43
Transport a sekrece bakterií a kvasinek Proteiny prokaryotních mikroorganismů určené k transportu obsahují na N- terminálním konci sekvence nazývané signální peptidy.

Transport a sekrece bakterií a
kvasinek
Proteiny prokaryotních mikroorganismů určené k transportu obsahují na N-
terminálním konci sekvence nazývané signální peptidy.
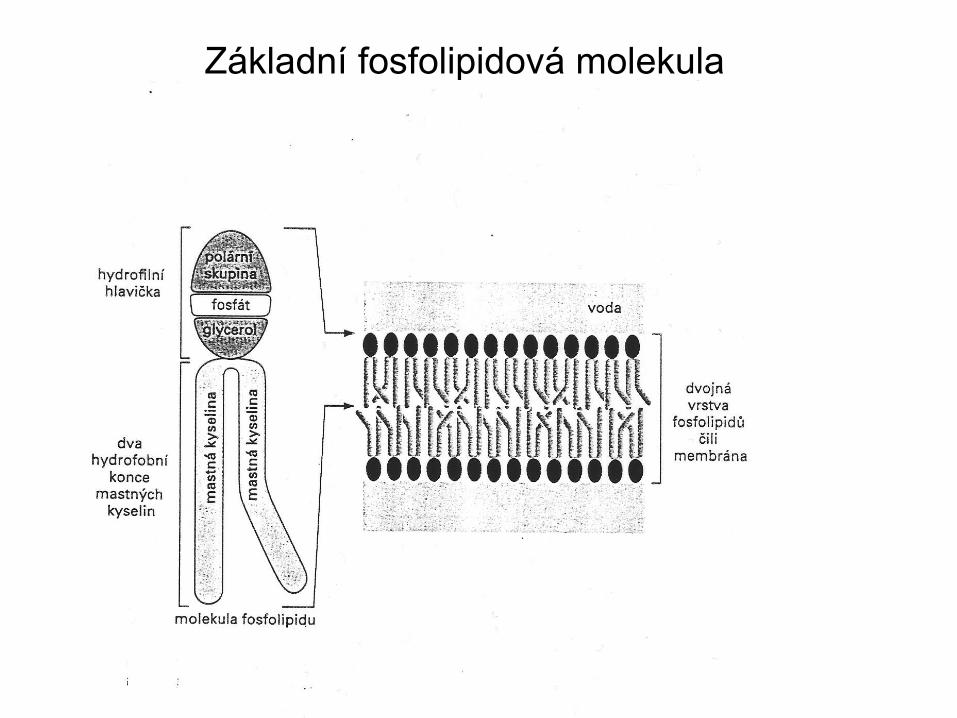
Základní fosfolipidová molekula
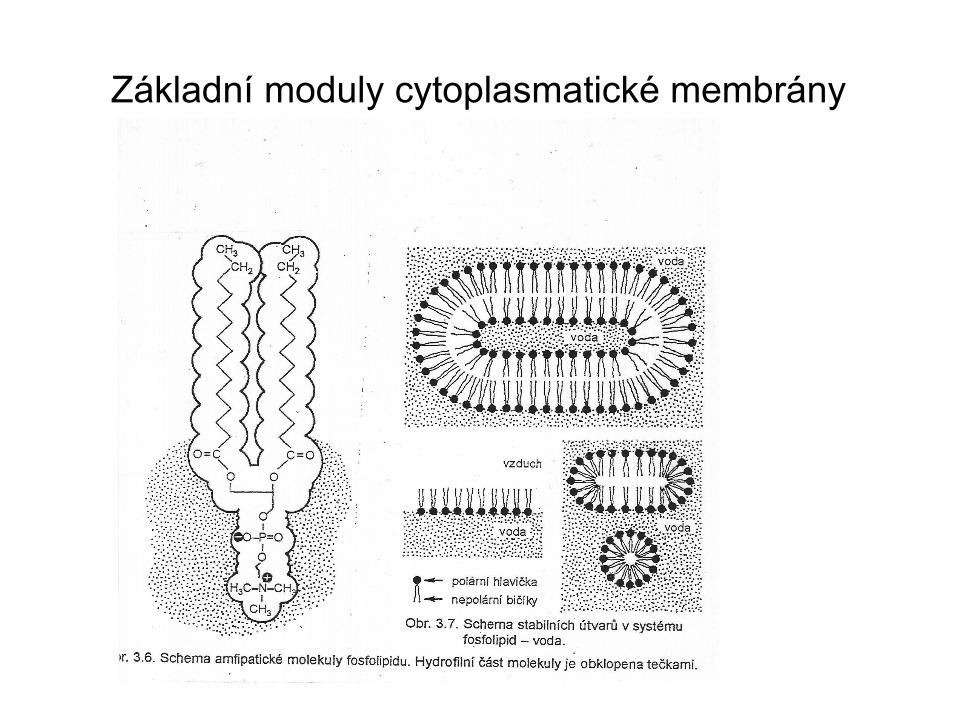
Základní moduly cytoplasmatické membrány
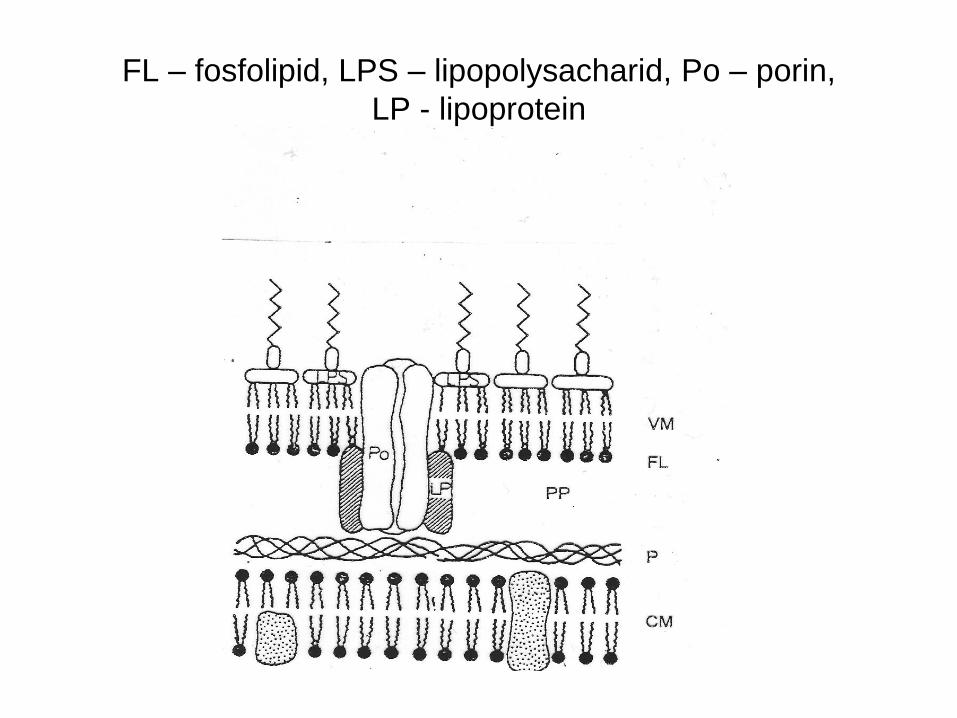
FL – fosfolipid, LPS – lipopolysacharid, Po – porin,
LP - lipoprotein

Typy porinů

Schema sekrečního kanálu

Sekrece proteinů
• Syntéza
• Zanoření do membrány
• Translokace
• Rozpoznání signálního peptidu
• Odštěpení signálního peptidu speciální peptidasou
• Konečné umístění zralého peptidu
• Signální sekvence : 18 – 40 AK
• Počátek: 5 - 6 AK , pozitivní náboj
• Střední část: průměrně 12 AK, hydrofobní bez náboje, α-helix
• Koncová část:C-terminální, 6 AK, neutrální AK v pozici -1 a -3,
často prolin nebo glycin

Signální peptidy - funkce

Proteiny vnější membrány (VM) G- bakterií
• Protein funkce • OmpA stabilizace VM a konjugace F, receptor Tu II fága
• Murein lipoprotein nejhojnějšní protein u E.coli, S,typhimurium
• OmpB (porin) difuzní kanál pro metabolity včetně maltosy
• LamB (maltoporin) specifický porin pro maltosu, maltodextrin,
receptor bakterofága λ
OmpC (porin) difuzní kanál malých molekul, receptor fága TuIb
a T4
OmpF (porin) difuzní kanál malých molekul, receptor fága TuIa
a T2
OmpT proteasa
PhoE (protein E) anion-selektivní difuzní kanál indukovaný
limitací fosfátu
Protein P anion-selektivní difuzní kanál Ps. aeruginosa,
TolA udržení integrity VM, skupina A kolicinů
TonA vstup sideroforu (Fe), receptor T1, T5, 80, kolicin M
TonB vstup sideroforu (Fe), transport B12
Tsx kanál pro nukleosidy, receptor sudých fágů, kolicin K
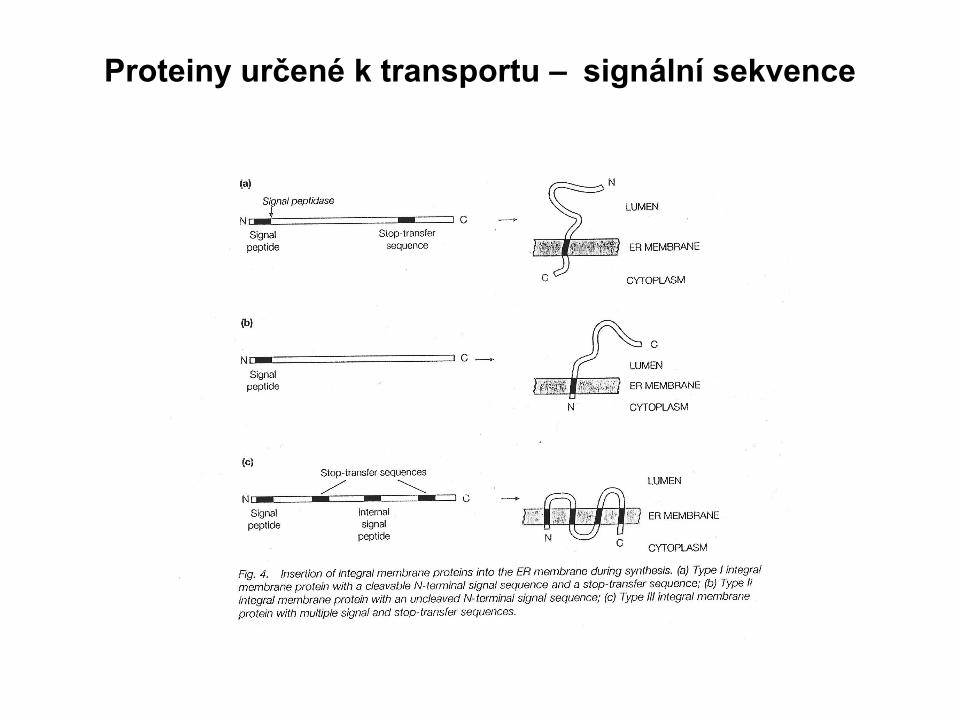
Proteiny určené k transportu – signální sekvence
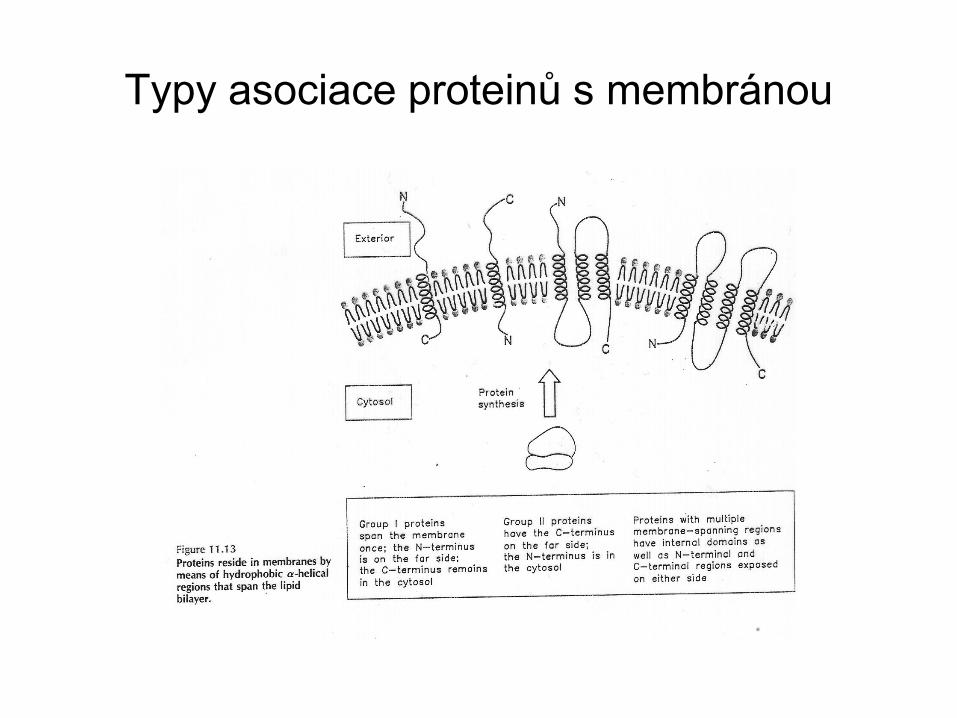
Typy asociace proteinů s membránou
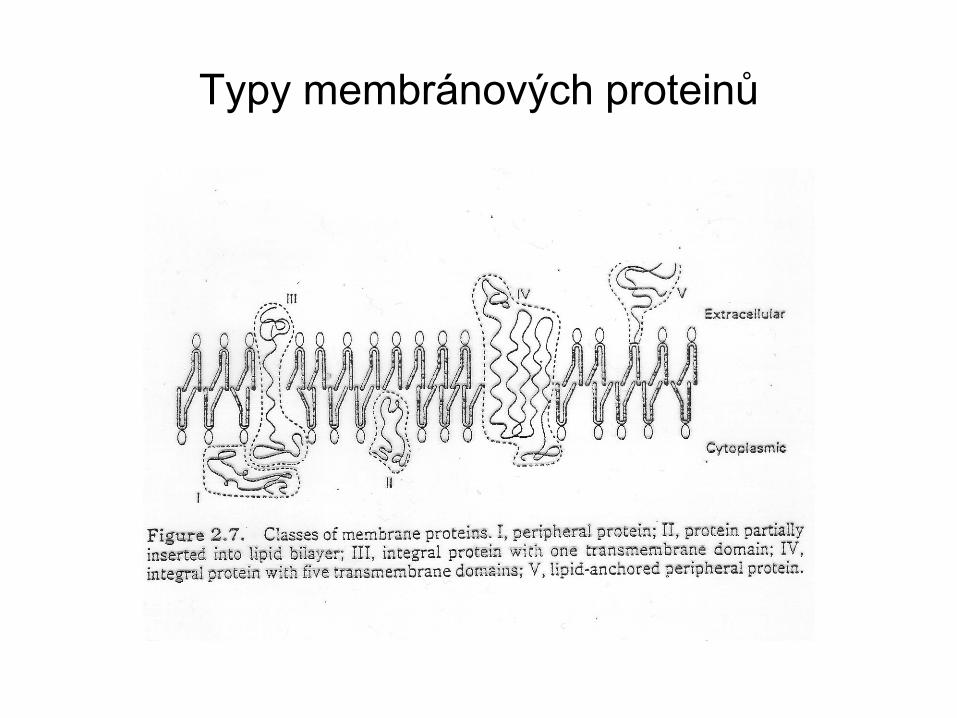
Typy membránových proteinů

Transport oligopeptidů
• Oligopermeasový systém
• opp mutanty špatně sporulují – fenotypicky se zastavují ve stadiu 0
• Sporulační defekt je výsledkem zablokování transportu oligopeptidů
• Vazebný protein pro peptid OppA je u B. subtilis zakotven v membráně jako lipoprotein
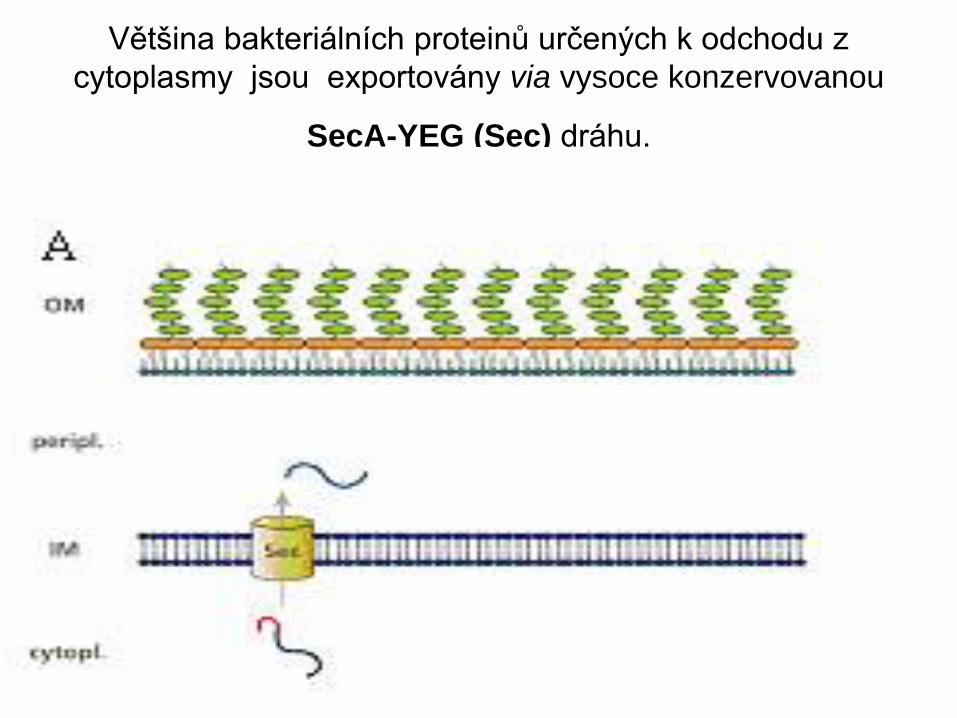
Většina bakteriálních proteinů určených k odchodu z
cytoplasmy jsou exportovány via vysoce konzervovanou
SecA-YEG (Sec) dráhu.
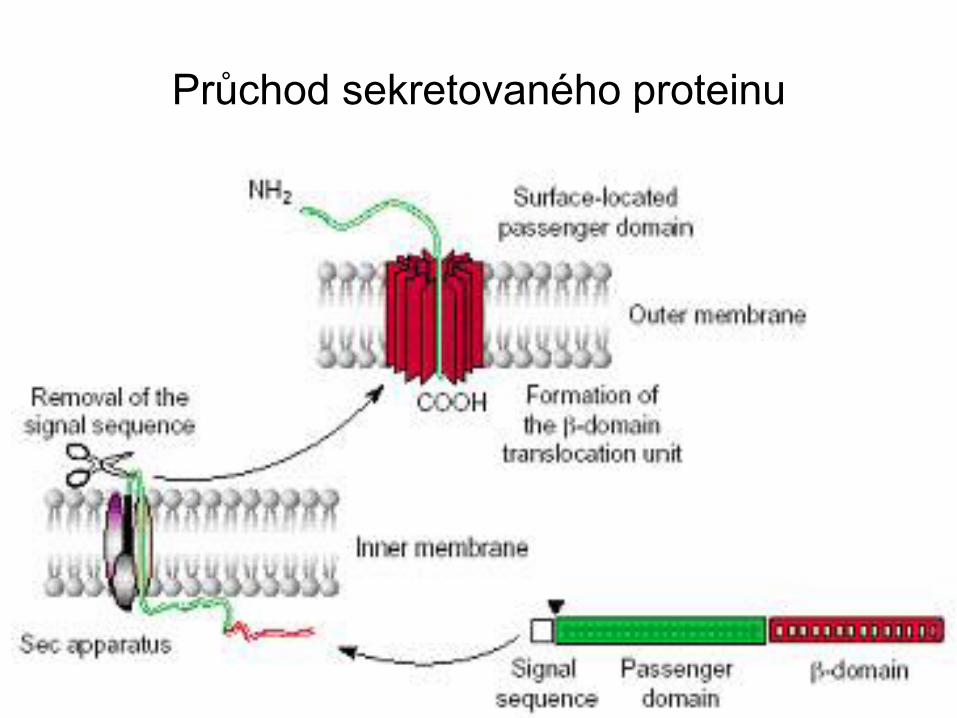
Průchod sekretovaného proteinu

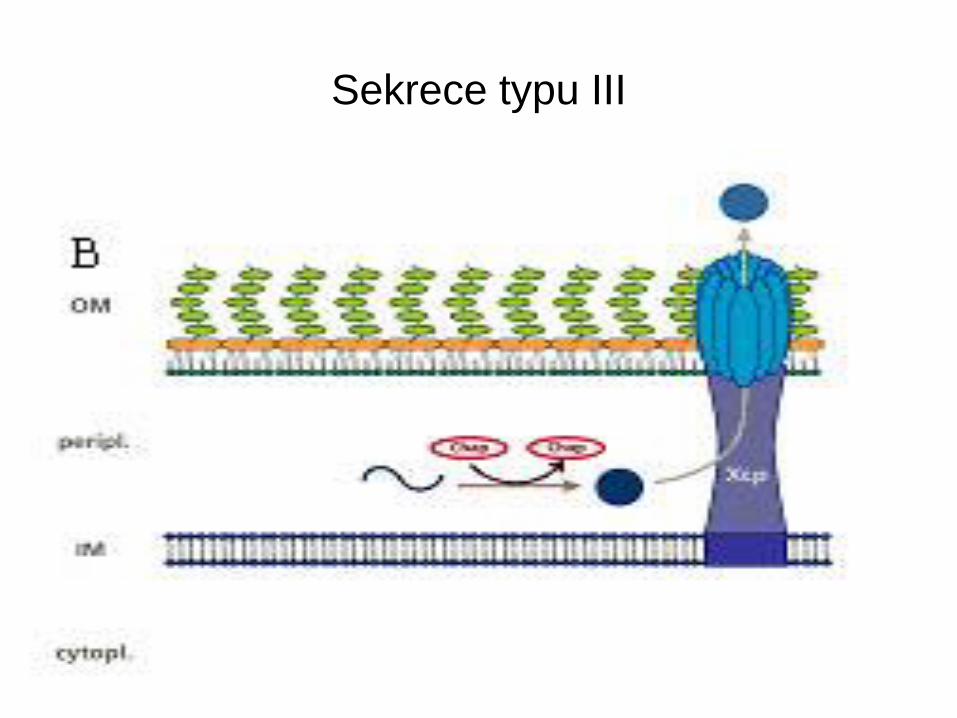
Sekrece typu III
• Vyskytuje se u G- bakterií patogenních pro lidi, zvířata i rostliny
• Tento typ sekrece podmiňuje jejich virulenci
• Systém se aktivuje kontaktem s eukaryotní cytoplasmatickou
membránou a umožňuje bakteriím injikovat své proteiny do
napadené buňky.
• Yersinia je vybavena virulonem Yop, který umožňuje obranu proti
primární imunitní obraně napadených buněk
• Sekreční aparát – 25 proteinů a soubor dalších 12 – Yop
• YopE, YopH, YopP, YopM, YopT, YopJ, YpkA/YopO, YopP/ YopJ
• Ničí cytoskeleton a blokují fagocytosu (E, H, T) zabraňují uvolnění
cytokinů(YopP/ YopJ) a indukují apoptosu

Translokace přes membránu
• Vyžaduje účast dalších proteinů – porační aktivita
• Významná je účast malého 11 kDa proteinu
LcrG, úloha při translokaci.
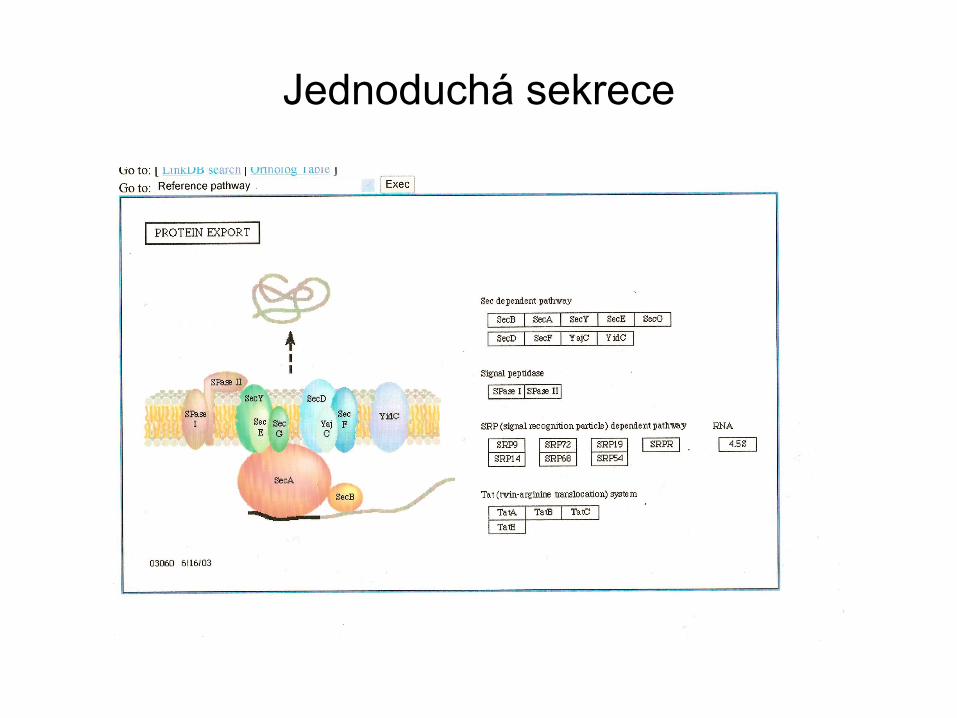
Jednoduchá sekrece
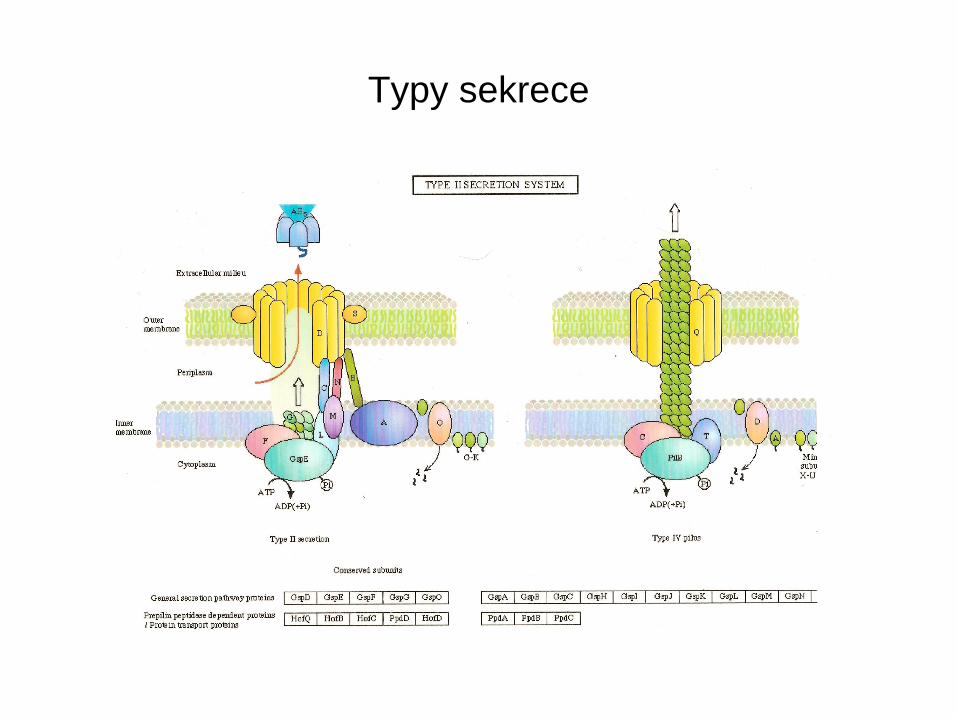
Typy sekrece
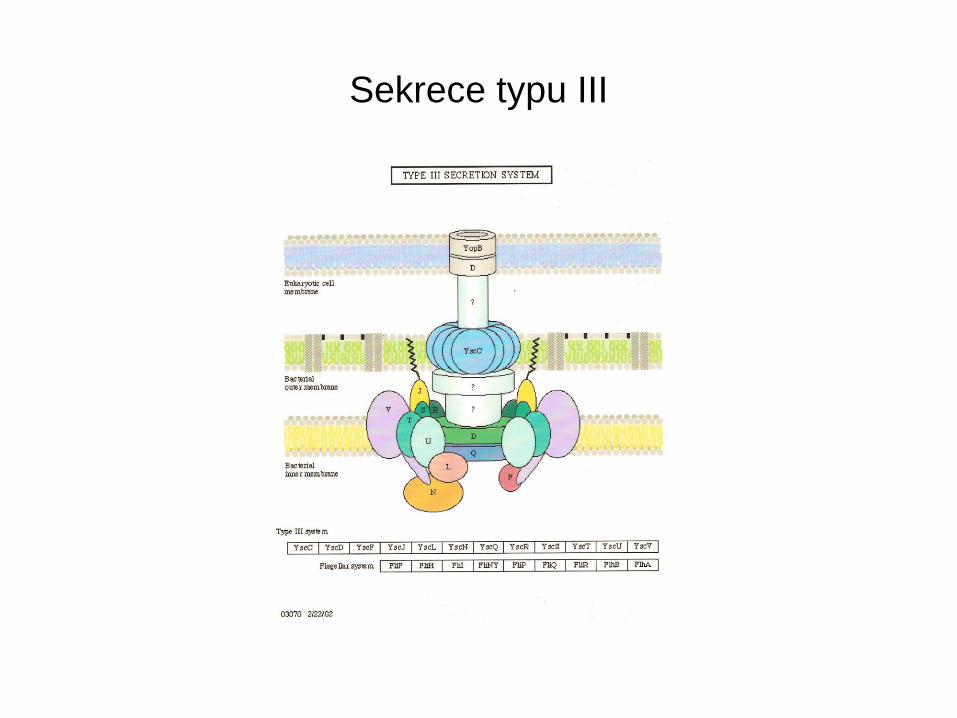
Sekrece typu III
Sekrece typu III

Sekrece typu IV – tvorba bičíků

Typy sekrece u Gram-negativních
bakterií
• Typ I. – je energizovaná hydrolýzou ATP, jedná se o jednokrokový přesun z cytoplasmy do periplasmatického prostoru ( α-hemolysin u E. coli ABC přenašeč)
• Typ II. obecná sekreční dráha vyžaduje energii z ATP a pmf, někdy GTP. První fáze přesunu přes cytoplasmatickou membránu je řízena geny anaolgními k sec genům. Proteiny mají N-terminální signální sekvenci, účast „signal recognition particles“ SRP.
• Typ III. Sekrece používá systému kanálů (pórů) vycházejících z cytoplasmy na povrch buňky. Je funkční u patogenů. Proteiny obsahují N-terminální signální sekvenci, ale ta není odštepovaná. Proteiny bazálního tělíska bičíku.
• Typ IV. Proteiny obsahují N-terminální signální sekvenci, α-doménu, C-terminální β-doménu, bičíky, proteásy patogenů.
• Typ V. Přenos T-DNA z Agrobacter tumefaciens do hostitele a sekrece toxinu Bordetella pertussis

Bakteriální neklasicky sekretované
proteiny
• Modelové vyhledávací metody byly použity k vyhledávání sekvencí
pro identifikaci údajné signální sekvence nebo motivu
odpovědného za sekreci. Žádný signál ani charakteristický motiv
nebyly nalezeny pro velký počet neklasicky sekretovaných proteinů.
Je zřejmé, že tyto proteiny jsou sekretovány novým „způsobem“.
• Nicméně bylo nalezeno, že zjevně neklasicky sekretované proteiny
jsou rozlišitelné od buněčných proteinů svým složením
aminokyselin, sekundární strukturou a neuspořádanými oblastmi
(regiony).
• Je zřetelné, že předpověď neuspořádanosti odhaluje, že bakteriální
sekretované proteiny jsou více strukturně neuspořádané než jejich
cytoplasmatické analogy.

Bakteriální neklasicky sekretované
proteiny • Sekrece proteinů přes biologické membrány je většinou
zprostředkovaná translokačními mechanismy rozpoznávajícími specifické sekvenční značky (tag) nebo motivy označujícími proteiny k sekreci.
• U bakterií, the klasický tripartitně strukturovaný Sec signální peptid řídí cíle sekrečních drah. Kromě Sec-závislé sekrece, různé další sekreční dráhy byly objeveny,pracují způsobem nezávislým na Sec. Převládajícím typem je twin-arginine translokace (Tat) tato sekreční dráha využívá twin-arginine konsensus motivu a je uložena uvnitř vlastního signálního peptidu.
• Zatímco Sec- a Tat-závislá seckreční dráha translokuje proteiny pouze přes vnitřní membránu u Gram-negativních bakterií, přídavné translokační mechanismy byly nalezeny ve vnější membráně těchto bakterií. N-terminální signální peptid má centrální roli v těchto sekrečních systémech jako tag-signální sekrece.
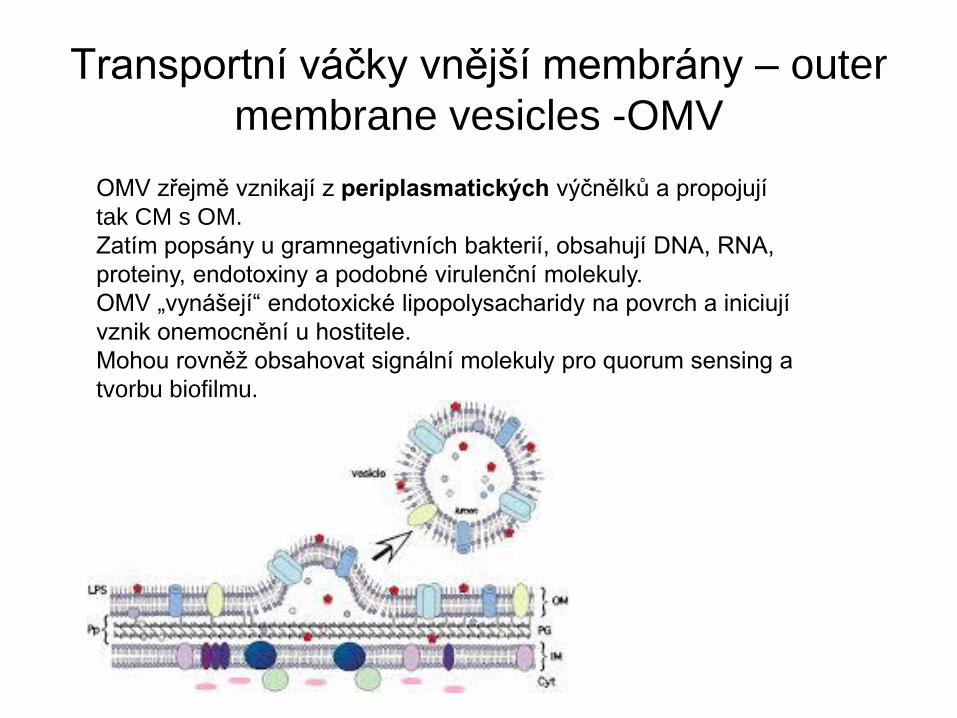
Transportní váčky vnější membrány – outer
membrane vesicles -OMV
OMV zřejmě vznikají z periplasmatických výčnělků a propojují
tak CM s OM.
Zatím popsány u gramnegativních bakterií, obsahují DNA, RNA,
proteiny, endotoxiny a podobné virulenční molekuly.
OMV „vynášejí“ endotoxické lipopolysacharidy na povrch a iniciují
vznik onemocnění u hostitele.
Mohou rovněž obsahovat signální molekuly pro quorum sensing a
tvorbu biofilmu.
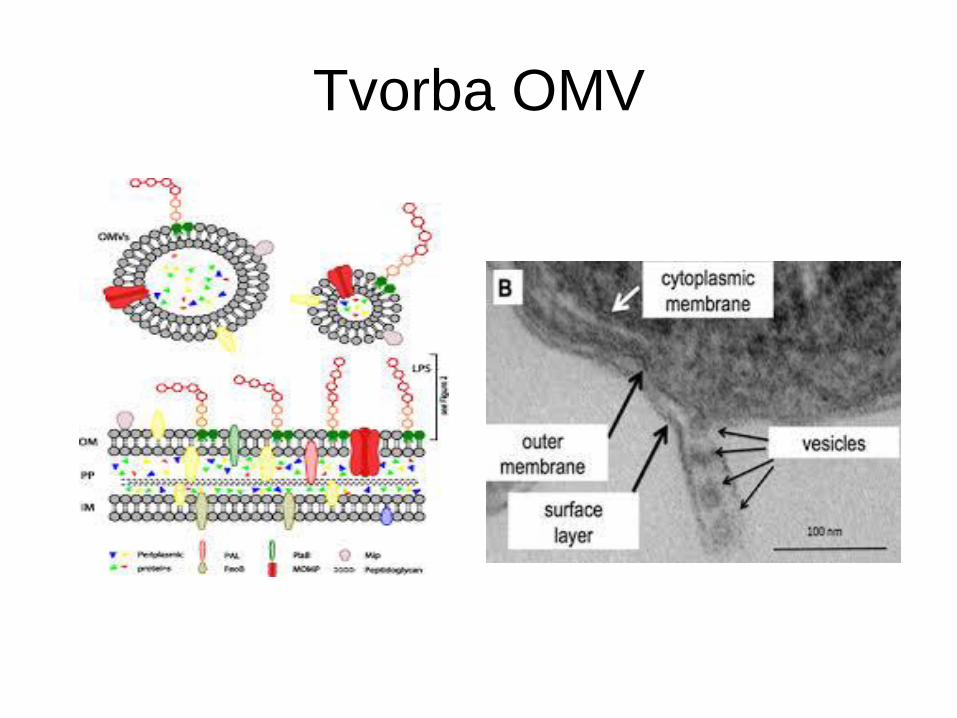
Tvorba OMV
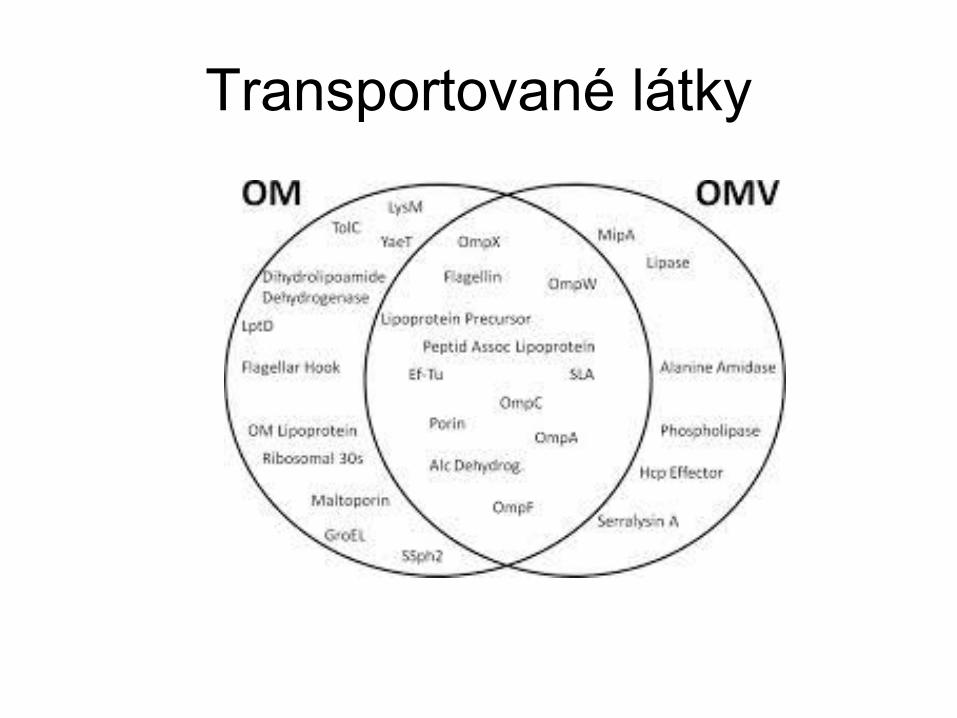
Transportované látky
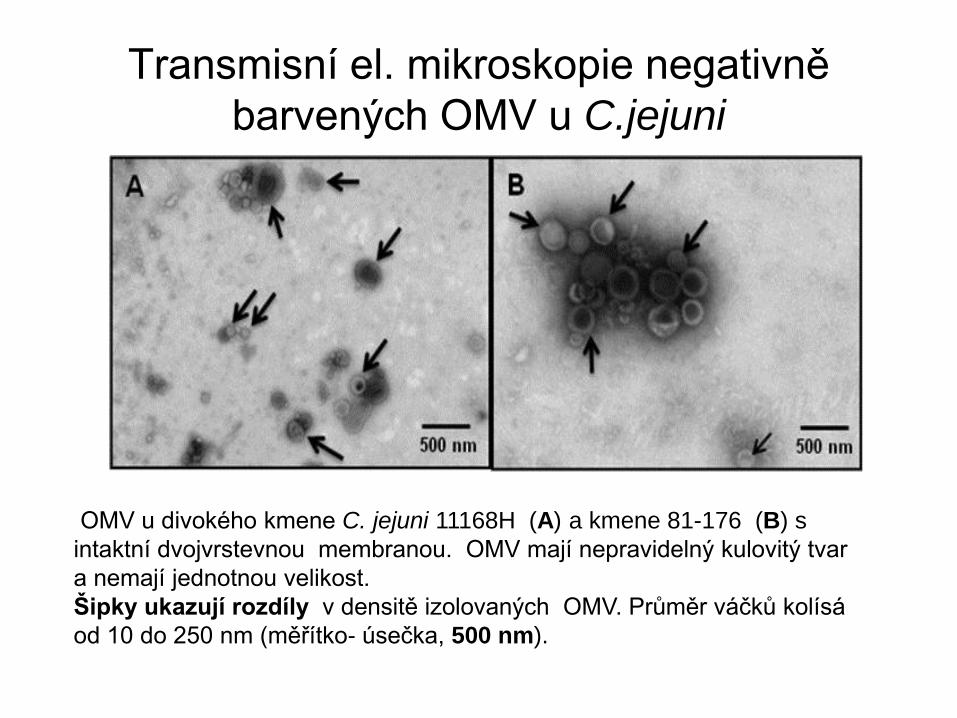
Transmisní el. mikroskopie negativně
barvených OMV u C.jejuni
OMV u divokého kmene C. jejuni 11168H (A) a kmene 81-176 (B) s
intaktní dvojvrstevnou membranou. OMV mají nepravidelný kulovitý tvar
a nemají jednotnou velikost.
Šipky ukazují rozdíly v densitě izolovaných OMV. Průměr váčků kolísá
od 10 do 250 nm (měřítko- úsečka, 500 nm).

Proteomická analýza C. jejuni 11168H
.
Bylo identifikováno 151 proteinů, včetně periplasmatických a
s vnější membránou asociovaných proteinů, ale také proteiny
důležité pro přežití a patogenezi, včetně CDT (cytoletální
distendní toxin).
OMV obsahovaly 16 N-vázané glykoproteiny, což naznačuje, jak
periplasmaticky lokalizované imunogenní proteiny mohou
interagovat s buňkami hostitele.
Váčky C.jejuni mají cytotoxickou aktivitu a indukují imunitní
odpověď hostitele na T84 intestinálních epiteliálních buňkách. Tato
odpověď nebyla zeslabena působením proteinázy K nebo
polymyxinem B před jejich inkubací s intestinálními epiteliálními
buňkámi.

Modelový systém (artificial neural networks) byl použit ke konstrukci znaků
proteinů k identifikaci neklasicky sekretovaných proteinů u Gram-pozitivních a
Gram-negativních bakterií
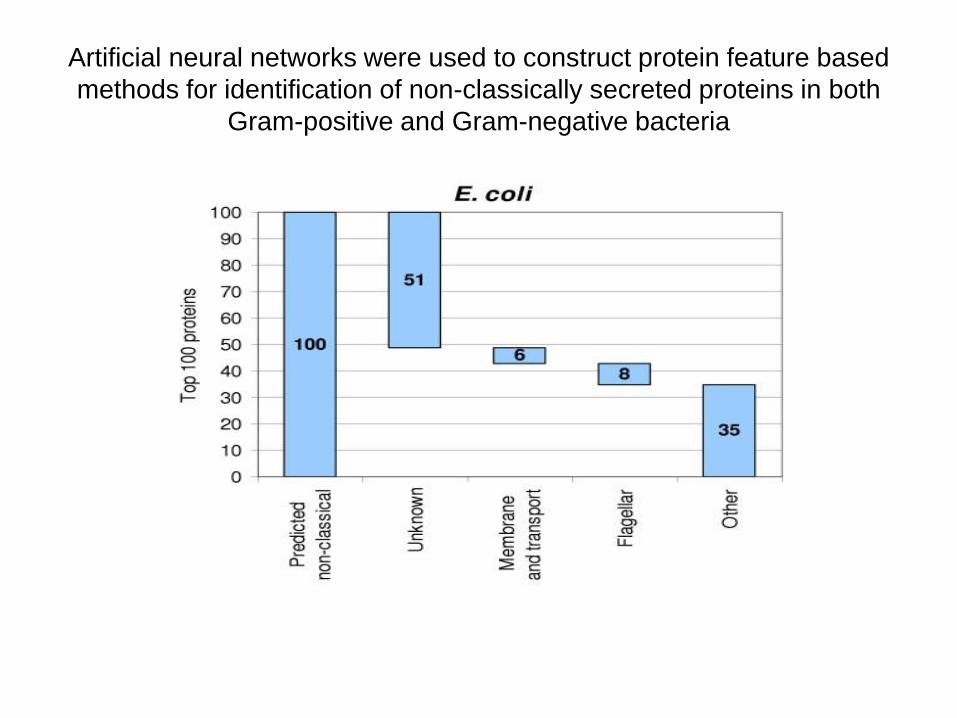
Artificial neural networks were used to construct protein feature based
methods for identification of non-classically secreted proteins in both
Gram-positive and Gram-negative bacteria
Účast Golgi aparátu na transportu
Účast dalších vesikulů na transportu
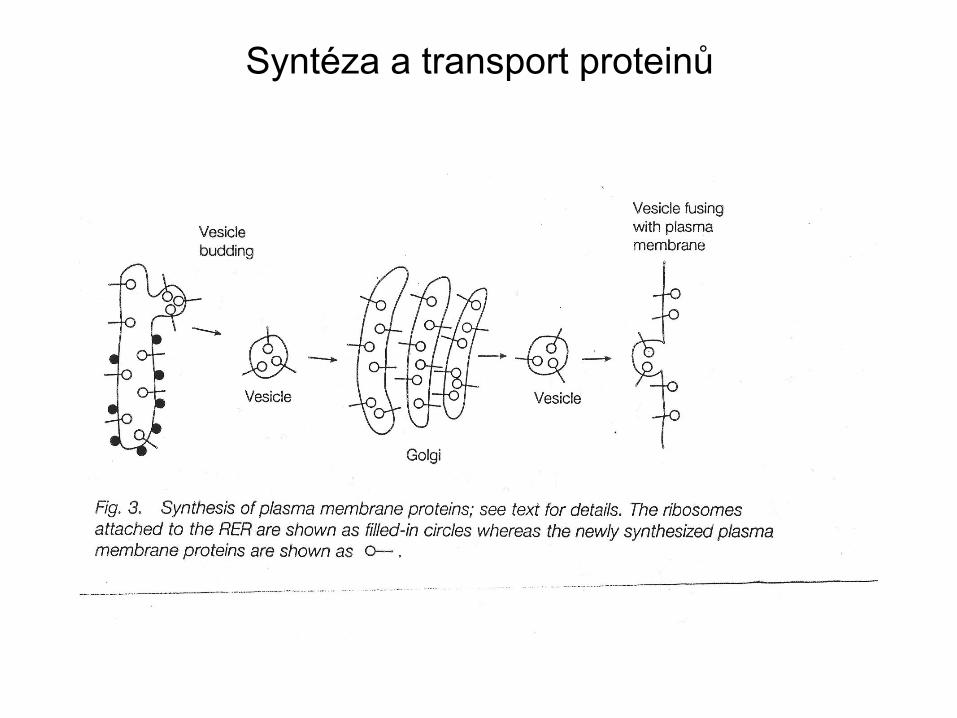
Syntéza a transport proteinů
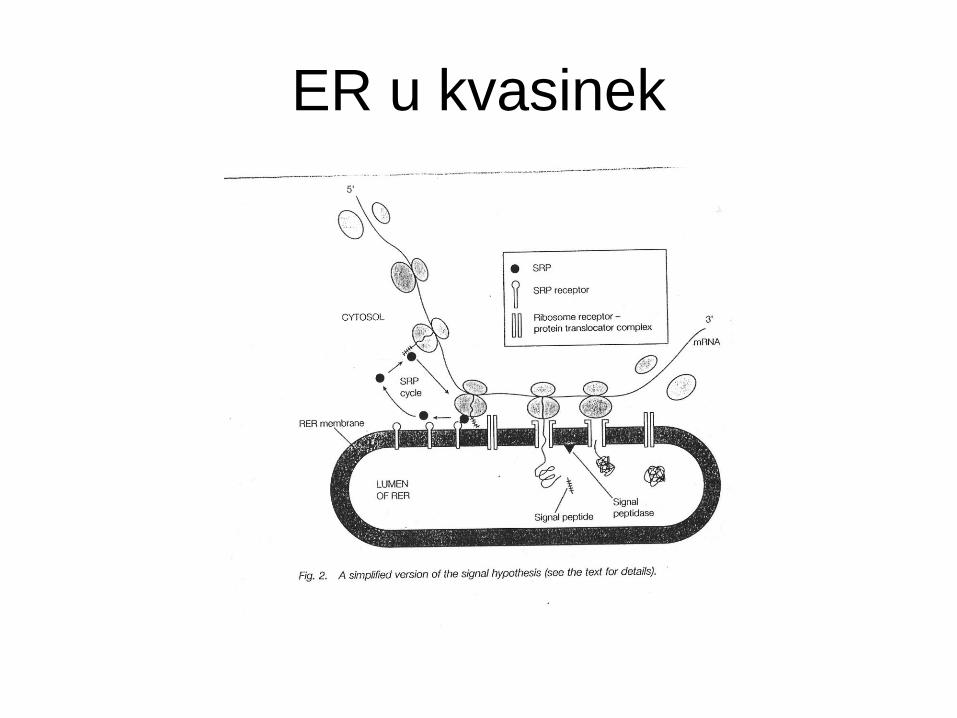
ER u kvasinek

Transport u kvasinek
• Endoplasmatické retikulum (ER) - cytoplasmatická organela -
počátek sekreční dráhy.
• Sekrece proteinů začíná spolu s kotranslační inzercí
polypeptidového řetězce do ER.
• Proteiny jsou glykosylované a skládají se v lumen ER a přepravují
se v obalených váčcích (COPII coated vesicles), které pučí ER a
dopravují svúj náklad do Golgi aparátu.
• Proteiny po projití Golgi aparátem jsou směrovány buď do vakuol
nebo do cytoplasmatické membrány.
• Ačkoli se ER a jádro liší strukturou, jejich membrány a lumen jsou
are spojité a oba systémy jsou místy přenosu proteinů.

Kvasinkový feromon
• Kvasinkový „kopulační“ feromon alfa
faktor inhibuje adenylát cyklasu
• in vitro, alfa faktor inhibuje membránově
vázanou adenylát cyklasu buněk v
závislosti na koncentraci.
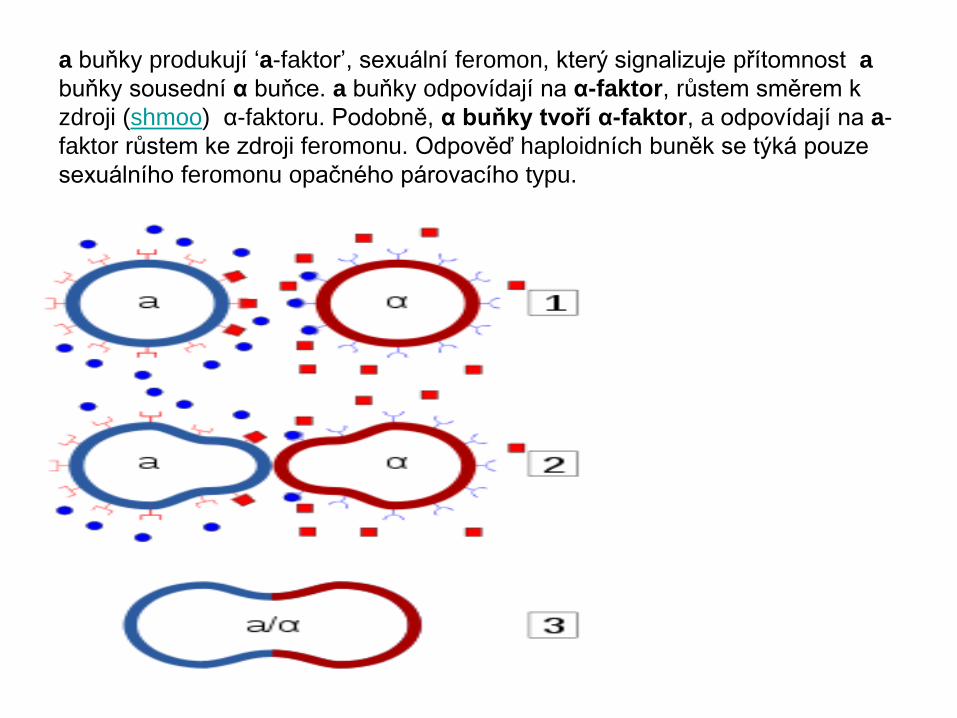
a buňky produkují ‘a-faktor’, sexuální feromon, který signalizuje přítomnost a
buňky sousední α buňce. a buňky odpovídají na α-faktor, růstem směrem k
zdroji (shmoo) α-faktoru. Podobně, α buňky tvoří α-faktor, a odpovídají na a-
faktor růstem ke zdroji feromonu. Odpověď haploidních buněk se týká pouze
sexuálního feromonu opačného párovacího typu.

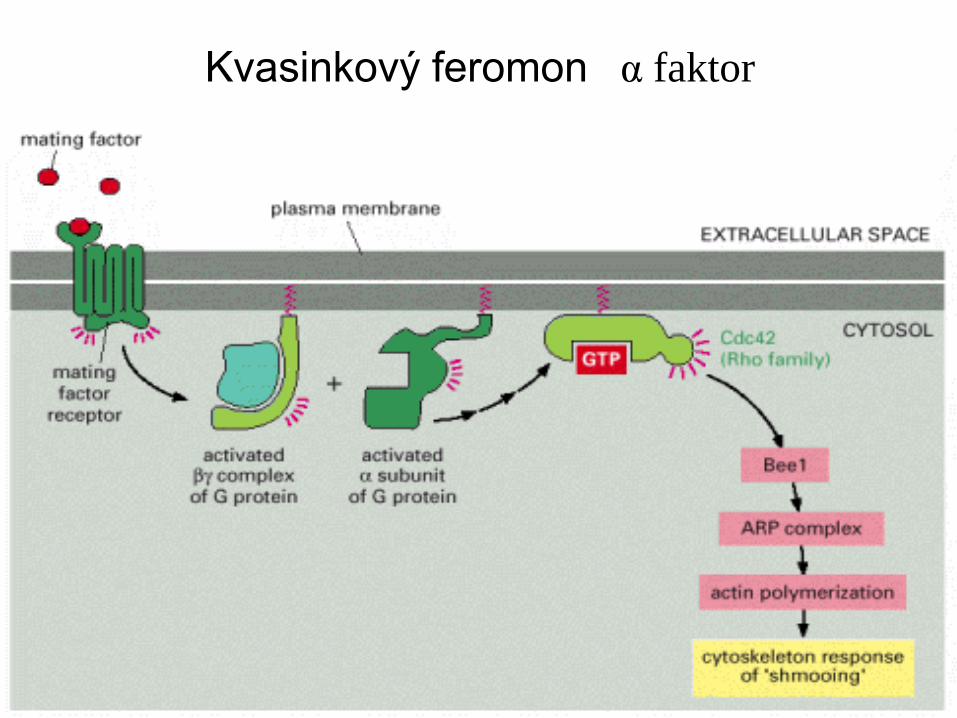
Kvasinkový feromon α faktor

Signální dráha kvasinkového sexuálního faktoru
Extracelulární „mating factor“ se váže na G-protein vázaný receptor v cytoplasma-
tické membráně.
Aktivace receptoru spouští disociaci podjednoty Ga, která váže GTP z hetero-
trimerického G-proteinu. Ten obratem aktivuje GTP-vazebný protein z Rho family,
Cdc42, který je kvasinkovým homologem malé savčí GTPase, která aktivuje
spouštění za tvorby filopodie.
Tak jako v savčích buňkách Cdc42 aktivuje protein (Bee1, homolog WASp) který
aktivuje ARP komplex, vedoucí k místní nukleaci aktinu, v místě vazby „mating“
Faktoru. Místní nukleace aktinu a růst vláken vede k polarizovanému růstu a získání
tvaru „shmoo“. Nadto aktivace receptoru spouští další odpovědi přes MAP kinasovou
Kaskádu a připravuje tak haploidní buňku na spájení („mating).

















