Page 1
Kobe University Repository : Kernel
タイトルTit le
転写共役因子Peroxisome proliferator-act ivated receptor γ coact ivator-1 (PGC-1α)の新規アイソフォームの同定(Ident ificat ion of a novelisoform of peroxisome proliferator-act ivated receptor g coact ivator-1(PGC-1α))
著者Author(s) 金, 世煜
掲載誌・巻号・ページCitat ion 神戸大学医学部紀要=Medical journal of Kobe University,65(1/2/3/4):1-9
刊行日Issue date 2005-03
資源タイプResource Type Departmental Bullet in Paper / 紀要論文
版区分Resource Version publisher
権利Rights
DOI
JaLCDOI
URL http://www.lib.kobe-u.ac.jp/handle_kernel/00422382
PDF issue: 2021-05-20
Page 2
l
転写共役因子 Peroxisomeproliferator-activated receptor γ
coactivator-l (PGC-lα)の新規アイソフォームの同定
金 世建
神戸大学大学院医学系研究科応罵分子医学講座
結束病代語・消化器・腎臓内科
連絡先:金 世霊
神戸大学大学誌医学系研究科応用分子医学講窪
糖尿病代認・消化器・腎臓内科
神戸市中央区橋町 7-5-1
電話:078-382-5861
Fax: 078-382-2080
〈平成16年 5月27日受付〉
【要約】
マウスゲノム靖報の解析かち peroxisomeproliferator-
activated receptor γcoactivator-1α(PGC-1α)
の新規アイソフォームを同定した。この薪規アイソフォー
ムの mRNAのアミノ駿コード領域は,諜報の PGC-
1αm豆NAの 5'嬬の 16アミノ酸分の塩基配列が,こ
れとは異なった 12アミノ離分の配列に置換されてお
り,既報の第 lエクソンの上流に存在する新規エクソ
ンからの転写顎始によって生じるスプライシングバリ
アントと考えられた。薪農アイソフォームは骨諮筋お
よび心筋に特異的iこ強発現していた。アデノウイルス
ベクターを用いて C2C12培養筋篤縮胞に本アイソフォー
ムを強制発現させると, ミトコンドワア関連莞缶子の
発現が増加し組識学的検討でもミトコンドワアの増加
を認めた。また, リポーターアッセイにより,転写!z9
子 Nuclearrespiratory factor-1の転写活性を増強
させる{乍患を持つことが明らかとなった。 PGC-1α の
新規アイソフォームは,骨格筋のエネルギ一代謝変化
の制御に関わる可能性が示唆された。
【緒
Peroxisome proliferator-activated receptorγ
coactivator-1α(PGC-1α)は,転写冨子Peroxisome
proliferator-activated receptor γ(PPARγ 〉に結
合し,その転写活性を増強させる転写共投国子として
罰定された該蛋白である (1)0 PGC-lα は操々な転写茜
子の活性を制調することにより,多彩な生物学的作用
を発揮することが報告されているが,中でも生体のエ
ネルギ一代謝の制御における作用が注目されてい
るは, 3)。
PGC-1α は謁色脂訪組識に発現し,寒冷暴露により
その発現は強く誘導される。梅色脂肪中では, PGC-1
αはPPARγ や甲状腺ホルモン受容体の転写活性を
刺激することにより, uncoupling protein (UCP) 1を
はじめとした熱産生iこ関与する遺伝子の発現を増強さ
せる作用を有する(1)。また,肝臓では PGC-lα は絶
食により発現が誘導され,摂食によりその発現は抵下
するは〕。肝細胞で PGC-lα はHNF-4α,グルココル
チコイド受容体, Foxむなどの転写国子に結合し,
これらの転写活性を刺激することによりフォスフォエ
ノールピルどン酸カルボキシキナーゼやグルコース 8
リン酸脱リン酸化酵素といった糖薪生系遺伝子む発現
を誘導する(4一九
骨格筋の筋線維は 1型線維(遅筋)と 2型線維(速
筋)に分類される(7)0 1型隷維にはミトコンドリアが
豊富であり,エネルギー需要を酸化的リン酸化経籍iこ
より強く依存するのに対し 2聖線維はグリコーゲン
分解からのエネルギーにより強く抜存する。骨格筋で
はPGC-1α 辻 1型線維に高発現するが, PGC-lα を
筋肉に通乗j発現させたトランスジェニックマウスでは,
本来 2型線維が主鉢の筋において 1型線維を詩徴づ
ける種々の遺伝子の発現が増加し, ミトコンドワアも
増加する (8)。また骨格筋生挨む解析から 2型轄尿病
患者の骨搭筋では PGC-lα によって誘導される遺長
【キーワード]PGC-lα,スプライシングパワアント,第肉,転写共役因子
( 1 )
Page 3
2
子群の発現が低下することも報告されている (9・ヘ
すなわち, PGC-1α は骨絡筋のエネノレギ一代謝制梅に
重要な役割を果たす転写共役因子であるとともに,骨
格筋における PGC-1α む機能や発現の低下は 2型
糖尿痛む病謹と関連する可能性が示唆される。
今@J,著者はマウスゲノム?膏報の解析により筋肉及
び心筋に高発現する PGC-1α の新規アイソフォーム
を同定し,その発現特性及び機能について検言ました。
本研究により, この新規アイソフォームは骨格筋のヱ
ネルギ一代謝変化の制御に重要な機能を果たす可能性
が示唆された。
【方 法】
蛋自発現ベクター
マウス PGC-1α の新規アイソフォーム (PGC-1αb)
の cDNAは, PGC-1α1b特異領域を含むセンスプラ
イマー (5'-tga gtg aca tgg atg ttg gga ttg t-3')
及び既報のマウス PGC-1α のカルボキシル末端の匡
子山こ本自民なアンチセンスプライマー (5'-caa agc tga
cac ccg tga at-3')を用いて,マウス膏搭筋 cDNA
ライブラリー〈タカラバイオ〉を鋳主として PCRに
て増幅した。既報の PGC-1α(GC-1αa) 0) cDNA
も同じライブラワーより PCRにて増幅した。これら
の cDNAを蛋自発現ベクタ-pcDNA3.1 (インピト
ロジェン)!こサブクローニングし,それぞれのベクター
を pcDNAjPGC-1αb,pcDNAjPGC-1αaと名づけ
た。 PGC-1αbをコードするアデノウイルスベクター
は TakaraAdenovirus Expressionキット〔タカラ
バイオ)を用いて,既報の手技(叫に従い作成し, Ax
CAPGC-1αbと名づけた。 βガラクトシダーゼ遺伝
子をコードするアデノウイルスベクタ-AxCALacZ
辻東京大学童科学研究所,斉藤泉薄士より供与を受け
た。ヒト Nuclearrespiratory factor-1 (NRF-l)
cDNAはPCR法によりヒト骨格筋 cDNA(タカラバ
イオ)を鋳型として増幅し. pcDNA3.1にサブクロー
ニングした。このベクターは pcDNAjNRF-1と名づ
けた。また,ヒトmitochondrialtranscription factor
A (mTFA)遺伝子の NRF-1反応性領域詰既報と同
隷にゆヒトゲノム DNA(タカラバイオ社)を鋳型と
して PCRIこて増幅し,ホタルルシフエラーゼ遺伝子
をコードするベクタ-pGL3 (インピトロジェン)に
サブクローニンクーした O 本プラスミドは mTFA-
1ucjpGL3と名づけた。
リポーターアッセイ
lμgの mTFA-1ucjpGL3,1μzの pcDNAjPGC-1
く2)
αaもしくは pcDNAjPGC-1αb,1μgの pcDNAj
NRF-1, 0.1μgの・ガラクトシダーゼ遺伝子をコー
ドする SRα(SRαjLacZ)の各プラスミドを混合後,
LipofectAMINE (インビトロジェン)を用いて, 24
ウェルプレートで培養した 293細抱iこ導入した。 pcD
NAjPGC-1αaもしくは pcDNAjPGC-1αb及び、
pcDNAjNRF-1の導入を行わない場合は,遺伝子を
コードしない pcDNA.3.1を方日え, トランスフェクショ
ンに用いるプラスミド量は常に 3.1μgとなるよう謁
整した。 48時間後に組抱を呂収し,可解化分画のル
シフエラーゼ活性及び・ガ、ラクトシダーゼ活性をホタ
ルルシフエラーゼアッセイシステム及びβガラクトシ
ダーゼアッセイシステム〈プロメガ〉により測定し,
βガラクトシダーゼ活性あたりのルシフエラーゼ活性
により N豆F-1の転写活性を算出した。
C2C12筋詩紹抱への PGC-lαbの過剰発現
C2C12筋芽縮胞は既報のごとく培養し,筋誇紹抱
への分化誘導を行った(ヘ C2C12筋筒細胞に以前に
記載した方法に従って(ωAxCAPGC-lαbもしくは対
照として AxCALacZを感染させ,感染 48時間後に
細抱を回収し,細胞から totalRNAを抽出し,
RT-PCR Iこ供した。また,@J収した細胞を電子顕微
鏡による解析に供した。
マウス各臓器の PGC-lα各アイソフォームの発現
5匹む 10適齢雄性 ICRマウスの各臓器を話時摂金
下で採取し totalRNAを揺出した。各語体か0t辱た
total RNAを等量づ、つ還合した後,定量的 RT-PCR
によち, PGC-lα各アイソフォームの発現量を解析し
fこ。
定量的 RT-PCR
細抱,マウス各騒器より抽出した約 3μgの total
RNAを鋳型として逆転写を行い, cDNAを毎た。こ
の cDNAを鋳型として,以前iこ記載した手技に従
い<t4)SYBIミGreenPCR Masterキット(ノfーキンエ
ルマー), Sequence Detector Mode17900 (PEアプ
ライドセイエンス)を用いて定量的 PCR解析を行い,
36B4遺伝子に対する相対発現量として各撞遺伝子の
発現量を定量した。遺伝子発現の検出に用いたプライ
マーベアは以下の通りである。 PGC-1αb,5' -tga
gtg aca tgg atg坑ggga ttg t-3' (センス)及び
5' -tcg cag gct cat tgt tgt act ggt-3' (アンチセ
ンス); ATP合成酵素 βサブユニット (subunitof
A TP synthetase: b A TPase), 5' -gag acc ttg ggc
aga atc atg aat gtc a-3' (センス〉及び、 5' -ggg
Page 4
cca gca gat cca caa cct tta tc-3' (アンチセンス);
チトクロームc喜変化欝素サブユニット II(cytochrome c
oxidase 8ubunit宜:COX II), 5' -gcc gac taa atc
aag caa cag taa-3' (センス)及び 5' -tct agg aca
atg ggc ata aag cta t-3' (アンチセンス); mtTF
A, 5' -agt tcc cac gct gg七agtgt-3' (センス〉及
び 5' -gcg cac atc tcg acc c-3' (アンチセンス);
NRF-1, 5' -cag caa ccc tga tgg cac cgt gtc g-3'
(センス)及び、 5' -ggc ctc tga tgc ttg cgt cgt ctg
g-3' (アンチセンス)0PGC-1αa及び 36B4mRNA
の検出には既報のプライマーペアを用いた(14slao
【結 果】
PGC-lα新規アイソフォームの同定
マウス ESTデータベースを竣索の結果,窺報む P
GC-1αcDNAと5'剖のみが異なった塩基配列を有す
るESTクローンが存在することを見出した (ESTア
クセッション番号 AW107131及び BB853729)。これ
らの ESTクローンの塩基配列から推定されるアミノ
酸記列は,既報の PGC-1α のアミノ末端 16アミノ按
分の塩基が欠失し, これとは異なる 12アミノ酸分の
塩基に量換された講造を持っていた(図 1)。マウス
ゲノム清報データベースを検索した結果, これら
ESTクローンにおいて既報の PGC-1αcDNA塩基配
列とは異なる部生と完全に一致する配列が,既報の P
GC-1α遺{云子の第一エクソン上流約 15kbの位墨に存
在した。この梧同性領域の 3'末端にはスプライシン
グ受容部のコンセンサス配列に合致する塩基配列が存
在した。以上の知克から,今回同定した ESTクロー
ンがコードする mRNAは, PGC-1α遺伝子の未開定
のスプライシングパワアントに由来すると考えられた。
以下,本論文では,既報の PGC-1α を PGC-1αaと
今回同定したスプライシングバリアントを PGC-1αb
と呼ぶ。
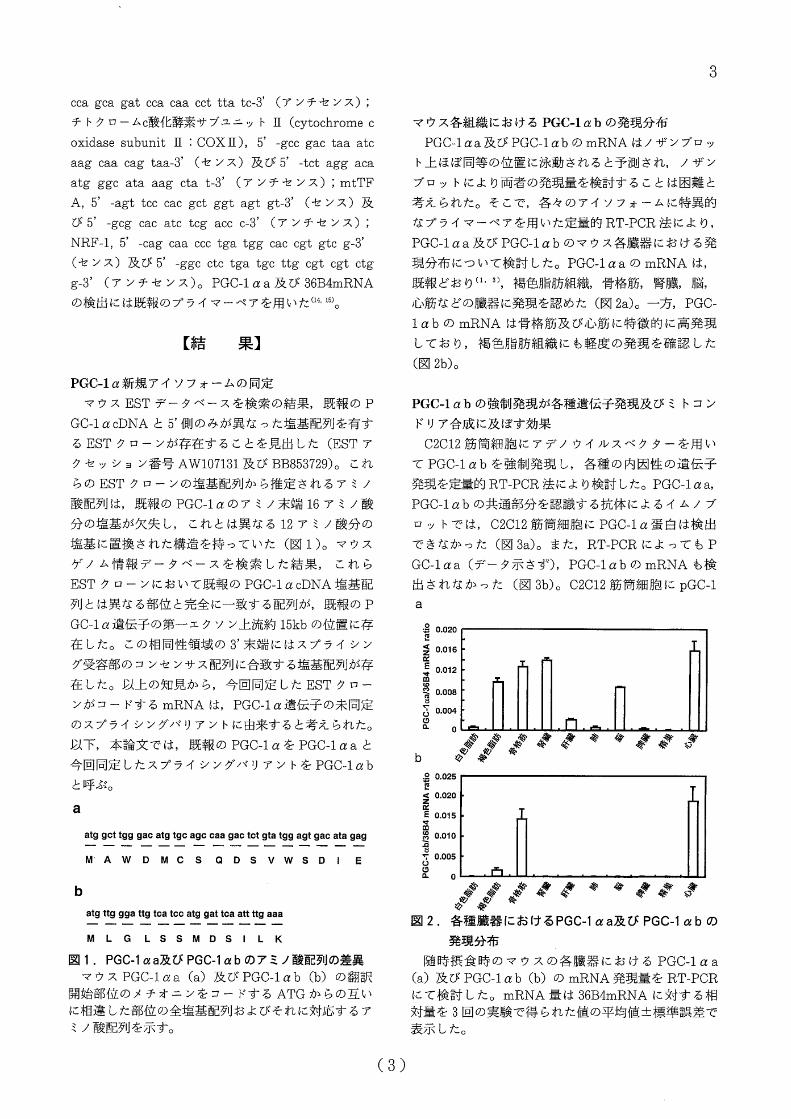
a
atg gct tgg gac atg tgc agc caa gac tct gta tgg agt gac ata gag
MAWDMCS QDSVWSDI E
b
atg ttg gga ttg tca tcc atg gat tca att ttg aaa
MLG L SSMDSILK
匿 1. PGC・1αa及び PGC-lαbのアミノ接配列の差異
マウス PGC-1αa(a)及び PGC-1αb(b)の翻訳
関桧部位のメチオニンをコードする ATGからの互い
に相違した部位む全塩基配列およびそれに対応するア
ミノ酸配列を示す。
(3)
3
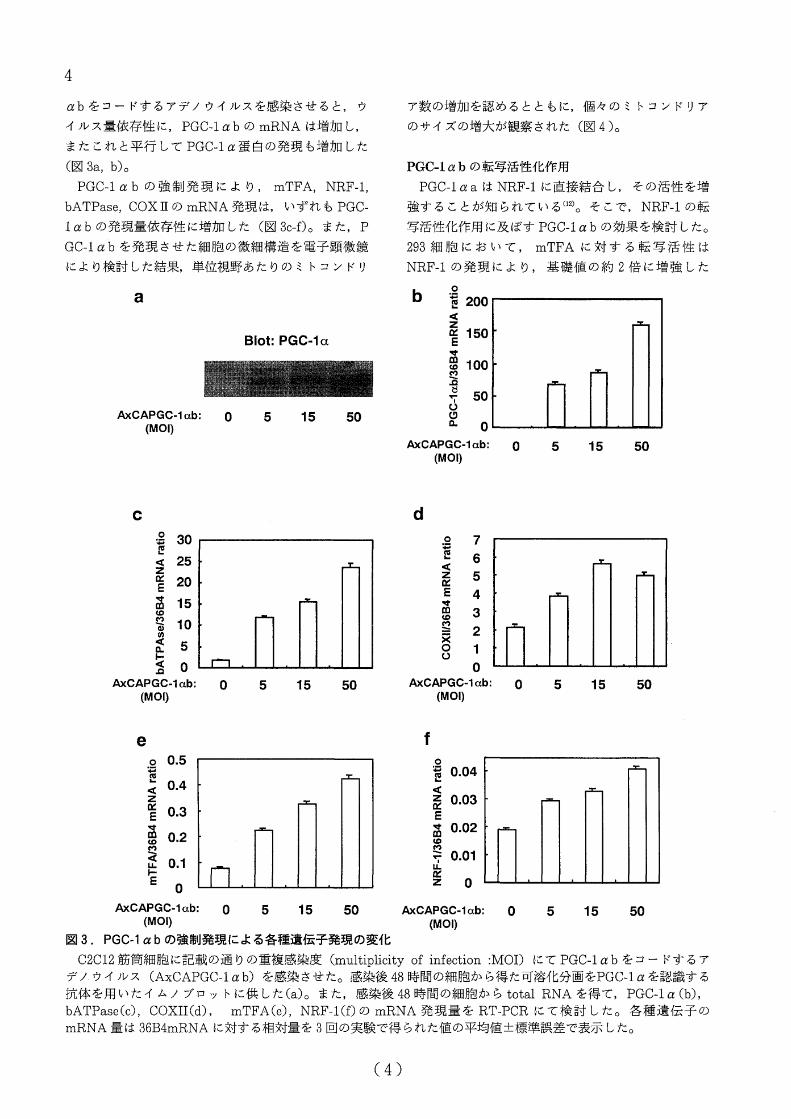
マウス各組織におぜる PGC-lαbの発現分布
PGC-1αa及び PGC-1αbのmRNAはノザンプロッ
ト上ほぼ同等の位置に泳動されると予瀕され,ノザ、ン
プロットにより両者の発現量を験討することは困難と
考えられた。そこで,各々のアイソフォームに特異的
なプライマーベアを患いた定量的 RT-PCR法により,
PGC-1αa及び PGC-1αbのマウス各臓器における発
現分布について検討した。 PGC-1αaのmRNAは,
既報どおり(1.3) 掲色詰訪組識,骨格筋,腎臓,脳,
心篇などの臓器に発現を認めた(璽 2a)。一方, PGC-
lαbの mRNAIま骨格筋及び心筋iこ特徴的に高発現
しており,褐急詣訪組織にも軽震の発環を確認した
(国2b)。
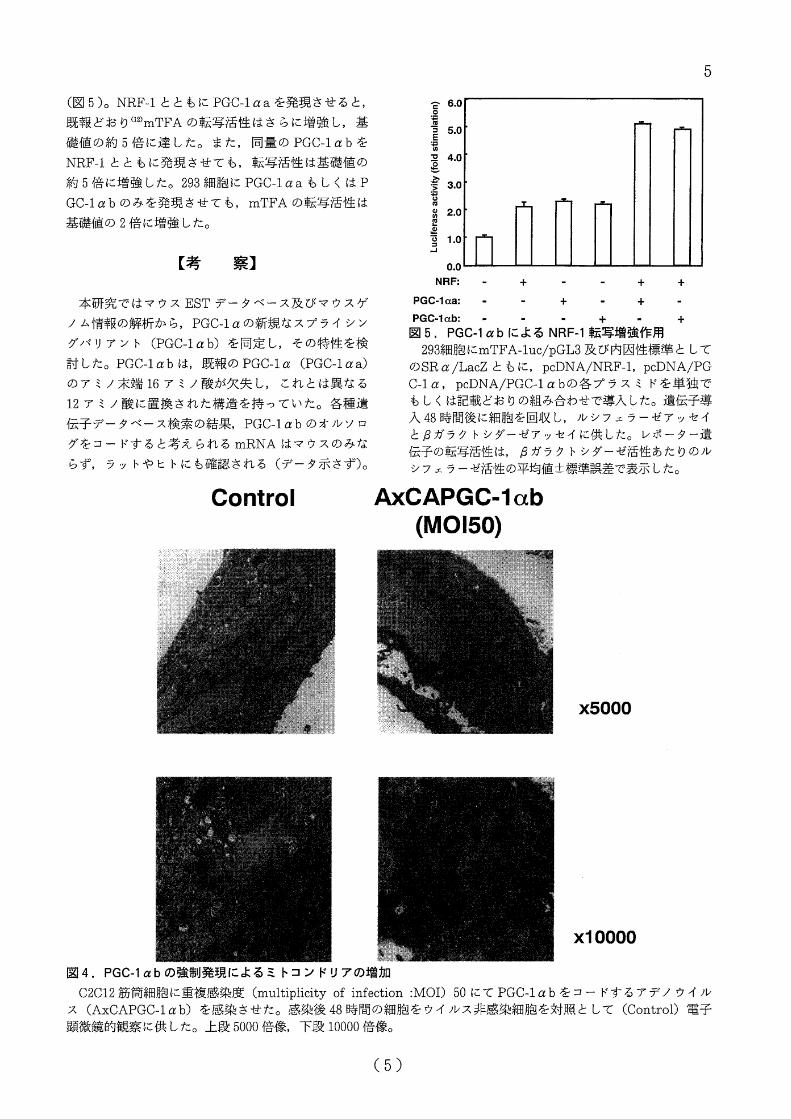
PGC-lαbの強制発現が各謹遺{云子発現及びミトコン
ドリア合成に及ぼす効果
C2C12筋笥紐抱にアデノウイノレスベクターを罵い
て PGC-1αbを強制発現し,各撞の内因性の遺伝子
発現を定量的 RT-PCR法により検討した。 PGC-1αa,
PGC-1αbの共通部分を認識する抗体によるイムノプ
ロットでは, C2C12箆篤鱈胞に PGC-1α蛋白は検出
できなかった(図 3a)。また, RT-PCRによっても P
GC-1αa (データ示さず), PGC-1αbのmRNAも検
出されなかった(密 3b)0C2C12筋笥組胞に pGC-1
a
b
,g 0.020 ",
‘司芸0.016D::
jom 毛D
39GOB
U 0.004 0 a.
,2 0.025
2 2962B
E 0.015
z ~ 0.010 :o で 0.005u 。a. 0
命令命令命令.. )参考 98 タ点々々 々々o--ゼ「
密 2. 各種臓器におけるPGC・1αa及び PGC-lαbの
発現分布
槌時摂食時のマウスの各議器における PGC-1αa
(a)及び PGC-lαb(b)の mRNA発現量を RT-PCR
にて検討した。 mRNA量は 36B4mRNAに対する相
対量を 3@]の実験で得ちれた植の平均値土標準誤差で
表示した。
Page 5
ア数の増加を認めるとともに,掴々のミトコンドリア
のサイズの増大が観察された〈図 4)。
PGC-lαbO)転写活性化作用
PGC-lαaはNRF-llこ亘接結合し,その活性を増
強することが知られている(12)。そこで, NRF-IO)転
写活性化作用に及ぼす PGC-lαbの効果を検討したO
293細抱において, mTFAに対する転写活性は
NRF-lの発現iこより,基礎誼の約 2倍に増強した
αbをコードするアデノウイルスを感染させると,
イノレス量依存性iこ, PGC-lαb 0) mRNAは増加し,
またこれと平行して PGC-lα蛋白の発現も増加した
〈図 3a,b)。
PGC-lαbの強制発現によち, mTFA, NRF-l,
bA TPase, COX IIのmRNA発現は,いずれも PGC-
1αbの発現量抜毒性に増加した〈図 3c-f)。また, P
GC-lαbを発現させた細胞の徴細構造を電子顕微鏡
iこより換討した結果,単位視野あたりのミトコンドワ
ウ
4
n
u
n
u
n
u
《U
民
叫
《
》
司
4
4
Z
噌E
。=Ed『ZNhE-玄出品甲山口a6FEOO&
50
b
Blot: PGC.1α
a
50 15 5 G
o AxCAPGC圃 1αb:
(MOI)
50 15 5 G AxCAPGC・1αb:(話Ol}
d
7
6
5
4
3
2
1
8 AxCAPGC-1αb:
(MOI)
。一割弱』〈ZNhEマ図@品川『一宗一00
c 。場..
<<l 』
《z 0:: E マ∞ u) F事
@
自陣
《c.. ト・
~ 0 AxCAPGC・1αb:
(MOI)
30
25
20
15
10
5
50 15 5 G 50 15 5 9
aHTqu
司4
4
z
nunununuw
n
u
n
u
n
U
A
U
。-H語《ZHh呈・マ包@目立で
Hhgz
0.5
0.4
0.1
0.3
0.2
。ぉ酬と〈
ZU5守剛志円ミHhaFE
e
e AxCAPGC・1αb:
(MOI)
璽 3. PGCぺαbの強制発現による各蓮遺缶子発現の変化
C2C12筋筒細抱に記載の通りの重複感染度 (multiplicityof infection :MOI) Iこて PGC-lαbをコードするア
デノウイルス (AxCAPGC-lα訟を感染させた。感染後48時間の紐抱から得た可溶化分画をPGC-lαを認識する
抗体を用いたイムノプロットに供した(心。また,感染後 48時間の細抱かる totalRNAを得て, PGC-lα(b),
bATPase(c), COXII(心, 血 TFA (e), NRF -1(f)の mRNA発現量を RT-PCRにて検討した。各種遺伝子の
mRNA量iま36B4mRNAに対する相対量を 3呂の実験で得られた値の平均鐘士課準誤差で表示した。
(4)
50 15 5 。AxCAPGC・1αb:
(MOI) 50 15 5 Q
8
Page 6
(図 5)0NRF-1とともに PGC-1αaを発現させると,
既報どおり(12)mTFAの転写活性はさらに増強し,基
礎僅の約 5fきに達した。また,同量の PGC-1αbを
NRF-1とともに発現させても,転写活性は基礎僅の
約 5倍に増強した。 293組抱iこPGC-1αaもしくは P
GC-1αbのみを発現させても, mTFAの転写活性は
基礎櫨の 2倍に増強した。
【考 察】
本研究ではマウス ESTデータベース及びマウスゲ
ノム清報の解析から, PGC-1α の新規なスプライシン
グバリアント (PGC-1α訟を同定し,その特性を検
討した。 PGC-1αbは,既報の PGC-1α(PGC-1αa)
のアミノ末端 16アミノ酸が欠失し,これとは異なる
12アミノ駿iこ置換された構造を持っていた。各種遺
伝子データベース検索の結果, PGC-1αbOオルソロ
グをコードすると考えられる mRNAはマウスのみな
らず, ラットやヒトにも確認される(データ示さず)。
Control
言 6.00 .... 国
~ 5.0 :;:; 韓
三 4.00
匂阻
害3.0z 。惇
m 2.0 暗... 世
匂回
三1.0J
0.0
NRF:
PGC・1αa:
+ +
÷ +
PGC.1αb: ・・・+・図 5.PGC・1αbによる NRF-l転写増強作用
5
+
÷
293細胞にmTFA-1ucjpGL3及び内密性諜準として
のSRαjLacZともに, pcDNAjNRF-l, pcDNAjPG
C-1α , pcDNAjPGC-1αbの各プラスミドを単独で
もしくは記載どおりの組み合わせで導入した。遺伝子導
入 48時間後に細胞を冨寂し,ルシフエラーゼアッセイ
とβガラクトシダーゼアッセイに供した。レポーター遺
缶子む転写活性は, βガラクトシダーゼ活性あたりのル
シフエラーゼ活性む平均鐘±諜準誤差で表示した。
AxCAPGC-1αb (MOI50)
x5000
x10000
密 4. PGC・1αbの強制発現によるミトコンドワアの増加
C2C12筋筒細屈に重複惑染震 (multiplici匂T of infection :MOI) 50にて PGC-1αbをコードするアデノウイル
ス (AxCAPGC-lαb)を感染させた。惑染後48時間の椙抱をウイルス非感染細胞を対照として (Control)電子
顕徴鏡的観察に供した。上段 5000倍畿,下設 10000f音イ象。
(5)
Page 7
6
これ辻, PGC-lαbがマウスのみならず,時乳動物に
普遍的;こ存在する PGC-lα遺{云子のスプライシング
バリアントである可能性を示唆するものであるO
PGC-lαaと辻異なり, PGC-lαbの臓器分布は,
ほぼ骨諮筋及び心筋に限局していた。 PGC-lαaは,
骨格筋では呼吸鎖における接化的リン酸化に関わる遺
伝子の誘導に関与するとともに, ミトコンドリアの生
成にも重要な磯能を果たすと考えられているω。そこ
で,本萌究では,士吾養筋笥縮抱に PGC-lαbを過剰
発現し,各種の内菌性遺伝子の発現を検討した。
C2C12詰養篇簡細胞にアデノウイルスベクターを用
いて PGC-lαbを発現させると,酸化的リン酸化に
必須の欝素である bATPase及び COX豆 0)mRNA
が PGC-lαb蛋自の発現量の増加に伴って強く誘導
された。 bATPaseむ遺託子は核内ゲノム上に存在す
るのに対しく16),COX宜遺伝子はミトコンドワア遺伝
子にコードされる的。 mTFAは核内ゲノムにコード
される転写因子であるが, ミトコンドりア iこ移行して
ミトコンドワアむ各謹の遺伝子の発現誘導やミトコン
ドリア遺伝子の謹製iこ関与することが知られている(へ
また,転写冨子 NRF-lは各種の酸化的ワン喜変化iこ関
わる酵素の遺伝子を誘導するとともに m工nTFAむ遺
伝子発現も活性化する cl路へ§
り弘, m工nTF、A及びび、 NRF-lの発現lはま毘量依存性に増加し,
さら iにこミトコンドリアの生成も誘導し7たこ。以上の結果
よち, PGC-lαb は, PGC-lαaと同様に,箭肉にお
ける酸化的リン酸化に関わる遺伝子の誘導能を有する
とともに, NRF-ljmTFAカスケードを介してミトコ
ンドリアの生戒を増加させることが明ちかとなった。
293細胞を用いた一通性発現系により, PGC-lαa
とPGC雌 1αbむ転写共役活笹を検討したところ, PG
C-lαaと同様に PGC-lαbもNRF-lの転写活性を刺
激した。 PGC-lαa,PGC-lαbともに NRF-l非導入
細胞においても, mTFA 0)プロモーター活性を基礎
鐘に比して約 2倍に増加させたが,これは 293縮抱に
存在する内菌性0)NRF-lの活性を刺激した結果であ
ると考えらえる。導入したプラスミド量あたちの
NRF-l tこ対する転写共役活J詮iこは, PGC問 1αaとPG
C-lαb寵で差は認めなかった。すなわち, PGC-lαa
とPGC-lαbは,少なくとも NRF-l1こ対しては,ほ
ぼ同等の活性を有するものと考えられた。
PGC-lαa及び PGC-lαbが機能的iこ同等であると
仮定すると,これらのアイソフォームのうちいずれが
生理的に重要かは,南アイソフォームの発現量の多寡
に依存すると考えられる o RT田 PCR法では両者の発
現量の絶対鐘を比較することは困難であり,骨諮筋に
おいていずれむアイソフォームが, ミトコンドリアの
(6)
生成や酸化的リン酸化に関わる各種の遺伝子の発現に
対し主要な役割を果たすかは明らかではなし可。アイソ
フォーム特異的な抗体を作成し,蛋白レベルで両アイ
ソフォームの発現量C絶対値を比較することは, PG
C-l αbむ生理的意義のさらなる検討にとって重要で
あろう。また,各アイソフォームを特異的に欠失する
遺伝子欠損マウスを作成すれば,橿体レベルでのエネ
ルギ一代謝における生理的機能む差異が暁らかになる
と考えられる。また,運動急需により骨格筋の酸化的
リン酸化能やインスワン抜葎性む糖取り込み能は増強
する (20,2九また,運動負需によって骨権第0)PGC-l
αの発現量が増加すること (22,23)も知られており,運動
負荷による骨格筋の糖指費代諜の変化には, PGC-lα
の発現増強が重要な投書日を果たす可能性がある O 運動
負荷において PGC-lα のいずれのアイソフォームが
増加するのかという点も,今後む重要な検討課題と考
えられる。
最近, DNAマイクロアレイを黒いた解析により,
2聖糖尿病患者の骨格窮では PGC-lα及び、 PGC-lα
いよって誘導される遺缶子群の発現が低下していると
り報告がなされたけ, 1九また 2型糖尿病患者の近親
者では,耐糖能が正常であっても,骨格筋におけるミ
トコンドリア機能が低下していることも報告されてい
る位九すなわち骨格筋における PGC-lα の発現の抵
下や,それによって生じるミトコンドリア機能,すな
わち酸化的ワン酸化反症の抵下は 2型糖尿病の擢患
感受性を決定付ぜる遺伝的要菌の一つである可能性が
示唆される o DNAマイクロアレイを用いた解析では
PGC-lαbとPGC-lαaをIRl.IlJできず 2型糖尿病患
者でいずれのアイソフォームの低下が生じているか辻
明らかではない。 PGC-lαb,PGC-lαaの各アイソ
フォームの発現量を 2型糖尿病患者の骨格筋で検討す
ることにより 2型糖京病の遺伝素因の解明にとって
有用な情報が得与れる可能性があろう。
【謝辞】
本研究にあたり, AxCALacZを撰与いただきまし
た斉藤泉博士に深く惑謝いたします。また,ご指導を
賜りました神戸大学大学説志期分子医学講座,糧京病
代露首・消化器・腎E車内科春日雅人教授に厚くお礼申し
上げます。
【文献】
1) Puigserver, P., Wu, Z., Park, C.W., Graves,
R., Wright, M., Spiegelman, B.M.: A cold-
Page 8
inducible coactivator of nuclear receptors
linked to adaptive thermogenesis. Cell 党:
829-839, 1998.
2) Knutti, D., Kralli, A.:PGC-1, a versatile
coactivator. Trend. Endocrinol. Metab. 12:
360-365, 2001.
3) Puigserver, P., Spiegelman, B.M.: Peroxisome
proliferator-activated receptor-gamma coactivator
lα(PGC-1α): transcriptional coactivator and
metabolic regulaもor.Endocr. Rev. 24: 78-90,
2003.
4) Yoon, J.C., Puigserver, P勺 Chen,G., Donovan,
J., Wu, Z., Rhee, J., Adelmant, G., Stafford,
J., Kahn, C.R., Granner, D.K., Newgard, C.
B., Spiegelman, B.M.: Control of hepatic
gl uconeogenesisもhroughthe transcriptional
coactivator PGC-1. Nature. 413: 131-138, 2001.
5) Herzig, S., Long, F., Jhala, U.S., Hedrick, S.,
Quinn, R., Bauer, A., Rudolph, D., Schutz,
G., Yoon, C., Puigserver, P., Spiegelman, B.,
Montminy, 主主 CREB regulates hepatic
gluconeogenesis through the coactivator
PGC-1. Nature. 413: 179-183, 2001.
6) Puigserver, P., Rhee, J., Donovan, J., Walkey,
C.J., Yoon, J.C., Oriente, F., Kitamura, Y.,
Altomonte, J., Dong, H., Accili, D., Spiegelman,
B.拡 Insulin-regulatedhepatic gluconeogenesis
through FOX01-PGC-1αinteraction. Nature.
423: 550-555, 2003.
7) Booth, F.W. & Thomason, D.B.: Molecular
and cellular adaptation of muscle in response
to exercise: perspectives of various models.
Physiol. Rev. 71: 541-585, 1991.
8) Lin, J.,羽Tu,五円 Tarr,P.T., Zhang, C.Y.,
Wu, Z., Boss, 0., Michael, L.G., Puigserver, P.,
Isotani, E., Olson, E.N., Lowell, B.B., Bassel-
Duby,豆., Spiegelman, B.M.: Transcriptional
co-activator PGC-1 alpha drives the formation
of slow-twitch muscle fibres. Nature. 418:
797-801. 2002.
9) 1¥在ootha,V.K., Lindgen, C.M., Eriksson, K.F.,
Subramanian, A., Sihag, S., Lehar, J., Puigserver,
P., Carlsson, E., Ridderstrale, M., Laurila,
E., Houstis, N., Daly, M.J., Patterson, N.,
Mesirov, J.P., Golub, T.R., Tamayo, P.,
Spiegelman, B., Lander, E.S., Hirschhorn,
J.N., Altshuler, D., Groop, L.C.: PGC-1a1pha-
(7)
7
responsive genes involved in oxidative
phosphorylation are coordinately downregulated
in human diabetes. Nat Genet. 34: 267-273,
2003.
10) Patti, M.E., Butte, A.J., Crunkhorn, S., Cusi,
K., Berria, R., Kashyap, S., 主主iyazaki,Y.,
Kohane, 1., Costello, M., Saccone, R., Landaker,
E.J., Goldfine, A.B., 五在un,E., DeFronzo, R.,
Finlayson, J., Kahn, C.R., Mandarino, L.J.:
Coordinated reduction of genes of oxidative
metabolism in humans with insulin resistance
and diabetes: Potential role of PGC1 and
NRF1. Proc Natl Acad Sci USA. 100: 8466-
8471, 2003.
11) Sakaue, H., Ogawa, W., Takata, M., Kuroda,
S., Kotani, K., Matsumoto, M., Sakaue,
hι, Nishio, S., Ueno,五 Kasuga, 1¥ι:
Phosphoinositide 3-kinase is required for
insulin-induced but not for growth hormone-
or hyperosmolarity-induced glucose uptake
in 3T3-L1 adipocytes. Mol Endocrinol. 11:
1552-1562, 1997.
12)羽Tu,Z., Puigserver, P., Andersson, U., Zhang,
C., Adelmant, G., Mootha, V., Troy,
A., Cinti, S., Lowell, B., Scarpulla, R.C.,
Spiegelman, B.M.: Mechanismas Controlling
E在itochondrial Biogenesis and Respiration
through the Thermogenic Coactivator PGC同1.
CelL 98: 115-124, 1999.
13) Takata, M., Ogawa, W., Kitamura, T., Hino,
Y., Kuroda, S., Kotani, K., Klip, A.,
Gingras, A.C., Sonenberg, N., Kasuga,話J
Requiremnet for Akt (protein kinase B) in
insulin-induced activation of glycogen synthase
and phosphorylation of 4E-BP1 (PHAS-l). J
Biol Chem. 274: 20611-20618, 1999.
14)話iyake,1ζ, Ogawa, W., Matsumoto, M.,
Nal王amura,T., Sakaue,五., Kasuga, M.:
Hyperinsulinemia, glucose intolerance, and
dyslipidemia induced by acute inhibition of
phosphoinositide 3-kinase signaling in the
liver. J. Clin. Invest. 110: 1483-1491, 2002.
15) Inoue, H., Ogawa, W., Ozaki, M., Haga, S.,
Matsumoto, M., Firukawa, K., Hashimoto,
N., Kido, Y., Mori, T., Sakaue,五.,
Teshigawara, K., Jin, S., Iguchi, H.,
Hiramatsu,豆., LeRoith, D., Takeda, K.,
Page 9
8
Akira, S., Kasuga, M.: R01e of ST A T -3 in
regu1ation of hepatic gluconeogenic genes
and carbohydrate metabolism in vivo. Nat
Med. 10: 168-174, 2004.
16) Das, A.M.:豆egulationof the mitochondrial
ATP-synthase in health and disease. 五在01
Genet Metab. 79: 71-82, 2003.
17) Gnaiger, E., Lassnig, B., Kuznetsov, A.,
Rieger, G., Margreiter, R.: Mitochondrial
oxygen affinity, respiratory flux control
and excess capacity of cytochrome c oxidase.
J Exp Biol. 201: 1129-1139, 1998.
18) Shadel, G.S., Clayton, D.A.: Mitochondrial
transcription initiation. Variation and conserva-
tion. J. Biol. Chem. 268: 16083-16086, 1993.
19) Scarpulla, R.C.: Nuclear control of
respiratory chain expression in mammalian
cells. J. Bioenerg. Biomembr. 29: 109-119,
1997.
20) Holloszy, J.O., Coyle, E.F.: Adaptations of
skeletal muscle to endurance exercIse and
their metabolic consequences. J Appl Physiol.
56: 831-838, 1984.
21) Galbo, H.: Influence of aging and exercise
on endocrine function. Int J Sport Nutr
Exerc Metab. 11 Suppl: S49-57, 2001.
22) Baar, K., Wende, A.R., Jones, T.E., Marison,
M., Nolte, L.A., Chen, M., Kelly, D.P.,
Holloszy, J.O.: Adaptations of skeletal
muscle to exercise: rapid increase in the
transcriptional coactivator PGC-1. FASEB J.
16: 1879-1886, 2002.
23) Russell, A.P., Feilchenfeldt, J., Schreiber, S.,
Praz, M., Crettenand, A., Gobelet, C., Meier,
C.A., Bell, D.R., Kralli, A., Giacobino, J.P.,
Deriaz, 0.: Endurance training in humans
leads to fiber type-specific increases in levels
of peroxisome proliferator-activated receptor-
ga盟国acoactivator-1 and peroxisome proliferator-
activated receptor-alpha in s註eletalmuscle.
Diabetes. 52: 2874-2881, 2003.
24) Petersen, K.F., Dufour, S., Befroy, D.,
Garcia, R., Shulman, G.1.: Impaired
mi七ocondrialactivity in the insulin-resistant
offspring of patients with type 2 diabetes.
N. Engl. J. 乱在ed.350: 664-671, 2004.
(8)
Page 10
2
Identification of a novel isoform of peroxisome proliferator-activated receptor g coactivator-1 (PGC・-1a)
Shiyu Jin
Division of Diabetes and Digestive and Kidney Diseases, Department of Clinical Molecular Medicine,
Kobe University Graduate School of Medicine,豆obe650-0017, Japan
We have identified a novel splicing variant of peroxisome proliferator-activated receptor γ
coactivator-l (PGC-lα), derived from a previously unidentified exon of the PGC-lαgene. This
isoform was abundantly expressed in skeletal muscle and heart. Ectopic expression of the isofrom of
the PGC-lαinto C2C12 myotubes with the use of adenovirus-mediated gene transfer increased the
mRNA levels of genes involved in mitochondrial respiratory chain and stimulated biogenesis of
mitochondria. Reporter gene assays revealed that the isofrom stimulated transactivation activity of
nuclear respiratory factor-1. These results suggest that the novel isoform of PGC-lαplays an
important role in energy homeostasis in skeletal muscle.
(9)