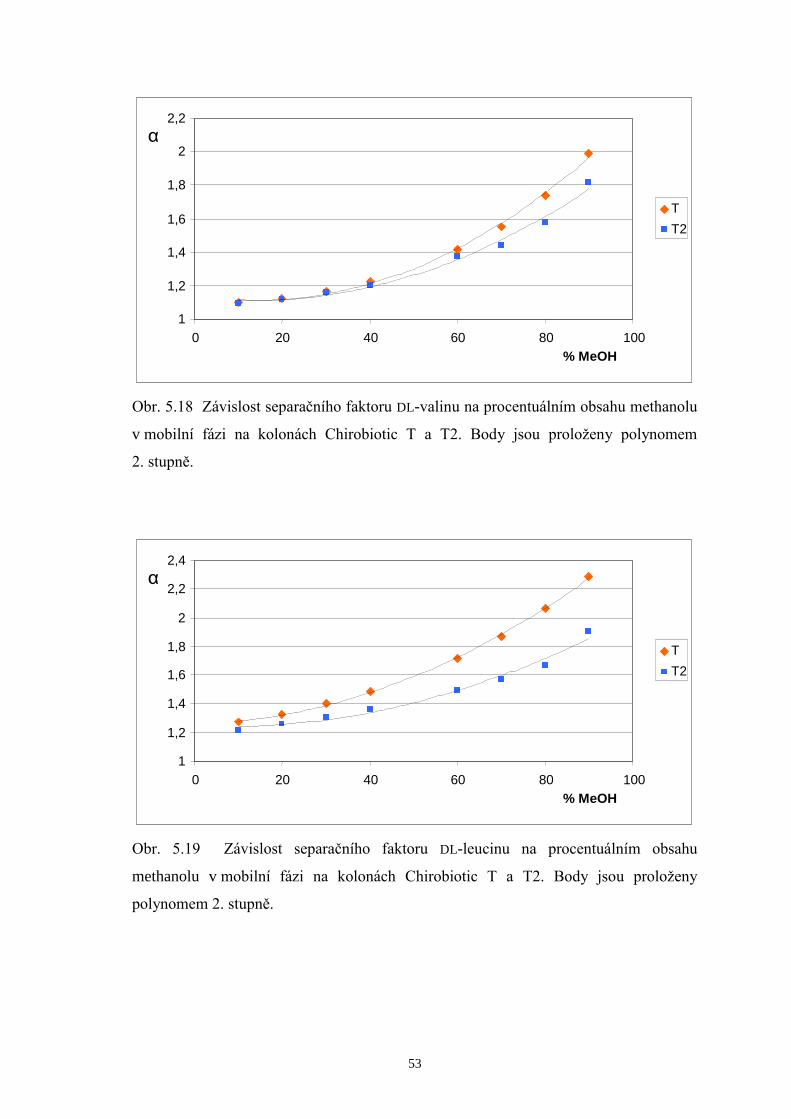
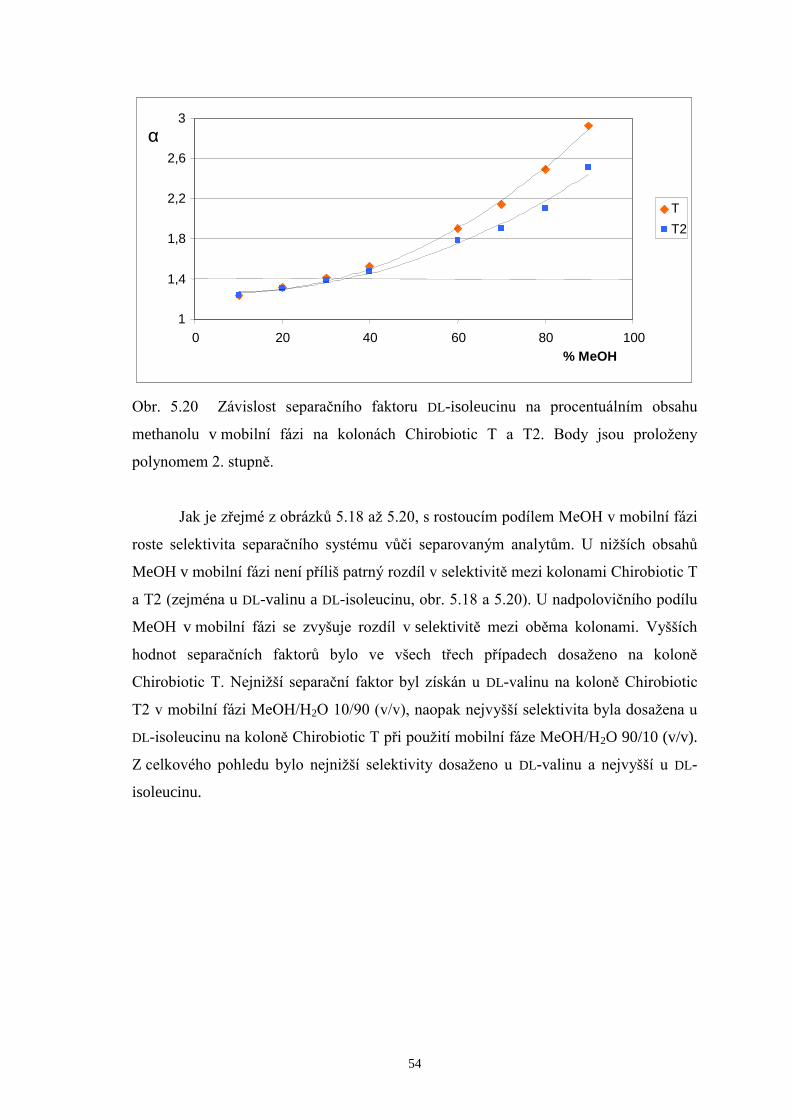
Na chirální kolony Chirobiotic T a T2 byla taktéž dávkována směs zásobních
roztoků DL-valinu, DL-leucinu a DL-isoleucinu (v poměru 1/1/1) za použití mobilních
fází MeOH/H2O v rozmezí 10/90 až MeOH/H2O 90/10 (v/v). Podrobnější srovnání
doplněná chromatogramy a příslušné diskuze výsledků jsou uvedeny v další části této
kapitoly (str. 38 až 42). Stejně jako u kolony Chirobiotic TAG eluovaly ve všech
případech L-enantiomery aminokyselin před D-analogy. Chirální separace provedené
v systémech s mobilními fázemi s nižším podílem methanolu (10 až 40 %obj.)
neposkytly na žádné z obou kolon dostatečná rozlišení všech šesti enantiomerů
aminokyselin. Optimalizace metody byla následně koncipována tak, aby došlo k eluci
všech L-enantiomerů v jednom píku a jednotlivé D-aminokyseliny (minoritní složky ve
vzorcích) se rozdělily až na základní linii. Hodnoty rozlišení, kterých bylo dosaženo na
kolonách Chirobiotic T a T2 při použití mobilních fází s vyšším podílem MeOH (60 až
80 %obj.) jsou shrnuty v tab. 5.1.
Page 39
38
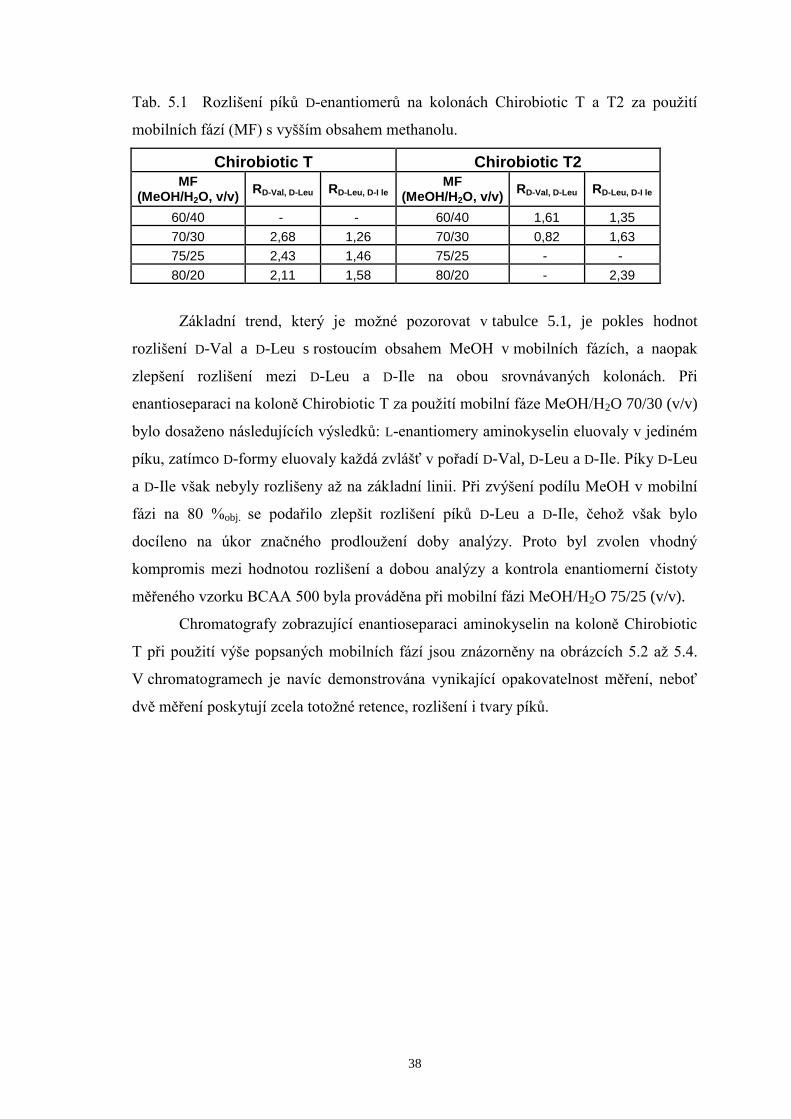
Tab. 5.1 Rozlišení píků D-enantiomerů na kolonách Chirobiotic T a T2 za použití
mobilních fází (MF) s vyšším obsahem methanolu.
Chirobiotic T Chirobiotic T2 MF
(MeOH/H2O, v/v) RD-Val, D-Leu RD-Leu, D-I le
MF (MeOH/H2O, v/v)
RD-Val, D-Leu RD-Leu, D-I le
60/40 - - 60/40 1,61 1,35
70/30 2,68 1,26 70/30 0,82 1,63
75/25 2,43 1,46 75/25 - -
80/20 2,11 1,58 80/20 - 2,39
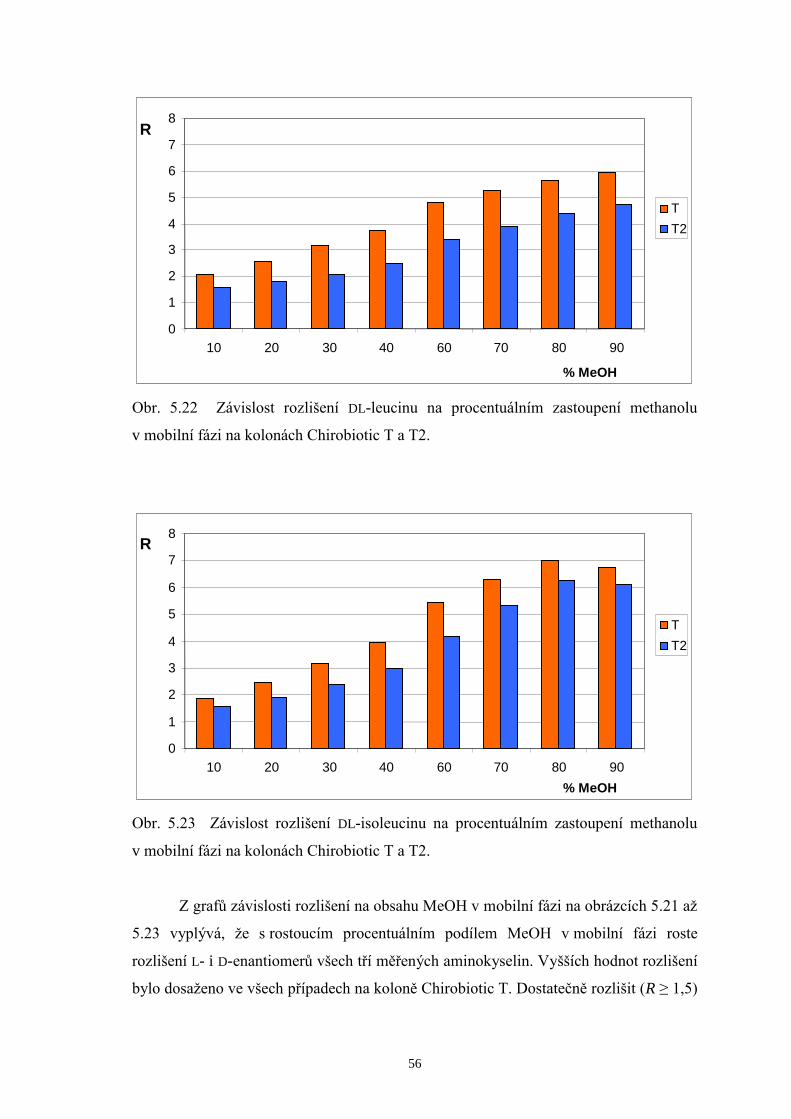
Základní trend, který je možné pozorovat v tabulce 5.1, je pokles hodnot
rozlišení D-Val a D-Leu s rostoucím obsahem MeOH v mobilních fázích, a naopak
zlepšení rozlišení mezi D-Leu a D-Ile na obou srovnávaných kolonách. Při
enantioseparaci na koloně Chirobiotic T za použití mobilní fáze MeOH/H2O 70/30 (v/v)
bylo dosaženo následujících výsledků: L-enantiomery aminokyselin eluovaly v jediném
píku, zatímco D-formy eluovaly každá zvlášť v pořadí D-Val, D-Leu a D-Ile. Píky D-Leu
a D-Ile však nebyly rozlišeny až na základní linii. Při zvýšení podílu MeOH v mobilní
fázi na 80 %obj. se podařilo zlepšit rozlišení píků D-Leu a D-Ile, čehož však bylo
docíleno na úkor značného prodloužení doby analýzy. Proto byl zvolen vhodný
kompromis mezi hodnotou rozlišení a dobou analýzy a kontrola enantiomerní čistoty
měřeného vzorku BCAA 500 byla prováděna při mobilní fázi MeOH/H2O 75/25 (v/v).
Chromatografy zobrazující enantioseparaci aminokyselin na koloně Chirobiotic
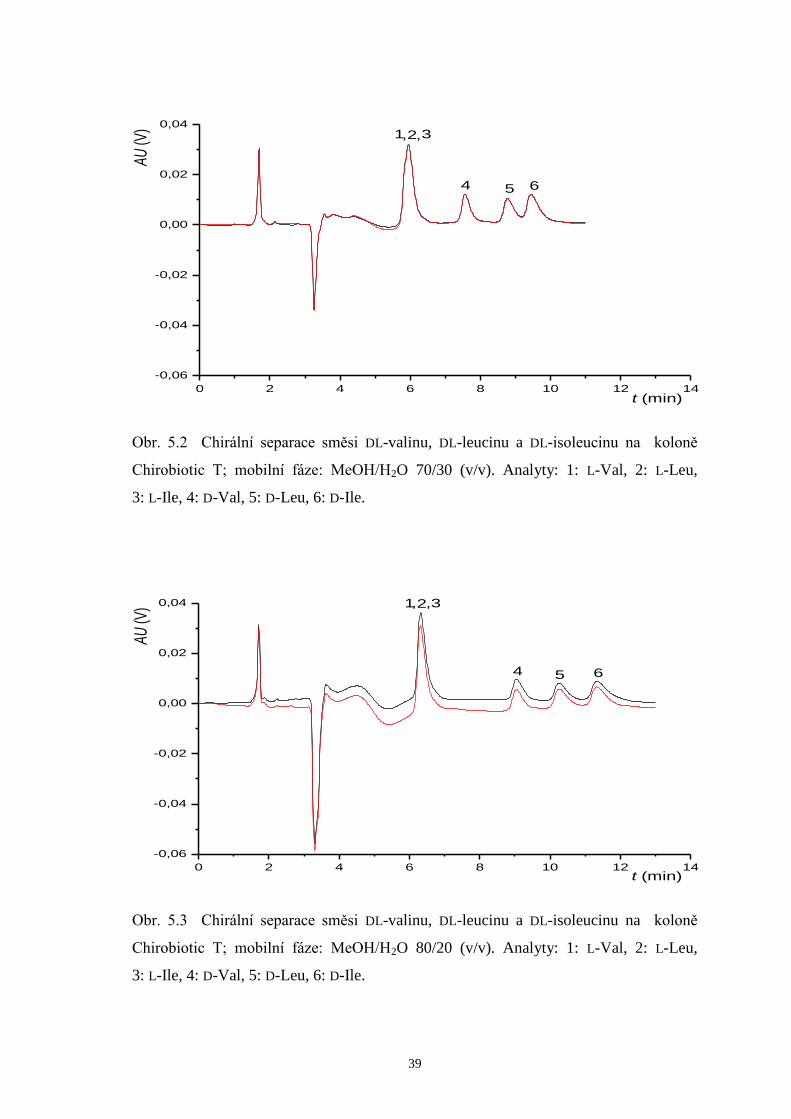
T při použití výše popsaných mobilních fází jsou znázorněny na obrázcích 5.2 až 5.4.
V chromatogramech je navíc demonstrována vynikající opakovatelnost měření, neboť
dvě měření poskytují zcela totožné retence, rozlišení i tvary píků.
Page 40
39
0 2 4 6 8 10 12 14
-0,06
-0,04
-0,02
0,00
0,02
0,04
65
, ,
4
321AU
(V)
t (min)
Obr. 5.2 Chirální separace směsi DL-valinu, DL-leucinu a DL-isoleucinu na koloně
Chirobiotic T; mobilní fáze: MeOH/H2O 70/30 (v/v). Analyty: 1: L-Val, 2: L-Leu,
3: L-Ile, 4: D-Val, 5: D-Leu, 6: D-Ile.
0 2 4 6 8 10 12 14
-0,06
-0,04
-0,02
0,00
0,02
0,04
65
,,
4
321
AU (V
)
t (min)
Obr. 5.3 Chirální separace směsi DL-valinu, DL-leucinu a DL-isoleucinu na koloně
Chirobiotic T; mobilní fáze: MeOH/H2O 80/20 (v/v). Analyty: 1: L-Val, 2: L-Leu,
3: L-Ile, 4: D-Val, 5: D-Leu, 6: D-Ile.
Page 41
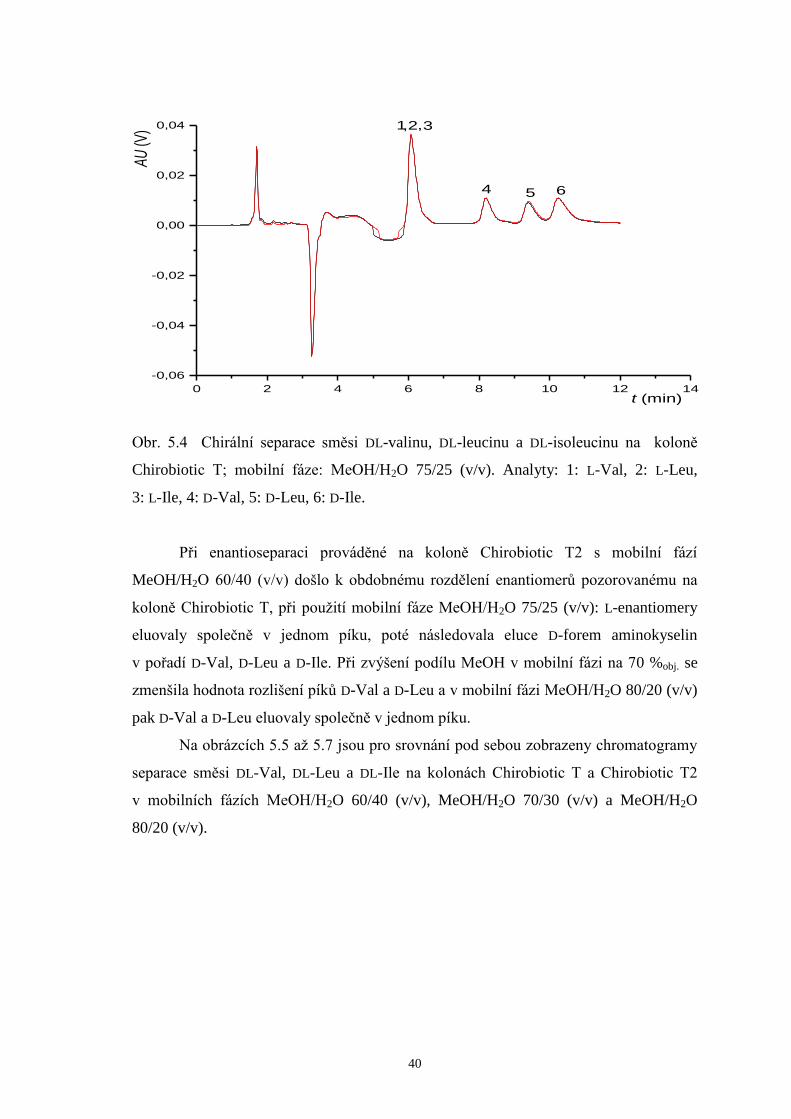
40
0 2 4 6 8 10 12 14
-0,06
-0,04
-0,02
0,00
0,02
0,04 ,,
654
321
AU (V
)
t (min)
Obr. 5.4 Chirální separace směsi DL-valinu, DL-leucinu a DL-isoleucinu na koloně
Chirobiotic T; mobilní fáze: MeOH/H2O 75/25 (v/v). Analyty: 1: L-Val, 2: L-Leu,
3: L-Ile, 4: D-Val, 5: D-Leu, 6: D-Ile.
Při enantioseparaci prováděné na koloně Chirobiotic T2 s mobilní fází
MeOH/H2O 60/40 (v/v) došlo k obdobnému rozdělení enantiomerů pozorovanému na
koloně Chirobiotic T, při použití mobilní fáze MeOH/H2O 75/25 (v/v): L-enantiomery
eluovaly společně v jednom píku, poté následovala eluce D-forem aminokyselin
v pořadí D-Val, D-Leu a D-Ile. Při zvýšení podílu MeOH v mobilní fázi na 70 %obj. se
zmenšila hodnota rozlišení píků D-Val a D-Leu a v mobilní fázi MeOH/H2O 80/20 (v/v)
pak D-Val a D-Leu eluovaly společně v jednom píku.
Na obrázcích 5.5 až 5.7 jsou pro srovnání pod sebou zobrazeny chromatogramy
separace směsi DL-Val, DL-Leu a DL-Ile na kolonách Chirobiotic T a Chirobiotic T2
v mobilních fázích MeOH/H2O 60/40 (v/v), MeOH/H2O 70/30 (v/v) a MeOH/H2O
80/20 (v/v).
Page 42
41
0 2 4 6 8 10 12 14
-0,08
-0,06
-0,04
-0,02
0,00
0,02
0,04 0 2 4 6 8 10 12 14
-0,06
-0,04
-0,02
0,00
0,02
0,04
0,06
65
, ,
(B)
4
321
AU
(V)
t (min)
6
,
(A)
54
321A
U (V
)
t (min)
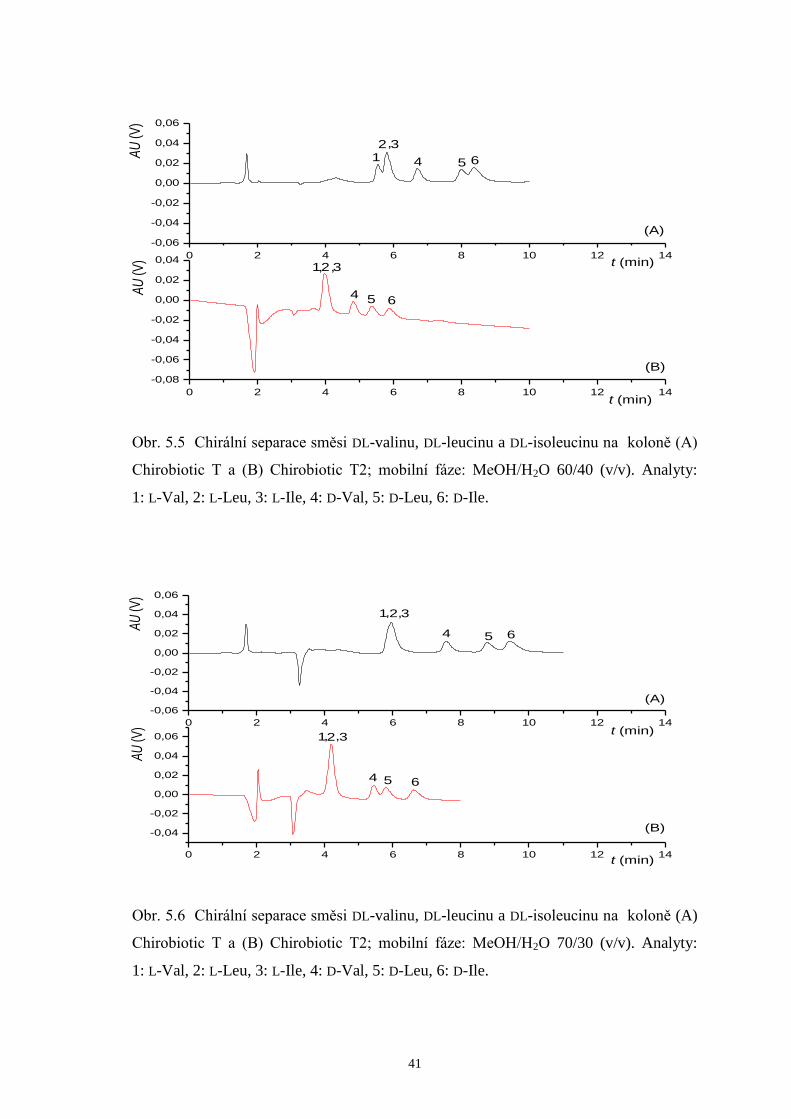
Obr. 5.5 Chirální separace směsi DL-valinu, DL-leucinu a DL-isoleucinu na koloně (A)
Chirobiotic T a (B) Chirobiotic T2; mobilní fáze: MeOH/H2O 60/40 (v/v). Analyty:
1: L-Val, 2: L-Leu, 3: L-Ile, 4: D-Val, 5: D-Leu, 6: D-Ile.
0 2 4 6 8 10 12 14
-0,04
-0,02
0,00
0,02
0,04
0,06
0 2 4 6 8 10 12 14
-0,06
-0,04
-0,02
0,00
0,02
0,04
0,06
65
,,
4
321
(B)
AU
(V)
t (min)
65
,,
4
321
(A)
AU
(V)
t (min)
Obr. 5.6 Chirální separace směsi DL-valinu, DL-leucinu a DL-isoleucinu na koloně (A)
Chirobiotic T a (B) Chirobiotic T2; mobilní fáze: MeOH/H2O 70/30 (v/v). Analyty:
1: L-Val, 2: L-Leu, 3: L-Ile, 4: D-Val, 5: D-Leu, 6: D-Ile.
Page 43
42
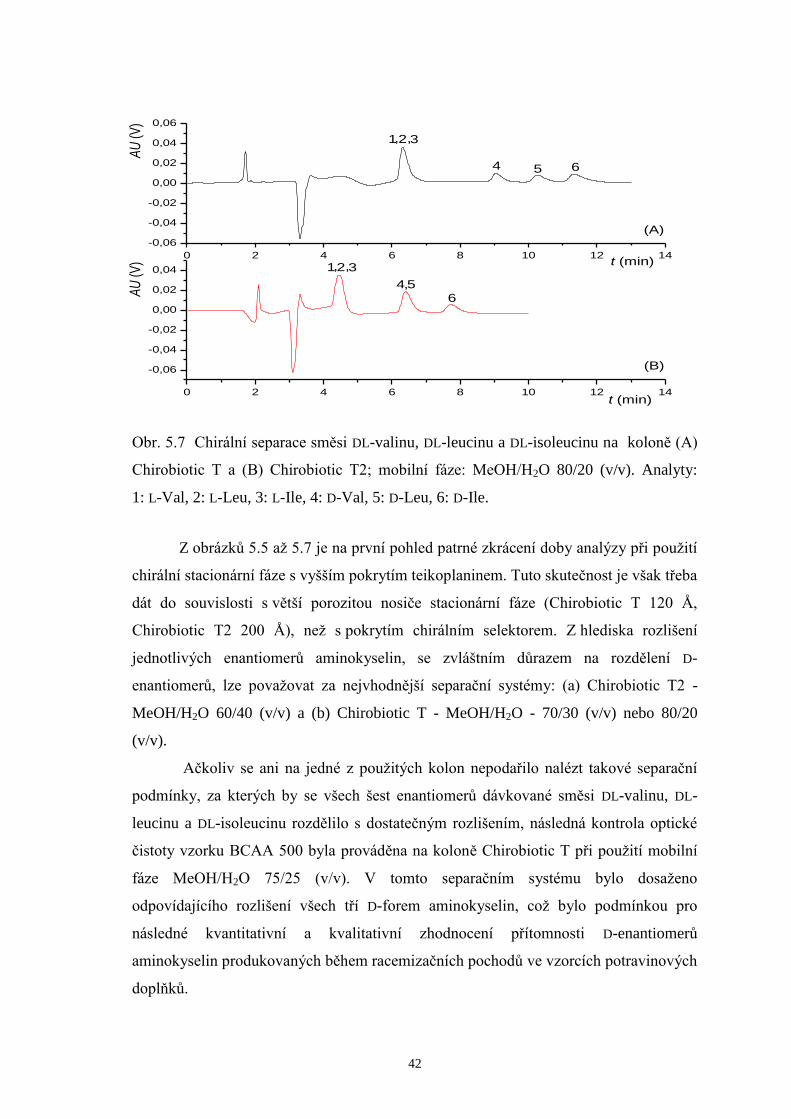
0 2 4 6 8 10 12 14
-0,06
-0,04
-0,02
0,00
0,02
0,04
0 2 4 6 8 10 12 14
-0,06
-0,04
-0,02
0,00
0,02
0,04
0,06
6
,54
,, 321
(B)
AU
(V)
t (min)
65
,,
4
321
(A)
AU
(V)
t (min)
Obr. 5.7 Chirální separace směsi DL-valinu, DL-leucinu a DL-isoleucinu na koloně (A)
Chirobiotic T a (B) Chirobiotic T2; mobilní fáze: MeOH/H2O 80/20 (v/v). Analyty:
1: L-Val, 2: L-Leu, 3: L-Ile, 4: D-Val, 5: D-Leu, 6: D-Ile.
Z obrázků 5.5 až 5.7 je na první pohled patrné zkrácení doby analýzy při použití
chirální stacionární fáze s vyšším pokrytím teikoplaninem. Tuto skutečnost je však třeba
dát do souvislosti s větší porozitou nosiče stacionární fáze (Chirobiotic T 120 Å,
Chirobiotic T2 200 Å), než s pokrytím chirálním selektorem. Z hlediska rozlišení
jednotlivých enantiomerů aminokyselin, se zvláštním důrazem na rozdělení D-
enantiomerů, lze považovat za nejvhodnější separační systémy: (a) Chirobiotic T2 -
MeOH/H2O 60/40 (v/v) a (b) Chirobiotic T - MeOH/H2O - 70/30 (v/v) nebo 80/20
(v/v).
Ačkoliv se ani na jedné z použitých kolon nepodařilo nalézt takové separační
podmínky, za kterých by se všech šest enantiomerů dávkované směsi DL-valinu, DL-
leucinu a DL-isoleucinu rozdělilo s dostatečným rozlišením, následná kontrola optické
čistoty vzorku BCAA 500 byla prováděna na koloně Chirobiotic T při použití mobilní
fáze MeOH/H2O 75/25 (v/v). V tomto separačním systému bylo dosaženo
odpovídajícího rozlišení všech tří D-forem aminokyselin, což bylo podmínkou pro
následné kvantitativní a kvalitativní zhodnocení přítomnosti D-enantiomerů
aminokyselin produkovaných během racemizačních pochodů ve vzorcích potravinových
doplňků.
Page 44
43
5.2 Charakterizace a vzájemné porovnání chirálních stacionárních fází
na bázi teikoplaninu
5.2.1 Waltersův test
Waltersův test se používá pro klasifikaci reverzních achirálních stacionárních
fází. Je založen na zhodnocení převládajících retenčních interakčních mechanismů v
RPLC, hydrofobicitě a silanofilicitě stacionární fáze. Hydrofóbní interakce, které
závisejí na četnosti a délce uhlovodíkových řetězců, jsou vyjádřeny poměrem retenčních
faktorů anthracenu a benzenu v mobilní fázi složené z acetonitrilu a vody (tzv. index
hydrofobicity HI). Silanolové interakce, které jsou závislé na množství přístupných
silanolových skupin, jsou vyjádřeny poměrem retenčních faktorů N,N-diethyltoluamidu
(N,N-DETA) a anthracenu v mobilní fázi tvořené čistým acetonitrilem (tzv. silanolový
index SI). Právě v čistém acetonitrilu jsou dobře solvatovány alkylové řetězce (např.
C18, C8). Zadržování N,N-DETA souvisí se silanolovou aktivitou stacionární fáze,
zatímco retence anthracenu je spojována výlučně s hydrofóbními interakcemi67
.
Waltersův test byl proveden na kolonách Chirobiotic T a T2. Vypočtené indexy
hydrofobicity a silanolové indexy jsou znázorněny v tabulce 5.2.
Tabulka 5.2 Index hydrofobicity (HI) a silanolový index (SI) změřené na kolonách
Chirobiotic T a Chirobiotic T2.
kolona HI
k (anthracen)/k (benzen) SI
k (N,N-DETA)/k (anthracen)
Chirobiotic T 0,95 2,17
Chirobiotic T2 1,18 1,76
Waltersův test sice není navržen pro porovnávání kolon na bázi teikoplaninu a
podobných, a také reverzní systém s použitím teikoplaninových CSP není typickým
reverzním systémem, přesto lze z hodnot uvedených v tabulce 5.2 učinit určité závěry.
Relativně nízké hodnoty HI potvrzují skutečnost, že teikoplaninové kolony se od
reverzních fází typu C18, kde se hodnoty HI pohybují v rozmezí 3 - 4 (podle konkrétní
kolony), značně liší. Přitom CSP Chirobiotic T2 s vyšším pokrytím chirálním
selektorem má hodnotu HI poněkud vyšší než kolona Chirobiotic T. Rozdíl mezi oběma
chirálními stacionárními fázemi z pohledu indexu hydrofobicity však není velký. Tato
Page 45
44
skutečnost vyvolává otázku, jak moc se pokrytí obou stacionárních fází teikoplaninem
liší. Výrobce totiž hodnoty pokrytí neudává.
Srovnání hodnot silanolového indexu koresponduje s výsledky získanými pro
HI, tj. vyšší hodnotě HI pro Chirobiotic T2 odpovídají nižší hodnoty SI, zatímco pro
kolonu Chirobiotic T je tomu naopak. Silanolové interakce se v retenčním mechanismu
tedy zřejmě uplatňují více při použití kolony s menším pokrytím chirálním selektorem.
Větší rozdíl v hodnotách SI pro zkoumané kolony oproti hodnotám HI má však
pravděpodobně souvislost i s delším řetězcem („spacerem“), kterým je teikoplanin
vázán na silikagelový nosič v případě kolony Chirobiotic T2, kde lze tudíž předpokládat
větší odstínění silanolových skupin, než v případě kolony Chirobiotic T.
5.2.2 Porovnání retenčních faktorů enantiomerů valinu, leucinu a isoleucinu na
kolonách Chirobiotic T, T2 a TAG
Retenční faktor charakterizuje retenci, tzn. jak moc jsou analyty na koloně
zadržovány a zpožďovány.
Porovnání retenčních faktorů jednotlivých enantiomerů aminokyselin valinu,
leucinu a isoleucinu v závislosti na procentuálním obsahu MeOH v mobilní fázi na
kolonách Chirobiotic T, T2 a TAG jsou znázorněna na obrázcích 5.8 až 5.13. Při měření
na koloně Chirobiotic TAG byla dávkována směs DL-valinu, DL-leucinu a DL-
isoleucinu, v případě kolon Chirobiotic T a T2 byly racemáty těchto tří aminokyselin
dávkovány zvlášť.
Page 46
45
0
0,5
1
1,5
2
2,5
3
3,5
4
10 20 30 40 60 70 80 90
% MeOH
k
T
T2
TAG
Obr. 5.8 Porovnání retenčních faktorů aminokyseliny L-valinu na kolonách Chirobiotic
T, T2 a TAG při použití mobilních fází MeOH/H2O 10/90 (v/v) až MeOH/H2O 90/10
(v/v).
Obr. 5.9 Porovnání retenčních faktorů aminokyseliny L-leucinu na kolonách Chirobiotic
T, T2 a TAG při použití mobilních fází MeOH/H2O 10/90 (v/v) až MeOH/H2O 90/10
(v/v).
0
0,5
1
1,5
2
2,5
3
3,5
4
10 20 30 40 60 70 80 90
% MeOH
k
T
T2
TAG
Page 47
46
Obr. 5.10 Porovnání retenčních faktorů aminokyseliny L-isoleucinu na kolonách
Chirobiotic T, T2 a TAG při použití mobilních fází MeOH/H2O 10/90 (v/v) až
MeOH/H2O 90/10 (v/v).
Obr. 5.11 Porovnání retenčních faktorů aminokyseliny D-valinu na kolonách
Chirobiotic T, T2 a TAG při použití mobilních fází MeOH/H2O 10/90 (v/v) až
MeOH/H2O 90/10 (v/v).
0
0,5
1
1,5
2
2,5
3
3,5
4
10 20 30 40 60 70 80 90
% MeOH
k
T
T2
TAG
0
1
2
3
4
5
6
7
8
9
10 20 30 40 60 70 80 90
% MeOH
k
T
T2
TAG
Page 48
47
Obr. 5.12 Porovnání retenčních faktorů aminokyseliny D-leucinu na kolonách
Chirobiotic T, T2 a TAG při použití mobilních fází MeOH/H2O 10/90 (v/v) až
MeOH/H2O 90/10 (v/v).
Obr. 5.13 Porovnání retenčních faktorů aminokyseliny D-isoleucinu na kolonách
Chirobiotic T, T2 a TAG při použití mobilních fází MeOH/H2O 10/90 (v/v) až
MeOH/H2O 90/10 (v/v).
0
1
2
3
4
5
6
7
8
9
10 20 30 40 60 70 80 90
% MeOH
k
T
T2
TAG
0
1
2
3
4
5
6
7
8
9
10 20 30 40 60 70 80 90
% MeOH
k
T
T2
TAG
Page 49
48
Porovnáme-li kolony Chirobiotic T, T2 a TAG z hlediska retence analytů (obr.
5.8 až 5.13), je zřejmé, že nejnižších retenčních faktorů bylo pro všechny enantiomery
dosaženo na koloně Chirobiotic T2. Nejvyšší retence bylo naopak dosaženo na koloně
Chirobiotic T pro L-enantiomery a pro D-enantiomery na koloně Chirobiotic TAG. Ze
závislostí retenčních faktorů na množství MeOH v mobilní fázi dále vyplývá, že s
rostoucím podílem MeOH v mobilní fázi většinou roste retence u všech šesti
enantiomerů na všech třech použitých kolonách. Tento trend je dobře patrný zejména na
koloně Chirobiotic T, kde u mobilních fází s nižším procentem MeOH retenční faktor
vzrůstá jen mírně, u L-leucinu a L-isoleucinu dokonce nepatrně klesá (obr. 5.9 a 5.10),
avšak u mobilních fází s nadpolovičním podílem MeOH již retence u všech šesti
enantiomerů prudce roste.
Na koloně Chirobiotic T2 již není výše popsaný trend tolik výrazný. Při nižším
procentu MeOH v mobilní fázi retenční faktor (s výjimkou L-leucinu, obr. 5.9) mírně
vzrůstá. Při porovnání retence za použití mobilní fáze MeOH/H2O 10/90 (v/v) a
MeOH/H2O 20/80 (v/v) lze konstatovat, že zadržování všech analytů (kromě D-valinu a
D-isoleucinu, obr. 5.11 a 5.13) na této koloně je vyšší při použití mobilní fáze s nižším
obsahem MeOH. V mobilních fázích s nadpolovičním obsahem organické fáze se
retence analytů na této koloně, stejně tak jako u Chirobiotic T, zvyšuje. Výjimku lze
nalézt u retenčních faktorů L-leucinu, L-isoleucinu a D-leucinu (obr. 5.9, 5.10 a 5.12) v
mobilní fázi s 90 %obj. MeOH. Důvodem změny retence může být fakt, že při použití
mobilní fáze s nadbytkem organického modifikátoru se mohou při separačním
mechanismu spíše uplatňovat interakce, které jsou charakterističtější pro polárně-
organický separační mod.
Na koloně Chirobiotic TAG bylo měřeno pouze za použití mobilních fází s 30 až
70 %obj. MeOH. Retenční faktory se na této koloně také zvyšují s rostoucím podílem
MeOH v mobilní fázi, dobře patrné je to zejména u všech tří D-enantiomerů (obr. 5.11
až 5.13). Retence L-leucinu v mobilních fázích 30 až 60 %obj. MeOH také vzrůstá, při
70 % MeOH však již klesá (obr. 5.9). V poslední zmíněné mobilní fázi eluuje L-leucin
v jednom píku společně s L-valinem a L-isoleucinem (viz obr. 5.1). Retence
jednotlivých L-enantiomerů, jejichž retenční časy jsou velmi blízké, může být navíc
vzájemně ovlivňována.
Za nejvýraznější výsledek, vyplývající z obr. 5.8 až 5.13, lze považovat
minimální vliv složení mobilní fáze na retenci všech sledovaných analytů na koloně
Chirobiotic T2. Z analytického pohledu má tato skutečnost pozitivní dopad na
Page 50
49
robustnost metody. Naopak negativním důsledkem je menší prostor pro optimalizaci
separace.
Grafy na obrázcích 5.14 až 5.17 slouží k vzájemnému porovnání retenčních
faktorů L- resp. D-enantiomerů aminokyselin na procentuálním podílu MeOH
v mobilních fázích na kolonách Chirobiotic T a T2.
1,5
1,7
1,9
2,1
2,3
2,5
2,7
2,9
3,1
3,3
0 20 40 60 80 100
% MeOH
k
L-Val
L-Leu
L-Ile
Obr. 5.14 Porovnání retenčních faktorů aminokyselin L-valinu, L-leucinu a L-isoleucinu
na koloně Chirobiotic T při použití mobilních fází MeOH/H2O 10/90 (v/v) až
MeOH/H2O 90/10 (v/v).
Page 51
50
1
2
3
4
5
6
7
8
9
0 20 40 60 80 100
% MeOH
k
D-Val
D-Leu
D-Ile
Obr. 5.15 Porovnání retenčních faktorů aminokyselin D-valinu, D-leucinu a
D-isoleucinu na koloně Chirobiotic T při použití mobilních fází MeOH/H2O 10/90 (v/v)
až MeOH/H2O 90/10 (v/v).
Obr. 5.16 Porovnání retenčních faktorů aminokyselin L-valinu, L-leucin a L-isoleucinu
na koloně Chirobiotic T2 při použití mobilních fází MeOH/H2O 10/90 (v/v) až
MeOH/H2O 90/10 (v/v).
1,2
1,3
1,4
1,5
1,6
1,7
1,8
1,9
0 20 40 60 80 100
% MeOH
k
L-Val
L-Leu
L-Ile
Page 52
51
Obr. 5.17 Porovnání retenčních faktorů aminokyselin D-valinu, D-leucinu a
D-isoleucinu na koloně Chirobiotic T2 při použití mobilních fází MeOH/H2O 10/90
(v/v) až MeOH/H2O 90/10 (v/v).
Jak je patrné z grafů na obrázcích 5.14 až 5.17, retence aminokyselin valinu,
leucinu a isoleucinu v mobilních fázích s nižším obsahem methanolu na obou
studovaných kolonách velmi dobře korelují s hodnotami log P (tab. 4.1).
Nejhydrofóbnější DL-leucin je při použití mobilních fází s nadpolovičním podílem H2O
zadržován nejvíce. Naopak nejnižší hodnotu log P má DL-valin, u něhož se hydrofóbní
interakce uplatňují již v menší míře a je v těchto mobilních fázích zadržován nejméně.
Z grafů závislosti retenčních faktorů L-enantiomerů a D-enantiomerů na podílu
MeOH v mobilní fázi dále vyplývá, že na koloně Chirobiotic T i T2 dochází při
zvyšujícím se množství MeOH v mobilní fázi k záměně elučního pořadí některých
enantiomerů.
Na koloně Chirobiotic T se při použití mobilní fáze obsahující přibližně 66 %obj.
MeOH mění eluční pořadí L-valinu a L-isoleucinu, od ~ 78 %obj. MeOH v mobilní fázi
byly získány stejné hodnoty retenčních faktorů L-leucinu a L-valinu (obr. 5.14). Rovněž
eluční pořadí mezi D-enantiomery se na koloně Chirobiotic T mění; při ~ 45 %obj.
MeOH v mobilní fázi se zaměňuje eluce mezi D-leucinem a D-isoleucinem (obr. 5.15).
Zatímco v případě CSP Chirobiotic T jsou získané závislosti (obr. 5.14 a 5.15)
relativně konzistentní, tj. trend závislostí je jednoznačný, pouze dochází při určitém
složení mobilní fáze k záměně elučního pořadí dvou enantiomerů, grafy zkonstruované
pro CSP s vyšším pokrytím teikoplaninem (obr. 5.16 a 5.17) jsou mnohem
1
1,5
2
2,5
3
3,5
4
0 20 40 60 80 100
% MeOH
k
D-Val
D-Leu
D-Ile
Page 53
52
komplikovanější (obzvláště v případě závislostí pro L-enantiomery), a tudíž obtížněji
interpretovatelné. Získané výsledky je možné přibližně popsat následujícím způsobem.
Na koloně Chirobiotic T2 lze, ve srovnání s Chirobiotic T, ještě častěji pozorovat
změnu elučního pořadí se zvyšujícím se množstvím MeOH v mobilní fázi. Při použití
mobilní fáze obsahující ~ 53 %obj. MeOH se zaměňuje eluce mezi L-valinem a L-
isoleucinem (L-Ile eluuje před L-Val), dále se při ~ 60 %obj. MeOH mění pořadí eluce
mezi L-valinem a L-leucinem (L-Leu eluuje před L-Val). Se zvýšením podílu MeOH na
72 %obj. se zaměňuje eluční pořadí mezi L-leucinem a L-isoleucinem (L-Leu eluuje před
L-Ile) (obr. 5.16). Také mezi D-analogy dochází k záměně eluce; při ~ 42 %obj. MeOH
v mobilní fázi se zaměňuje eluce D-leucinu a D-isoleucinu (D-Leu eluuje před D-Ile), při
79 %obj. MeOH se mění pořadí D-valinu a D-leucinu (D-Leu eluuje před D-Val)
(obr. 5.17).
5.2.3 Porovnání separačních faktorů enantiomerů valinu, leucinu a isoleucinu na
kolonách Chirobiotic T a T2
Separační faktor vyjadřuje selektivitu separace.
Na obrázcích 5.18 až 5.20 jsou znázorněny grafy závislosti separačního faktoru
DL-valinu, DL-leucinu a DL-isoleucinu na složení mobilní fáze na kolonách Chirobiotic
T a T2.
Page 54
53
Obr. 5.18 Závislost separačního faktoru DL-valinu na procentuálním obsahu methanolu
v mobilní fázi na kolonách Chirobiotic T a T2. Body jsou proloženy polynomem
2. stupně.
Obr. 5.19 Závislost separačního faktoru DL-leucinu na procentuálním obsahu
methanolu v mobilní fázi na kolonách Chirobiotic T a T2. Body jsou proloženy
polynomem 2. stupně.
1
1,2
1,4
1,6
1,8
2
2,2
0 20 40 60 80 100
% MeOH
α
T
T2
1
1,2
1,4
1,6
1,8
2
2,2
2,4
0 20 40 60 80 100
% MeOH
α
T
T2
Page 55
54
Obr. 5.20 Závislost separačního faktoru DL-isoleucinu na procentuálním obsahu
methanolu v mobilní fázi na kolonách Chirobiotic T a T2. Body jsou proloženy
polynomem 2. stupně.
Jak je zřejmé z obrázků 5.18 až 5.20, s rostoucím podílem MeOH v mobilní fázi
roste selektivita separačního systému vůči separovaným analytům. U nižších obsahů
MeOH v mobilní fázi není příliš patrný rozdíl v selektivitě mezi kolonami Chirobiotic T
a T2 (zejména u DL-valinu a DL-isoleucinu, obr. 5.18 a 5.20). U nadpolovičního podílu
MeOH v mobilní fázi se zvyšuje rozdíl v selektivitě mezi oběma kolonami. Vyšších
hodnot separačních faktorů bylo ve všech třech případech dosaženo na koloně
Chirobiotic T. Nejnižší separační faktor byl získán u DL-valinu na koloně Chirobiotic
T2 v mobilní fázi MeOH/H2O 10/90 (v/v), naopak nejvyšší selektivita byla dosažena u
DL-isoleucinu na koloně Chirobiotic T při použití mobilní fáze MeOH/H2O 90/10 (v/v).
Z celkového pohledu bylo nejnižší selektivity dosaženo u DL-valinu a nejvyšší u DL-
isoleucinu.
1
1,4
1,8
2,2
2,6
3
0 20 40 60 80 100
% MeOH
α
T
T2
Page 56
55
5.2.4 Porovnání rozlišení enantiomerů valinu, leucinu a isoleucinu na kolonách
Chirobiotic T a T2
Rozlišení charakterizuje míru rozdělení dvou analytů (píků), popř. míru
vzájemného překrývání dvou sousedních píků.
Na obrázcích 5.21 až 5.23 jsou znázorněny grafy závislosti rozlišení DL-valinu,
DL-leucinu a DL-isoleucinu na složení mobilní fáze na kolonách Chirobiotic T a T2.
Obr. 5.21 Závislost rozlišení DL-valinu na procentuálním zastoupení methanolu
v mobilní fázi na kolonách Chirobiotic T a T2.
0
1
2
3
4
5
6
7
8
10 20 30 40 60 70 80 90
% MeOH
R
T
T2
Page 57
56
Obr. 5.22 Závislost rozlišení DL-leucinu na procentuálním zastoupení methanolu
v mobilní fázi na kolonách Chirobiotic T a T2.
Obr. 5.23 Závislost rozlišení DL-isoleucinu na procentuálním zastoupení methanolu
v mobilní fázi na kolonách Chirobiotic T a T2.
Z grafů závislosti rozlišení na obsahu MeOH v mobilní fázi na obrázcích 5.21 až
5.23 vyplývá, že s rostoucím procentuálním podílem MeOH v mobilní fázi roste
rozlišení L- i D-enantiomerů všech tří měřených aminokyselin. Vyšších hodnot rozlišení
bylo dosaženo ve všech případech na koloně Chirobiotic T. Dostatečně rozlišit (R ≥ 1,5)
0
1
2
3
4
5
6
7
8
10 20 30 40 60 70 80 90
% MeOH
R
T
T2
0
1
2
3
4
5
6
7
8
10 20 30 40 60 70 80 90
% MeOH
R
T
T2
Page 58
57
enantiomery DL-valinu se podařilo na koloně Chirobiotic T při použití mobilní fáze
s obsahem vyšším než 30 %obj. MeOH, na Chirobiotic T2 s obsahem min. 40 %obj.
MeOH. U enantiomerů DL-leucinu a DL-isoleucinu bylo dostatečného rozlišení
dosaženo na obou kolonách již v mobilní fázi s nejnižším zkoumaným množstvím
MeOH, tedy 10 %obj..
V grafu na obr. 5.23 (DL-isoleucin) je na obou separačních kolonách zřejmý
pokles hodnoty rozlišení v mobilní fázi MeOH/H2O 90/10 (v/v). Tento jev lze vysvětlit
v souvislosti s výsledky uvedenými v kap. 5.2.3, tj. s jednoznačným trendem závislosti
pro separační faktor, zhoršením symetrie píků (viz kap. 5.2.5) resp. účinnosti separace
při vyšších hodnotách retence (viz rovnice (4) na str. 34). Dalšími faktory, které mohou
mít vliv na pozorovaný pokles rozlišení enantiomerů isoleucinu, může být zatížení
výsledků získaných při použití mobilní fáze s vysokým obsahem MeOH větší chybou,
neboť malé a rozmyté píky bylo obtížné integrovat. Lze také zvážit i změnu separačního
mechanismu při přechodu k polárně organickému modu.
5.2.5 Porovnání symetrie píků enantiomerů valinu, leucinu a isoleucinu na
kolonách Chirobiotic T a T2
Faktor symetrie charakterizuje tvar píku daného analytu. Pokud je hodnota
faktoru symetrie rovna jedné, pík je zcela symetrický (má gaussovský tvar), pokud je
menší než jedna, pík je nesymetrický vpředu, pokud je větší než jedna, pík je rozmytý,
tak zvaně chvostuje.
Na obrázcích 5.24 až 5.29 jsou znázorněny grafy závislosti symetrie píků
enantiomerů valinu, leucinu a isoleucinu na složení mobilní fáze na kolonách
Chirobiotic T a T2.
Page 59
58
Obr. 5.24 Závislost symetrie píku L-valinu na procentuálním obsahu methanolu
v mobilní fázi na kolonách Chirobiotic T a T2.
0
0,5
1
1,5
2
2,5
10 20 30 40 60 70 80 90
% MeOH
S
T
T2
Obr. 5.25 Závislost symetrie píku L-leucinu na procentuálním obsahu methanolu
v mobilní fázi na kolonách Chirobiotic T a T2.
0
0,5
1
1,5
2
2,5
10 20 30 40 60 70 80 90
% MeOH
S
T
T2
Page 60
59
0
0,5
1
1,5
2
2,5
10 20 30 40 60 70 80 90
% MeOH
S
T
T2
Obr. 5.26 Závislost symetrie píku L-isoleucinu na procentuálním obsahu methanolu
v mobilní fázi na kolonách Chirobiotic T a T2.
0
0,5
1
1,5
2
2,5
10 20 30 40 60 70 80 90
% MeOH
S
T
T2
Obr. 5.27 Závislost symetrie píku D-valinu na procentuálním obsahu methanolu
v mobilní fázi na kolonách Chirobiotic T a T2.
Page 61
60
0
0,5
1
1,5
2
2,5
10 20 30 40 60 70 80 90
% MeOH
S
T
T2
Obr. 5.28 Závislost symetrie píku D-leucinu na procentuálním obsahu methanolu
v mobilní fázi na kolonách Chirobiotic T a T2.
0
0,5
1
1,5
2
2,5
10 20 30 40 60 70 80 90
% MeOH
S
T
T2
Obr. 5.29 Závislost symetrie píku D-isoleucinu na procentuálním obsahu methanolu
v mobilní fázi na kolonách Chirobiotic T a T2.
Z grafů na obrázcích 5.24 až 5.29 vyplývá, že hodnoty faktoru symetrie jsou u
všech enantiomerů na obou kolonách větší, než jedna, čili u všech píků docházelo
k jejich rozmývání. Jedinou výjimkou je pouze L-valin v mobilní fázi MeOH/H2O 10/90
(v/v) (obr. 5.24), kde faktor symetrie dosahoval hodnoty mírně pod jedna. Z celkového
pohledu se zdá být poněkud lepší symetrie píků na koloně Chirobiotic T2, což je patrné
Page 62
61
zejména u mobilních fází s vyšším obsahem MeOH. Při nižších obsazích MeOH
v mobilních fázích je naopak nižších hodnot faktoru symetrie u většiny enantiomerů
dosahováno na koloně Chirobiotic T.
V případě píku L-valinu (obr. 5.24) je na koloně Chirobiotic T zřejmý růst
hodnoty faktoru symetrie se zvyšujícím se podílem MeOH v mobilní fázi. To má také
souvislost s retenčním faktorem - čím déle je analyt na koloně zadržován, tím více
dochází k rozmývání jeho zóny. Na koloně Chirobiotic T2 rovněž u nižších obsahů
MeOH faktor symetrie roste, s nadpolovičním podílem MeOH v mobilní fázi však
zůstává přibližně konstantní resp. klesá.
Symetrie píku L-leucinu (obr. 5.25) na koloně Chirobiotic T opět mírně vzrůstá, i
když hodnoty faktoru symetrie jsou u nižších a vyšších obsahů MeOH vcelku
vyrovnané. Na koloně Chirobiotic T2 symetrie píku L-leucinu s rostoucím množstvím
MeOH spíše klesá.
Hodnoty faktoru symterie píku L-isoleucinu (obr. 5.26) jsou na koloně
Chirobiotic u nižších obsahů MeOH opět vyrovnané, s nadpolovičním podílem MeOH
se faktor symetrie zvyšuje. Na koloně Chirobiotic T2 faktor symetrie s rostoucím
množstvím MeOH mírně klesá, i když hodnoty jsou u všech používaných mobilních
fází velmi vyrovnané. Lze říci, že výsledky získané pro leucin a isoleucin jsou
analogické.
V grafu na obrázku 5.27 nejsou vyneseny hodnoty faktoru symetrie u mobilních
fází s 10 až 30 %obj. MeOH. Při těchto obsazích MeOH se totiž píky L- a D-leucinu
natolik překrývaly, že software nebyl schopen vypočítat faktor symetrie píku D-leucinu.
U mobilních fází s vyšším obsahem MeOH je však již patrný růst faktoru symetrie se
stoupajícím procentem MeOH v mobilní fázi, zejména na koloně Chirobiotic T.
Pro symetrie píků D-leucinu a D-isoleucinu (obr. 5.28, 5.29) nelze jednoznačně
říci, zda-li má závislost faktoru symetrie na složení mobilní fáze stoupající nebo
klesající charakter. Vzhledem k tomu, že se píky D-leucinu a D-isoleucinu při nižších
obsazích MeOH v mobilní fázi překrývaly s píky jejich L-analogů, nejsou výpočty
hodnot faktorů symetrie zcela přesné.
Page 63
62
5.3 Určení meze detekce a meze stanovení DL-valinu, DL-leucinu
a DL-isoleucinu
Kalibrace byla provedena na koloně Chirobiotic T při použití mobilní fáze
optimalizovaného složení MeOH/H2O 75/25 (v/v). V tomto separačním systému
eluovaly L-aminokyseliny v jednom píku, zatímco D-enantiomery eluovaly každý
zvlášť. Z tohoto důvodu jsou hodnoty meze detekce (LOD) a meze stanovení (LOQ)
vypočítány pro každou D-aminokyselinu zvlášť, zatímco hodnoty LOD a LOQ pro L-
enantiomery představují sumu koncentrací všech tří L-forem.
Na kolonu byla dávkována směs standardů DL-valinu, DL-leucinu a DL-
isoleucinu rozpuštěných v H2O o pěti různých koncentracích získaných postupným
ředěním výchozí koncentrace směsi aminokyselin. Jednotlivá měření byla provedena
vždy třikrát. Koncentrace jednotlivých enantiomerů aminokyselin použitých k
proměření kalibrační závislosti jsou uvedeny v tabulce 5.3.
Tabulka 5.3 Koncentrace standardů aminokyselin valinu, leucinu a isoleucinu použité
pro kalibraci.
l á t k o v á k o n c e n t r a c e (mg/ml)
L-Val, L-Leu, L-Ile D-Val D-Leu D-Ile
0,3137 0,1047 0,1041 0,1050
0,6275 0,2094 0,2081 0,2100
1,2550 0,4188 0,4163 0,4200
2,5100 0,8375 0,8325 0,8400
5,0200 1,6750 1,6650 1,6800
Kalibrační přímky pro jednotlivé enantiomery byly sestrojeny ze závislosti
výšky píku (μV) na koncentraci analytu (mg/ml). Odečtená výška šumu základní linie
(σ) činila 4 μV. Z jednotlivých kalibračních rovnic byly vypočítány limity detekce a
limity stanovení pro každý enantiomer dosazením hodnot 3σ resp. 10σ za y.
Výsledky získané z kalibračních závislostí jsou uvedeny v tabulce 5.4.
Page 64
63
Tabulka 5.4 Kalibrační rovnice, koeficienty spolehlivosti, limity detekce (LOD) a
limity stanovení (LOQ) enantiomerů aminokyselin valinu, leucinu a isoleucinu pro
optimalizovanou metodu k hodnocení enantiomerní čistoty měřeného vzorku.
aminokyselina kalibrační rovnice R LOD
(10-4
mg/ml) LOQ
(10-4
mg/ml)
L-Val, L-Leu, L-Ile y = 46913x + 13874 0,998 2,558 6,395
D-Val y = 41001x + 4221 0,998 2,927 7,317
D-Leu y = 21931x + 2677 0,997 5,472 13,679
D-Ile y = 15134x + 3312 0,991 7,929 19,823
Hodnoty limitů detekce se pohybovaly řádově v desetinách µg/ml a hodnoty
limitů stanovení dosahovaly v případě D-enantiomerů aminokyselin spíše jednotek
µg/ml. Z tabulky 5.4 je zřejmé, že hodnoty LOD i LOQ rostou v korelaci s elučním
pořadím enantiomerů, což souvisí se zhoršováním tvarů píků analytů více zadržovaných
na koloně.
5.4 Studium procesů vyvolávajících racemizaci aminokyselin ve vzorku
Jedním z cílů této části práce bylo určit podmínky prostředí, které se případně
mohou podílet na racemizaci L-aminokyselin obsažených v měřeném vzorku. Studován
byl vliv teploty, světla a rovněž kyselého pH (po orálním příjmu BCAA v lidském
žaludku).
Nejprve byl na kolonu nadávkován zásobní roztok vzorku skladovaného v
chladu a temnu. Po kontrole jeho enantiomerní čistoty byl vzorek vystaven výše
zmíněným podmínkám a byl zkoumán jejich potenciální vliv na racemizaci L-valinu, L-
leucinu a L-isoleucinu ve vzorku. Použité pracovní postupy jsou přesně popsány
v kap. 4.9.
Přítomnost D-enantiomerů však v žádném ze sledovaných vzorků prokázána
nebyla. Pro tento fakt jsou dvě možná vysvětlení:
Aminokyseliny obsažené ve vzorku nepodléhají racemizaci za daných
podmínek, což je příznivou zprávou pro ty, kteří výživové BCAA doplňky
užívají. V práci64
, zabývající se enantioseparací DL-aminokyselin s rozvětveným
řetězcem metodou kapilární elektroforézy, byla také potvrzena optická čistota
vybraných komerčních BCAA doplňků pro sportovní výživu.
Page 65
64
V úvahu připadá také možnost, že proces racemizace proběhl, ale množství
vzniklých D-analogů aminokyselin bylo nízké a vzhledem k malé citlivosti
používaného UV/VIS detektoru byla hodnota koncentrace D-enantiomerů pod
mezí detekce (LOD).
Pro ověření jedné či druhé hypotézy by bylo nutné provést derivatizaci vhodným
činidlem, které by zvýšilo citlivost detekce.
Page 66
65
6 ZÁVĚR
V této práci byla optimalizována metoda vysokoúčinné kapalinové
chromatografie pro separaci enantiomerů aminokyselin valinu, leucinu a isoleucinu.
L-formy těchto aminokyselin jsou obsaženy v BCAA potravinových doplňcích pro
sportovce.
Měření byla postupně prováděna na separačních kolonách Chirobiotic T, T2 a
TAG. Pro účely následné kontroly enantiomerní čistoty vzorku bylo nejlepší chirální
separace dosaženo na koloně Chirobiotic T s mobilní fází MeOH/H2O 75/25 (v/v). Při
tomto složení chromatografického systému se sice nepodařilo vzájemně oddělit L-
enantiomery aminokyselin, ovšem D-formy byly dostatečně rozlišeny, což bylo pro
kontrolu přítomnosti D-enantiomerů v měřeném vzorku velmi vhodné.
Všechny tři používané chirální stacionární fáze byly porovnány z hlediska
retence analytů. Na koloně Chirobiotic T byly L-enantiomery aminokyselin zadržovány
ze studovaných kolon nejvíce. Naopak retenční faktory D-enantiomerů byly nejvyšší na
koloně Chirobiotic TAG.
Dále byly podrobně studovány a porovnávány kolony Chirobiotic T a T2
z hlediska elučního pořadí, separačních faktorů, rozlišení a rozmývání zón analytů.
Eluční pořadí bylo na všech kolonách na bázi teikoplaninu ve všech zkoumaných
separačních systémech vždy stejné - L-enantiomer eluoval před D-aminokyselinou. Při
změně složení mobilní fáze docházelo, především na stacionární fázi Chirobiotic T2,
k záměně retenčního pořadí jednotlivých D-aminokyselin. Na koloně Chirobiotic T bylo
dosaženo lepší selektivity separace enantiomerů. Vyšších hodnot rozlišení píků D- a L-
enantiomerů jednotlivých aminokyselin bylo jednoznačně dosaženo rovněž na koloně
Chirobiotic T. Dále byly tyto dvě kolony porovnávány z hlediska symetrie píků. Z
výsledků nelze vyvodit zcela jednoznačný závěr, na které koloně docházelo k menšímu
rozšiřování zón měřených aminokyselin. Z celkového pohledu je však patrné, že na
koloně Chirobiotic T2 bylo dosaženo účinnější separace enantiomerů studovaných
aminokyselin.
Na kolonách Chirobiotic T a T2 byl také proveden Waltersův test, ze kterého
vyplynulo, že poněkud vyšší index hydrofobicity měla stacionární fáze s větším
pokrytím chirálním selektorem. Rozdíl v hodnotách HI mezi oběma kolonami však
nebyl velký. Naopak vyšší hodnotu silanolového indexu měla stacionární fáze
Page 67
66
Chirobiotic T - silanolové interakce se v retenčním mechanismu tedy zřejmě uplatňují
více při použití kolony s menším pokrytím chirálním selektorem.
Dalším z cílů této práce bylo sledování procesů, které mohou ve vzorku vyvolat
racemizaci aminokyselin. Přítomnost D-enantiomerů však v žádném ze vzorků
detekována nebyla.
Page 68
67
SEZNAM LITERATURY
1. Tesařová E., Armstrong D. W., v knize: Advanced Chromatographic and
Electromigration Methods in BioSciences (Deyl Z., ed.), kap. 5. Elsevier, Amsterdam
1998.
2. Streblová E.: Souhrnné texty z chemie, II. díl. Univerzita Karlova v Praze, Praha
2000.
3. Trnka T., Klinotová E., Kotora M., Sejbal J.: Organická chemie. Univerzita Karlova
v Praze, Praha 2003.
4. Červinka O.: Chem. Listy 93, 294 (1999).
5. Cahn R. S., Ingold C. K., Prelog V.: Angew. Chem. 5, 385 (1966).
6. Maier N. M., Franco P., Lindner W.: J. Chromatogr. A 906, 3 (2001).
7. Marchelli R., Dossena A., Palla G.: Trends Food Sci. Technol. 7, 113 (1996).
8. MacDermott A. J.: Origins Life Evol. Biosphere 25, 191 (1995).
9. Fassihi A. R.: Int. J. Pharm. 92, 1 (1993).
10. Knocke B., Blaschke G.: J. Chromatogr. A 666, 235 (1994).
11. Srkalová S.: Bakalářská práce, PřF UK, Praha 2006.
12. Srkalová S., Kalíková S., Tesařová E.: Chem. Listy, přijato k tisku.
13. Ekborg –Ott K. H., Armstrong D. W., v knize: Chiral Separations: Applications and
Technology (Ahuja S., ed.), kap. 9. American Chemical Society, Washington, DC
1997.
14. Pickenhagen W., v knize: Flavor Chemistry – Trends and Developments (Teranishi
R., Buttery R. G., Shahidi F., eds.). American Chemical Society, Washington, DC
1989.
15. Simó C., Barbas C., Cifuentes A.: Electrophoresis 24, 2431 (2003).
16. Lekh R. J., Djong-Chi C., Tsutomu O., Yukiko N., Hidehiko Y.: Trends Food Sci.
Technol. 10, 199 (1999).
17. Ekborg-Ott K. H., Armstrong D. W., Taylor A. D.: J. Agric. Food Chem. 50, 473
(2002).
18. Liardon R., Lederman S.: J. Agric. Food Chem. 34, 557 (1986).
19. Tovar L. R., Schwass D. E., v knize: Xenobiotics in Foods and Feeds (Finley J. W.,
Schwass D. E., eds.). American Chemical Society, Washington, DC 1983.
Page 69
68
20. Nehring U. P.: Proceedings of the Symposium on Chemical Reactions in Foods
(Velíšek J., ed.). VUPP-STI, Praha 1992.
21. Brückner H., Jaek P., Langer M., Godel H.: Amino Acids 2, 271 (1992).
22. Simó C., Barbas C., Cifuentes A.: J. Agric. Food Chem. 50, 5288 (2002).
23. Friedman M. J.: J. Agric. Food Chem. 47, 3457 (1999).
24. Poncini L., Wimmer F. L., Vakamoce V.: Fiji Agric. J. 45, 65 (1983).
25. Casal S., Alves A. R., Mendes E., Oliveira M. B. P. P., Ferreira M. A.: J. Agric.
Food Chem. 51, 6495 (2003).
26. Pantó F: v knize: Progress in Food Fermentation, Proceedings of the Euro Food
Chem. (Benedito de Barber C., ed.), sv. VII. IATA-CSIC, Valencia 1993.
27. Brückner H., Justus J., Kirschbaum J.: Amino Acids 21, 429 (2001).
28. Zagon J., Dehne L. I., Bögl K. W.: Nutr. Res. 14, 445 (1994).
29. D´Aniello A., D´Onofrio G., Pischetola M., D´Aniello G., Vetere A., Petrucelli L.,
Fisher G. H.: J. Biol. Chem. 268, 16949 (1993).
30. D´Aniello A., Di Fiore M., Fisher G.: Trends Compar. Biochem. Physiol. 4, 1
(1998).
31. http://www2.zf.jcu.cz/public/departments/koz/vyz/pred_08.pdf, staženo 27.11.2007.
32. Stratil P., Kubáň V.: Chem. Listy 98, 379 (2004).
33. Stratil P., Kubáň V.: Chem. Listy 99, 3 (2005).
34. http://kulturistika.ronnie.cz/c-2263-bcaa-fenomen-nebo-mytus-.html, staženo
5.11.2007.
35. Baracos V. E., Mackenzie M. L.: J. Nutr. 136, 237 (2006).
36. http://www.lekarnaexpres.cz/?page=detail&id=1097, staženo 25.2.2008.
37. http://obchod.symbinatur.com/produkt68-BCAA-500-Starlife.html#, staženo
25.2.2008.
38. Crowe M. J., Weatherson J. N., Bowden B. F.: Eur. J. Appl. Physiol. 97, 664 (2006).
39. Blomstrand E.: J. Nutr. 136, 544 (2006).
40. Newsholme E. A., Blomstrand E.: J. Nutr. 136, 274 (2006).
41. Barchas J. D., Freedman D. X.: Biochem. Pharmacol. 12, 1232 (1963).
42. Sato Y., Eriksson S., Hagenfeldt L., Wahren J.: Clin. Physiol. 1, 151 (1981).
43. Gomez-Merino D., Béquet F., Berthelot M., Riverain S., Chennaoui M., Guezennec
C. Y.: Int. J. Sports Med. 22, 317 (2001).
44. Wilson W. M., Maughan R. J.: Exp. Physiol. 77, 921 (1992).
Page 70
69
45. Struder H. K., Hollmann W., Platen P., Donike M., Gotzmann A., Weber K.: Horm.
Metab. Res. 30, 188 (1998).
46. Meeusen R., Piacentini M. F.: Adv. Exp. Med. Biol.: 527, 521 (2003).
47. Strachan A. T., Leiper J. B., Maughan R. J.: Exp. Physiol. 89, 657 (2004).
48. Varnier M.: Eur. J. Appl. Physiol. Occup. Physiol. 69, 83 (1994).
49. Wagenmakers A. J. M.: Muscle fatigue mechanisms in exercise and training
(Marconnet P., Komi P. V., Saltin B., Sejersted O. M., eds.), str. 69. Karger AG,
Basel 1992.
50. http://cs.wikipedia.org/wiki/Serotonin, staženo 3.12.2007.
51. http://www.chm.bris.ac.uk/motm/serotonin/home1.htm, staženo 3.12.2007.
52. http://cs.wikipedia.org/wiki/Fenylketonurie, staženo 21.3.2008.
53. Choudry H. A., Pan M., Karinch A. M., Souba W. W.: J. Nutr. 136, 314 (2006).
54. Freund H., Hoover H. C. Jr., Atamian S., Fischer J. E.: Ann. Surg. 190, 18 (1979).
55. Cerra F. B., Mazuski J., Teasley K., Nuwer N., Lysne J., Shronts E., Konstantinides
F.: Crit. Care Med. 11, 775 (1983).
56. Cerra F. B., Upson D., Angelico R., Wiles 3rd C., Lyons J., Faulkenbach L.,
Paysinger J.: Surgery 92, 192 (1982).
57. Jaing Z. M., Sugarbaker S., Wilmore D.: Surg. Gynecol. Obstet. 166, 115 (1988).
58. Štulík K. a kol.: Analytické separační metody. Univerzita Karlova v Praze, Praha
2004.
59. Tesařová E.: Analýza chirálních sloučenin (Tesařová E., ed.), str. 31. Pražské
analytické centrum inovací, Praha 2007.
60. Kalíková K.: Diplomová práce, PřF UK, Praha 2005.
61. Berthod A., Youbang L., Bagwill C., Armstrong D. W.: J. Chromatogr. A 731, 123
(1996).
62. Ilisz I., Ballet
S., Van Rompaey K., De Wachter R., Tourwé D., Armstrong D. W.,
Péter A.: J. Sep. Sci. 30, 1881 (2007).
63. Berthod A., Chen X. H., Kullman J. P., Armstrong D. W., Gasparrini F.,
D’Acquarica I., Villani C., Carotti A.: Anal. Chem. 72, 1767 (2000).
64. Boniglia C., Carratu B., Sanzini E.: J. Food Sci. 67, 1352 (2002).
65. Walters M. J.: J. Assoc. Off. Anal. Chem. 70, 465 (1987).
66. http://www.jascofrance.fr/pdf/hplc.pdf, staženo 15.4.2008.
67. Kafková B., Tesařová E., Suchánková J., Bosáková Z., Coufal P.: J. Sep. Sci. 26,
686 (2003).